 Open access peer-reviewed chapter
Open access peer-reviewed chapter


Abstract
The goat is an important part of livestock farming due to their meat, milk, wool, and other products. The understanding of the goat genome has opened drastic opportunities for productivity improvement. Many important genomic technologies have been developed, including microsatellites, single nucleotide polymorphism, and whole genome sequencing, and these techniques are being used to identify important genomic regions in the goat genome. Identification of important genes related to meat, milk, and wool can help design breeding programs for increasing the productivity of goat farming. Recent advances in genome engineering tools like zinc finger nuclease, TALENS, and CRISPR/Cas9 have also made it easier to engineer farm animal genomes. Medically and commercially important genes are being engineered in farm animals for medicinal and commercial purposes. This chapter will focus on some of these technologies being applied in goat breeding to increase animal health and the commercial economy.
Keywords
- Capra hircus
- genetics
- microsatellites
- SNP
- GWAS
- genome engineering
- productivity
1. Introduction
Goats (
The past 100 years have seen technological modification and new scientific methods, which have caused an immense increase in the outcome of livestock globally. The selective breeding programs in livestock animals to produce animals better suited to the environment, management systems, and better productivity drove the manipulation of animal genetic resources. These modifications directed at increasing the genetic potential of livestock resulted in new breeds for major livestock animals, which contributed to increasing the income of farmers. Initially, inventions in reproduction techniques made it possible to deliver high-merit genetics for breeding programs resulting in increased selection pressure. In addition, improvements in computing methods, selection accuracy, and breeding value estimation were observed affecting the animal selection programs, the combination of these approaches with quantitative and qualitative genetics resulted in the development of genomic tools, which resulted in increasing the production potential of farm animals by many folds over time. Therefore, these developments caused a drastic change in approaches directed toward improving livestock productivity. For instance, during the period of 1957–2005, an increase of 400% in broiler growth rate and an increase of 50% in food conversion rate has been recorded [10]. These approaches and tools have not only resulted in accelerating the progress rate in the livestock industry but also have been a major reason for understanding the lifespan of animals and complex biological pathways controlling the productivity-related traits in farm animals. In addition, the breeding programs focused on increasing the production and yield of livestock not only achieved productivity but also the lifespan of these animals under selection saw improvement [11]. Over time these breeding programs have expanded their goals targeting multiple characteristics in animal health, welfare, survival, fertility, and other welfare-affecting characteristics in animals (Figure 1) [12, 13, 14].
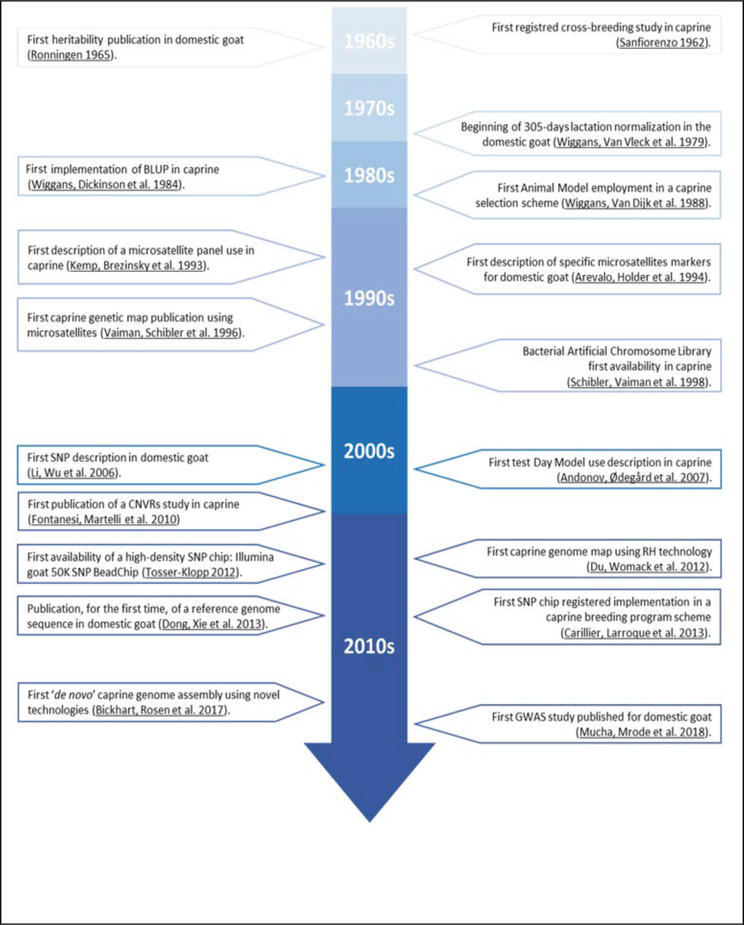
Figure 1.
Chronology and timeline of caprine genetic advances milestones [
All of these notable and important characteristics affecting the commercial value of livestock are defined by the genomic makeup of animals. The vast majority of the genes in sheep and goat genomes affecting productivity traits have been identified. These genes are controlling important characteristics like disease resistance, sensitivity, production performance, and reproductive performance. Their identification has resulted in controlling the targets for improving economic traits through genetic variation [32, 33]. The discovery coupled with molecular genetics techniques has provided the possibility of increasing the selection accuracy in the early stages of the breeding programs [34]. In addition, molecular genetics has provided information on individual candidate genes related to individual economic traits. These individual candidate genes targeting approaches help to identify important qualitative traits loci (QTLs) in the genes [35, 36].
A number of studies have reported the candidate genes influencing milk, wool, reproductive, disease resistance, and growth traits in goats [37]. However, there are also genes that control more than one characteristic in goats, for example, the GH gene (growth hormone) is influencing both milk and growth traits. The candidate genes are involved in sex determination, disease resistance, reproduction, metabolism, and productivity in goats proving to be economically important [38, 39, 40, 41, 42, 43, 44]. The techniques involved in the candidate gene studies have the ability to identify the region of genes where genetic variation at QTL is present and how it is affecting the trait [45]. Molecular genetics provides us with the ability to identify this genetic variation at specific loci and manipulates them to increase goat productivity.
