Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
This chapter was designed to figure out the basic knowledge about domestication, adaptation and immigration of goat breeds, with a spotlight on modern versus old strategies to enhance genetic performance along with recognizing their role in the livestock production industry with a special focus on the position adaptive selection in view of the new high technologies of investigating genome and building of selective comparison between goat breeds. Also, this chapter focused on goat production throughout the world, the vital role played by goats and the biodiversity of goat genetic resources and the special characteristics of goats under different conditions. In addition, this chapter is concerned with modern strategies to enhance goat genetic performance, including different molecular tools, besides mentioning the outcomes of utilizing advanced molecular tools in goat breeding, as well as identification of candidate genes related to important economic traits, detection of signatures of selection and quantitative trait loci, applied genome-wide association studies and the methodology of genomic selection, where the recent findings of genomic studies on goats are listed in a logical and sequential fashion.
Faculty of Agriculture (Alshatby), Animal and Fish Production Department, Alexandria University, Alexandria City, Egypt
Amr M.A. Rashad
Faculty of Agriculture (Alshatby), Animal and Fish Production Department, Alexandria University, Alexandria City, Egypt
Nada N.A.M. Hassanine
College of Animal Science and Technology, Chongqing Key Laboratory of Forage and Herbivore, Chongqing Engineering Research Centre for Herbivores Resource Protection and Utilization, Southwest University, Chongqing, P.R. China
Mahmoud A. Sharaby
Faculty of Agriculture (Alshatby), Animal and Fish Production Department, Alexandria University, Alexandria City, Egypt
Sobhy M.A. Sallam
Faculty of Agriculture (Alshatby), Animal and Fish Production Department, Alexandria University, Alexandria City, Egypt
*Address all correspondence to: elemlak1339@gmail.com
1. Introduction
Goats play an important role in the livelihood of a large proportion of small and marginal holders [1, 2, 3]. Moreover, the genetic diversity of goat breeds is indispensable to meet current production needs under various environments, allows sustainable genetic improvement and facilitates rapid adaptation to changing breeding objectives. Characterization and determination of genetic differences between and within goat breeds are potential tools to help the rapid improvement of economically important traits [4, 5, 6].
Attempts were made to increase meat, milk and fibre (hair and cashmere) production of goats. Most of the attempts were made traditionally through changing production systems and reproductive management but recently, by introducing high prolificacy genes and detecting the changes in the whole genome which affect fertility, fibre and growth traits [7, 8].
There are many minor genes scattered across the genome that influence most goat traits of economic importance, as well as specific environmental factors. Traditionally, phenotypic information was used to improve quantitative traits, but now phenotypic and genotypic information is needed to improve quantitative traits. Traditionally, pedigree-assisted selection and progeny testing have been used widely to improve many of these production traits. However, these traits are mostly low or medium heritable which may slow down improvement rates. Recent researches proved that some of such traits could be controlled by major genes or candidate genes, signatures of selection (SS), molecular markers, selective sweeps and quantitative trait loci (QTL) which can be traced along with their influence by the application of some advanced molecular tools, such as next-generation sequencing (NGS) or whole-genome sequencing (WGS) or high-throughput single-nucleotide polymorphism (SNP) genotyping [9, 10].
In general, traditional animal breeding techniques for improving such traits of concern are usually costly and take a long time and some undesirable traits may appear in offspring throughout the process of improvement. The genomic selection (GS) based on second-generation sequencing (next-generation sequencing) and third-generation sequencing (single-molecule real-time, SMRT) are promising alternatives for improvement that enable the animal breeders to select eligible animals with desirable traits such as longevity, fertility, litter size and disease resistance at early ages. This will result in an increase in the accuracy of the selection response [11].
During the past few decades, advances in molecular genetics have led to the identification of multiple genes, genetic markers, signatures of selection, selective sweeps, candidate genes associated with traits of interest in livestock also, detection of QTLs, obtaining the mitochondrial DNA (mtDNA) and describing the whole-genome sequencing (WGS), thus, marker-assisted selection (MAS), genome-wide studies (GWS), genome-wide association studies (GWAS) and genomic selection (GS).
This has provided opportunities to enhance the response of selection, in particular for traits that are difficult to improve by conventional selection such as low heritable traits or those whose phenotype measurement on selection candidates is difficult, expensive, only possible late in life or not possible [12, 13].
The general aims of this chapter were to introduce fundamental knowledge about the history, immigration of goat and modern versus old strategies to enhance goat genetic performance. Also, investigating the vital role played by goat, genetic diversity in goat, molecular tools in goat breeding, major genes related to economic traits and adaptation, the methodology of genomic selection (GS) and recent genome studies on goat by extracting the most important findings in the reliable studies with focusing on the recent studies.
There were several objectives in the current chapter: 1) investigate the worldwide goat distribution, 2) assess the worldwide goat production between the past and the present, 3) take a deep spotlight on the history and immigration of goats, 4) follow the goat domestication from the domestication centre to the different continents in the world, 5) evaluate modern versus old strategies to enhance goat genetic performance, 6) searching deeply in the recent reliable studies and genetic databases about the major genes or candidate genes, signatures of selection (SS), molecular markers, selective sweeps and quantitative trait loci (QTL) which associated with most important economic traits in the goats, 7) proposing a methodology to apply the genomic selection in the view of recent and reliable genome-wide association studies, 8) investigating the outcomes of utilizing advanced molecular tools in goat breeding and 9) finally, preparing a remarkable chapter as a reference guide for preserving the effort and time required to search hundreds of investigations and studies related to goat science.
