Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
Osteoporosis is a skeleton disease characterized by low bone mass, bone tissue deterioration, and alterations in bone microarchitecture. It is estimated that there are more than 500 million patients worldwide who suffer from osteoporosis-related fractures. There are several risk factors for osteoporosis. These comprise common risk factors like aging and steroid deficiency, as well as specific risk factors such as glucocorticoid use, decreased bone quality, and modifications in bone microarchitecture. However, the pivotal role of the immune system in osteoporosis was initially sidelined in the pathophysiology of osteoporosis but has gained much attention in recent years. Current discoveries have highlighted the existence of a vast network of communication between bone and the immune system in the maintenance of bone homeostasis. Moreover, it is observed that the majority of first-line therapies currently used to treat osteoporosis have immunomodulatory potential thereby preventing bone resorption by regulating the “Immunoporotic axis.” Therefore, these findings would potentially lead to a dramatic shift in the treatment modalities for the management of inflammatory bone loss observed in osteoporosis. To emphasize the crucial role of the immune system in the pathology of osteoporosis, our group coined the term “Immunoporosis” in 2018. In this chapter, we will comprehensively review the pivotal role of the immune system in the pathophysiology of osteoporosis. Furthermore, we will discuss several novel immunomodulatory strategies that can be exploited for the management and prevention of osteoporosis.
Translational Immunology, Osteoimmunology and Immunoporosis Lab, Department of Biotechnology, All India Institute of Medical Sciences (AIIMS), New Delhi, India
Taranjeet Kaur
Translational Immunology, Osteoimmunology and Immunoporosis Lab, Department of Biotechnology, All India Institute of Medical Sciences (AIIMS), New Delhi, India
Asha Bhardwaj
Translational Immunology, Osteoimmunology and Immunoporosis Lab, Department of Biotechnology, All India Institute of Medical Sciences (AIIMS), New Delhi, India
Leena Sapra
Translational Immunology, Osteoimmunology and Immunoporosis Lab, Department of Biotechnology, All India Institute of Medical Sciences (AIIMS), New Delhi, India
Sumedha Yadav
Translational Immunology, Osteoimmunology and Immunoporosis Lab, Department of Biotechnology, All India Institute of Medical Sciences (AIIMS), New Delhi, India
Rupesh K. Srivastava*
Translational Immunology, Osteoimmunology and Immunoporosis Lab, Department of Biotechnology, All India Institute of Medical Sciences (AIIMS), New Delhi, India
*Address all correspondence to: rupesh_srivastava13@yahoo.co.in
1. Introduction
Bone is a connective tissue that consists of two main components: extracellular matrix and cellular component. The extracellular matrix comprises organic components such as collagen type 1 fibrils and non-collagenous glycoproteins and proteoglycans, and inorganic components calcium and phosphate carbonate which are organized as hydroxyapatite crystals. The cellular component includes the bone cells (osteoclasts, osteoblasts, and osteocytes) which drive the process of bone remodeling as each cell has its distinct function.
Osteoblasts are anabolic cells that are derived from pluripotent bone mesenchymal stem cells (BMSCs). They are regarded as “Bone Forming Cells” that are cuboidal in shape when present in active form. Osteoblast differentiation from MSCs is genetically controlled by the expression of many genes, such as runt-related transcription factor-2 (Runx2), distal-less homeobox-5 (Dlx5), and osterix (Osx), etc. [1]. Upon aging, the osteoblasts can either undergo apoptosis or transform into osteocytes and bone-lining cells. The bone lining cells depict the resting phase in the bone remodeling process. The prominent functions of osteoblasts are the synthesis of proteins comprising the extracellular matrix, the regulation of osteoclasts via paracrine signaling, and bone matrix mineralization.
Osteoclasts are bone-eating multinucleated large cells that are formed from colony-forming unit-macrophage (CFU-M) lineage progenitors. Macrophage colony-stimulating factor (MCSF) and receptor activator of nuclear factor kappa-B ligand (RANKL) are crucial regulators of osteoclast differentiation. While RANKL enhances the differentiation of progenitors into osteoclasts, MCSF encourages the proliferation and survival of osteoclast progenitors. The role of osteoblasts in osteoclast differentiation and proliferation is associated with the expression of MCSF and RANKL. The resorption by osteoclasts is mediated by dissolving the hydroxyapatite crystals due to the acidification of resorption lacunae which eventually release calcium, minerals, and growth factors residing in the bone matrix.
The entrapped osteoblasts that are embedded in bone matrix are called osteocytes. They are regarded as the most mature form of osteoblasts and have a peculiar role in regulating bone remodeling. Osteocytes have mechanical stress sensors and respond to them via secretion of insulin-like growth factor 1 (IGF-1). The entrapment of osteocytes is essential for their growth and occurs due to the secretion of chemicals in response to mechanical stress only. Bone formation and resorption are also under the influence of osteocytes in response to external mechanical stimuli. Osteocytes have a distinctive spider-like morphology due to the presence of branched cellular processes that form a network of interconnected canaliculi and permit cell-to-cell communication via gap junctions. Upon maturation, the osteoblastic surface markers are lost, while osteocytes gain specific markers namely dentin matrix acidic phosphoprotein (DMP)-1 and Sclerostin (sost). A study on sost-deficient mice suggests that sclerostin negatively regulates osteoblast differentiation via the canonical Wnt pathway and hence controls bone formation [2]. Additionally, osteocytes maintain mineral homeostasis by expressing RANKL expression resulting in osteoclast activity during high calcium demand.
The dynamic process of bone remodeling requires several interactions between osteoblasts and osteoclasts. Bone remodeling preserves the structure of the bone and constantly swaps out the old bone for a new one. Several skeleton malformations are caused by abnormalities in the bone remodeling process. The intricate process of bone remodeling is controlled by several biochemical and mechanical factors. Important elements that play a role in bone regulation are parathyroid hormone (PTH), estrogen, thyroid hormone and glucocorticoids, IGFs, prostaglandins, and bone morphogenetic proteins (BMPs) and vitamin D. Another crucial factor that has a significant role in the regulation of bone health is the immune system. Recent research has shown that the immune and skeleton system actively interacts with one another. Osteoimmunology is a novel branch of immunology that deals with the intricate link between the bone and the immune system [3]. According to experimental data, pro-inflammatory cytokines such as interleukin (IL)-1, IL-6, and tumor necrosis factor (TNF)-α are detrimental to the bone. These discoveries paved the way for the emergence of a distinct discipline known as “Immunoporosis,” or the immunology of osteoporosis, which was named by our group in 2018. In clinical applications, understanding immunology is more crucial than ever for the treatment and management of osteoporosis.
This chapter discussed in particular the role of the immune system in osteoporosis i.e., Immunoporosis. Additionally, we suggest that in the long run, focusing on the Immunoporotic-nexus would be a much better immunotherapeutic approach to managing and treating the inflammatory bone loss seen in osteoporosis.
Osteoporosis is a disease of the bone in which enhanced bone loss, low bone mineral density (BMD), and increased risk of fractures are observed. This bone loss is caused due to many factors. Some of these include dietary intake, smoking, genetics, etc. Mainly post-menopausal women are at a greater risk of suffering from osteoporosis, this is because the hormone estrogen offers an osteoprotective role as it increases osteoclast apoptosis. Estrogen deficiency in post-menopausal women thus leads to osteoporosis. It can occur in males as well. It has affected nearly 500 million people worldwide [4]. This disease is further classified into primary and secondary osteoporosis. Primary osteoporosis is of two types i.e., type 1 (known as post-menopausal osteoporosis) and type 2 (known as senile osteoporosis). Secondary osteoporosis is caused due to certain defined etiological mechanisms. These mechanisms can be ranging from lifestyle factors like excessive smoking, and alcohol consumption to endocrine disorders like hypogonadism, diabetes, etc. The use of drugs like methotrexate and glucocorticoids is also linked with secondary osteoporosis [5]. Often the disease is undetected and remains silent until it is manifested in the form of a fracture. These fractures occur most commonly at the hip, leading to loss of mobility. The primary diagnostic measure for the screening of osteoporosis is the T score obtained by the dual-energy x-ray absorptiometry (DEXA) scan. A score of 2.5 or any score lower than 2.5 marks osteoporosis [4]. As it is now a well-established fact that there is an appraisable interplay between the bone and the skeletal system further sections of this chapter emphasized the immunology of osteoporosis i.e., Immunoporosis.
3. Immunoporosis: the role of the immune system in osteoporosis
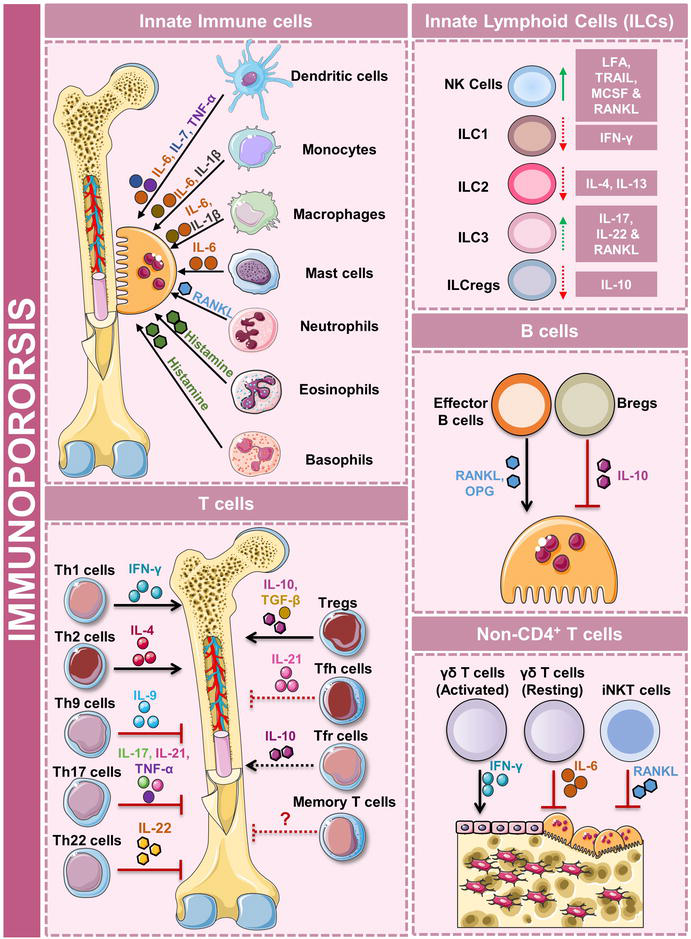
In 2002, Osteoimmunology was established as a novel field in the sciences of immunology to study the interaction between the immune system and the skeletal system. In 2018, our group coined the term “Immunoporosis” and highlighted the importance of the “Immune-skeletal” interface in the development of osteoporosis. Cells of the immune system, mainly lymphocytes, have a profound impact on the skeletal system. These cells can either enhance the activity of osteoclasts and lead to more bone resorption or inhibit the activity of osteoclasts, leading to more bone formation. Immune cells carry out this function by the secretion of certain factors, which are known as cytokines. Cells of the lymphoid lineage, mainly subsets of CD4+T helper cells, B cells, and regulatory B cells (Bregs) have a profound impact on the bone. In addition, cells of the innate immune system also play a crucial role in osteoporosis. In further sections, we will describe the role of innate and adaptive immune cells in the pathophysiology of osteoporosis (Figure 1).
