Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
Beer is a world-famous beverage, second only to tea and coffee, where the yeasts traditionally used are Saccharomyces cerevisiae and Saccharomyces pastorianus for the production of ale and lager beer, respectively. Their production, especially craft beer production, has grown in recent years, as has the development of new products. For this reason, research has focused on the selection of yeasts with good fermentation kinetics, as well as beers with outstanding aromatic profiles. The final flavor and aroma of beer is a combination of hundreds of active aroma compounds produced mostly during fermentation as a result of yeast metabolism (higher alcohols, esters, aldehydes, and vicinal diketones). Likewise, several studies have demonstrated the potential of wild yeasts of the genus Saccharomyces, both in aromatic production and in the production of healthy compounds of interest such as melatonin. This chapter therefore focuses on non-conventional Saccharomyces yeasts as they have the capacity to produce outstanding aroma compounds, as well as compounds that can provide health benefits, under moderate consumption.
Department of Agrifood Research, Madrid Institute for Rural, Food and Agriculture Research and Development (IMIDRA), Spain
Margarita García
Department of Agrifood Research, Madrid Institute for Rural, Food and Agriculture Research and Development (IMIDRA), Spain
Teresa Arroyo
Department of Agrifood Research, Madrid Institute for Rural, Food and Agriculture Research and Development (IMIDRA), Spain
*Address all correspondence to: vanesa.postigo@madrid.org
1. Introduction
Nowadays, there is a growing global interest in craft beer, as well as in the production of new beers that meet market needs. For this reason, brewers have focused especially on the use of yeasts, especially non-conventional yeast, to innovate in the brewing sector [1, 2, 3, 4, 5]. Wild yeast strains can provide different aroma and flavor characteristics with which to obtain varieties and styles of beer that are alternative and new to existing ones [6]. However, the use of these domesticated yeasts may show variable fermentative characteristics and affect the consistency and quality of the beers produced [4, 7]. Unlike the current commercial strains of Saccharomyces yeast, their domestication has occurred over millennia, resulting in brewers obtaining consistent beers with established aromas, flavours and fermentation conditions [4, 8]. Nowadays, however, all the steps leading to yeast domestication can be done in a faster way [2, 9], since the metabolism pathways of yeast are better known, as well as the production pathways of the different aromatic compounds (higher alcohols, esters, phenols, acids, and monoterpenes) [2, 10, 11, 12, 13]. It is for this reason that brewers and researchers have focused on the search for new yeasts from different environments using current techniques [14]. Determining the suitability of new yeast strains can be done by evaluation by expert panels; however, this must be coupled with their analysis using physicochemical analytical techniques to show correlation with the aroma compounds produced. Such compounds are produced in smaller proportions than ethanol and carbon dioxide, which are the main products of fermentation, but are nevertheless of great importance [10, 15]. Secondary metabolites can be divided into different categories and include sulfur-containing aroma compounds, undesirable carbonyl compounds, volatile phenols, organic acids, fusel alcohols, esters, and monoterpene alcohols [10, 15, 16, 17]. These groups of aromatic compounds play a number of roles in the impact of flavor on beer, as many of them can act synergistically, thus increasing flavors, despite being below their threshold [11, 15, 16].
The present review focuses on the use of non-conventional yeasts of the genus Saccharomyces for fermenting beer wort, obtaining an outstanding organoleptic profile, as well as the production of secondary metabolites with healthy characteristics for potential use by brewers.
Beer is a worldwide-known beverage and the most popular after tea and water. However, people do not focus their attention on how it is brewed, as well as on its ingredients, which are barley as the main cereal, water, hops, and yeast. Other cereals or adjuncts may also be added [18, 19].
Water is the main component of beer, accounting for 85–92% of beer, as well as serving as the basis for brewing the beer wort. Water quality is an important factor to take into account since the presence of any odor, taste, and impurity is unacceptable because of its influence on the sensory properties of the beer. The enzymatic and colloidal reactions that occur during brewing are influenced by the content of dissolved ions found in water such as Ca2+, Mg2+, Na+1, K+1, SO4−2, Cl−, HCO3−1, and CO3−2 [20]. Calcium (Ca2+) is involved in protein coagulation and polyphenol and carbohydrate formation, forms and precipitates calcium oxalate and calcium carbonate, reduces pH during mashing and boiling, and promotes yeast metabolism and flocculation, and its concentration is key to residual hardness and alkalinity. Magnesium (Mg2+) is a vital nutrient for yeast and necessary for the isomerization of humulone to isohumulone in hops. Sodium (Na+) can contribute to the beer body. Chloride (Cl−) in low amounts enhances malt sweetness. Bicarbonate (HCO3−) functions as a buffering agent, buffering pH variations [21].
The basis of beer production is malted barley. The malting process is carried out for the yeast fermentation, as well as proteins that promote and enhance foam stability [22]. Malt can provide the beer with aromas of biscuit, honey, cinnamon, bread, chocolate, cocoa, coffee, and roasted and/or smoked, among others [23].
Hops (Humulus lupulus L.) is the ingredient that will mainly provide the bitterness of the beer due to the iso-α-acids (iso-normal-humulone, iso-co-humulone, and iso-ad-humulone) obtained by the isomerization of the α-acids (normal-humulone, co-humulone, and ad-humulone) during the wort boiling and whirlpool process in brewing. These α-acids are contained in the resins of hop flowers [24]. In addition to bitterness, hop also contributes to beer aromas, such as citrus, floral (geraniol, linalool), herbaceous, and fruity (limonene, myrcene, β-pyrene), due to the resins, proteins, and essential oils they possess [25]. Hops are sensitive to oxidation in contact with air, so it is important that they are stored under vacuum in a cool, dry place; otherwise they can transmit unpleasant aromas to the beer (such as isovaleric acid with rancid cheese aroma).
Yeast is an important ingredient in beer brewing, as it is responsible for fermentation and thus for the conversion of the sugars in the beer wort into ethanol and carbon dioxide (CO2). However, it will also produce different by-products, the main ones being aromatic compounds that will characterize the aromatic profile of the beers.
In the past, these four ingredients were not well-known, but it was known that yeast was necessary for the production of fermented beverages. It was not until the development of lager beers that the exact composition of yeast was discovered. Although yeasts have been used for centuries to brew beer, they were first identified as responsible for the fermentation of malted barley water in the 19th century. The first principles of yeast function were discovered during the 17th and 18th centuries [26]; however, it was not until the mid-19th century that the French scientist Louis Pasteur was able to demonstrate that yeast is made up of living cells responsible for the fermentation process [27]. In the same vein, sugars were also not known to be another essential ingredient and can be classified according to the type of fermented beverage, whether from sugar cane (sucrose), milk (lactose), fruit or honey (fructose and glucose), or cereals (maltose) [19].
3. Beginnings in the use of yeast for beer production
The beginnings of beer production date back to the Neolithic civilization. Beer is a traditional product and valued for its physicochemical properties that give it quality. Therefore, the history of brewing is not only the history of scientific and technological developments but also the history of people: their government, their culture, and their daily life [28]. Early brewers, winemakers, and bakers realized that by using small portions of finished products that had already been fermented, it was possible to obtain products with faster and more predictable fermentation. Thus, the ability of Saccharomyces cerevisiae yeast to convert sugars into ethanol, as well as into other compounds that provide organoleptic characteristics to foods, in addition to alcohol to beverages, began to be exploited [2, 29]. However, the use of pure yeast cultures did not begin until sometime after the pioneering work of Pasteur and Hansen in the 19th century [27]. It should be noted that it has not yet been determined whether the yeast lineages used industrially originated from the loss of contact with their natural niches and limited dispersal or from adaptation to the industrial niche (domestication) [30, 31]. Domestication therefore is about artificially selecting and breeding wild species with improved characteristics adapted to artificial environments. However, this domestication can lead to genome breakdown, polyploidy, chromosomal rearrangements, gene amplifications and deletions, horizontal gene transfer, and loss of genetic diversity due to bottlenecking [32, 33].
The most studied yeast at the industrial level is Saccharomyces cerevisiae, where its diversity lies both in genetic drift by bottleneck and small isolated populations as well as in selection and niche adaptation. In yeasts used for winemaking, horizontal gene transfer events [34, 35, 36] and copy number variations [37, 38, 39, 40] have been described that increase sugar and nitrogen metabolic activity, which favors fermentation of grape must, as well as better tolerance to chemicals used in vineyards and in wine [41]. In contrast, the most significant changes are found in the yeasts used for brewing, as yeasts are generally reused for several fermentation batches. This continuous growth in a very specific industrial niche has resulted in a continuous selection imposed by the brewing environment. Therefore, the yeasts currently used at the industrial level could be considered as the result of a centuries-long evolutionary experiment in a highly selective niche.