In this chapter, our aim is to discuss the technologies involved in underlining the importance of genetics in increasing the productivity of goat productivity. In addition, we will also focus on the areas of genetics that provide a great service to better the production and productivity of goats.
2. Use of genomic tools
The genomics approaches started in the 1980s, and the major focus for developing this technology was to develop standalone genomic markers that can be used against inherited diseases and for parentage testing [46, 47]. Thereafter, the focus shifted from parentage testing and inherited disease toward the more economical traits affecting QTLs to be used in marker-assisted breeding, through combining quantitative genomic technologies and marker-assisted selection (MAS). Furthermore, the fact was then realized that these commercially important traits are not controlled by the expression of a small number of genes instead, it is controlled by hundreds of genes involved in the expression of these economic traits, this made way for a further intensive approach for development. The recent 15 years saw the refining and implementation of different methods to be used in genomic selection. The advances in the genomic selection methods combined with data analytic techniques and computing methods have helped to generate a large amount of information for predicting breeding value efficiently.
2.1 Microsatellites methodology and application
Short tandem repeats also known as microsatellites are simple sequence repeats, which are present in the genome of all mammals. These are identified by designing specific primers according to specific sequence repeats for DNA flanking microsatellite regions and are amplified through a polymerase chain reaction. The microsatellites unit number may vary depending on the microsatellites and the number of repeats can change from 2 to 30.
The methodology of microsatellites is related to designing specific PCR primers that are related to specific specie and their specific place in the genome, two primers are designed for microsatellites on either side of repeats. After applying the primers, the segments are amplified in the PCR, the PCR amplified segments are then analyzed either on capillary electrophoresis or gel electrophoresis. However, the investigator can determine the times’ the CA dinucleotide was repeated and its size for the individual allele. Furthermore, it is more desirable to get two bands on the data but sometimes the data also shows minor bands in addition to two major bands, this difference is mostly of two nucleotides from the major bands, and these are called stutter bands [48].
Using PCR primers in research of two multiplex systems consisting of 11 microsatellites associated with parentage testing in goats was characterized [48]. Of these microsatellites, 18 were found to be located on 16 different chromosomes, and these were identified in different animals: five from sheep, nine from cattle, and eight from goats. The parentage exclusion probability was calculated to be higher than 0.99999 and two identical genotypes found probability of less than 10–15. This shows the reliability of microsatellites for parentage testing. The effective number of PCR primers is discussed periodically and currently, the accepted number is 14 [49]. In addition to parentage testing, pedigree verification is also another thing where microsatellites have been a handful [50]. The Murciano-Granadina goat nucleus herd pedigree verification study in Spain of 388 animals resulted in 16.2% (63) being incompatible and 71.9% (279) compatible. The incompatible animals were considered due to data transfer errors, or the archaic system used. These results suggest using the microsatellites for reducing the errors in the breeding programs of goats. A 10% parentage misidentification can lead up to a reduction of 4% in genetic progress [50]. Microsatellites can be a good assistance for the genetic conservation of animal resources, particularly for endangered species, and are being used as a tool for these experiments (Figure 2) [51, 52, 53].
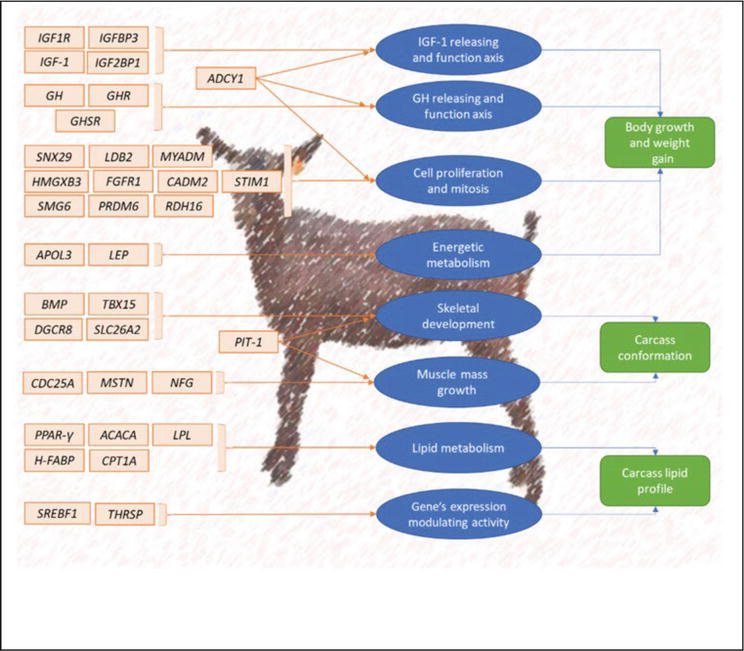
Figure 2.
Figure showing the important candidate genes for meat production in goat genome [
2.2 SNP chip methodology and application
The techniques that followed microsatellites in genome exploring are single nucleotide polymorphisms also known as SNPs. An SNP is a difference of one nucleotide on specific loci on a chromosome. This can happen after a nucleotide is replaced by another at the original place for example thymine (t) being replaced by cytosine (c). The SNPs are present in the genome of all living species, including goats.
The several research projects conducted according to International Goat Genome Consortium (IGGC) guidance, the goat genome was sequenced successfully helping to identify 12 million high-quality variants of SNPs in the genome [55]. This resulted in the creation of an SNP database which contains the technical and biological characteristics information from IGGC (International Goat Genome Consortium) by using advanced SNP detection and bioinformatic tools. The important feature of the SNP database created by IGGC included the selection of minor allele frequencies for diverse breeds, the technological success rate of SNP design, and evenly spaced SNPs in the genome.
The methodology of detecting SNP using the chip is a high throughput, most automated procedure [56]. These are designed on the DNA microarray principle, which contains specific probes depending on the target genome. These SNP probes are hybridized into a DNA sample to check the target allele for SNP. However, the data collected from these arrays are not as complicated as WGS for analysis, requiring bioinformatical software for data processing and analysis. In addition, these SNP arrays can only detect those SNP whose locus on the genome is already characterized, requiring prior genetic information.