Figure 1.
Role of immune system in osteoporosis i.e., Immunoporosis. Schematic diagram representing the role of different immune cells in the pathophysiology of osteoporosis.
3.1 Role of innate immune cells in osteoporosis
Innate immune cells include dendritic cells (DCs), mast cells, eosinophils, basophils, neutrophils, monocytes, macrophages, and innate lymphoid cells (ILCs), etc. Out of these cells, DCs, monocytes, and macrophages have been reported to enhance osteoclast differentiation. Also, these cells, including osteoclasts are derived from a common progenitor. DCs are precursors of osteoclasts and can develop into fully functional multi-nucleated osteoclasts. It has been reported that when CD11c+ DCs interact with CD4+ T cells, they develop into multinucleated osteoclasts [6]. Classical and intermediate monocytes can also differentiate into functional osteoclasts; however, their differentiation depends on the inflammatory conditions. Under physiological conditions, classical monocytes develop into osteoclasts whereas under inflammatory conditions intermediate monocytes develop into osteoclasts. The role of macrophages in bone remodeling is still under investigation. A study carried out by Yamaguchi et al. demonstrated that upon induction with lipopolysaccharide (LPS) and interferon (IFN)-γ M1 macrophages suppressed RANKL-dependent osteoclastogenesis. However, the same results were not observed with M2 macrophages [7]. Another study also reported that M2 macrophages have better osteoclast-forming capabilities than M1 macrophages [8]. Some studies have also correlated allergy with increased bone loss. Mast cells are known to secrete histamine, it has been reported that histamine aids in osteoclastogenesis by directly acting on osteoclasts [9]. Also, a high number of mast cells have been reported in post-menopausal osteoporotic patients. Eosinophils and basophils are also actively involved in allergic inflammation. The major cytokines observed during allergy are IL-31 and IL-33. IL-31 is increased in post-menopausal osteoporotic patients with low BMD [10]. IL-33 on the other hand has an inhibitory effect on osteoclast differentiation, by the release of IL-10 [11].
Natural killer (NK) cells play a crucial role in tumor immunity and protect virus-infected cells. Interesting findings about these cells have been reported in patients suffering from rheumatoid arthritis (RA). Soderstom et al. reported that NK cells produce MCSF and RANKL and promote osteoclast differentiation [12]. However, in 2015, the same group reported that NK cells can inhibit osteoclastogenesis when they get activated by IL-15. Interestingly, IL-15 has been reported to have a dual role. It can promote the apoptosis of osteoclasts as well as osteoblasts [13, 14].
ILCs are classified into three groups i.e., ILC1, ILC2, and ILC3. These cells are mirror images of the T helper subset cells (Th1, Th2, and Th17) and just like them, are dependent on the master transcription factors T-bet, GATA-3, and RORγt and IFN-γ, IL-4, and IL-17 are moreover their distinguishing cytokines [4]. It has been reported that ILCs produce RANKL and might lead to enhanced osteoclast development. ILC1, via the release of IFN-γ, might regulate the bone remodeling process. ILC2 can also release IL-4 and IL-13, thereby offering an osteoprotective role. ILC3 is similar to Th17 cells. They secrete IL-17, which has been associated with increased osteoclast differentiation and therefore might promote bone loss. However, further studies are required to explore the role of ILCs in the progression of osteoporosis. Collectively, these findings suggest that developing strategies that target the innate immune system might be beneficial in the management and treatment of osteoporosis.
3.2 Role of the adaptive immune system in osteoporosis
3.2.1 T helper cells
A subset of CD4+ lymphoid cells is known as T helper cells. They are divided into the Th1, Th2, Th17, Th9, Th22, T-regulatory cells (Tregs), T-follicular helper cells (Tfh), and memory T cells based on surface markers, their cytokine profile, and transcription factors.
Upon stimulation with IL-12, naïve CD4+ T cells differentiate into Th1 cells. The transcription factor T-bet is involved in the development of this subset. The signature cytokine released by Th1 cells is IFN-γ. It has been observed that IFN-γ has a dual role in osteoclastogenesis. It inhibits osteoclastogenesis, but it also induces RANKL expression in T cells [15]. The dual nature of this cytokine is because its effect depends on the stage of osteoclast maturation. In the early stages, IFN-γ inhibits osteoclastogenesis by suppressing RANK-RANKL-mediated signaling. Interestingly, in later stages, it promotes osteoclastogenesis by encouraging the fusion of monocytes to form multi-nucleated osteoclasts. It can carry out its inhibitory effect via other mechanisms as well. Studies have reported that IFN-γ collaborates with toll-like receptors (TLRs) and inhibit osteoclastogenesis [16]. This cytokine can also upregulate the expression of guanylate-binding proteins (GBP2 and GBP5) which have an inhibitory effect on osteoclast differentiation [17]. Since low levels of IFN-γ have also been reported in the serum of post-menopausal osteoporotic patients, this cytokine could be used as a therapeutic agent that offers an osteoprotective role and potentially alleviates bone deterioration [4].
The transcription factor GATA-3 is responsible for the differentiation of naïve CD4+ T cells into effector CD4+ Th2 cells. This subset secretes IL-4, IL-5, and IL-13. Studies have reported that out of these three signature cytokines, IL-4 and IL-13 are actively involved in the regulation of the Immunoporotic axis. Similar to IFN-γ, low levels of IL-4 have been observed in the serum of post-menopausal osteoporotic patients [18]. The cytokine IL-4 can carry out its effector functions via a diverse array of mechanisms. IL-4 suppresses osteoclastogenesis by inhibiting three main signaling pathways needed for osteoclast function i.e., NF-κB signaling, calcium signaling, and NFATc1 signaling [19]. It has been reported that IL-4 can suppress osteoclastogenesis by inducing the peroxisome proliferator-activated receptor (PPAR-γ) ligand and by blocking the interaction of TNF-α with signal transducers and activators of transcription (STAT)-6. IL-13 and IL-4 can have a combined suppressive effect on osteoclastogenesis. They encourage the production of osteoprotegerin (OPG) by osteoblasts, thereby leading to suppressed osteoclastogenesis [20].
Th9 and Th22 cells are said to be close relatives of Th17 cells and they secrete IL-9 and IL-22 respectively. Th9 cells have both inflammatory and anti-inflammatory potential. The transcription factors involved in Th9 cell differentiation are Foxo-1, STAT6, IRF4, and GATA3. In individuals suffering from RA high levels of IL-9 have been found in the synovial fluid [21]. Studies have shown that Th9 levels are positively correlated with RA. IL-9 also carries out its effector functions by modulating the genes involved in osteoclastogenesis. Th22 cells, like Th9 cells, also promote osteoclast differentiation by the secretion of their signature cytokine IL-22 [22]. However, further research should still be carried out to explore the Immunoporotic potential of these cells in case of osteoporosis.
The transcription factor B cell lymphoma-6/Bcl-6 is involved in the differentiation of Tfh cells. This subset of T cells is involved in B cell development. However, Tfh cells are reported to be highly correlated in RA patients. Ma et al. have observed that these cells promote inflammation in RA by the secretion of cytokine IL-21 [23]. A recently identified T follicular regulatory cell (Tfr), has been reported to have a suppressing effect on the Tfh cells observed in RA patients. A study carried out by Niu et al. in 2018 revealed that the dysregulation of the “Tfh-Tfr” ratio in RA patients is caused due to enhanced IL-6/pSTAT3 signaling [24]. Studies have also shown that estrogen is responsible for inhibiting Tfh activity. Estrogen mediates this activity by suppressing the expression of Bcl-6, which is important in Tfh differentiation [25]. The research on Tfh is still limited and the role of this distinct subset of T cells in osteoporosis can be further explored.
Memory T cells are the surviving population of T cells generated after the pool of antigen-activated effector cells declines in population. The population of memory T cells is maintained by the cytokines IL-7 and IL-15 [26]. Memory T cells are subdivided into further populations. These mainly include central memory T cells, effector memory T cells, and tissue-resident memory T cells. Memory T cells act as a linker between allergy and bone loss. A study reported that Th2 cells increased bone loss in the allergic enteropathy mice model by increasing the population of CD4+CD44hiCD62Llo effector memory T cells. This data might support the hypothesis that there might be an osteoporosis-specific antigen that leads to the development of memory T cells [27].