The increase in the consumption of craft beer [42], as well as consumers’ interest in trying new beer styles [43], has encouraged the application of new yeasts in brewing [44, 45]. These yeasts include the wild yeast Saccharomyces eubayanus (ancestor of today’s lager yeast hybrid), as well as various species such as Torulaspora delbrueckii, Lachancea thermotolerans, and Mrakia gelida [46, 47, 48]. These non-conventional yeasts have functional advantages over commercial yeasts as they provide novel flavor profiles, production of flavorful beers with low alcohol content, reduced caloric content, and primary acidification, in addition to their use to improve existing beer styles or create new styles. Currently, the use of new yeasts focuses not only on the isolation of wild yeasts but also on mutagenesis and breeding. However, the most attractive option for brewers remains the isolation of yeast strains from different niches [2]. The advantages offered by wild yeast isolation are extensive:
The natural biodiversity of microorganisms representing the habitat of a geographic region can be tapped.
These new species can bring added value to beer in terms of organoleptic qualities.
Some yeasts isolated from nature already have the status of Generally Recognized as Safe/Qualified Presumption of Safety.
Current regulations favor the use of unmodified genetic stocks, as they add identity and uniqueness to differentiate the production line.
On an industrial level, not only ethanol and glycerol production but also the utilization of available sugars in the wort, hop tolerance and resistance to low temperatures, and the relative production of aromatic compounds such as esters and higher alcohols, as well as low levels of acetic acid and hydrogen sulfide, are important [2, 45, 49]. Not all species are able to ferment the sugars present in the wort (glucose, fructose, maltose, and maltotriose). In the case of S. eubayanus, it seems highly efficient at utilizing maltose in addition to presenting an efficient production of biomass at low temperature (ideal for lager brewing), thus balancing with the non-capacity to ferment maltotriose. Wild yeasts are noted for their potential to produce remarkable aroma compounds in brewing [50]. The type strain of Saccharomyces arboricola has been found to produce high levels of ethyl esters [50]. However, it should also be noted that numerous Saccharomyces wild yeast strains tend to produce significant perceivable levels of unpleasant phenolic aroma (POF), mainly through the production of 4-vinyl guaiacol from ferulic acid [51], imparting a clove-like aroma to beer. This is considered undesirable in most beer styles. In some cases, POF aromas do not represent a problem, but are desired in the final fermented product, such as wheat beer, where clove aroma is part of the normal flavor profile [52]. Therefore, the ability of Saccharomyces wild yeasts to ferment at low or high temperatures, to obtain new aromatic profiles, high or low ethanol levels and a reduction in H2S production, as well as to supporting stressful wort conditions, represents a potential for new processing strategies. Cubillos et al. [53] collect in their study the application of various strains of wild yeast for application in beer.
Another advantage of wild yeasts is that they can be subjected to different fermentation conditions to observe their behavior. In this case, from a technological point of view, the application of aeration during fermentation is an interesting tool for controlling yeast metabolism during fermentation [54]. First, due to Crabtree-negative yeasts, when the oxygen concentration in the medium is saturated, the yeast metabolism begins to be predominantly oxidative, thus reducing the ethanol content and increasing the yeast biomass. On the other hand, aeration may also affect aromatic compounds by reducing or increasing their concentration in beer (the acetaldehyde content may decrease, and the concentration of higher alcohols increases) [55, 56, 57]. Within the non-conventional strains of Saccharomyces, there are strains that may present the Kluyver effect with aeration during fermentation, as in the study conducted by Postigo et al. [58]. In this study, during fermentation with continuous aeration, it was observed that one of the tested strains (CLI 1057) that under anaerobic conditions only fermented glucose, fructose, and sucrose, when changing to aeration conditions, began to ferment part of the maltose present in the must [59, 60]. This behavior resulted in a variation in the concentration of aromatic compounds where isoamyl acetate, DMS, diacetyl, and 2,3-pentanedione decreased considerably, as observed in other studies such as Mauricio et al. [61]. However, the rest of the compounds increased, resulting in a more phenolic beer than without aeration.
The beverage industry has focused on the search for fruity and floral aromas, which is why consumers demand beers with fruitier aromatic profiles. The ingredients in beer that can provide such aromas and flavors are hops, but mostly yeast during the fermentation process, as it will provide a fruitier organoleptic profile to beers. Various aroma compounds can be found in beer, although studies mainly focus on the alcohols and esters produced by yeast as they will provide the main aromas found in beer [10, 62, 63].
Meilgaard elaborated the “beer aroma wheel,” where all important beer aromas were included, including all yeast-derived aromas, as well as all other raw materials used during brewing [64].
Main aromatic compounds relevant in beer:
Sulfur compounds: yeasts can produce sulfur dioxide (SO2) and hydrogen sulfide (H2S) during fermentation [65, 66]. SO2 has several functionalities, including acting as an antimicrobial agent, bleaching agent, oxygen scavenger, reducing agent, and enzyme inhibitor [67]. However, the maximum levels of SO2 allowed in beer depend on the regulations of each country, always taking into account its perception threshold (20 mg/L), as it can impart unpleasant aromas to beer [68]. Although it should be noted that it is a positive characteristic desired in some bottom-fermented beers [69], this compound is usually found in concentrations of less than 10 mg/L, being a secondary or feedback inhibition product of amino acid anabolism [65, 69]. On the other hand, H2S (rotten egg aroma) has a lower perception threshold (0.005 mg/L) and can mask desired aromas in beer [70, 71]. Its production occurs mainly during the cell maturation cycle, but it is assimilated later in the budding cycle [66].
Carbonyl compounds: within this group, the most prominent compounds that produce undesirable aromas in beer are vicinal diketones and acetaldehyde, produced by yeasts during the fermentation process. Vicinal diketones include diacetyl and 2,3-pentanedione. Both have low thresholds (0.15 mg/L and 0.9 mg/L, respectively [72]) and a buttery or toffee-like aroma. Diacetyl is a by-product of valine anabolism during glucose metabolism. Later, during the beer maturation phase, yeasts take up diacetyl and reduce it to 2,3-butanediol, which has no unpleasant aroma [73]. 2,3-pentanedione, on the other hand, is produced during the synthesis of the amino acid isoleucine in the mitochondria of yeast cells, but like diacetyl, its content is reduced during maturation [73, 74]. Acetaldehyde is produced during the yeast growth phase as a result of sugar metabolism and is subsequently converted to ethanol [75]. It has an undesirable green apple or grassy taste, with a threshold of 30 mg/L [72, 76, 77].
Phenolic compounds: these compounds are generally referred to as phenolic off-flavors (POFs) for most beers but are accepted in Belgian-style beers or wheat beers. These compounds contribute clove, smoky, spicy, medicinal, and burnt aromas [78, 79]. The compounds responsible for these aromas are generally 4-vinylguaiacol, 4-vinylphenol, 4-ethylguaiacol, 4-ethylphenol, 4-vinylsyringol, styrene, eugenol, guaiacol, and vanillin [80]. Although they depend on the presence of certain precursors in the wort for their production, generally the type of yeast used is largely responsible for their formation. The precursors that can be found in the wort are phenolic acids with a high flavor threshold, such as ferulic, coumaric, and cinnamic acids, which are derived from malt [51, 80]. These compounds usually have low thresholds: 4-ethylphenol, 0.9 mg/L (phenolic aroma, astringent); 4-ethylguaiacol, 0.13 mg/L (phenolic aroma, sweet); 4-vinylguaiacol, 0.3 mg/L (phenolic aroma, bitter, clove); guaiacol, 3.88 μg/L (smoky aroma); and 4-vinylphenol, 0.2 mg/L (phenolic aroma, smoky) [72, 81].
Organic acids: within the organic acids are non-volatile and volatile acids, the latter being the most prominent in beer as they generally contribute along with the rest of the off-flavors [82]. In beer, the main organic acids that can be found are propionic, isobutyric, butyric, isovaleric, valeric, caproic, acetic, caprylic, caprylic, capric, and lauric, the last four being the ones that most influence the flavor of beer [83, 84]. Their concentration plays an important role since in high concentrations, they can contribute acidic, salty, or even cheesy and sweaty flavors [16, 83, 84, 85]. Acetic acid, with a vinegar-like aroma, has a high threshold of 175 mg/L, whereas caprylic acid, with a goat-like aroma, has a much lower threshold of 15 mg/L. Capric acid contributes waxy-like aroma with a threshold of 10 mg/L, and lauric acid is described as soapy above the threshold of 6.1 mg/L [16]. On the other hand, the most prominent non-volatile organic acids are oxalic acid, citric acid, malic acid, fumaric acid, succinic acid, lactic acid, and pyruvic acid, with thresholds higher than those of the volatile acids, ranging from 220 to 700 mg/L, most of which provide acidic flavors [16, 83, 84, 86, 87, 88]. This group of organic acids is dependent on the yeast strain used and is a by-product of glycolysis, the citric acid cycle, and the metabolism of amino acids and fatty acids [88].