The SNPs identified between and within six breeds, Savanna, Alpine, Boer, Saanen, Creole, and Katjang goats, could also be used for the breeds not included in the experiment [57]. The validation for these SNP was conducted by using 52,295 SNPs in the ten goats and was successfully genotyped, which led to a 52 k SNP chip (Illumina, Sandiego, California, USA). These 52 K SNP chip developments created acceleration for advanced goat genome studies. Thus, the turning point came with the evaluation of the economic production trait of goats [24, 58]. A genome-wide association study (GWAS) of the goat genome for important economic traits in the UK followed shortly after the manufacturing of 52 k SNP chip [48]. The GWAS combines phenotypic data like the meat yield of goats with the information collected from the SNP chip [59]. The study carried out in the UK focused on the udder conformation and milk yield traits.
2.3 Use of whole-genome resequencing and methodology
The new developments in next-generation sequencing methods and their reduced cost have gained increased interest in whole-genome genome resequencing as compared to its alternative of genotyping by using SNP chips in breeding programs. Resequencing of whole genome offers a large number of specific variations in the target population as compared to its rival SNP genotyping, which is based on common SNP selected from different populations. Initially, the use of low-cost genotyping by sequencing was tempered after the discovery of the quality of variations obtained from genotyping by sequencing was lower as compared to the quality of variations obtained from genotyping by SNPs due to the lower depth of genome coverage. However, when the genotyping quality was increased with increased genome depth the cost also went up [53]. The characterization of common variants is also available on SNP chips; the use of whole genome sequencing provides advantages that include the characterization of copy number variations, structural variations, and other rare variants.
The basic procedure for next-generation-based whole genome resequencing includes these principal steps, DNA extraction, target enrichment, sequencing, and library preparation. The data obtained from these steps after the sequence is raw data and it further undergoes quality control, demultiplexing, variant identification, mapping for reading the reference genome, and annotation. The above-explained procedure leads to the generation of a variant call file, and after the consistent exhibition of differences in multiple reads, the SNP is called [60].
In goats, comprehensive studies have been carried out to underline the polymorphisms in economically important genes. These candidate genes control the metabolism, physiological pathways, and expression of phenotypes. The important genes are sex determination and proliferation are the SRY gene of the Y chromosome, amelogenin (AMEL), the reproduction-controlling gene FOXL2, and the melatonin receptor gene MTNR1A. The genes bone formation BMP (bone morphogenetic protein), POU1F1 gene for caprine pituitary specific transcription, LEP (leptin), MSTN caprine myostatin, IGF insulin-like growth factor, GH growth hormone, and GHR growth hormone receptor are responsible for body weight muscle growth, body condition, birth weight, body condition, bone formation, and weaning weight. The genes like MC1R melanocortin 1 receptor and KAP keratin-associated proteins are involved in wool production traits. The casein gene family is the major gene that is involved in milk production in goats. The important gene family involved in the immune system response is MHC major histocompatibility complex. The normal function and expression of all these genes are keys to better production in goat farming [37].
The important gene for milk-related traits and milk yield is casein gene family. The genes involved in wool production are melanocortin1 receptor and keratin-associated protein. Major histocompatibility gene family is known to be involved in developing the immune system of animals against the disease and disease resistance [37].
The major candidate genes for milk yield and milk composition traits are the casein gene and their family. Keratin-associated protein (KAP) and melanocortin 1 receptor (MC1R) genes are candidate genes for wool traits. The major histocompatibility complex (MHC) gene is considered important for the immune system and disease-resistance traits. The functions of these genes on economically important traits are different (Figure 3) [37].
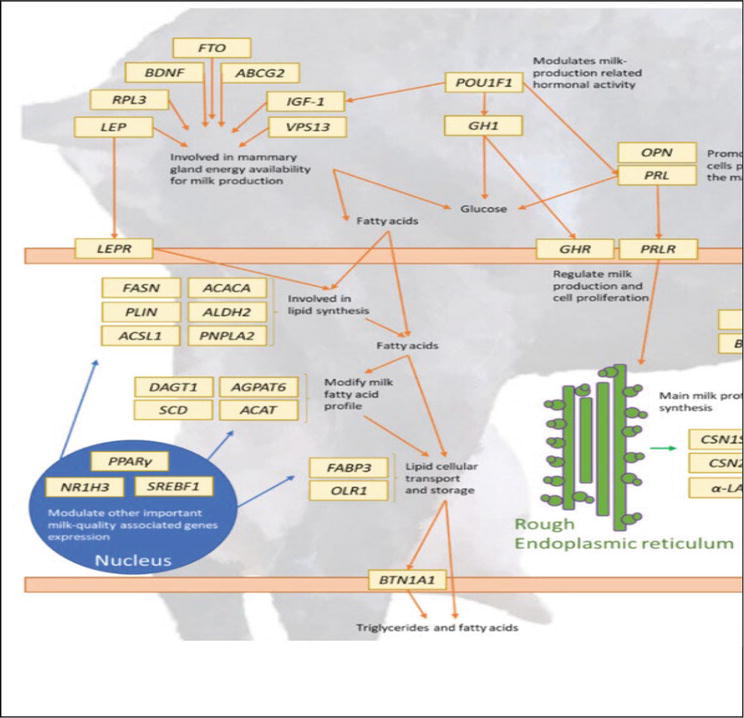
Figure 3.
Schematic diagram showing the milk-related different genes and their relation to each other [
2.4 Genome engineering by the applications of CRISPR/Cas9 in goats
Genome engineering has been revolutionized due to modern tools, which make it possible to engineer genomes more precisely and efficiently with desired results as compared to conventional genome modification tools. One of these advanced genome engineering tools is CRISPR/cas9, which has been applied to goat and sheep genomes to fulfill the desired targets. Until today, numerous models for sheep and goats have been engineered through CRISPR/cas9 systems. Further studies are continuing to provide useful models of sheep and goats for the service of biomedicine and agricultur (Figure 4).