The balance of Treg and Th17 cells in osteoporosis is the crucial determining factor in the development of this disease. Th17 cells secrete IL-17, which promotes osteoclastogenesis, whereas Tregs secrete IL-10, which is anti-osteoclastogenic in nature. CD4+CD25+FoxP3+ Tregs, via secreting IL-10 and transforming growth factor (TGF)-β inhibit osteoclastogenesis [28]. Apart from carrying out their effector functions via cytokines, they can suppress osteoclast differentiation via expressing Cytotoxic T lymphocyte antigen-4 (CTLA-4), an inhibitory ligand, which interacts with CD80/86 on the precursors of osteoclasts [29]. Th17 cells are osteoclastogenic in nature. They secrete inflammatory cytokines like IL-6, IL-17, TNF-α, and RANKL, which further enhance osteoclastogenesis. IL-6 supports osteoclastogenesis as it is reported to help in the conversion of Tregs to Th17 cells. This cross-talk between Tregs and Th17 cells generates the “Treg-Th17” axis, which heavily influences osteoclastogenesis and ultimately bone resorption [30].
3.3 Non-CD4+ T cells involved in osteoporosis
γδ T cells (Gamma- Delta T cells) and natural killer T cells have been reported to play a role in certain auto-immune diseases where increased bone loss is observed [31]. Unfortunately, their direct role in osteoporosis is still unknown. Most of the T cells express the αβ receptor on their surface. However, a small subpopulation of T cells (1–5% in peripheral circulation) express the unconventional γδ receptor. The γδ T cells are known to secrete IL-6 and IFN-γ, both of these cytokines harness pro and anti-osteoclastogenic roles [32]. A distinct γδ T cell population which also secretes IL-17 (γδ17+ T cells) has been reported to improve effector functions of the cells from lymphoid and myeloid lineage. Their differentiation is supported by IL-23 and literature suggests that these cells might possess some osteoprotective functions. Unfortunately, no studies to date have investigated the role of γδ17+ T cells in post-menopausal osteoporosis [4].
Invariant Natural killer T cells or iNKT cells can promote osteoclastogenesis in a RANKL-dependent manner. These cells express high levels of RANKL in osteoporotic patients [33]. They can indirectly enhance osteoclastogenesis by activating myeloid DCs and macrophages, which are osteoclast precursors.
3.4 Role of B cells in osteoporosis
B cells are classified into B-1 B cells and B-2 B cells. B-1 B cells are derived from hematopoietic stem cells (HSCs) found in the fetal liver, whereas B-2 B cells are derived from HSCs in the bone marrow. They are further classified as marginal zone and follicular B cells. B cells can directly impact the RANKL/OPG ratio, which is a very important contributing factor in osteoporosis. It has been reported that B cells release RANKL, which promotes not only osteoclastogenesis but also increases the population of B cells. This forms a vicious cycle resulting in bone loss [34, 35]. Studies also report that B cells harm osteoblasts as well. B cells release osteoblast-inhibiting factors which include chemokine ligand (CCL)-3 and TNF that activate NF-κB and ERK signaling in osteoblasts, ultimately leading to dysfunctional osteoblasts [36]. These findings suggest that B cells might be an interesting target for the treatment of osteoporosis.
In the early 2000s, the role of regulatory B cells (Bregs) in the modification of immune responses during inflammatory conditions has been well established. Based on different phenotypic markers, Bregs are further divided into distinct subsets. Studies on murine Bregs and human Bregs have established the fact that Bregs offer an osteoprotective role. Their signature cytokines include IL-10, IL-35, and TGF-β. IL-10 and TGF-β can have a direct impact on the Treg/Th17 balance. Our group has also reported that there is a subsequent decrease in the number of IL-10 secreting Bregs (CD19+CD1dhiCD5+) in the osteoporotic mice model [37]. IL-35 secreted by Bregs has also been shown to suppress osteoclastogenesis. IL-35 induces the production of OPG, which is a decoy receptor for RANKL [38]. Therefore, Bregs have a pivotal role in the pathogenesis of osteoporosis.
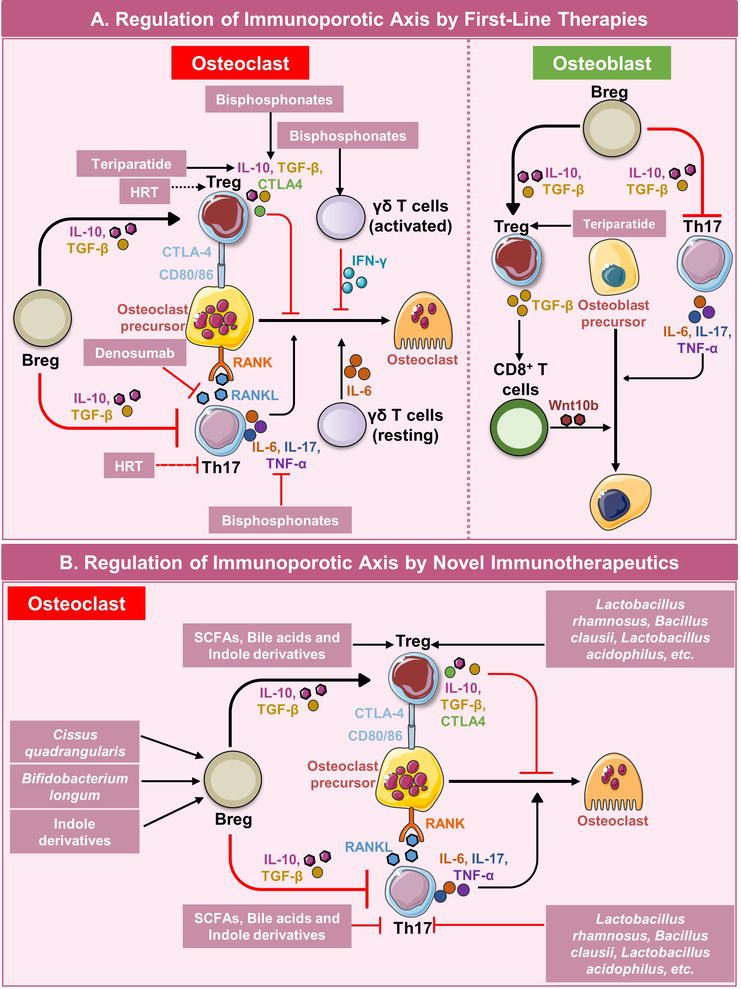
Therapies for osteoporosis include anti-resorptive and anti-osteoclastogenic drugs. First-line anti-resorptive drugs include bisphosphonates, denosumab, and teriparatide. Bisphosphonates are the most common first-line therapy for osteoporosis. They inhibit osteoclast differentiation [39]. Denosumab is an antibody against RANKL. It is a fully humanized, IgG2 resembling, monoclonal antibody. The administration of denosumab has been shown to reduce bone resorption and enhance trabecular bone microstructure [40]. Teriparatide is a synthetic version of the parathyroid hormone. Intermittent dosage of teriparatide has been shown to enhance osteoblast activity [41]. Apart from their direct effect on the bone cells, it is observed that the majority of first-line therapies for osteoporosis have immunomodulatory potential. Therefore, it is plausible to suggest that these drugs are also inhibiting bone resorption by regulating the “Immunoporotic axis.” Below the role of various drugs in regulating the Immunoporotic axis has been explained along with this we have also suggested various novel immunotherapeutics that can be utilized as a treatment modality for osteoporosis by exploiting the immunoporotic axis (Table 1) (Figure 2).
First Line Therapies
S. No.
Therapies
Immunoporotic mechanism of Action
1.
Bisphosphonates
Regulate the cytokine balance between Tregs and Th17 cells (decrease levels of IL-6, IL-17, and IFN-γ while enhancing the levels of IL-4, IL-10, and TGF-β in plasma).
Role of various first-line therapies and novel immunotherapeutics in regulating the bone health by modulating the immunoporotic axis.
Figure 2.
Role of immune cells in regulating bone remodeling and potential therapeutic checkpoints. (A) Immune cells regulate the activity of osteoclasts and osteoblasts by producing various cytokines. First-line therapies can regulate the bone remodeling by further regulating these immune cells. (B) Various novel immunotherapeutics such as phototherapeutics, probiotics, and their metabolites like short-chain fatty acids (SCFAs), bile acids, and tryptophan metabolites have strong immunomodulatory properties and are observed to regulate the bone remodeling by modulating the immune system.
4.1 Effect of already existing first-line therapies on immunoporotic axis
4.1.1 Bisphosphonates
Bisphosphonates are drugs that inhibit the enzyme farnesyl diphosphate synthase i.e., FPP synthase. This enzyme is responsible for the synthesis of isoprenoid lipids, which further are necessary for the post-translational prenylation of GTPases (mostly of Ras, Rho, and Rab families). Since this enzyme is blocked, the prenylation of the GTPases is inhibited and their function is ultimately blocked. Further, in-vivo studies have reported that when bisphosphonates inhibit FPP synthase, the substrates of this enzyme isopentenyl diphosphate (IPP) and dimethylallyl phosphate (DMAPP) accumulate in the cytosol, and by an unknown mechanism, activate a specific subset of γδ T cells (VγVδ2+ T cells). It has been reported that IPP and DMAPP are activators of VγVδ2+ T cells. These cells also secrete IFN-γ, when treated with zoledronic acid. Therefore, these cells can be a potential target for the treatment of osteoporosis [42]. Apart from exerting their effect on VγVδ2+ T cells, bisphosphonates also can modulate the cytokine balance between Th17 cells and Treg cells. According to a study, there was a significant decrease of IL-6, IL-17, and IFN-γ in the plasma levels of osteoporotic patients who received yearly bisphosphonate treatment. There was a significant increase in IL-4, IL-10, and TGF-β plasma levels as well. Since it was earlier mentioned that IFN-γ plays a dual role in the pathophysiology of osteoporosis, this study has not reported the exact correlation of IFN-γ with treated osteoporotic patients. Zoledronic acid inhibits the proliferation of Treg cells in vitro. The immunosuppressive functions of Treg cells are thus heavily impaired due to the action of zoledronic acid [43]. A study reported that bisphosphonates can directly lead to the expansion of antibody-producing B cells. This study concludes that bisphosphonates can enhance humoral immune response [44]. However, the effect of bisphosphonates on Bregs and ultimately on the “Breg-Th17-Treg” axis is still undetermined. Even though studies have shown the effect of bisphosphonates on the immune system, the immune-modulatory properties of these drugs should be further investigated.