Higher alcohols: higher alcohols are produced by yeasts as a by-product of amino acid metabolism and catabolism. During catabolism, yeasts take up the amino acids and they are transaminated by four transaminase enzymes [89]. The resulting product is an α-keto acid, which through an irreversible reaction forms the higher alcohols, being known as the Ehrlich route [90, 91]. The main aromas provided by the higher alcohols are floral, fruity, or herbaceous, the most important in beer being n-propanol, isobutanol, isoamyl alcohol, and 2-phenylethanol [10, 16, 92]. N-butanol provides a sweet taste, but its threshold is high (600 mg/L) [92]. Isobutanol and amyl alcohol have solvent-like aromas, with thresholds of 100 and 50–70 mg/L, respectively. Isoamyl alcohol and 2-phenylethanol give fruitier aromas, whereas isoamyl alcohol shows more banana and alcohol flavors with a threshold of 50–65 mg/L. The 2-phenylethanol compound has a gummy bear and rose flavor at a threshold of 40 mg/L [16, 92].
Esters: in addition to higher alcohols, esters are also an important group of aromatic compounds because they have low thresholds and provide fruity aromas to beer [10, 11]. The esters can be divided into two groups, the acetate esters whose concentration is majority in beers and the medium-chain fatty acid ethyl esters [10, 11]. The most important esters that can be found in beer are ethyl acetate (solvent flavor with a threshold of 33 mg/L), isoamyl acetate (banana flavor with a threshold of 1.6 mg/L), isobutyl acetate (fruity and sweet flavor with a threshold of 1.6 mg/L), and phenylethyl acetate (aroma of rose, apple, and honey with a threshold of 3.8 mg/L) [16, 93, 94]. Moreover, medium-chain fatty acid ethyl esters are formed from a medium-chain fatty acid and an ethanol radical. The most prominent are ethyl hexanoate (apple, anise flavor, with a threshold of 0.23 mg/L) and ethyl octanoate (acid apple flavor, with a threshold of 0.9 mg/L) [10, 11, 16].
The use of non-conventional yeasts in winemaking has been extensively studied in both Saccharomyces and non-Saccharomyces, as they have shown great potential in the production of aromatic esters during vinification, in both mixed and sequential inoculation [95, 96, 97]. However, despite the great aromatic potential of non-conventional yeasts, there are fewer studies on beer than on wine [6, 98]. In Saccharomyces species, mainly the Ehrlich pathway for fusel alcohols or the enzymes responsible for ester formation have been studied as main pathways for the formation of aroma compounds [99, 100, 101]. Gamero et al. [102] also carried out their studies with non-Saccharomyces yeasts with a prominent organoleptic profile, so these non-conventional yeasts should also present such metabolic pathways involved in the formation of this type of flavor compounds, such as the Ehrlich pathway, or the specific enzymes responsible for the synthesis of esters (Atf1p, Atf2p in S. cerevisiae). However, the regulation of their expression or the functionality of these enzymes may vary from non-Saccharomyces yeast to Saccharomyces yeast.
The study carried out by Postigo et al. [103] with 114 Saccharomyces cerevisiae yeast strains isolated from wine environment resulted in the production of aromatic compounds similar to those produced by commercial strains such as SafAle S-04 (Fermentis, Lesaffre, France). However, the relative proportions between the specific volatiles, or the flavor profiles, were different. This diversity of aromatic profiles was immense, and, interestingly, some of these strains could produce greater amounts of aromatic compounds than the commercial Saccharomyces strain. The strains studied were characterized by the production of higher concentrations of alcohols, such as isoamyl alcohol, isobutanol, and β-phenylethanol, as well as esters such as isoamyl acetate and ethyl butyrate compared to the commercial strain. On the other hand, it was also observed that some of these strains showed guaiacol production above the threshold (3.88 ppb) (smoked flavor) [104], giving a phenolic character to beers. A particularity of non-conventional yeasts is the production of POFs, generally unwanted in some beer styles but appreciated in wheat beers [52]. In other studies, such as the one carried out by Rossi et al. [105] with 12 yeast strains isolated from environments such as grape must, bakery, wine, and apple stillage showed behaviors similar to those observed in the previous study. Table 1 shows the different yeast strains isolated from non-conventional environments and the production of different aromatic by-products, the most important of which are shown below. Some of the strains studied exhibited fermentative behaviors close to commercial strains (Safbrew-S33 and Nottingham), with concentrations of aromatic compounds higher in some cases than those in commercial ones. Another peculiarity of the non-conventional yeasts of Saccharomyces is that there are some strains that will only ferment in the wort glucose, fructose, and sucrose but not maltose [58, 103]. This makes them ideal candidates for brewing low ethanol beers, as they can produce levels of aromatic compounds similar to those of other Saccharomyces strains that do ferment maltose, which can cause the characteristic wort flavor of some beers with low ethanol content [108, 109]. This also prevents the use of mechanical methods to remove ethanol content. Furthermore, residual sugars that may remain in beer can influence aspects such as the viscosity that contributes to the sensation in the mouth and body of beer [110, 111]. Likewise, this type of strains can be used in cofermentation with other strains of Saccharomyces to improve the aroma of beer [106].
Functional beer is defined as a beer that can provide health benefits under moderate consumption. Functional beers include those that have a low ethanol content, as well as those that provide high concentrations of compounds such as fiber, vitamins, minerals, polyphenols, and probiotics [1].
Recent years have seen an increase in consumption and interest in low-alcohol beers. This is mainly due to health and safety reasons, in addition to an increase in strict social regulations [112]. Low-ethanol beers can have health benefits due to the healthy components they contain, besides a lower energy intake and the total absence of negative effects of alcohol consumption.
Several studies have shown that with the use of non-conventional yeast, functional beers can be obtained, as they not only possess the ability to produce remarkable aromatic compounds, as well as other by-products such as melatonin [6, 113].
Functional beers also include beers that are gluten-free, thus covering consumers suffering from coeliac disease, which is a gluten-sensitive, immune-mediated enteropathy.
Probiotic beers are also included in functional beers. Probiotics include those live microorganisms that are added to food and that under certain doses can be potentially beneficial for human health, especially for the intestinal microbial balance [114]. Therefore, a probiotic beer is one obtained by using probiotic microorganisms during the fermentation process. The best known microorganisms used for their probiotic characteristics are lactic acid bacteria (Lactobacillus, Bifidobacterium, Enterococcus, or Streptococcus) [115]. However, studies have also been extended to other microorganisms such as yeasts, where experiments have been carried out with Saccharomyces cerevisiae var. boulardii, since it has been found to have the ability to prevent diarrhea associated with the use of antibiotics, as well as helping with infections caused by Escherichia coli or Clostridium difficile [116, 117]. In addition, beer trials have also been carried out with S. cerevisiae var. boulardii, where the results have shown an increase in the antioxidant capacity due to metabolite secretion by the yeasts in the craft beers obtained [118]. However, studies have focused not only on the study of non-conventional yeasts of the Saccharomyces genus but also on the use of other non-Saccharomyces genera (Lachancea, Kluyveromyces, Torulaspora, Metschnikowia, Kazachstania, Brettanomyces, Pichia, Candida, Hanseniaspora, Rhodotorula, Rodosporidobolus) since they have been found to have great potential for use in the production of craft beers with probiotic characteristics [119]. Craft beer is generally not unfiltered or unpasteurized; thus, the yeast cells remaining after fermentation can provide a probiotic character to the beer and can be considered beneficial to health. This could be seen in the studies carried out by Mulero et al. [118] comparing unfiltered beers fermented with S. cerevisiae var. boulardii and a commercial S. cerevisiae strain. According to the studies of Hill et al. [120], the minimum recommended dose of live probiotic cell count per product sample is 9 Log colony forming units (CFU). On the other hand, industrial beers, after being subjected to high temperatures during pasteurization, may eliminate such probiotics. For this reason, craft rather than industrial brewing would be more appropriate since viability is crucial for the efficacy of probiotics [106]. However, it should be noted that the presence of yeasts in beer can give beer a unique taste but reduce its shelf life as during storage, cell lysis can negatively affect its quality. This is why yeast strain selection is important in the production of craft beers [121].