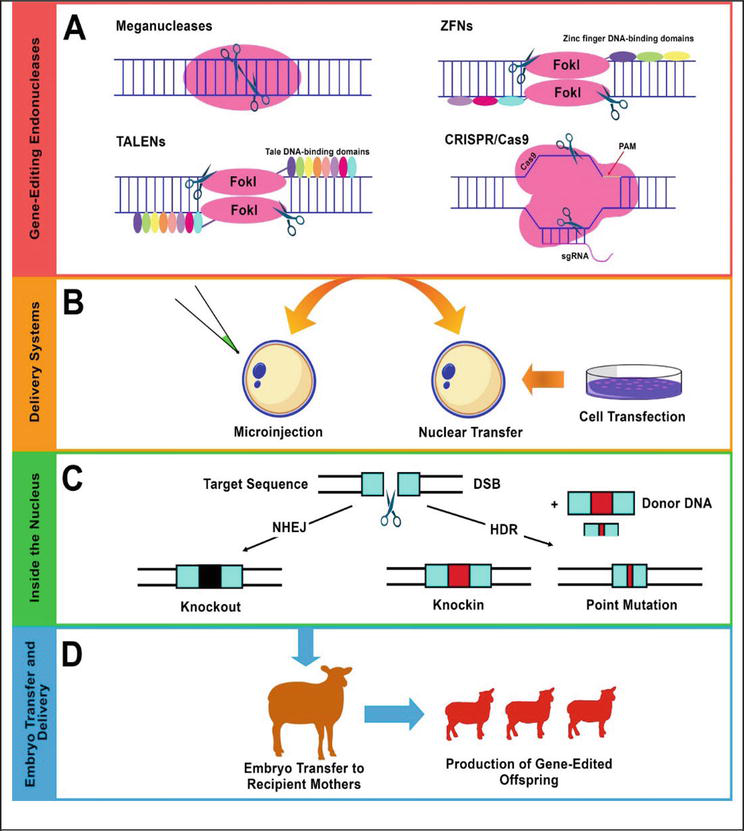
Figure 4.
The visualization of procedure involving TALENS, ZFNs, and CRISPR/cas9a [
Accelerating the growth rate and enhancing the body weight of livestock animals have been the key goals of farm agriculture. The genes that are responsible for these economic traits are the key targets for advanced genome engineering technologies. The first gene to be targeted by CRISPR/Cas9 is MSTN, in accordance to achieve the goal of engineering the most important gene modification in goats and sheep.
Furthermore, the steps are being checked in order to make it safe for engineering the genome in big animals, for this, a trio-based sequencing has been carried out to investigate the variations discovered in edited samples, which could have been naturally obtained, parentally inherited or a result from specific target occurrence [60]. The results of the experiment showed a negligible amount of off-target editing, which does not affect the use of CRISPR/Cas9 in large livestock. These results provide information about the potential of multiplex editing by CRISPR/Cas9 in large animals.
The methodology of the CRISPR/Cas9 system of gene editing involves creating a double-strand break in the DNA by the Cas9 protein. The target gene that needs to be disrupted is cut by DSB. The cleaved DNA segment can be repaired by two pathways NHEJ (non-homologous end joining) and HDR (homology-directed repair). Nonhomologous end joining is used to create knockouts for the genes by deleting some nucleotides from the DNA sequence of the gene. However, the homology-directed repair will lead to the insertion of predicted DNA segments. The DSBs created in the DNA sequence of a gene make it possible to delete one or more nucleotides and also can be used to insert a few nucleotides or a segment of DNA in the genome of the target animal at the specific target site [62].
CRISPR/Cas9 was used to edit 4 important genes in goats PrP, MSTN, nucleoporin 155 (NUP155), and BLG, in goat fibroblast, and three knouts for MSTN in goats were generated by using SCNT [63]. The end results of the study showed an efficiency ranging from 9 to 70% by using CRISPR/Cas9, which indicates the potential of this technology to be used in the caprine system. Later than this further knouts for fibroblast growth factor and MSTN in goats or for both have been reported [64]. From the total of 98 individual animals obtained, only 14 lambs died shortly after birth while 79 lambs were alive and five lambs were aborted. Furthermore, 10.2 % (10/98) showed disruption in both genes, 21.4% ( 21/98) showed disruption in FGF5, and 15.3% (15/98) showed disruption in MSTN. These results approve the ability about the efficacy and efficiency of CRISPR/Cas multiplex targeting in large farm animals. Specifically, because the economic traits are controlled by multiple genes in different locations. Further investigations were also conducted on these founders’ edited mutants from the above study to check the viability and authenticity of knockout alleles transmission and gene disruption [65]. In addition, the MSTN-disrupted goats were also analyzed for transcriptomic changes [66]. The transmission and occurrence of the disrupted genes were confirmed, and also considerable changes at the transcriptome level and gene expression level were also confirmed. The changes at the expression level were recorded in the unsaturated fatty acid biosynthesis and fatty acid metabolism, which suggests that MSTN plays and regulatory role in the expression of these genes. Moreover, the researchers also conducted trio-based family sequencing of the engineered progenies and goats to look for any indels, structural variants, and other
The occurrence of FGF5 knockout simultaneously at genetic and morphological levels has been confirmed and an increased secondary hair follicle number and high fiber length were also seen [68]. Another study was conducted to check the disruption effect and hair follicle development and growth phenotype by creating a goat knockout through SCNT, CRISPR/Cas9 was used to disrupt the EDAR ectodysplasin receptor gene. The knockout EDAR genes generated showed the absence of top hairs on heads and primary abnormal hair follicles, these are the specific features of EDAR knockouts. These founders of EDAR knouts generated by CRISPR/Cas9 provide a model to study the relationship between hair follicle growth and development with the EDAR gene. The study of hair follicle growth and development genes is of primary importance because they are the key features of cashmere and wool-producing goat breeds. FGF5 gene engineering using CRISPR/Cas was carried out by introducing nonsense codon introgression in the gene to increase the production of cashmere hair goats [69].