4.1.2 Teriparatide
Teriparatide is a synthetic version of the human parathyroid hormone. It is made up of the first 34 amino acids of the parathyroid hormone. It was first approved for medical use in 1987 and later, in November 2002, was approved for the treatment of osteoporosis by the Food and Drug administration (FDA). Unlike bisphosphonates, teriparatide has fewer side effects and can even offer protection against bisphosphonate-related osteonecrosis of the jaw i.e., BRONJ [45]. Teriparatide has an anabolic effect on bone. This synthetic peptide has a direct effect on osteocytes, osteoblasts, and T cells. It is either administered continuously or intermittently. This therapy is therefore known as cPTH or iPTH therapy. iPTH treatment has been reported to reduce the risk of fractures in humans, however, cPTH infusion might harm bone health. It has been reported that cPTH treatment results in cortical bone loss, mainly by activating osteoclast formation [46]. Sclerostin has an inhibitory effect on osteoblast activity and it has been reported that there exists an inverse correlation between sclerostin and PTH levels in healthy women [47]. iPTH treatment leads to the repression of sclerostin and enhances the activity of osteoblasts which results in improved bone health. T cells bearing the receptor for PTH (PPR) act as key mediators in carrying out the effector functions of iPTH. It has been shown that when PTH interacts with the PPR receptor on T cells, Wnt10b signaling is enhanced which lowers the production of sclerostin, whilst increasing osteoblastogenesis [46]. As mentioned earlier, bisphosphonates exert their effect mainly via CD4+ T helper cells. Interestingly, CD8+ T cells are involved in carrying out the effector functions of teriparatide. The engagement of CD8+ T cells became apparent when it was seen that iPTH was unable to show its anabolic effect in the absence of CD8+ T cells. Therefore, literary evidence suggests that Wnt10b, which is an important ligand for osteoblast activity along with CD8+ T cells is important for carrying out the anabolic function of teriparatide [48, 49]. To further support this fact, a study carried out by Terauchi et al. showed that iPTH increased serum levels of osteocalcin (a marker of bone formation) in T cell-reconstituted mice but not in T cell-deficient mice [49]. A similar effect on bone resorption was seen, via observing the serum C-terminal telopeptide of collagen (CTX) levels. The same group also reported that iPTH increased the production of RANKL by both CD4+ T cells and CD8+ T cells [49]. Since RANKL is a known inducer of osteoclastogenesis, the use of iPTH for the treatment of osteoporosis can be questioned. Together these findings establish the cross-talk between the anabolic effects of teriparatide and T cells. Since Tregs are an important component of osteoimmunology, the effect of this synthetic peptide on these subsets can be of immense value. An in vitro study carried out by Yu et al. reported that teriparatide did not increase the number of CD4+CD25+FoxP3+ T cells in cultures of peripheral blood CD4+CD25+ T cells. Yu et al. suggest that teriparatide might affect the Treg population via unknown, indirect mechanisms [50]. Although much data is not available on the effect of teriparatide on B lymphocytes, recent studies show that the PPR receptor is important for B cell maturation and function. It has been demonstrated that the loss of PPR on osteoprogenitors leads to impaired B cell maturation and further causes their egress from bone marrow [51]. This finding very much correlates with the fact that T cells also utilize the PPR receptor for enhancing osteoblast activity. These findings together suggest that the teriparatide actively harnesses the adaptive immune system to carry out its anabolic effects on bone, and this crosstalk between teriparatide and cells of the adaptive immune system might be a good candidate for Immunotherapeutics.
4.1.3 Denosumab
Denosumab is a fully humanized monoclonal antibody resembling IgG2. It has a high affinity for RANKL and its mode of action is similar to the naturally produced RANKL inhibitor OPG. It was manufactured by Amgen, USA, and was approved by FDA for use in 2010. According to Lewiecki yearly use of denosumab marked an increased BMD at the lumbar spine (from 3.0 to 6.7%) and at the hip (from 1.9 to 3.6%). Data from many, phase II randomized studies reveal that denosumab enhances bone health. A significant increase in BMD at the lumbar spine was observed in all the randomized studies [52]. Although no significant effect of denosumab on immune cells was reported. However, Rossini et al. reported that during 1-year administration of denosumab, the number of B cells and CD4+ T cells increased and remained at a higher percentage for 1 year [53]. The RANK receptor, apart from osteoclasts, is also expressed in immune cells which release pro-inflammatory cytokines and are important in mounting an immune response. The inhibition of this system by the use of denosumab has been linked with an increased risk of infections and side effects due to the suppression of the immune system. Even though increased BMD was observed in patients receiving denosumab treatment, the severe risk of infections remains an issue. Therefore, a deeper understanding of the RANK/RANKL/OPG axis can lead to the development of better strategies that harness the Immune-skeletal interface and thus help in the efficient management of osteoporosis.
4.1.4 Anti-sclerostin antibody
Sclerostin is a glycoprotein released by osteocytes that blocks the Wnt signaling needed for osteoblast formation. More sclerostin means less bone formation, which ultimately leads to bone loss. Since sclerostin binds to Wnt molecules, it became an attractive pharmacological target. Eventually, Amgen released romosozumab in 2019 which is a fully humanized monoclonal antibody against sclerostin. It binds to sclerostin and prevents its interaction with Wnt signaling molecules. Wnt then interacts with its transmembrane receptor LRP5/6, which dephosphorylates the protein axin. This leads to the rise of β-catenin, which enhances the transcription of Runx2 and OPG. Ultimately, sclerostin inhibition enhances the Wnt-signaling cascade, which leads to osteoblast differentiation and proliferation [54]. Studies reported that the use of romosozumab led to increased BMD in the lumbar spine, decreased incidence of fractures, and increased cortical bone architecture [55, 56]. When combined with teriparatide and bisphosphonates, a significant increase in BMD at the lumbar spine and hip was also observed. However, no significant difference was observed when romosozumab was combined with denosumab [54]. Another anti-sclerostin antibody, blomosozumab, offered similar results i.e., an increase in BMD and reduced risk of fractures. Unfortunately, the use of romosozumab is associated with adverse side effects as well. It was observed that the use of romosozumab was associated with increased cardiovascular risks [57].
4.1.5 Cathepsin K inhibitors
Cathepsin K is a proteolytic enzyme that belongs to the class of cysteine proteases and is coded by the gene CstK. It is abundant in osteoclasts and is highly efficient in cleaving the complex triple helix of type I collagen. Since this enzyme plays a crucial role in bone remodeling, it has been identified as a potential therapeutic target. This rationale has been proven by animal studies. Pennypacker et al. have demonstrated that cathepsin K knockout mice have enhanced bone health i.e., increased BMD, increased trabecular bone, and overall higher bone strength [58]. On the other hand, overexpression of cathepsin K in mice led to increased bone turnover [59]. Some of the most widely used cathepsin K inhibitors are odanacatib, relacatib, and balicatib. Out of all the three drugs, odanacatib has the highest cathepsin K inhibiting potential with very limiting side effects. It has been reported that odanacatib does not have an inhibitory effect on other closely related cathepsins (cathepsin L, S, and V), which are present in different cells and are important for homeostasis [60].
4.1.6 Hormone replacement therapy (HRT)
Estrogen insufficiency is the most common cause of postmenopausal osteoporosis, resulting in rapid bone loss that peaks within the first 2–3 years after menopause. Hormone replacement therapy (HRT) has been considered one of the first-line therapy. HRT replaces hormones that women lose throughout the menopausal transition. To alleviate the symptoms of menopause, conventional HRT contains estrogen and progesterone to mimic hormones produced by the human ovary. It includes only estrogen therapies and estrogen-progesterone or progestin hormone therapy. HRT inhibits not only bone loss and the degeneration of bone microarchitecture but also reduces the risk of fracture by 20–40% at all bone sites. However, the preventive impact of HRT on BMD reduces at an unexpected rate after medication discontinuation, albeit some degree of fracture protection may exist following HRT discontinuation [61]. HRT’s osteoprotective and fracture-prevention effects were diminished and non-significant in women over the age of 60. Also, the adverse effects of using HRT include an increased risk of breast cancer, endometrial cancer, blood clots, and stroke with long-term use [62]. There are numerous estrogen medications available, including those produced by the human ovary, such as estradiol and estriol. Conjugated equine estrogen (CEE), the most often prescribed estrogen in the United States, is one of the several estrogenic chemicals [63]. Estrogen is easily absorbed by the gastrointestinal (GI) tract, skin, and mucous membranes. Estrogen has strong immunomodulatory properties and has a role in preventing osteoporosis by influencing the Breg-Treg-Th17 cell axis i.e., the immunoporotic axis [37, 64].
4.2 Novel immunotherapeutics for osteoporosis
As already discussed, most of the currently available first-line therapies have the capability of modulating the immune system and therefore might be preventing bone loss by influencing the immune system. However, these therapies are associated with several side effects and thus there is an urgent need for effective therapeutics with fewer side effects. Below we have suggested some compounds that have shown phenomenal immunomodulatory properties and therefore can be utilized for the prevention and management of osteoporosis (Figure 2).
4.2.1 Phytotherapy
Recently, emerging research has shown that many plants and herbs harness immunomodulatory properties. Bioactive compounds from the plants Moringa oleifera, Curcuma longa, and Cissus quadrangularis, modulate the immune system and play a therapeutic role in osteoporosis. Our group has reported that the administration of C. quadrangularis has a positive effect on bone health, by modulating the immune cells involved in osteoporosis [65]. Sitosterol-3-O-D-glucoside, niazirin, and marumoside A are compounds found in the seeds of Moringa oleifera. Ma et al. have reported that all three compounds suppress the secretion of pro-inflammatory cytokines IL-17A and IL-22. The leaves of the same plant also contain a compound quercetin, which inhibits RANKL production and the secretion of IL-17 from Th17 cells. Another such RANKL inhibitor is a compound called 18-glycyrrhetinic acid (isolated from the roots of Glycrrhiza glabra), which blocks RANKL-induced osteoclast differentiation [66]. This compound inhibits the interaction of TRAF6 with RANKL, which further prevents the activation of MAPK and NF-κB pathways. Many other plants like Cucurma longa, Psoralea corylifolia, and Crocus sativus suppress RANKL-induced osteoclastogenesis [67, 68, 69, 70, 71]. Therefore, phytotherapy can be considered an excellent treatment option for osteoporosis however further scientific evidence is required for employing phytotherapy in an osteoporosis` treatment regime.