6.1 Melatonin production
Melatonin is a mammalian hormone that regulates sleep and has antioxidant properties. It is produced by yeasts during fermentation and can therefore be a source of exogenous melatonin for the body, since as people age, less melatonin is produced in the body. In addition, it provides beer with antioxidant, antiaging, anti-inflammatory, antitumor, and immunomodulatory capacities [122, 123, 124].
Much of the food and beverages we consume on a daily basis contain melatonin. Therefore, their intake helps to increase the melatonin level in the body and its antioxidant status in human serum, which is the reason that this molecule is absorbed in the gastrointestinal tract [123, 125, 126] and readily crosses all morphophysiological barriers and tissue and cell membranes [124, 125, 126, 127, 128, 129]. Likewise, melatonin interacts with toxic reagents, generating other metabolites that are in turn direct free radical scavengers. The combined actions of melatonin and its derivatives greatly enhance the efficacy of melatonin in protecting against free radical damage and reducing the likelihood of human disease [130].
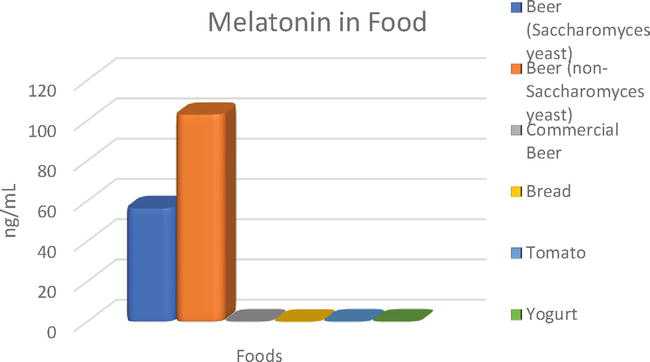
Melatonin is a by-product of yeast that is produced in the final stages of fermentation and is excreted into the medium during the stationary phase of the yeast fermentation [131]. In the studies carried out by Postigo et al. [103, 132] with different strains of wild Saccharomyces and non-Saccharomyces yeast, it was generally observed that Saccharomyces yeasts produce melatonin in a higher percentage than non-Saccharomyces yeasts. However, the concentration of non-Saccharomyces yeasts (ranged between 6.69 and 102.98 ng/mL) is in some cases double that of Saccharomyces yeasts (between 5.04 and 56.51 ng/mL). Furthermore, in studies carried out with sequential fermentations, where both yeast genera participate, melatonin concentrations were detected in almost all beers (values from 33.63 to 66.57 ng/mL) [133]. On the other hand, in the studies carried out by García-Moreno et al. [122], the melatonin values found in different commercial beers ranged from 58 to 169 pg/mL. As Valera et al. [134] determined in their studies, the presence of Saccharomyces seems to promote a higher presence of melatonin in the medium. In the studies carried out by Fernandez-Cruz et al. [135] at the intracellular level with wine yeasts, melatonin was only detected in non-Saccharomyces yeasts. Also, Juhnevica-Radenkova et al. [131] mention in their work that melatonin is produced in the final stages of fermentation and is excreted into the medium during the stationary phase of the yeast fermentation. In comparison with other types of food such as bread, tomato, or yogurt, we can find different melatonin values of 28.9, 138.1, and 126.7 pg/mL, respectively, being therefore lower than what we could find in some beers fermented with non-conventional yeasts [136]. The comparison of the different concentrations can be seen in Figure 1.
Figure 1.
Melatonin concentrations found in different foods. Beer (Saccharomyces yeast) [103], beer (non-Saccharomyces yeast) [132], commercial beer [122], bread, tomato, and yogurt [136].
These levels of melatonin that can be found in various foods, as well as in beer, are concentrations below those studied that have supposed positive effects on health (1–10 mg) [137, 138]; however, if ingested together with other foods, they can contribute to increase its concentration in the human serum. Studies carried out by Maldonado et al. [123] determined that the intake of beer with high melatonin content (169.7 pg/ml) contributed to increase the antioxidant properties of human serum. However, although melatonin can act as a strong antioxidant, it can be degraded in the presence of oxygen, light, or free radicals that are present during the aging process [139]. This fact could be observed in the studies of lambic beer carried out by Postigo et al. [140], where the analysis at different maturation times of the lambic beers brewed determined their degradation over time. It should also be taken into account whether the final product is subjected to final heat treatments to prolong the stability of the product (such as pasteurization), since it has been shown that high temperatures can also degrade it and reduce its concentration in food [136].
6.2 Antioxidant capacity
Different compounds such as phenolics can be found in beer [141]. These substances can also be found naturally in fruits, vegetables, nuts, seeds, and beverages [142]. When studying the antioxidant fraction of beer, phenolic compounds are the most studied, which are found in hops (20–30%) as well as in malt (70–80%) [141]. Hops provide the beer with phenolic acids, prenylated chalcones, flavonoids, catechins, and proanthocyanidins [141]. Malt contains an overall phenolic mass of 1.0–1.9 mg g−1 dry matter [143]. Likewise, the yeast used in brewing can also influence the phenolic composition and antioxidant capacity of the final product [106]. Several studies, such as the one carried out by Viana et al. [144], showed that the use of certain yeast strains for the production of Pale Ale beer significantly influences the antioxidant capacity of the beers.
Antioxidant capacity is related to parameters such as total phenolic and flavonoid content; 2,2-Diphenyl-1-picrylhydrazyl (DPPH) radical scavenging activity; and 2,2′-azinobis-(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) radical cation scavenging activity [145]. In the studies carried out by Postigo et al. [103] with different strains of wild yeast, it was observed that there were no major differences between the yeasts of Saccharomyces (9.50 to 13.67 mmol TE/L) and the commercial strain S-04 (11.18 mmol TE/L), as well as in sequential fermentations (9.63 to 13.70 mmol TE/L) (Table 2). However, in those strains where maltose was not fermented, or where there was competition between species in sequential culture, the antioxidant capacity was lower [133]. In contrast, in the studies carried out by Granato et al. [113], the values found in beers (ale and lager) were lower, between 424.77 and 10,508.47 μmol TE/L. These antioxidant capacity values may also vary depending on the method used for their analysis, since by the Oxygen Radical Absorption Capacity (ORAC) method in another study, values between 3.70 and 29.11 mmol TE/L were obtained. The ORAC method is based on the absorption capacity of oxygen radicals, whereby the decrease in fluorescence emission is measured [146, 147]. Another method used to determine the antioxidant capacity is that using the ferric-reducing antioxidant power, whose principle is the determination of the reduction of a ferric-tripyridyltriazine complex to its ferrous form, colored, in the presence of antioxidant components, where values of 3125 μmol Fe2+/L can be found in ale beer.
The use of non-conventional Saccharomyces yeasts provides beer with additional natural flavor variants that differentiate it from other commercial beers. These characteristics can be obtained either in pure culture, in sequential culture, or with aeration during the fermentation, which makes them very versatile. Furthermore, such wild yeasts can provide bio-healthy properties to beer under moderate consumption, as they may present production of compounds such as melatonin, with concentrations higher than those of commercial strains and even those of other foods.