The functional role of some genes like acetyl CoA acyltransferase 2 ACAA2 in sheep adipocyte cells and in the mammary epithelial cells of goat gene stearoyl-CoA desaturase 1 has also been investigated by using CRISPR/Cas9 [70, 71]. These genes are known to affect directly or indirectly related to fatty acid metabolism and milk traits. CRISPR/Cas9 has also been used to produce sheep and goats with modified milk production characteristics that can be very important for the large-scale production of important pharmaceuticals and proteins in their milk.
In another study, a defined point mutation was introduced in the goat genome by using CRISPR/Cas9 in the GDF9 growth differentiation factor 9, which is largely related to litter size and ovulation rate [72]. The study showed a targeting efficiency of 22.2% with 4 kids carrying the mutation out of 18 injected. In short, both of the above studies showed very successful cases of inducing a reliable and specific point mutation in livestock by using CRISPR/Cas9-induced HDR. In another investigation, they checked the effect and efficiency of open-pulled straw vitrification as a technique for preserving microinjected embryos over the reproductive capacity of AANAT transgenic offspring and AANAT microinjected embryo development [73].
Furthermore, CRISPR/Cas9 technology has been used to target fibroblast PrP gene and generate PrP gene knockout donor cells, which will then be applied with SCNT to produce goats with PrP resistance [60, 74, 75]. The target efficiency was recorded at 20% when both of the genes MSTN and PrP were targeted simultaneously, and this target efficiency was increased to 70% when only a single PrP was targeted. Thus, the results of these experiments suggest CRISPR/Cas9 technology can be highly used to produce disease resistance in domestic and commercial livestock.
3. Conclusion
The recent past has seen numerous technological advancements in the goat genetics and breeding field. These technologies have been successful in helping goat breeders to increase the productivity of goats. Goat genome studies have been the key focus of most scientists, which helped the technologies to research through genomic regions and exploit them for increasing the meat, milk, and wool production in the goat. Genome modification tools have become necessary in recent years for research in agriculture, biomedicine, and model studies. Genome engineering tool such as CRISPR/Cas9 provides an immense number of opportunities for revolutionizing farm and agriculture research. Therefore, applying these research tools to goats will create significant results. The use of CRISPR/Cas9 to generate genetically modified animals like goats, sheep, and cattle is currently going on in the world and the list will keep on growing.
References
- 1.
FAO. The second report on the state of the world’s animal genetic resources for food and agriculture. In: Scherf BD, Pilling D, editors. FAO Commission on Genetic Resources for Food and Agriculture Assessments. Rome: FAO; 2015. Available from: http://www.fao.org/3/a-i4787e/index.html - 2.
Naderi S, Rezaei HR, Pompanon F, Blum MG, Negrini R, Naghash HR, et al. The goat domestication process inferred from large-scale mitochondrial DNA analysis of wild and domestic individuals. Proceedings of the National Academy of Sciences of the United States of America. 2008; 105 (46):17659-17664. DOI: 10.1073/pnas.0804782105 - 3.
Gkiasta M, Russell T, Shennan S, Steele J. Neolithic transition in Europe: The radiocarbon record revisited. Antiquity. 2003; 77 (295):45-62. DOI: 10.1017/S0003598X00061330 - 4.
Newman JL. The Peopling of Africa: A Geographic Interpretation. New Haven and London: Yale University Press; 1995. p. 252. ISBN:0-300-06003-3. Available from: https://books.google.pl/books?hl=en&lr=&id=pDjlC1ws158C&oi=fnd&pg=PA1&dq=Newman+JL.+The+Peopling+of+Africa:++A+Geographic+Interpretation.+Yale++University+Press%3B+1995.+p.+252&ots=vcOLhEjjsA&sig=ndDzuX2cNc29yRzV3T0FOy6sWBg&redir_esc=y#v=onepage&q&f=false - 5.
Clutton-Brock J. Cattle, sheep, and goats south of the Sahara: An archaeo- zoological perspective. In: Blench RM, MacDonald KC, editors. The Origins and Development of African Livestock: Archaeology, Genetics, Linguistics and Ethnography. London: UCL Press; 2000. pp. 30-37 - 6.
Hassan FA. Climate and cattle in North Africa: A first approximation. In: Blench RM, MacDonald KC, editors. The Origins and Development of African Livestock: Archaeology, Genetics, Linguistics and Ethnography. London: UCL Press; 2000. pp. 61-86 - 7.
Yuan J, Jian-Lin H, Blench R. Livestock in ancient China: An archaeo zoological perspective. In: Past Human Migrations in East Asia. Routledge; 2008. pp. 116-136 - 8.
Porter V. Goats of the world. Ipswich, UK: Farming Press; 1996. p. 179 - 9.
Meadow RH. The origins and spread of agriculture and pastoralism in northwestern South Asia. In: The Origins and Spread of Agriculture and Pastoralism in Eurasia. Edited by D.R. Harris. 1996. pp. 390-412. ISBN: 9781857285383 - 10.
Zuidhof MJ, Schneider BL, Carney VL, Korver DR, Robinson FE. Growth, efficiency, and yield of commercial broilers from 1957, 1978, and 2005. Poultry Science. 2014; 93 (12):2970-2982. DOI: 10.3382/ps.2014-04291 - 11.
Rauw WM, Kanis E, Noordhuizen-Stassen EN, Grommers FJ. Undesirable side effects of selection for high production efficiency in farm animals; a review. Livestock Production Science. 1998; 56 (1):15-33. DOI: 10.1016/S0301-6226(98)00147-X - 12.
Merks JW, Mathur PK, Knol EF. New phenotypes for new breeding goals in pigs. Animal. 2012; 6 (4):535-543. DOI: 10.1017/S1751731111002266 - 13.