4.2.2 Probiotics
The probiotics are live non-pathogenic microorganisms that harbor health benefits to individuals when administered in adequate amounts. Numerous bacterial genera, including Lactobacillus, Enterococcus, Bacillus, Escherichia, and Bifidobacterium, have been employed as probiotics due to their advantageous effects. A microbe must exhibit certain traits to be labeled as a probiotic, including the ability to tolerate acid and bile, remain stable in terms of genotype and phenotype, adhere to mucosal surfaces, be resistant to antibiotics, produce antimicrobial substances, and be able to inhibit known pathogens.
Several studies have highlighted the role of probiotics in preventing bone loss by modulating the immune system. In healthy animal model studies, the effect of probiotics was observed to be strain and gender dependent. When Lactobacillus reuteri ATCC 6475 was given orally for about 4 weeks to mice, an increased bone mineral density was observed only in male mice. Furthermore, there was enhanced osteocalcin expression as well as an increased rate of bone formation while reduced expression of inflammatory cytokine TNF-α in the jejunum and ileum. In females, under a mild inflammatory state, oral administration of probiotics resulted in enhanced bone density after 8 weeks rather than 4 weeks (as in males). The conclusion from this experiment was that, in healthy conditions, an anti-inflammatory environment is experienced by females which prevented any effect of L. reuteri administration. But as soon as a pro-inflammatory environment is induced, the benefactor role of L. reuteri can be witnessed [72].
The immune system has a prominent role in regulating bone health and osteoporosis [4, 73, 74]. Bifidobacterium longum (BL) via enhancing anti-osteoclastogenic and immunomodulating potential of Bregs ameliorates the bone health in Ovx (ovariectomized) mice [64]. The differentiation and functional activity of RANKL-induced osteoclastogenesis are suppressed by BL in both mouse bone marrow cells and human PBMCs. In comparison to control Bregs, BL-induced Bregs were found to be much more effective at inhibiting osteoclastogenesis and altering the balance of Treg-Th17 cells [64]. Other studies reveal that probiotics like Lactobacillus rhamnosus, Lactobacillus acidophilus, and Bacillus clausii maintain the Tregs-Th17 balance, hence preventing bone loss [75, 76, 77]. Therefore, it can be concluded that supplements like probiotics and fermented products can enhance bone health. Hence, probiotics can serve as an adjunct therapy for patients suffering from primary or secondary osteoporosis as they are actively involved in the regulation of the “Breg-Treg-Th17” cell axis.
4.2.3 Postbiotics
The interplay between the immune system, bone, and the gut microbiota is known as the “Gut-Immune-Bone” axis. It is very important in osteoporosis. The immune cells participating in the “Immunoporotic” axis are directly affected by the metabolites secreted from the gut microbiota. Bregs are encouraged to differentiate by a number of gut-associated metabolites (GAMs), including butyrate, 5-hydroxy indole-3-acetic acid (HIAA), acetate, propionate, and pentanoate. These compounds boost the AhR transcription factor’s activity, which is essential for Bregs differentiation. So, after their investigation into the effects of probiotics, researchers focused on microbial metabolites such as short-chain fatty acids (SCFAs) like acetate, propionate, butyrate, and tryptophan metabolites, among others. In this chapter, the immunomodulatory capabilities of SCFAs, bile acids, and tryptophan metabolites are covered.
4.2.3.1 Short-chain fatty acids (SCFAs)
SCFAs are widely studied fermented metabolites of gut microbiota that diffuse in the bloodstream and regulate effector functions of distant organs. Butyrate is a histone deacetylase inhibitor and it is known to inhibit osteoclastogenesis. The immune function of SCFAs includes the antimicrobial metabolite production by intestinal macrophages as well as NLR3 inflammasome activation via receptor binding. They also down-regulate the proinflammatory mediators such as Nitric Oxide (NO), IL-6, and IL-12, induced by lipopolysaccharide (LPS) hence, controlling the systemic inflammation. Treg differentiation and proliferation are also controlled by SCFAs with butyrate being the most potent followed by propionate while acetate regulates B cell functions [78]. The negative regulation of osteoclasts by butyrate and propionate occurs via a receptor-independent mechanism along with enhanced glycolysis in osteoclasts which was specified by blocking glycolysis which reverted the anti-osteoclastogenic potential of these SCFAs [79]. A research study by Tyagi et al. reported that butyrate-induced Tregs promote CD8+ T cells to secrete Wnt10b, a bone anabolic Wnt ligand. All these studies suggest that supplementation of SCFAs in patients with osteoporosis can be a novel therapeutic strategy and it can act as an adjunct therapy along with first-line immunotherapies.
4.2.3.2 Bile acids
The primary bile acids, synthesized in the liver and secreted into the duodenum by the host are essential for lipid and glucose metabolism, and confer antibacterial defenses when converted into secondary bile acids by gut microbiota. Cholic acid and chenodeoxycholic acid are two majorly synthesized bile acids in the human liver. The gut microbiota converts cholic acid to deoxycholic acid and lithocholic acid, regarded as secondary bile acids, and has immunomodulatory effects. It has been reported that two derivatives of LCA i.e., 3-oxo-Lithocholic acid and isoallo-Lithocholic acid have been found to affect adaptive immunity. Former has an inhibitory effect on Th17 cells and later influences the enhanced differentiation of T regulatory cells by increasing FoxP3 expression in low TGFβ concentrations via mitochondrial reactive oxygen species (mitoROS) [80, 81]. The enhanced differentiation of Tregs by IsoalloLCA occurs via the nuclear hormone receptor, NR41A. Secondary bile acids induce the immune system to produce anti-inflammatory cytokines in the gastrointestinal tract. Ursodeoxycholic acid, a secondary bile acid has been found to enhance osteoblastogenesis in Saos-2 a human osteosarcoma cell line [82]. A study revealed that targeting bile acid receptors FXR and TGR5 receptors attenuated not only osteoclastogenesis but also enhanced osteoblastogenesis [83]. Further studies need to be done to observe the role of bile acids in osteoporotic patients, either directly or via the immune system. Our group is also exploring the role of secondary bile acids, modified by gut microbiota in the pathophysiology of Osteoporosis.
4.2.3.3 Tryptophan metabolites
Tryptophan, an essential amino acid is catabolized into its derivatives by the gut microbiota that maintains the immune cell homeostasis and function. There are two pathways by which tryptophan is converted to its metabolites, direct pathway which results in indole, tryptamine, indole ethanol (IE), indole lactic acid (ILA), indole propionic acid (IPA), indole acetic acid (IAA), indole aldehyde (IAld), etc. whereas kynurenine and 3-hydroxyanthranilic acid (HAA) are products of Kynurenine pathway. Tryptophan metabolites regulate the differentiation of immune cells through transcription factors such as the aryl hydrocarbon receptor (AhR) that are expressed by immune cells such as macrophages, T-cells, B-cells, ILCs, etc. The tryptophan metabolites control the differentiation and proliferation of Tregs, Bregs, and ILC3 along with anti-inflammatory macrophages. When the AhR ligand binds to its receptor, it translocates from the cytoplasm into the nucleus and forms heterodimers with the nuclear translocator of AhR, and binds to the gene promoter.
Studies reveal that AhR ligands and receptors are potentially involved in the induction of Tregs to produce anti-inflammatory cytokines i.e., IL-10 and IL-22 [84]. The AhR ligands 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) induce Tregs proliferation while 2-(1’H-indolo-3′-carbonyl)-thiazole-4-carboxylic acid methyl ester (ITE) is an AhR agonist and has a role in controlling autoimmunity in Treg dependent manner.
As per Abron et al. [85], the activation of the AhR ligand with ITE effectively suppresses inflammatory bowel disease and ameliorates encephalomyelitis symptoms. Additionally, kynurenine and its metabolites are also involved in promoting FoxP3+ immunosuppressive Tregs instead of proinflammatory Th17 cells [86, 87]. In post-menopausal osteoporosis, the imbalance between Tregs and Th17 nexus results in an inflammatory environment. Tryptophan metabolite 5 hydroxy indole acetic acid (5-HIAA) has been observed to prevent bone loss in RA by inducing the Bregs [88]. Supplementing the patient with postbiotics like tryptophan and its metabolites or kynurenine and its metabolites can potentially suppress the inflammation via AhR receptor induction in immune cells.
4.2.4 Phytoestrogens
Phytoestrogens are estrogen-like substances produced from plants that resemble 17-estradiol structurally. Phytoestrogens are classified as isoflavones, stilbene, coumestan, and lignan. Isoflavones are the most studied type of phytoestrogens. Genistein, daidzein, glycitein, formononetin, and biochanin A found in soybeans are the primary isoflavone phytoestrogens [89]. Phytoestrogens are found effective in preventing bone loss due to osteoporosis. Treatment of Ovx rats with resveratrol slowed down the deterioration of trabecular bone [90]. Higher BMD values at the spine and hip regions are seen in postmenopausal women who regularly consume a lot of dietary isoflavones [91]. The trabecular microarchitecture of the proximal tibia in the orchidectomized rat model of male osteoporosis is improved by genistein therapy [92]. Phytoestrogens show immunomodulatory properties due to their ability to block intracellular signaling pathways linked to immunological responses. For instance, genistein can reduce allergic inflammation [89]. Isoflavone extract can lessen the activation of MAPK, NF-κB, and JAK-STAT caused by IL-22, IL-17A, and TNF in healthy human epidermal keratinocytes [93]. Phytoestrogens are also reported to prevent bone loss by modulating the immune system. Treatment with formononetin (formo) and isoformononetin (isoformo) prevented bone loss in ovariectomized mice. In contrast to the Ovx group treated with vehicle, formo/isoformo-treated Ovx mice had lower levels of the pro-osteoclastogenic subset Th17 and B cells. Treatment with formo and isoformo also increased T-regulatory cell differentiation and lowered the expression of Th17 differentiation factors in Ovx mice [94]. Thus, it can be concluded that phytoestrogens can prevent osteoporosis by regulating the immunoporotic axis, but additional research is needed to substantiate this claim.