1.Yeo HQ , Liu SQ. An overview of selected specialty beers: Developments, challenges and prospects. International Journal of Food Science and Technology. 2014;49(7):1607-1618. DOI: 10.1111/ijfs.12488
2.Steensels J, Verstrepen KJ. Taming wild yeast: Potential of conventional and nonconventional yeasts in industrial fermentations. Annual Review of Microbiology. 2014;68(1):61-80. DOI: 10.1146/annurev-micro-091213-113025
3.James SA, Stratford M. Zygosaccharomyces. In: The Yeasts. London: Elsevier; 2011. pp. 937-947. DOI: 10.1016/B978-0-444-52149-1.00084-7
4.Lentz M, Putzke T, Hessler R, Luman E. Genetic and physiological characterization of yeast isolated from ripe fruit and analysis of fermentation and brewing potential. Journal of the Institute of Brewing. 2014;120(4):559-564. DOI: 10.1002/jib.154
5.Petruzzi L, Rosaria Corbo M, Sinigaglia M, Bevilacqua A. Brewer’s yeast in controlled and uncontrolled fermentations, with a focus on novel, nonconventional, and superior strains. Food Review International. 2016;32(4):341-363. DOI: 10.1080/87559129.2015.1075211
6.Basso RF, Alcarde AR, Portugal CB. Could non-Saccharomyces yeasts contribute on innovative brewing fermentations? Food Research International. 2016;86:112-120. DOI: 10.1016/j.foodres.2016.06.002
7.Vanderhaegen B, Neven H, Coghe S, Verstrepen KJ, Derdelinckx G, Verachtert H. Bioflavoring and beer refermentation. Applied Microbiology and Biotechnology. 2003;62(2-3):140-150. DOI: 10.1007/s00253-003-1340-5
8.Piskur J, Rozpedowska E, Polakova S, Merico A, Compagno C. How did Saccharomyces evolve to become a good brewer? Trends in Genetics. 2006;22(4):183-186. DOI: 10.1016/j.tig.2006.02.002
9.Lodolo EJ, Kock JLF, Axcell BC, Brooks M. The yeast Saccharomyces cerevisiae-the main character in beer brewing. FEMS Yeast Research. 2008;8(7):1018-1036. DOI: 10.1111/j.1567-1364.2008.00433.x
10.Pires EJ, Teixeira JA, Brányik T, Vicente AA. Yeast: The soul of beer’s aroma—A review of flavour-active esters and higher alcohols produced by the brewing yeast. Applied Microbiology and Biotechnology. 2014;98(5):1937-1949. DOI: 10.1007/s00253-013-5470-0
11.Verstrepen KJ, Derdelinckx G, Pierre DJ, Winderickx J, Thevelein JM, Pretorius IS, et al. Flavor-active esters: Adding fruitiness to beer. Journal of Bioscience and Bioengineering. 2003;96(2):110-118. DOI: 10.1016/S1389-1723(03)90112-5
12.Procopio S, Qian F, Becker T. Function and regulation of yeast genes involved in higher alcohol and ester metabolism during beverage fermentation. European Food Research and Technology. 2011;233(5):721-729. DOI: 10.1007/s00217-011-1567-9
13.Procopio S, Brunner M, Becker T. Differential transcribed yeast genes involved in flavour formation and its associated amino acid metabolism during brewery fermentation. European Food Research and Technology. 2014;239(3):421-439. DOI: 10.1007/s00217-014-2236-6
14.Steensels J, Snoek T, Meersman E, Picca Nicolino M, Aslankoohi E, Christiaens JF, et al. Selecting and generating superior yeasts for the brewing industry. Cerevisia. 2012;37(2):63-67. DOI: 10.1016/j.cervis.2012.08.001
15.Verstrepen KJ, Derdelinckx G, Delvaux FR. Esters in beer-part 1: The fermentation process: More than ethanol formation. Cerevisia. 2003;28(3):41-49
16.Meilgaard MC. Flavor chemistry in beer: Part I: Flavor interaction between principal volatiles. Technical Quarterly—Master Brewers Association. 1975;12(2):107-117
17.Kollmannsberger H, Biendl M, Nitz S. Occurrence of glycosidically bound flavour compounds in hops, hop products and beer. Monatsschrift fur Brauwissenschaft. 2006;5(6):83-89
18.Bogdan P, Kordialik-Bogacka E. Alternatives to malt in brewing. Trends in Food Science and Technology. 2017;65:1-9. DOI: 10.1016/j.tifs.2017.05.001
19.Nelson M. The Barbarian’s Beverage [Internet]. New York: Routledge; 2005. DOI: 10.4324/9780203309124
20.Belitz H, Grosch W. Alcoholic beverages. In: Food Chemistry. Berlin, Heidelberg: Springer Berlin Heidelberg; 2004. pp. 892-937. DOI: 10.1007/978-3-540-69934-7_21
21.Eumann M. Water in brewing. In: Brewing. London: Elsevier; 2006. pp. 183-207. DOI: 10.1533/9781845691738.183
22.Niu C, Han Y, Wang J, Zheng F, Liu C, Li Y, et al. Malt derived proteins: Effect of protein Z on beer foam stability. Food Bioscience. 2018;25:21-27. DOI: 10.1016/j.fbio.2018.07.003
23.Parr H, Bolat I, Cook D. Modelling flavour formation in roasted malt substrates under controlled conditions of time and temperature. Food Chemistry. 2021;337:127641. DOI: 10.1016/j.foodchem. 2020.127641
24.Kishimoto T, Teramoto S, Fujita A, Yamada O. Evaluation of components contributing to the international bitterness unit of wort and beer. Journal of the American Society of Brewing Chemists. 2022;80(1):53-61. DOI: 10.1080/03610470.2021.1878684
25.Astray G, Gullón P, Gullón B, Munekata PES, Lorenzo JM. Humulus lupulus L. as a natural source of functional biomolecules. Applied Sciences. 2020;10(15):5074. DOI: 10.3390/app10155074
26.Barnett JA. Beginnings of microbiology and biochemistry: The contribution of yeast research. Microbiology. 2003;149(3):557-567. DOI: 10.1099/mic.0.26089-0
27.Barnett JA. A history of research on yeasts 2: Louis Pasteur and his contemporaries, 1850-1880. Yeast. 2000;16(8):755-771. DOI: 10.1002/1097-0061 (20000615)16:8<755::AID-YEA587>3.0.CO;2-4
28.Meussdoerffer FG. A comprehensive history of beer brewing. In: Handbook of Brewing. Weinheim, Germany: Wiley-VCH Verlag GmbH & Co. KGaA; 2009. pp. 1-42. DOI: 10.1002/ 9783527623488.ch1
29.Michel RH, McGovern PE, Badler VR. Chemical evidence for ancient beer. Nature. 1992;360(6399):24-24
30.Goddard MR, Greig D. Saccharomyces cerevisiae: A nomadic yeast with no niche? FEMS Yeast Research. 2015;15(3):1-6
31.Warringer J, Zörgö E, Cubillos FA, Zia A, Gjuvsland A, Simpson JT, et al. Trait variation in yeast is defined by population history. PLoS Genetics. 2011;7(6):e1002111
32.Driscoll CA, Macdonald DW, O’Brien SJ. An evolutionary view of domestication. PNAS. 2009;106:9971-9978
33.Purugganan MD, Fuller DQ. The nature of selection during plant domestication. Nature. 2009;457(7231):843-848
34.Marsit S, Mena A, Couloux A, Guy J, Luc LJ, Barrio E, et al. Evolutionary advantage conferred by an eukaryote-to-eukaryote gene transfer event in wine yeasts. Molecular Biology and Evolution. 2015;32(7):1695-1707
35.Marsit S, Sanchez I, Galeote V. Horizontally acquired oligopeptide transporters favour adaptation of Saccharomyces cerevisiae wine yeast to oenological environment. Environmental Microbiology. 2016;18:1148-1161
36.Almeida P, Barbosa R, Zalar P, Imanishi Y, Shimizu K, Turchetti B, et al. A population genomics insight into the Mediterranean origins of wine yeast domestication. Molecular Ecology. 2015;24(21):5412-5427
37.Marsit S, Dequin S, Montpellier F, Supagro M, Montpellier F, Montpellier F. Diversity and adaptive evolution of Saccharomyces wine yeast: A review. FEMS Yeast Research. 2015;15:fov067
38.Dunn B, Richter C, Kvitek DJ, Pugh T, Sherlock G. Analysis of the Saccharomyces cerevisiae pan-genome reveals a pool of copy number variants distributed in diverse yeast strains from differing industrial environments. Genome Research. 2012;22:908-924
39.Steenwyk J, Rokas A. Extensive copy number variation in fermentation-related genes among Saccharomyces cerevisiae wine strains. G3 (Bethesda). 2017;7(May):1475-1485
40.Strope PK, Skelly DA, Kozmin SG, Mahadevan G, Stone EA, Magwene PM, et al. The 100-genomes strains, an S. cerevisiae resource that illuminates its natural phenotypic and genotypic variation and emergence as an opportunistic pathogen. Genome Research. 2015;25:762-774