Byrne TJ, Ludemann CI, Amer PR, Young MJ. Broadening breeding objectives for maternal and terminal sheep. Livestock Science. 2012; 144 (1-2):20-36. DOI: 10.1016/j.livsci.2011.10.010 - 14.
Cole JB, Eaglen SAE, Maltecca C, Mulder HA, Pryce JE. The future of phenomics in dairy cattle breeding. Animal Frontiers. 2020; 10 (2):37-44. DOI: 10.1093/af/vfaa007 - 15.
Hayward SJ. Evaluation of six breeding lines of milk goats. The Journal of Agriculture of the University of Puerto Rico. 1962; 46 (3):205-212. DOI: 10.46429/jaupr.v46i3.12336 - 16.
Ronningen K. Causes of variation in the flavour intensity of goat milk. Acta Agriculturæ Scandinavica. 1965; 15 (3-4):301-342. DOI: 10.1080/00015126509433125 - 17.
Wiggans GR, Van Vleck LD, Dickinson FN. Projection factors for goat lactation records. Journal of Dairy Science. 1979; 62 (5):797-801. DOI: 10.3168/jds.S0022-0302(79)83328-7 - 18.
Wiggans GR, Dickinson FN, King GJ, Weller JI. Genetic evaluation of dairy goat bucks for daughter milk and fat. Journal of Dairy Science. 1984; 67 (1):201-207. DOI: 10.3168/jds.S0022-0302(84)81285-0 - 19.
Wiggans GR, van Dijk J, Misztal I. Genetic evaluation of dairy goats for milk and fat yield with an animal model. Journal of Dairy Science. 1988; 71 (5):1330-1337. DOI: 10.3168/jds.S0022-0302(88)79690-3 - 20.
Kemp SJ, Brezinsky L, Teale AJ. A panel of bovine, ovine and caprine polymorphic microsatellites. Animal Genetics. 1993; 24 (5):363-365. DOI: 10.1111/j.1365-2052.1993.tb00341.x - 21.
Vaiman D, Schibler L, Bourgeois F, Oustry A, Amigues Y, Cribiu EP. A genetic linkage map of the male goat genome. Genetics. 1996; 144 (1):279-305. DOI: 10.1093/genetics/144.1.279 - 22.
Arevalo E, Holder DA, Derr JN, Bhebhe E, Linn RA, Ruvuna F, et al. Caprine microsatellite dinucleotide repeat polymorphisms at the SR-CRSP-1, SR-CRSP-2, SR-CRSP-3, SR-CRSP-4 and SR-CRSP-5 loci. Animal Genetics. 1994; 25 (3):202. DOI: 10.1111/j.1365-2052.1994.tb00124.x - 23.
Schibler L, Vaiman D, Oustry A, Guinec N, Dangy-Caye AL, Billault A, et al. Construction and extensive characterization of a goat bacterial artificial chromosome library with threefold genome coverage. Mammalian Genome. 1998; 9 (2):119-124. DOI: 10.1007/s003359900701 - 24.
Mucha S, Mrode R, Coffey M, Kizilaslan M, Desire S, Conington J. Genome-wide association study of conformation and milk yield in mixed-breed dairy goats. Journal of Dairy Science. 2018; 101 (3):2213-2225. DOI: 10.3168/jds.2017-12919 - 25.
Bickhart DM, Rosen BD, Koren S, Sayre BL, Hastie AR, Chan S, et al. Single-molecule sequencing and chromatin conformation capture enable de novo reference assembly of the domestic goat genome. Nature Genetics. 2017; 49 (4):643-650. DOI: 10.1038/ng.3802 - 26.
Carillier C, Larroque H, Palhière I, Clément V, Rupp R, Robert-Granié C. A first step toward genomic selection in the multi-breed French dairy goat population. Journal of Dairy Science. 2013; 96 (11):7294-7305. DOI: 10.3168/jds.2013-6789 - 27.
Dong Y, Xie M, Jiang Y, Xiao N, Du X, Zhang W, et al. Sequencing and automated whole-genome optical mapping of the genome of a domestic goat (Capra hircus). Nature Biotechnology. 2013; 31 (2):135-141. DOI: 10.1038/nbt.2478 - 28.
Tosser-Klopp G. Goat Genome Assembly, Availability of an International 50K SNP Chip and RH Panel: An Update of the International Goat Genome Consortium projects. Workshop Plant et Animal Genome; 2012; San Diégo - Californie, United States - 29.
Fontanesi L, Martelli PL, Beretti F, Riggio V, Dall'Olio S, Colombo M, et al. An initial comparative map of copy number variations in the goat ( Capra hircus ) genome. BMC Genomics. 2010;11 (1):1-15. DOI: 10.1186/1471-2164-11-639 - 30.
Li XL, Wu ZL, Liu ZZ, Gong YF, Zhou RY, Zheng GR. SNP identification and analysis in part of intron 2 of goat MSTN gene and variation within and among species. The Journal of Heredity. 2006; 97 (3):285-289. DOI: 10.1093/jhered/esj026 - 31.
Andonov S, Ødegård J, Svendsen M, Ådnøy T, Vegara M, Klemetsdal G. Comparison of random regression and repeatability models to predict breeding values from test-day records of Norwegian goats. Journal of Dairy Science. 2013; 96 (3):1834-1843. DOI: 10.3168/jds.2012-5910 - 32.
Barbieri ME, Manfredi E, Elsen JM, Ricordeau G, Bouillon J, Grosclaude F, et al. Influence of the αs1 casein locus on milk performance and genetic parameters of Alpine goats. Genetics Selection Evolution. 1995; 27 (5):437-450 - 33.
Sise JA, Penty JM, Montgomery GW, Gootwine E. The duplicated gene copy of the ovine growth hormone gene contains a PvuII polymorphism in the second intron. Animal Genetics. 1993; 24 (4):319-321. DOI: 10.1111/j.1365-2052.1993.tb00321.x - 34.
Barillet F, Manfredi E, Elsen JM. Genetic variations and associations for improving milk production and milk product qualities in sheep and goats. In: Proceedings of the Sixth World Congress on Genetics Applied to Livestock Production; Armidale, Australia. 1998. pp. 129-136 - 35.