4.2.5 Biologic therapies
For several systemic inflammatory autoimmune disorders such as RA, inflammatory bowel disease, and psoriasis, biologic therapies targeting crucial elements of the dysregulated immune response have significantly improved patient outcomes and changed treatment paradigms. There are already medicines in the clinic that target specific immune cells like B and T cells or released mediators like proinflammatory cytokines such as TNF, IL-1, IL-6, IL-12, IL-17, and IL-23 [95]. Biologic agents such as etanercept (soluble TNF receptor IgG Fc fusion protein), infliximab, adalimumab, and golimumab (anti-TNF mAbs), certolizumab pegol (pegylated antibody fragment), anakinra (IL-1 receptor antagonist), tocilizumab (anti–IL-6 receptor mAb), ustekinumab (anti–IL-12/IL-23 mAb) and, secukinumab (anti–IL-17A mAb) are permitted for several autoimmune diseases [95]. In the case of RA reduced bone loss is associated with biologic medication therapy. Studies using anti-TNF drugs have shown preservation or growth in the BMD of the spine and the hip, as well as an improved profile of bone markers [96]. The first biologic drug approved for the treatment of osteoporosis was denosumab which increases BMD and reduces fracture risk [97]. Another biologic agent used for osteoporosis treatment is romosozumab [98]. Anti-Dkk-1 monoclonal antibody (PF04840082) is predicted to enhance bone mass and reduce the risk of osteoporotic fractures by encouraging osteoblasts to produce more bone [99]. Thus, these modern antiresorptives or anabolic biologic therapies have the potential to build bone mass quickly to normal levels and decrease fracture rates. Therefore, treatment for osteoporosis can be revolutionized by these more recent medicines [100]. However, more clinical studies are necessary to further prove the effectiveness of biological therapy in preventing osteoporosis while also taking into account their adverse side effects.
Recent research has demonstrated the critical part the immune system serves in the pathogenesis of osteoporosis. These results open the door for further research in the emerging topic of immunoporosis. Numerous medications used to treat osteoporosis have deadly adverse effects such as nephrotoxicity, ulceration, and BRONJ. A monoclonal antibody targeting RANKL is denosumab. Treatment with denosumab results in a decrease in RANKL-induced osteoclastogenesis. However, many studies have reported that denosumab use causes adverse effects and potentially suppresses the immune system. Many FDA-approved classes of drugs are used for treatment as well, which include selective estrogen receptor modulators (SERMs) and antibodies (the most recent antibody approved was a sclerostin-inhibiting antibody called romosozumab), but side effects with the use of this antibody are also prevalent. Since so many side effects are observed, it is crucial to further investigate the effects of these drugs on the immune system and develop safer therapeutic strategies for targeting the disease. By regulating the immune system, phytotherapy and probiotics, together with their metabolites, have demonstrated extraordinary osteoprotective capabilities and can thus be potential future therapeutic candidates. To confirm their effectiveness in humans, further study is still required because the majority of studies on phytotherapy and probiotics are conducted on mice.
VP, TK, AB, LS, SY, and RKS acknowledge the Department of Biotechnology, AIIMS, New Delhi-India for providing infrastructural facilities. AB thanks ICMR, LS thanks UGC, and SY thanks DBT for the research fellowship. Figures are created with the help of Servier Medical Art, provided by Servier, licensed under a Creative Commons Attribution 3.0 unported license (https://smart.servier.com).
This work was financially supported by projects: DBT(BT/PR41958/MED/97/524/2021), ICMR (61/05/2022-IMM/BMS), and CCRH-AYUSH (614/2022–2023) Govt. of India, and an intramural grant (AC-21) sanctioned to RKS.
RKS contributed to the conceptualization and writing of the manuscript. VP, TK, AB, LS, and SY participated in the writing and editing of the review. RKS suggested and AB created the illustrations.
References
1.Cappariello A, Ponzetti M, Rucci N. The “soft” side of the bone: Unveiling its endocrine functions. Hormone Molecular Biology and Clinical Investigation. 2016;28(1):5-20
2.Winkler DG. Osteocyte control of bone formation via sclerostin, a novel BMP antagonist. The EMBO Journal. 2003;22(23):6267-6276
3.Walsh MC, Kim N, Kadono Y, Rho J, Lee SY, Lorenzo J, et al. Osteoimmunology: Interplay between the immune system and bone metabolism. Annual Review of Immunology [Internet]. 2006;24:33-63. Available from: http://www.ncbi.nlm.nih.gov/pubmed/16551243
4.Srivastava RK, Sapra L. The rising era of “Immunoporosis”: Role of immune system in the pathophysiology of osteoporosis. Journal of Inflammation Research [Internet]. 2022;15:1667-1698. Available from: https://www.dovepress.com/the-rising-era-of-immunoporosis-role-of-immune-system-in-the-pathophys-peer-reviewed-fulltext-article-JIR
5.Sözen T, Özışık L, Başaran NÇ. An overview and management of osteoporosis. European Journal of Rheumatology [Internet]. 2017;4(1):46-56. Available from: http://www.ncbi.nlm.nih.gov/pubmed/28293453
6.Alnaeeli M, Teng YTA. Dendritic cells differentiate into osteoclasts in bone marrow microenvironment in vivo. Blood. 2009;113(1):264-265
7.Yamaguchi T, Movila A, Kataoka S, Wisitrasameewong W, Ruiz Torruella M, Murakoshi M, et al. Proinflammatory M1 macrophages inhibit RANKL-induced Osteoclastogenesis. Infection and Immunity. 2016;84(10):2802-2812
8.Yang J, Park O, Kim J, Kwon Y, Yun C, Han SH. Modulation of macrophage subtypes by IRF5 determines osteoclastogenic potential. Journal of Cellular Physiology. 2019;234(12):23033-23042
9.Biosse-Duplan M, Baroukh B, Dy M, de Vernejoul MC, Saffar JL. Histamine promotes Osteoclastogenesis through the differential expression of histamine receptors on osteoclasts and osteoblasts. The American Journal of Pathology. 2009;174(4):1426-1434
10.Ginaldi L, De Martinis M, Ciccarelli F, Saitta S, Imbesi S, Mannucci C, et al. Increased levels of interleukin 31 (IL-31) in osteoporosis. BMC Immunology. 2015;16(1):60
11.De Martinis M, Sirufo MM, Suppa M, Ginaldi L. IL-33/IL-31 Axis in osteoporosis. International Journal of Molecular Sciences. 2020;21(4):1239
12.Söderström K, Stein E, Colmenero P, Purath U, Müller-Ladner U, Teixeira de Matos C, et al. Natural killer cells trigger osteoclastogenesis and bone destruction in arthritis. National Academy of Sciences of the United States of America. 2010;107(29):13028-13033
13.Feng S, Madsen SH, Viller NN, Neutzsky-Wulff AV, Geisler C, Karlsson L, et al. Interleukin-15-activated natural killer cells kill autologous osteoclasts via LFA-1, DNAM-1 and TRAIL, and inhibit osteoclast-mediated bone erosion in vitro. Immunology. 2015;145(3):367-379
14.Takeda H, Kikuchi T, Soboku K, Okabe I, Mizutani H, Mitani A, et al. Effect of IL-15 and natural killer cells on osteoclasts and osteoblasts in a mouse coculture. Inflammation. 2014;37(3):657-669
15.Gao Y, Grassi F, Ryan MR, Terauchi M, Page K, Yang X, et al. IFN-γ stimulates osteoclast formation and bone loss in vivo via antigen-driven T cell activation. Journal of Clinical Investigation. 2007;117(1):122-132
16.Tang M, Tian L, Luo G, Yu X. Interferon-gamma-mediated Osteoimmunology. Frontiers in Immunology. 2018;9:1508
17.Place DE, Malireddi RKS, Kim J, Vogel P, Yamamoto M, Kanneganti TD. Osteoclast fusion and bone loss are restricted by interferon inducible guanylate binding proteins. Nature Communications. 2021;12(1):496
18.Zhang J, Fu Q, Ren Z, Wang Y, Wang C, Shen T, et al. Changes of serum cytokines-related Th1/Th2/Th17 concentration in patients with postmenopausal osteoporosis. Gynecological Endocrinology. 2015;31(3):183-190
19.Mangashetti LS, Khapli SM, Wani MR. IL-4 inhibits bone-resorbing activity of mature osteoclasts by affecting NF-κB and Ca2+ Signaling. The Journal of Immunology. 2005;175(2):917-925
20.Yamada A, Takami M, Kawawa T, Yasuhara R, Zhao B, Mochizuki A, et al. Interleukin-4 inhibition of osteoclast differentiation is stronger than that of interleukin-13 and they are equivalent for induction of osteoprotegerin production from osteoblasts. Immunology. 2007;120(4):573-579
21.Kar S, Gupta R, Malhotra R, Sharma V, Farooque K, Kumar V, et al. Interleukin-9 facilitates Osteoclastogenesis in rheumatoid arthritis. International Journal of Molecular Sciences. 2021;22(19):10397
22.Miyazaki Y, Nakayamada S, Kubo S, Nakano K, Iwata S, Miyagawa I, et al. Th22 cells promote osteoclast differentiation via production of IL-22 in rheumatoid arthritis. Frontiers in Immunology. 2018;9:2901