41.Almeida P, Barbosa R, Bensasson D, Lisboa UN De. Adaptive divergence in wine yeasts and their wild relatives, Molecular Ecology. 2017;26(7):2167-2182
42.CJS G. Economics of the craft beer revolution: A comparative international perspective. In: Garavaglia C, Swinnen J, editors. Economic Perspectives on Craft Beer. Cham: Springer International Publishing; 2018. pp. 3-54. DOI: 10.1007/978-3-319-58235-1
43.Aquilani B, Laureti T, Poponi S, Secondi L. Beer choice and consumption determinants when craft beers are tasted: An exploratory study of consumer preferences. Food Quality and Preference. 2015;41:214-224. DOI: 10.1016/j.foodqual.2014.12.005
44.Hittinger CT, Steele JL, Ryder DS. Diverse yeasts for diverse fermented beverages and foods. Current Opinion in Biotechnology. 2018;49:199-206. DOI: 10.1016/j.copbio.2017.10.004
45.Gibson B, Geertman JMA, Hittinger CT, Krogerus K, Libkind D, Louis EJ, et al. New yeasts-new brews: Modern approaches to brewing yeast design and development. FEMS Yeast Research. 2017;17(4):1-13
46.De FG, Sannino C, Sileoni V, Marconi O, Filippucci S, Tasselli G, et al. Mrakia Gelida in brewing process: An innovative production of low alcohol beer using a psychrophilic yeast strain. Journal of Food Microbiology. 2018;76(April):354-362. DOI: 10.1016/j.fm.2018.06.018
47.Domizio P, House JF, Joseph CML, Bisson LF, Bamforth CW. Lachancea thermotolerans as an alternative yeast for the production of beer. Journal of the Institute of Brewing. 2016;122(4):599-604. DOI: 10.1002/jib.362
48.Michel M, Kopecká J, Meier- Dörnberg T, Zarnkow M, Jacob F, Hutzler M. Screening for new brewing yeasts in the non-Saccharomyces sector with Torulaspora delbrueckii as model. Yeast. 2016;33(4):129-144. DOI: 10.1002/yea.3146
49.Gallone B, Mertens S, Gordon JL, Maere S, Verstrepen KJ, Steensels J. Origins, evolution, domestication and diversity of Saccharomyces beer yeasts. Current Opinion in Biotechnology. 2018;49:148-155. DOI: 10.1016/j.copbio.2017.08.005
50.Nikulin J, Krogerus K, Gibson B. Alternative Saccharomyces interspecies hybrid combinations and their potential for low-temperature wort fermentation. Yeast. 2018;35(1):113-127. DOI: 10.1002/yea.3246
51.Coghe S, Benoot K, Delvaux F, Vanderhaegen B, Delvaux FR. Ferulic acid release and 4-vinylguaiacol formation during brewing and fermentation: Indications for feruloyl esterase activity in Saccharomyces cerevisiae. Journal of Agricultural and Food Chemistry. 2004;52(3):602-608. DOI: 10.1021/jf0346556
52.Gallone B, Steensels J, Prahl T, Soriaga L, Saels V, Herrera-Malaver B, et al. Domestication and divergence of Saccharomyces cerevisiae beer yeasts. Cell. 2016;166(6):1397-1410. DOI: 10.1016/j.cell.2016.08.020
53.Cubillos FA, Gibson B, Grijalva- Vallejos N, Krogerus K, Nikulin J. Bioprospecting for brewers: Exploiting natural diversity for naturally diverse beers. Yeast. 2019;36(6):383-398. DOI: 10.1002/yea.3380
54.Rodrigues AJ, Raimbourg T, Gonzalez R, Morales P. Environmental factors influencing the efficacy of different yeast strains for alcohol level reduction in wine by respiration. LWT. 2016;65:1038-1043. DOI: 10.1016/j.lwt.2015.09.046
55.Kirsop BH. Oxygen in brewery fermentation. Journal of the Institute of Brewing. 1974;80(3):252-259. DOI: 10.1002/j.2050-0416.1974.tb03614.x
56.Virkajärvi I, Lindborg K, Jukka Kronlöf EP. Effects of aeration on flavor compounds in immobilized primary fermentation. Monatsschrift für Brauwiss. 1999;52(1):9-28
57.Kucharczyk K, Tuszyński T. The effect of wort aeration on fermentation, maturation and volatile components of beer produced on an industrial scale. Journal of the Institute of Brewing. 2017;123(1):31-38. DOI: 10.1002/jib.392
58.Postigo V, O’Sullivan T, Elink Schuurman T, Arroyo T. Non-conventional yeast: Behavior under pure culture, sequential and aeration conditions in beer fermentation. Food. 2022;11(22):3717. DOI: 10.3390/foods11223717
59.Fukuhara H. The Kluyver effect revisited. FEMS Yeast Research. 2003;3(4):327-331. DOI: 10.1016/S1567-1356(03)00112-0
60.Sims AP, Barnett JA. The requirement of oxygen for the utilization of maltose, cellobiose and D-galactose by certain anaerobically fermenting yeasts (Kluyver effect). Journal of General Microbiology. 1978;106(2):277-288. DOI: 10.1099/00221287-106-2-277
61.Mauricio JC, Moreno J, Zea L, Ortega JM, Medina M. The effects of grape must fermentation conditions on volatile alcohols and esters formed by Saccharomyces cerevisiae. Journal of the Science of Food and Agriculture. 1997;75(2):155-160. DOI: 10.1002/(SICI)1097-0010(199710)75:2<155::AID-JSFA853>3.0.CO;2-S
62.Saerens SMG, Delvaux FR, Verstrepen KJ, Thevelein JM. Production and biological function of volatile esters in Saccharomyces cerevisiae. Microbial Biotechnology. 2010;3(2):165-177. DOI: 10.1111/j.1751-7915.2009.00106.x
63.Vanderhaegen B, Neven H, Coghe S, Verstrepen KJ, Verachtert H, Derdelinckx G. Evolution of chemical and sensory properties during aging of top-fermented beer. Journal of Agricultural and Food Chemistry. 2003;51(23):6782-6790. DOI: 10.1021/jf034631z
64.Meilgaard MC, Dalgliesh CE, Clapperton JF. Beer flavour terminology. Journal of the Institute of Brewing. 1979;85(1):38-42. DOI: 10.1002/j.2050-0416.1979.tb06826.x
65.Dvořák J, Dostálek P, Štěrba K, Čejka P, Kellner V, Čulík J, et al. Determination of total sulphur dioxide in beer samples by flow-through chronopotentiometry. Journal of the Institute of Brewing. 2006;112(4):308-313. DOI: 10.1002/j.2050-0416.2006.tb00736.x
66.Nagami K. Hydrogen sulfide in brewing. MBAA Technical Quarterly. 1980;17:64-68
67.Guido LF. Sulfites in beer: Reviewing regulation, analysis and role. Science in Agriculture. 2016;73(2):189-197. DOI: 10.1590/0103-9016-2015-0290
68.Meilgaard MC. Individual differences in sensory threshold for aroma chemicals added to beer. Food Quality and Preference. 1993;4(3):153-167. DOI: 10.1016/0950-3293(93)90158-3
69.Ilet DR. Aspects of the analysis, role, and fate of sulphur dioxide in beer—A review. Tech Quaterly. Master Brewers Association of the America. 1995;32:213-221
70.Oka K, Hayashi T, Matsumoto N, Yanase H. Decrease in hydrogen sulfide content during the final stage of beer fermentation due to involvement of yeast and not carbon dioxide gas purging. Journal of Bioscience and Bioengineering. 2008;106(3):253-257. DOI: 10.1263/jbb.106.253
71.Anderson RJ, Howard GA. The origin and occurrence of volatile sulphur compounds in British ales and lagers. Journal of the Institute of Brewing. 1974;80(4):357-370. DOI: 10.1002/j.2050-0416.1974.tb03630.x
72.Meilgaard M, Elizondo A, Moya E. A study of carbonyl compounds in beer, part II. Flavor and flavor thresholds of aldehydes and ketones added to beer. Technical Quarterly - Master Brewers Association. 1970;7(3):143-149
73.Krogerus K, Gibson BR. 125th anniversary review: Diacetyl and its control during brewery fermentation. Journal of the Institute of Brewing. 2013;119(3):86-97. DOI: 10.1002/jib.84
74.Ryan ED, Kohlhaw GB. Subcellular localization of isoleucine-valine biosynthetic enzymes in yeast. Journal of Bacteriology. 1974;120(2):631-637. DOI: 10.1128/jb.120.2.631-637.1974
75.Liu SQ. Impact of yeast and bacteria on beer appearance and flavor [Internet]. In: Hill AE, editor. Brewing Microbiology. Oxford, UK: Woodhead Publishing; 2015. pp. 357-374. DOI: 10.1016/B978-1-78242-331-7.00017-4
76.Canonico L, Agarbati A, Comitini F, Ciani M. Torulaspora delbrueckii in the brewing process: A new approach to enhance bioflavour and to reduce ethanol content. Food Microbiology. 2016;56:45-51. DOI: 10.1016/j.fm.2015.12.005
77.Eßlinger HM. Handbook of Brewing Processes Technology Markets-Wiley. Weinheim: Wiley; 2009. p. 779