Moioli B, D’Andrea M, Pilla F. Candidate genes affecting sheep and goat milk quality. Small Ruminant Research. 2007; 68 (1-2):179-192. DOI: 10.1016/j.smallrumres.2006.09.008 - 36.
Tambasco DD, Paz CCP, Tambasco-Studart M, Pereira AP, Alencar MM, Freitas AR, et al. Candidate genes for growth traits in beef cattle crosses Bos taurus× Bos indicus. Journal of Animal Breeding and Genetics. 2003; 120 (1):51-56. DOI: 10.1046/j.1439-0388.2003.00371.x - 37.
Supakorn C. The important candidate genes in goats—A review. Walailak Journal of Science and Technology. 2009; 6 (1):17-36 - 38.
Malveiro E, Marques PX, Santos IC, Belo C, Cravador A. Association between SSCPs at Algarvia goat GH gene and milk traits. Archivos de zootecnia. 2001; 50 (190):49-57 - 39.
Li XL, Wu ZL, Gong YF, Liu YQ , Liu ZZ, Wang XJ, et al. Single-nucleotide polymorphism identification in the caprine myostatin gene. Journal of Animal Breeding and Genetics. 2006; 123 (2):141-144. DOI: 10.1111/j.1439-0388.2006.00572.x - 40.
Gupta N, Ahlawat SPS, Kumar D, Gupta SC, Pandey A, Malik G. Single nucleotide polymorphism in growth hormone gene exon-4 and exon-5 using PCR-SSCP in Black Bengal goats–A prolific meat breed of India. Meat Science. 2007; 76 (4):658-665. DOI: 10.1016/j.meatsci.2007.02.005 - 41.
Lan XY, Pan CY, Chen H, Zhao M, Li JY, Yu J, et al. The novel SNPs of the IGFBP3 gene and their associations with litter size and weight traits in goat (brief report). Archives of Animal Breed. 2007; 50 :223-224. DOI: 10.5194/aab-50-223-2007 - 42.
Lan XY, Pan CY, Chen H, Zhang CL, Li JY, Zhao M, et al. An AluI PCR-RFLP detecting a silent allele at the goat POU1F1 locus and its association with production traits. Small Ruminant Research. 2007; 73 (1-3):8-12. DOI: 10.1016/j.smallrumres.2006.10.009 - 43.
Fang X, Xu H, Zhang C, Chen H, Hu X, Gao X, et al. Polymorphism in BMP4 gene and its association with growth traits in goats. Molecular Biology Reports. 2009; 36 (6):1339-1344. DOI: 10.1007/s11033-008-9317-1 - 44.
Vaiman D, Koutita O, Oustry A, Elsen JM, Manfredi E, Fellous M, et al. Genetic mapping of the autosomal region involved in XX sex-reversal and horn development in goats. Mammalian Genome. 1996; 7 (2):133-137. DOI: 10.1007/s003359900033 - 45.
Van Arendonk JA, Tier B, Kinghorn BP. Use of multiple genetic markers in prediction of breeding values. Genetics. 1994; 137 (1):319-329. DOI: 10.1093/genetics/137.1.319 - 46.
Rasmusen BA, Beece CK, Christian LL. Halothane sensitivity and linkage of genes for H red blood cell antigens, phosphohexose isomerase (PHI) and 6-phosphogluconate dehydrogenase (6-PGD) variants in pigs. Animal Blood Groups and Biochemical Genetics. 1980; 11 (2):93-107. DOI: 10.1111/j.1365-2052.1980.tb01499.x - 47.
Fujii J, Otsu K, Zorzato F, de Leon S, Khanna VK, Weiler JE, et al. Identification of a mutation in porcine ryanodine receptor associated with malignant hyperthermia. Science. 1991; 253 (5018):448-451. DOI: 10.1126/science.1862346 - 48.
Meuwissen T, Hayes B, Goddard M. Genomic selection: A paradigm shift in animal breeding. Animal Frontiers. 2016; 6 (1):6-14. DOI: 10.2527/af.2016-0002 - 49.
Luikart G, Biju-Duval MP, Ertugrul O, Zagdsuren Y, Maudet C, Taberlet P. Power of 22 microsatellite markers in fluorescent multiplexes for parentage testing in goats (Capra hircus). Animal Genetics. 1999; 30 (6):431-438 - 50.
White S, Genestout L, Penedo C. Applied genetics in sheep and goats. ISAG Standing Communication. 2012; 2012 :1-4 - 51.
Jiménez-Gamero I, Dorado G, Muñoz-Serrano A, Analla M, Alonso-Moraga A. DNA microsatellites to ascertain pedigree-recorded information in a selecting nucleus of Murciano-Granadina dairy goats. Small Ruminant Research. 2006; 65 (3):266-273. DOI: 10.1016/j.smallrumres.2005.07.019 - 52.
Pépin L, Amigues Y, Lépingle A, Berthier JL, Bensaid A, Vaiman D. Sequence conservation of microsatellites between Bos taurus (cattle), Capra hircus (goat) and related species. Examples of use in parentage testing and phylogeny analysis. Heredity (Edinb). 1995; 74 (Pt 1):53-61. DOI: 10.1038/hdy.1995.7 - 53.
Ajmone-Marsan P, Colli L, Han JL, Achilli A, Lancioni H, Joost S, et al. The characterization of goat genetic diversity: Towards a genomic approach. Small Ruminant Research. 2014; 121 (1):58-72. DOI: 10.1016/j.smallrumres.2014.06.010 - 54.
Salgado Pardo JI, Delgado Bermejo JV, González Ariza A, León Jurado JM, Marín Navas C, Iglesias Pastrana C, et al. Candidate genes and their expressions involved in the regulation of Milk and meat production and quality in goats (Capra hircus). Animals (Basel). 2022; 12 (8):988. DOI: 10.3390/ani12080988 - 55.