23.Ma J, Zhu C, Ma B, Tian J, Baidoo SE, Mao C, et al. Increased frequency of circulating follicular helper T cells in patients with rheumatoid arthritis. Clinical & Developmental Immunology. 2012;2012:1-7
24.Niu Q, Chun HZ, Juan WX, Xiong JY, Fei AY, Mei LY, et al. Enhanced IL-6/phosphorylated STAT3 signaling is related to the imbalance of circulating T follicular helper/T follicular regulatory cells in patients with rheumatoid arthritis. Arthritis Research & Therapy. 2018;20(1):200
25.Kim DH, Park HJ, Park HS, Lee JU, Ko C, Gye MC, et al. Estrogen receptor α in T cells suppresses follicular helper T cell responses and prevents autoimmunity. Experimental & Molecular Medicine. 2019;51(4):1-9
26.Rubinstein MP, Lind NA, Purton JF, Filippou P, Best JA, McGhee PA, et al. IL-7 and IL-15 differentially regulate CD8+ T-cell subsets during contraction of the immune response. Blood. 2008;112(9):3704-3712
27.Ono-Ohmachi A, Yamada S, Uno S, Tamai M, Soga K, Nakamura S, et al. Effector memory CD4+T cells in mesenteric lymph nodes mediate bone loss in food-allergic enteropathy model mice, creating IL-4 dominance. Mucosal Immunology. 2021;14(6):1335-1346
28.Luo C, Wang L, Sun C, Li D. Estrogen enhances the functions of CD4+CD25+Foxp3+ regulatory T cells that suppress osteoclast differentiation and bone resorption in vitro. Cellular & Molecular Immunology. 2011;8(1):50-58
29.Fischer L, Herkner C, Kitte R, Dohnke S, Riewaldt J, Kretschmer K, et al. Foxp3+ regulatory T cells in bone and hematopoietic homeostasis. Frontiers in Endocrinology (Lausanne). 2019;10:578
30.Kimura A, Kishimoto T. IL-6: Regulator of Treg/Th17 balance. European Journal of Immunology. 2010;40(7):1830-1835
31.Bank I. The role of Gamma Delta T cells in autoimmune rheumatic diseases. Cell. 2020;9(2):462
32.Phalke SP, Chiplunkar SV. Activation status of γδ T cells dictates their effect on osteoclast generation and bone resorption. Bone Reports. 2015;3:95-103
33.Tilkeridis K, Kiziridis G, Ververidis A, Papoutselis M, Kotsianidis I, Kitsikidou G, et al. Immunoporosis: A new role for invariant natural killer T (NKT) cells through overexpression of nuclear factor-κB ligand (RANKL). Medical Science Monitor. 2019;25:2151-2158
34.Onal M, Xiong J, Chen X, Thostenson JD, Almeida M, Manolagas SC, et al. Receptor activator of nuclear factor κB ligand (RANKL) protein expression by B lymphocytes contributes to Ovariectomy-induced bone loss. Journal of Biological Chemistry. 2012;287(35):29851-29860
35.Fujiwara Y, Piemontese M, Liu Y, Thostenson JD, Xiong J, O’Brien CA. RANKL (receptor activator of NFκB ligand) produced by osteocytes is required for the increase in B cells and bone loss caused by estrogen deficiency in mice. Journal of Biological Chemistry. 2016;291(48):24838-24850
36.Sun W, Meednu N, Rosenberg A, Rangel-Moreno J, Wang V, Glanzman J, et al. B cells inhibit bone formation in rheumatoid arthritis by suppressing osteoblast differentiation. Nature Communications. 2018;9(1):5127
37.Sapra L, Bhardwaj A, Kumar Mishra P, Garg B, Verma B, Mishra GC, et al. Regulatory B cells (Bregs) inhibit Osteoclastogenesis and play a potential role in ameliorating Ovariectomy-induced bone loss. Frontiers in Immunology. 2021;12:1. Available from: www.frontiersin.org
38.Yago T, Nanke Y, Kawamoto M, Kobashigawa T, Yamanaka H, Kotake S. IL-35 inhibits human osteoclastogenesis from monocytes induced by receptor-activator of NF-κB ligand. Central European Journal of Immunology. 2018;43(2):148-154
39.Xu Q, Zhan P, Li X, Mo F, Xu H, Liu Y, et al. Bisphosphonate-enoxacin inhibit osteoclast formation and function by abrogating RANKL-induced JNK signalling pathways during osteoporosis treatment. Journal of Cellular and Molecular Medicine. 2021;25(21):10126-10139
40.Kostenuik PJ, Nguyen HQ, McCabe J, Warmington KS, Kurahara C, Sun N, et al. Denosumab, a fully human monoclonal antibody to RANKL, inhibits bone resorption and increases BMD in knock-in mice that express chimeric (murine/human) RANKL. Journal of Bone and Mineral Research. 2009;24(2):182-195
41.Kuo SW, Rimando M, Liu YS, Lee O. Intermittent Administration of Parathyroid Hormone 1-34 enhances Osteogenesis of human mesenchymal stem cells by regulating protein kinase Cδ. International Journal of Molecular Sciences. 2017;18(10):2221
42.Nada MH, Wang H, Workalemahu G, Tanaka Y, Morita CT. Enhancing adoptive cancer immunotherapy with Vγ2Vδ2 T cells through pulse zoledronate stimulation. Journal for Immunotherapy of Cancer. 2017;5(1):9
43.Liu H, Wang SH, Chen SC, Chen CY, Lo JL, Lin TM. Immune modulation of CD4+CD25+ regulatory T cells by zoledronic acid. BMC Immunology. 2016;17(1):45
44.Tonti E, Jiménez de Oya N, Galliverti G, Moseman EA, Di Lucia P, Amabile A, et al. Bisphosphonates target B cells to enhance humoral immune responses. Cell Reports. 2013;5(2):323-330
45.Yamachika E, Matsui Y, Matsubara M, Matsumura T, Nakata N, Moritani N, et al. The influence of zoledronate and teriparatide on gamma delta T cells in mice. Journal of Dental Sciences. 2017;12(4):333-339
46.Pacifici R. T cells, osteoblasts, and osteocytes: Interacting lineages key for the bone anabolic and catabolic activities of parathyroid hormone. Annals of the New York Academy of Sciences. 2016;1364(1):11-24
47.Mirza FS, Padhi ID, Raisz LG, Lorenzo JA. Serum sclerostin levels negatively correlate with parathyroid hormone levels and free estrogen index in postmenopausal women. The Journal of Clinical Endocrinology and Metabolism. 2010;95(4):1991-1997
48.Bedi B, Li JY, Tawfeek H, Baek KH, Adams J, Vangara SS, et al. Silencing of parathyroid hormone (PTH) receptor 1 in T cells blunts the bone anabolic activity of PTH. Proceedings of the National Academy of Sciences. 2012;109(12):E725-E733
49.Terauchi M, Li JY, Bedi B, Baek KH, Tawfeek H, Galley S, et al. T lymphocytes amplify the anabolic activity of parathyroid hormone through Wnt10b signaling. Cell Metabolism. 2009;10(3):229-240
50.Yu M, D’Amelio P, Tyagi AM, Vaccaro C, Li J, Hsu E, et al. Regulatory T cells are expanded by teriparatide treatment in humans and mediate intermittent PTH-induced bone anabolism in mice. EMBO Reports. 2018;19(1):156-171
51.Panaroni C, Fulzele K, Saini V, Chubb R, Pajevic PD, Wu JY. PTH signaling in osteoprogenitors is essential for B-lymphocyte differentiation and mobilization. Journal of Bone and Mineral Research. 2015;30(12):2273-2286
52.Wensel TM, Iranikhah MM, Wilborn TW. Effects of denosumab on bone mineral density and bone turnover in postmenopausal women. Pharmacotherapy. 2011;31(5):510-523
53.Rossini M, Viapiana O, Adami S, Idolazzi L, Ghellere F, Tripi G, et al. Effects of denosumab on peripheral lymphocyte subpopulations. Endocrine. 2016;53(3):857-859
54.Fabre S, Funck-Brentano T, Cohen-Solal M. Anti-Sclerostin antibodies in osteoporosis and other bone diseases. Journal of Clinical Medicine. 2020;9(11):3439
55.Cosman F, Crittenden DB, Adachi JD, Binkley N, Czerwinski E, Ferrari S, et al. Romosozumab treatment in postmenopausal women with osteoporosis. New England Journal of Medicine. 2016;375(16):1532-1543
56.Geusens P, Oates M, Miyauchi A, Adachi JD, Lazaretti-Castro M, Ebeling PR, et al. The effect of 1 year of romosozumab on the incidence of clinical vertebral fractures in postmenopausal women with osteoporosis: Results from the FRAME study. JBMR Plus. 2019;3(10):e10211
57.Lim SY, Bolster MB. Clinical utility of romosozumab in the management of Osteoporosis: Focus on patient selection and perspectives. International Journal of Women's Health. 2022;14:1733-1747
58.Pennypacker B, Shea M, Liu Q, Masarachia P, Saftig P, Rodan S, et al. Bone density, strength, and formation in adult cathepsin K (−/−) mice. Bone. 2009;44(2):199-207
59.Kiviranta R, Morko J, Uusitalo H, Aro HT, Vuorio E, Rantakokko J. Accelerated turnover of metaphyseal trabecular bone in mice overexpressing cathepsin K. Journal of Bone and Mineral Research. 2001;16(8):1444-1452
60.Duong LT, Leung AT, Langdahl B. Cathepsin K inhibition: A new mechanism for the treatment of osteoporosis. Calcified Tissue International. 2016;98(4):381-397
61.Yates J, Barrett-Connor E, Barlas S, Chen YT, Miller PD, Siris ES. Rapid loss of hip fracture protection after estrogen cessation: Evidence from the National osteoporosis risk assessment. Obstetrics & Gynecology. 2004;103(3):440-446
62.Mehta J, Kling JM, Manson JE. Risks, benefits, and treatment modalities of menopausal hormone therapy: Current concepts. Frontiers in Endocrinology (Lausanne). 2021;12:564781
63.Gambacciani M, Levancini M. Featured editorial hormone replacement therapy and the prevention of postmenopausal osteoporosis. Menopausal Review. 2014;4:213-220