78.Thurston P. The phenolic off-flavour test: A method for confirming the presence of wild yeasts. Journal of the Institute of Brewing. 1986;92(1):9-10
79.Scholtes C, Nizet S, Collin S. Guaiacol and 4-Methylphenol as specific markers of torrefied malts. Fate of volatile phenols in special beers through aging. Journal of Agricultural and Food Chemistry. 2014;62(39):9522-9528. DOI: 10.1021/jf5015654
80.Vanbeneden N, Van Roey T, Willems F, Delvaux F, Delvaux FR. Release of phenolic flavour precursors during wort production: Influence of process parameters and grist composition on ferulic acid release during brewing. Food Chemistry. 2008;111(1):83-91. DOI: 10.1016/j.foodchem.2008.03.029
81.Thurston PA, Tubb RS. Screening yeast strains for their ability to produce phenolic off-flavours: A simple method for determining phenols in wort and beer. Journal of the Institute of Brewing. 1981;87(3):177-179. DOI: 10.1002/j.2050-0416.1981.tb04012.x
82.Rodrigues JEA, Erny GL, Barros AS, Esteves VI, Brandão T, Ferreira AA, et al. Quantification of organic acids in beer by nuclear magnetic resonance (NMR)-based methods. Analytica Chimica Acta. 2010;674(2):166-175. DOI: 10.1016/j.aca.2010.06.029
83.Clapperton JF, Brown DGW. Caprylic flavour as a feature of beer flavour. Journal of the Institute of Brewing. 1978;84(2):90-92. DOI: 10.1002/j.2050-0416.1978.tb03844.x
84.Siebert KJ. Modeling the flavor thresholds of organic acids in beer as a function of their molecular properties. Food Quality and Preference. 1999;10(2):129-137. DOI: 10.1016/S0950-3293(98)00059-7
85.Montanari L, Perretti G, Natella F, Guidi A, Fantozzi P. Organic and phenolic acids in beer. LWT - Food Science and Technology. 1999;32(8):535-539. DOI: 10.1006/fstl.1999.0593
86.Amata AI, Germain P. The effect of pitching yeast aeration on the production of acetic acid during fermentations with brewers’ yeast: An enzymatic approach. Journal of the Institute of Brewing. 1990;96(3):131-134. DOI: 10.1002/j.2050-0416.1990.tb01023.x
87.Li H, Liu F. The chemistry of sour taste and the strategy to reduce the sour taste of beer. Food Chemistry. 2015;185:200-204. DOI: 10.1016/j.foodchem.2015.03.135
88.Coote N, Kirsop BH. The content of some organic acids in beer and other fermented media. Journal of the Institute of Brewing. 1974;80(5):474-483. DOI: 10.1002/j.2050-0416.1974.tb06797.x
89.Eden A, Simchen G, Benvenisty N. Two yeast homologs of ECA39, a target for c-myc regulation, code for cytosolic and mitochondrial branched-chain amino acid aminotransferases. The Journal of Biological Chemistry. 1996;271(34):20242-20245. DOI: 10.1074/jbc.271.34.20242
90.Hazelwood LA, Daran JM, van Maris AJA, Pronk JT, Dickinson JR. The ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Applied and Environmental Microbiology. 2008;74(8):2259-2266. DOI: 10.1128/AEM.02625-07
91.Ehrlich F. Über die Bedingungen der Fuselölbildung und über ihren Zusammenhang mit dem Eiweissaufbau der Hefe. Berichte der Dtsch Chem Gesellschaft. 1907;40(1):1027-1047
92.Engan S. Organoleptic threshold values of some organic acids in beer. Journal of the Institute of Brewing. 1974;80(2):162-163. DOI: 10.1002/j.2050-0416.1974.tb03598.x
93.Engan S. Organoleptic threshold values of some alcohols and esters in beer. Journal of the Institute of Brewing. 1972;78(1):33-36. DOI: 10.1002/j.2050-0416.1972.tb03425.x
94.Peddie HA. Ester formation in brewery fermentations. Journal of the Institute of Brewing. 1990;96(5):327-331. DOI: 10.1002/j.2050-0416.1990.tb01039.x
95.Ciani M, Comitini F, Mannazzu I, Domizio P. Controlled mixed culture fermentation: A new perspective on the use of non-Saccharomyces yeasts in winemaking. FEMS Yeast Research. 2010;10(2):123-133. DOI: 10.1111/j.1567-1364.2009.00579.x
96.Ciani M, Comitini F. Non-Saccharomyces wine yeasts have a promising role in biotechnological approaches to winemaking. Annales de Microbiologie. 2011;61(1):25-32. DOI: 10.1007/s13213-010-0069-5
97.Jolly NP, Varela C, Pretorius IS. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Research. 2014;14(2):215-237. DOI: 10.1111/1567-1364.12111
98.Michel M, Meier-Dörnberg T, Jacob F, Methner F, Wagner RS, Hutzler M. Review: Pure non-Saccharomyces starter cultures for beer fermentation with a focus on secondary metabolites and practical applications. Journal of the Institute of Brewing. 2016;122(4):569-587. DOI: 10.1002/jib.381
99.Carrau FM, Medina K, Boido E, Farina L, Gaggero C, Dellacassa E, et al. De novo synthesis of monoterpenes by Saccharomyces cerevisiae wine yeasts. FEMS Microbiology Letters. 2005;243(1):107-115. DOI: 10.1016/j.femsle.2004.11.050
100.Stribny J, Gamero A, Pérez-Torrado R, Querol A. Saccharomyces kudriavzevii and Saccharomyces uvarum differ from Saccharomyces cerevisiae during the production of aroma-active higher alcohols and acetate esters using their amino acidic precursors. International Journal of Food Microbiology. 2015;205:41-46. DOI: 10.1016/j.ijfoodmicro.2015.04.003
101.Janssens L, De Pooter HL, Schamp NM, Vandamme EJ. Production of flavours by microorganisms. Process Biochemistry. 1992;27(4):195-215. DOI: 10.1016/0032-9592(92)80020-4
102.Gamero A, Quintilla R, Groenewald M, Alkema W, Boekhout T, Hazelwood L. High-throughput screening of a large collection of non-conventional yeasts reveals their potential for aroma formation in food fermentation. Food Microbiology. 2016;60:147-159. DOI: 10.1016/j.fm.2016.07.006
103.Postigo V, García M, Cabellos JM, Arroyo T. Wine Saccharomyces yeasts for beer fermentation. Fermentation. 2021;7(4):290. DOI: 10.3390/fermentation7040290
104.Sterckx FL, Missiaen J, Saison D, Delvaux FR. Contribution of monophenols to beer flavour based on flavour thresholds, interactions and recombination experiments. Food Chemistry. 2011;126(4):1679-1685. DOI: 10.1016/j.foodchem.2010.12.055
105.Rossi S, Turchetti B, Sileoni V, Marconi O, Perretti G. Evaluation of Saccharomyces cerevisiae strains isolated from non-brewing environments in beer production. Journal of the Institute of Brewing. 2018;124(4):381-388. DOI: 10.1002/jib.503
106.Capece A, Romaniello R, Pietrafesa A, Siesto G, Pietrafesa R, Zambuto M, et al. Use of Saccharomyces cerevisiae var. boulardii in co-fermentations with S. Cerevisiae for the production of craft beers with potential healthy value-added. International Journal of Food Microbiology. 2018;284(March):22-30. DOI: 10.1016/j.ijfoodmicro.2018.06.028
107.Preiss R, Tyrawa C, Krogerus K, Garshol LM, Van Der Merwe G. Traditional Norwegian Kveik are a genetically distinct group of domesticated Saccharomyces cerevisiae brewing yeasts. Frontiers in Microbiology. 2018;9:2137
108.De Francesco G, Turchetti B, Sileoni V, Marconi O, Perretti G. Screening of new strains of Saccharomycodes ludwigii and Zygosaccharomyces rouxii. Journal of the Institute of Brewing. 2015;121(1):113-121. DOI: 10.1002/jib.185
109.Saison D, De Schutter DP, Vanbeneden N, Daenen L, Delvaux F, Delvaux FR. Decrease of aged beer aroma by the reducing activity of brewing yeast. Journal of Agricultural and Food Chemistry. 2010;58(5):3107-3115. DOI: 10.1021/jf9037387
110.Hughes PS, Baxter ED. Beer: Quality, Safety and Nutritional Aspects. Cambridge: Royal Society of Chemistry; 2001. pp. 40-73
111.Langstaff SA, Lewis MJ. The mouthfeel of beer—A review. Journal of the Institute of Brewing. 1993;99(1):31-37. DOI: 10.1002/j.2050-0416.1993.tb01143.x
112.Brányik T, Silva DP, Baszczyňski M, Lehnert R, Almeidae Silva JB. A review of methods of low alcohol and alcohol-free beer production. Journal of Food Engineering. 2012;108(4):493-506. DOI: 10.1016/j.jfoodeng.2011.09.020