Tosser-Klopp G, Bardou P, Bouchez O, Cabau C, Crooijmans R, Dong Y, et al. Design and characterization of a 52K SNP chip for goats. PLoS One. 2014; 9 (1):e86227. DOI: 10.1371/journal.pone.0086227 - 56.
Hasan MU, Ceyhan A. Use of molecular markers in parentage testing, genetic diversity, marker-assisted breeding and disease identification. Journal of Agriculture, Food, Environment and Animal Sciences. 2022; 2 (1):1-15 - 57.
Lashmar SF, Visser C, Van Marle-Köster E. Validation of the 50k Illumina goat SNP chip in the South African Angora goat. South African Journal of Animal Science. 2015; 45 (1):56-59 - 58.
Juditsky AB, Nazin AV, Tsybakov AB, Vayatis N. Recursive aggregation of estimators by the mirror descent algorithm with averaging. Problems of Information Transmission. 2005; 41 (4):368-384. DOI: 10.48550/arXiv.math/0505333 - 59.
Gorjanc G, Cleveland MA, Houston RD, Hickey JM. Potential of genotyping-by-sequencing for genomic selection in livestock populations. Genetics Selection Evolution. 2015; 47 (1):1-14. DOI: 10.1186/s12711-015-0102-z - 60.
Jian Y, Li M. A narrative review of single-nucleotide polymorphism detection methods and their application in studies of Staphylococcus aureus. Journal of Bio-X Research. 2021; 4 (01):1-9. DOI: 10.1097/JBR.0000000000000071 - 61.
Wang X, Liu J, Niu Y, Li Y, Zhou S, Li C, et al. Low incidence of SNVs and indels in trio genomes of Cas9-mediated multiplex edited sheep. BMC Genomics. 2018; 19 (1):1-8. DOI: 10.1186/s12864-018-4712-z - 62.
Hasan MU, Kozakli Ö, Ceyhan A. Possibilities of using CRISPR-based genome editing Technologies in Livestock. Journal of Agriculture, Food, Environment and Animal Sciences. 2022; 3 (1):56-68 - 63.
Ni W, Qiao J, Hu S, Zhao X, Regouski M, Yang M, et al. Efficient gene knockout in goats using CRISPR/Cas9 system. PLoS One. 2014; 9 (9):e106718. DOI: 10.1371/journal.pone.0106718 - 64.
Wang X, Yu H, Lei A, Zhou J, Zeng W, Zhu H, et al. Generation of gene-modified goats targeting MSTN and FGF5 via zygote injection of CRISPR/Cas9 system. Scientific Reports. 2015; 5 (1):1-9. DOI: 10.1038/srep13878 - 65.
Wang X, Niu Y, Zhou J, Zhu H, Ma B, Yu H, et al. CRISPR/Cas9-mediated MSTN disruption and heritable mutagenesis in goats causes increased body mass. Animal Genetics. 2018b; 49 (1):43-51. DOI: 10.1111/age.12626 - 66.
Wang L, Cai B, Zhou S, Zhu H, Qu L, Wang X, et al. RNA-seq reveals transcriptome changes in goats following myostatin gene knockout. PLoS One. 2017; 1249 (12):e0187966. DOI: 10.1371/journal.pone.0187966 - 67.
Li C, Zhou S, Li Y, Li G, Ding Y, Li L, et al. Trio-based deep sequencing reveals a low incidence of off-target mutations in the offspring of genetically edited goats. Frontiers in Genetics. 2018; 4 (9):449. DOI: 10.3389/fgene.2018.00449 - 68.
Wang X, Cai B, Zhou J, Zhu H, Niu Y, Ma B, et al. Disruption of FGF5 in cashmere goats using CRISPR/Cas9 results in more secondary hair follicles and longer fibers. PLoS One. 2016; 11 (10):e0164640. DOI: 10.1371/journal.pone.0164640 - 69.
Li G, Zhou S, Li C, Cai B, Yu H, Ma B, et al. Base pair editing of goat embryos: Nonsense codon introgression into FGF5 to improve cashmere yield. bio. 2018; 2018 :348441. DOI: 10.1101/348441 - 70.
Zhang Y, Wang Y, Wang X, Ji Y, Cheng S, Wang M, et al. Acetyl-coenzyme a acyltransferase 2 promote the differentiation of sheep precursor adipocytes into adipocytes. Journal of Cellular Biochemistry. 2019d; 20 (5):8021-8031. DOI: 10.1002/jcb.28080 - 71.
Tian H, Luo J, Zhang Z, Wu J, Zhang T, Busato S, et al. CRISPR/Cas9-mediated stearoyl-CoA desaturase 1 (SCD1) deficiency affects fatty acid metabolism in goat mammary epithelial cells. Journal of Agricultural and Food Chemistry. 2018; 66 (38):10041-10052. DOI: 10.1021/acs.jafc.8b03545 - 72.
Niu Y, Zhao X, Zhou J, Li Y, Huang Y, Cai B, et al. Efficient generation of goats with defined point mutation (I397V) in GDF9 through CRISPR/Cas9. Reproduction, Fertility, and Development. 2018; 30 (2):307-312. DOI: 10.1071/RD17068 - 73.
Tian X, Lv D, Ma T, Deng S, Yang M, Song Y, et al. AANAT transgenic sheep generated via OPS vitrified-microinjected pronuclear embryos and reproduction efficiency of the transgenic offspring. PeerJ. 2018; 6 :e5420. DOI: 10.7717/peerj.5420 - 74.
Hu S, Yang M, Polejaeva I. 360 double knockout of goat myostatin and prion protein gene using clustered regularly interspaced short palindromic repeat (CRISPR)/Cas9 systems. Reproduction, Fertility and Development. 2015; 27 (1):268-268. DOI: 10.1071/RDv27n1Ab360 - 75.
Fan Z, Yang M, Regouski M, Polejaeva IA. Gene knockouts in goats using CRISPR/Cas9 system and somatic cell nuclear transfer. Methods in Molecular Biology. 2019; 1874 :373-390. DOI: 10.1007/978-1-4939-8831-0_22