64.Sapra L, Shokeen N, Porwal K, Saini C, Bhardwaj A, Mathew M, et al. Bifidobacterium longum ameliorates ovariectomy-induced bone loss via enhancing anti-osteoclastogenic and immunomodulatory potential of regulatory B cells (Bregs). Frontiers in Immunology. 2022;13:875788
65.Azam Z, Sapra L, Baghel K, Sinha N, Gupta RK, Soni V, et al. Cissus quadrangularis (Hadjod) inhibits RANKL-induced osteoclastogenesis and augments bone health in an estrogen-deficient preclinical model of osteoporosis via modulating the host osteoimmune system. Cell. 2023;12(2):216
66.Chen X, Zhi X, Yin Z, Li X, Qin L, Qiu Z, et al. 18β-glycyrrhetinic acid inhibits osteoclastogenesis in vivo and in vitro by blocking RANKL-mediated RANK–TRAF6 interactions and NF-κB and MAPK signaling pathways. Frontiers in Pharmacology. 2018;9:647
67.Yang C, Zhu K, Yuan X, Zhang X, Qian Y, Cheng T. Curcumin has immunomodulatory effects on RANKL-stimulated osteoclastogenesis in vitro and titanium nanoparticle-induced bone loss in vivo. Journal of Cellular and Molecular Medicine. 2020;24(2):1553-1567
68.Chen H, Fang C, Zhi X, Song S, Gu Y, Chen X, et al. Neobavaisoflavone inhibits osteoclastogenesis through blocking RANKL signalling-mediated TRAF6 and c-Src recruitment and NF-κB, MAPK and Akt pathways. Journal of Cellular and Molecular Medicine. 2020;24(16):9067-9084
69.Fu L, Pan F, Jiao Y. Crocin inhibits RANKL-induced osteoclast formation and bone resorption by suppressing NF-κB signaling pathway activation. Immunobiology. 2017;222(4):597-603
70.Shi L, Zhao S, Chen Q, Wu Y, Zhang J, Li N. Crocin inhibits RANKL-induced osteoclastogenesis by regulating JNK and NF-κB signaling pathways. Molecular Medicine Reports. 2018;17(6):7947-7951
71.Azam Z, Sapra L, Bhardwaj A, Yadav S, Mishra PK, Shukla P, et al. Crocin attenuates osteoclastogenesis and enhances bone health by skewing the immunoporotic “Treg-Th17” cell axis in post-menopausal osteoporotic mice model. Phytomedicine Plus. 2022;2(3):100302
72.McCabe LR, Irwin R, Schaefer L, Britton RA. Probiotic use decreases intestinal inflammation and increases bone density in healthy male but not female mice. Journal of Cellular Physiology. 2013;228(8):1793-1798
73.Srivastava RK, Dar HY, Mishra PK. Immunoporosis: Immunology of osteoporosis—Role of T cells. Frontiers in Immunology. 2018;9:657
74.Sapra L, Azam Z, Rani L, Saini C, Bhardwaj A, Shokeen N, et al. “Immunoporosis”: Immunology of osteoporosis. Proceedings of the National Academy of Sciences, India Section B: Biological Sciences [Internet]. 2021;91(3):511-519. Available from: https://link.springer.com/10.1007/s40011-021-01238-x
75.Sapra L, Dar HY, Bhardwaj A, Pandey A, Kumari S, Azam Z, et al. Lactobacillus rhamnosus attenuates bone loss and maintains bone health by skewing Treg-Th17 cell balance in Ovx mice. Scientific Reports [Internet]. 2021;11(1):1807. DOI: 10.1038/s41598-020-80536-2
76.Dar HY, Pal S, Shukla P, Mishra PK, Tomar GB, Chattopadhyay N, et al. Bacillus clausii inhibits bone loss by skewing Treg-Th17 cell equilibrium in postmenopausal osteoporotic mice model. Nutrition [Internet]. 2018;54:118-128. Available from: http://www.ncbi.nlm.nih.gov/pubmed/29793054
77.Dar HY, Shukla P, Mishra PK, Anupam R, Mondal RK, Tomar GB, et al. Lactobacillus acidophilus inhibits bone loss and increases bone heterogeneity in osteoporotic mice via modulating Treg-Th17 cell balance. Bone Reports [Internet]. 2018;8:46-56. Available from: http://www.ncbi.nlm.nih.gov/pubmed/29955622
78.Mariño E, Richards JL, McLeod KH, Stanley D, Yap YA, Knight J, et al. Gut microbial metabolites limit the frequency of autoimmune T cells and protect against type 1 diabetes. Nature Immunology. 2017;18(5):552-562
79.Lucas S, Omata Y, Hofmann J, Böttcher M, Iljazovic A, Sarter K, et al. Short-chain fatty acids regulate systemic bone mass and protect from pathological bone loss. Nature Communications [Internet]. 2018;9(1):55. Available from: http://www.nature.com/articles/s41467-017-02490-4
80.Hang S, Paik D, Yao L, Kim E, Trinath J, Lu J, et al. Bile acid metabolites control TH17 and Treg cell differentiation. Nature. 2019;576(7785):143-148
81.Li W, Hang S, Fang Y, Bae S, Zhang Y, Zhang M, et al. A bacterial bile acid metabolite modulates Treg activity through the nuclear hormone receptor NR4A1. Cell Host & Microbe. 2021;29(9):1366-1377.e9
82.Dubreuil M, Ruiz-Gaspà S, Guañabens N, Peris P, Álvarez L, Monegal A, et al. Ursodeoxycholic acid increases differentiation and mineralization and neutralizes the damaging effects of bilirubin on osteoblastic cells. Liver International. 2013;33(7):1029-1038
83.Li Z, Huang J, Wang F, Li W, Wu X, Zhao C, et al. Dual targeting of bile acid receptor-1 (TGR5) and Farnesoid X receptor (FXR) prevents estrogen-dependent bone loss in mice. Journal of Bone and Mineral Research. 2018;34(4):765-776; jbmr.3652
84.Gandhi R, Kumar D, Burns EJ, Nadeau M, Dake B, Laroni A, et al. Activation of the aryl hydrocarbon receptor induces human type 1 regulatory T cell–like and Foxp3+ regulatory T cells. Nature Immunology. 2010;11(9):846-853
85.Abron JD, Singh NP, Mishra MK, Price RL, Nagarkatti M, Nagarkatti PS, et al. An endogenous aryl hydrocarbon receptor ligand, ITE, induces regulatory T cells and ameliorates experimental colitis. American Journal of Physiology-Gastrointestinal and Liver Physiology. 2018;315(2):G220-G230
86.Bessede A, Gargaro M, Pallotta MT, Matino D, Servillo G, Brunacci C, et al. Aryl hydrocarbon receptor control of a disease tolerance defence pathway. Nature. 2014;511(7508):184-190
87.Masuda K, Kimura A, Hanieh H, Nguyen NT, Nakahama T, Chinen I, et al. Aryl hydrocarbon receptor negatively regulates LPS-induced IL-6 production through suppression of histamine production in macrophages. International Immunology. 2011;23(10):637-645
88.Rosser EC, Piper CJM, Matei DE, Blair PA, Rendeiro AF, Orford M, et al. Microbiota-derived metabolites suppress arthritis by amplifying aryl-hydrocarbon receptor activation in regulatory B cells. Cell Metabolism [Internet]. 2020;31(4):837-851.e10. Available from: https://linkinghub.elsevier.com/retrieve/pii/S1550413120301182
89.Desmawati D, Sulastri D. Phytoestrogens and their health effect. Open Access Macedonian Journal of Medical Sciences. 2019;7(3):495-499
90.Zaychenko G, Belenichev I, Hnatiuk V, Doroshenko A, Sinitsyna O, Sulaieva O, et al. Protective effect of vaginal resveratrol administration on joint tissues in ovariectomized rats: Targeting mTOR and сaspase 3. Biomedicine & Pharmacotherapy. 2023;165:115176
91.Mei J, Yeung SSC, Kung AWC. High dietary phytoestrogen intake is associated with higher bone mineral density in postmenopausal but not premenopausal women. The Journal of Clinical Endocrinology and Metabolism. 2001;86(11):5217-5221
92.Filipović B, Šošić-Jurjević B, Ajdžanović V, Živanović J, Manojlović-Stojanoski M, Nestorović N, et al. The phytoestrogen genistein prevents trabecular bone loss and affects thyroid follicular cells in a male rat model of osteoporosis. Journal of Anatomy. 2018;233(2):204-212
93.Li HJ, Wu NL, Lee GA, Hung CF. The therapeutic potential and molecular mechanism of Isoflavone extract against psoriasis. Scientific Reports. 2018;8(1):6335
94.Mansoori MN, Tyagi AM, Shukla P, Srivastava K, Dev K, Chillara R, et al. Methoxyisoflavones formononetin and isoformononetin inhibit the differentiation of Th17 cells and B-cell lymphopoesis to promote osteogenesis in estrogen-deficient bone loss conditions. Menopause. 2016;23(5):565-576
95.Her M, Kavanaugh A. Alterations in immune function with biologic therapies for autoimmune disease. Journal of Allergy and Clinical Immunology. 2016;137(1):19-27
96.Zerbini CAF, Clark P, Mendez-Sanchez L, Pereira RMR, Messina OD, Uña CR, et al. Biologic therapies and bone loss in rheumatoid arthritis. Osteoporosis International. 2017;28(2):429-446
97.Tu KN, Lie JD, Wan CKV, Cameron M, Austel AG, Nguyen JK, et al. Osteoporosis: A review of treatment options. P T. 2018;43(2):92-104
98.Rosen CJ, Ingelfinger JR. Building better bones with biologics — A new approach to osteoporosis? New England Journal of Medicine. 2016;375(16):1583-1584
99.Betts AM, Clark TH, Yang J, Treadway JL, Li M, Giovanelli MA, et al. The application of target information and preclinical pharmacokinetic/pharmacodynamic modeling in predicting clinical doses of a Dickkopf-1 antibody for osteoporosis. The Journal of Pharmacology and Experimental Therapeutics. 2010;333(1):2-13
100.Tella SH, Gallagher JC. Biological agents in management of osteoporosis. European Journal of Clinical Pharmacology. 2014;70(11):1291-1301
Written By
Vardhan Pol, Taranjeet Kaur, Asha Bhardwaj, Leena Sapra, Sumedha Yadav and Rupesh K. Srivastava
Submitted: 28 August 2023Reviewed: 23 October 2023Published: 13 December 2023
 Open access peer-reviewed chapter
Open access peer-reviewed chapter