113.Granato D, Branco GF, Faria JDAF, Cruz AG. Characterization of Brazilian lager and brown ale beers based on color, phenolic compounds, and antioxidant activity using chemometrics. Journal of the Science of Food and Agriculture. 2011;91(3):563-571. DOI: 10.1002/jsfa.4222
114.Moslehi-Jenabian S, Lindegaard L, Jespersen L. Beneficial effects of probiotic and food borne yeasts on human health. Nutrients. 2010;2(4):449-473. DOI: 10.3390/nu2040449
115.Butel MJ. Probiotics, gut microbiota and health. Médecine et Maladies Infectieuses. 2014;44(1):1-8. DOI: 10.1016/j.medmal.2013.10.002
116.Buts JP. Twenty-five years of research on saccharomyces boulardii trophic effects: Updates and perspectives. Digestive Diseases and Sciences. 2009;54(1):15-18. DOI: 10.1007/s10620-008-0322-y
117.Im E, Pothoulakis C. Progrès récents dans la recherche sur Saccharomyces boulardii. Gastroentérologie Clinique et Biologique. 2010;34(SUPPL. 1):S62-S70. DOI: 10.1016/S0399-8320(10)70023-3
119.Canonico L, Zannini E, Ciani M, Comitini F. Assessment of non-conventional yeasts with potential probiotic for protein-fortified craft beer production. LWT. 2021;145(March):111361. DOI: 10.1016/j.lwt.2021.111361
120.Hill C, Guarner F, Reid G, Gibson GR, Merenstein DJ, Pot B, et al. The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nature reviews. Gastroenterology & Hepatology. 2014;11(8):506-514. DOI: 10.1038/nrgastro.2014.66
121.Iorizzo M, Coppola F, Letizia F, Testa B, Sorrentino E. Role of yeasts in the brewing process: Tradition and innovation. PRO. 2021;9(5):839. DOI: 10.3390/pr9050839
122.Garcia-Moreno H, Calvo JR, Maldonado MD. High levels of melatonin generated during the brewing process. Journal of Pineal Research. 2013;55(1):26-30. DOI: 10.1111/jpi.12005
123.Maldonado MD, Moreno H, Calvo JR. Melatonin present in beer contributes to increase the levels of melatonin and antioxidant capacity of the human serum. Clinical Nutrition. 2009;28(2):188-191. DOI: 10.1016/j.clnu.2009.02.001
124.Rodriguez-Naranjo MI, Gil-Izquierdo A, Troncoso AM, Cantos-Villar E, Garcia-Parrilla MC. Melatonin is synthesised by yeast during alcoholic fermentation in wines. Food Chemistry. 2011;126(4):1608-1613. DOI: 10.1016/j.foodchem.2010.12.038
125.Benot S, Gobema R, Reiter RJ, Garcia-Mauriño S, Osuna C, Guerrero JM. Physiological levels of melatonin contribute to the antioxidant capacity of human serum. Journal of Pineal Research. 1999;27(1):59-64. DOI: 10.1111/j.1600-079X.1999.tb00597.x
126.Reiter RJ, Tan DX, Maldonado MD. Melatonin as an antioxidant: Physiology versus pharmacology. Journal of Pineal Research. 2005;39(2):215-216. DOI: 10.1111/j.1600-079X.2005.00261.x
127.Ramakrishna A, Giridhar P, Sankar KU, Ravishankar GA. Melatonin and serotonin profiles in beans of coffea species. Journal of Pineal Research. 2012;52(4):470-476. DOI: 10.1111/j.1600-079X.2011.00964.x
128.Rodriguez-Naranjo MI, Gil-Izquierdo A, Troncoso AM, Cantos E, Garcia-Parrilla MC. Melatonin: A new bioactive compound in wine. Journal of Food Composition and Analysis. 2011;24(4-5):603-608. DOI: 10.1016/j.jfca.2010.12.009
129.Venegas C, García JA, Escames G, Ortiz F, López A, Doerrier C, et al. Extrapineal melatonin: Analysis of its subcellular distribution and daily fluctuations. Journal of Pineal Research. 2012;52(2):217-227. DOI: 10.1111/j.1600-079X.2011.00931.x
130.Sanchez-Barcelo EJ, Mediavilla MD, Tan DX, Reiter RJ. Clinical uses of melatonin: Evaluation of human trials. Current Medicinal Chemistry. 2010;17(19):2070-2095. DOI: 10.2174/092986710791233689
131.Juhnevica-Radenkova K, Moreno DA, Ikase L, Drudze I, Radenkovs V. Naturally occurring melatonin: Sources and possible ways of its biosynthesis. Comprehensive Reviews in Food Science and Food Safety. 2020;19(6):4008-4030. DOI: 10.1111/1541-4337.12639
132.Postigo V, Sánchez A, Cabellos JM, Arroyo T. New approaches for the fermentation of beer: Non-Saccharomyces yeasts from wine. Fermentation. 2022;8(6):280. DOI: 10.3390/fermentation8060280
133.Postigo V, Sanz P, García M, Arroyo T. Impact of non-Saccharomyces wine yeast strains on improving healthy characteristics and the sensory profile of beer in sequential fermentation. Food. 2022;11(14):2029. DOI: 10.3390/foods11142029
134.Valera MJ, Morcillo-Parra MÁ, Zagórska I, Mas A, Beltran G, Torija MJ. Effects of melatonin and tryptophol addition on fermentations carried out by Saccharomyces cerevisiae and non-Saccharomyces yeast species under different nitrogen conditions. International Journal of Food Microbiology. 2019;289:174-181. DOI: 10.1016/j.ijfoodmicro.2018.09.013
135.Fernández-Cruz E, Álvarez-Fernández MA, Valero E, Troncoso AM, García-Parrilla MC. Melatonin and derived l-tryptophan metabolites produced during alcoholic fermentation by different wine yeast strains. Food Chemistry. 2017;217:431-437. DOI: 10.1016/j.foodchem.2016.08.020
136.Kocadağlı T, Yılmaz C, Gökmen V. Determination of melatonin and its isomer in foods by liquid chromatography tandem mass spectrometry. Food Chemistry. 2014;153:151-156. DOI: 10.1016/j.foodchem.2013.12.036
138.Koziróg M, Poliwczak AR, Duchnowicz P, Koter-Michalak M, Sikora J, Broncel M. Melatonin treatment improves blood pressure, lipid profile, and parameters of oxidative stress in patients with metabolic syndrome. Journal of Pineal Research. 2011;50(3):261-266. DOI: 10.1111/j.1600-079X.2010.00835.x
139.Que Z, Ma T, Shang Y, Ge Q , Zhang Q , Xu P, et al. Microorganisms: Producers of melatonin in fermented foods and beverages. Journal of Agricultural and Food Chemistry. 2020;68(17):4799-4811. DOI: 10.1021/acs.jafc.0c01082
140.Postigo V, García M, Arroyo T. Study of a first approach to the controlled fermentation for lambic beer production. Microorganisms. 2023;11(7):1681. DOI: 10.3390/microorganisms11071681
141.Gerhäuser C. Beer constituents as potential cancer chemopreventive agents. European Journal of Cancer. 2005;41(13):1941-1954. DOI: 10.1016/j.ejca.2005.04.012
142.Katalinić V, Milos M, Modun D, Musić I, Boban M. Antioxidant effectiveness of selected wines in comparison with (+)-catechin. Food Chemistry. 2004;86(4):593-600. DOI: 10.1016/j.foodchem.2003.10.007
143.Fegredo JA, Meynell R, Lai AKH, Wong MCY, Martin CR, Wiseman H, et al. The antioxidant capacity of beer: Relationships between assays of antioxidant capacity, color and other alcoholic and non-alcoholic beverages. In: Beer in Health and Disease Prevention. Amsterdam, The Netherlands: Elsevier; 2009. pp. 475-481. DOI: 10.1016/B978-0-12-373891-2.00046-8
144.Viana AC, Pimentel TC, do Borges Vale R, Clementino LS, Januario Ferreira ET, Magnani M, et al. American pale ale craft beer: Influence of brewer’s yeast strains on the chemical composition and antioxidant capacity. LWT. 2021;152(August):112317. DOI: 10.1016/j.lwt.2021.112317
145.Pai TV, Sawant SY, Ghatak AA, Chaturvedi PA, Gupte AM, Desai NS. Characterization of Indian beers: Chemical composition and antioxidant potential. Journal of Food Science and Technology. 2015;52(3):1414-1423. DOI: 10.1007/s13197-013-1152-2
146.Tafulo PAR, Queirós RB, Delerue-Matos CM, Sales MGF. Control and comparison of the antioxidant capacity of beers. Food Research International. 2010;43(6):1702-1709. DOI: 10.1016/j.foodres.2010.05.014
147.Munteanu IG, Apetrei C. Analytical methods used in determining antioxidant activity: A review. International Journal of Molecular Sciences. 2021;22(7):3380. DOI: 10.3390/ijms22073380
Written By
Vanesa Postigo, Margarita García and Teresa Arroyo
Submitted: 15 October 2023Reviewed: 20 October 2023Published: 29 November 2023
 Open access peer-reviewed chapter
Open access peer-reviewed chapter