2.2 Investigation sources
More than 590 references (varied between published papers, scientific periodicals, international books, master’s and doctoral dissertations) published via Springer, Elsevier, Intech Open, Wiley, Taylor & Francis publishers, etc. were thoroughly investigated to extract all possible knowledge about the goats.
Also, the global databases related to animal production sector (especially goats) and genome databases were also used, as follows:
Goat is one of the oldest domesticated animals. According to genetic data and archaeo-zoological references, goats have been domesticated 10,000 years ago, in the region from Eastern-Anatolia to the Mountains of Zagros north of Iran (Figure 1), while the archaeo-zoological evidence suggests that they were domesticated from 8000 to 9000 years ago in the near east [14, 15].
Figure 1.
Goat domestication in the region from eastern-Anatolia in Turkey to the mountains of Zagros in the north of Iran in the past 10,000 years (developed by the authors).
Goats are among the big five livestock species (chickens, goat, sheep, cattle and pigs) recognized by the FAO [16, 17]. FAO Database contains more than 800,000,000 specimens from more than 1200 goat breeds throughout the world. Globally, there are more than one billion goats. Goat is referred to as a poor man’s cow [18]. Domesticated goats are generally utilized for producing meat, milk, fat, skin, hides, fibre or cashmere [18] worldwide [19, 20] (Table 1), and are also used for transport [20].
Region
No. of goats
Meat Pro. (Tones)
Meat pro. (Head)
Fat pro. (Tones)
Fat pro. (Head)
Skin pro. (Tones)
Skin pro. (Head)
Milk pro. (Tones)
Milk pro. (Head)
World
—
6,142,140
495,108,884
247,626
495,108,884
1,232,880
495,108,884
20,629,610
220,921,370
Africa
489,021,886
—
—
40,541
131,262,796
263,922
131,262,796
4,487,005
86,250,504
Eastern Africa
183,591,469
503,788
48,064,791
16,730
48,064,791
111,463
48,064,791
1,683,703
26,597,994
Middle Africa
62,667,249
215,838
17,317,883
5483
17,317,883
32,579
17,317,883
171,813
2,780,743
Northern Africa
47,874,919
205,372
19,824,364
5765
19,824,364
38,763
19,824,364
1,582,893
23,969,051
Southern Africa
9,452,259
21,558
1,512,674
485
1,512,674
—
—
12,374
101,280
Western Africa
185,435,990
—
—
12,078
44,543,084
78,524
44,543,084
1,036,222
32,801,436
Americas
39,194,276
130,103
9,607,137
5022
9,607,137
25,582
9,607,137
801,285
8,906,470
Northern America
2,685,122
9207
624,400
359
624,400
1777
624,400
25,982
257,878
Central America
9,007,506
40,516
2,254,084
1413
2,254,084
8087
2,254,084
166,208
780,315
Caribbean
3,439,056
9053
621,356
149
621,356
1278
621,356
252,818
1,221,427
South America
24,062,592
71,327
6,107,297
3101
6,107,297
14,440
6,107,297
356,277
6,646,850
Asia
579,347,344
4,486,883
344,195,895
195,480
344,195,895
918,554
344,195,895
12,219,732
114,335,466
Central Asia
10,986,896
38,152
2,099,490
1102
2,099,490
5346
2,099,490
56,111
548,841
Eastern Asia
165,473,380
2,452,793
162,739,830
96,363
162,739,830
498,988
162,739,830
442,160
8,039,210
Southern Asia
326,303,601
1,477,785
142,467,519
79,171
142,467,519
318,717
142,467,519
10,146,318
—
South-eastern Asia
37,545,555
—
—
8875
18,929,297
49,997
18,929,297
399,648
9,329,661
Western Asia
39,037,912
272,881
17,959,759
9969
17,959,759
45,506
17,959,759
1,175,495
14,998,180
Europe
16,241,452
88,129
8,019,723
4211
8,019,723
16,600
8,019,723
3,121,548
11,427,563
Eastern Europe
4,752,310
29,676
2,121,799
1257
2,121,799
6778
2,121,799
—
—
Northern Europe
247,311
497
39,692
20
39,692
103
39,692
26,587
54,135
Southern Europe
8,875,054
47,738
4,855,172
2236
4,855,172
8752
4,855,172
1,120,143
6,560,893
Western Europe
2,366,777
10,218
1,003,060
698
1,003,060
967
1,003,060
1,199,130
1,862,640
Oceania
4,301,278
29,370
2,023,333
2372
2,023,333
8222
2,023,333
40
1367
Australia and New Zealand
3,977,140
29,052
1,992,243
2360
1,992,243
8166
1,992,243
—
—
Melanesia
23,173
204
23,173
8
—
36
23,173
40
1367
Micronesia
*****
14
1228
1
1228
3
1228
—
—
Polynesia
31,494
100
6689
3
6689
17
6689
—
—
Table 1.
Worldwide goat production (meat, milk, fat, skin, etc.) according to FAO, 2020.
Several factors have contributed to the differences among breeds. These differences are classified according to suitability to different purposes, breeding systems and adaptability to many environments [20]. Above 600 breeds of goat have been developed worldwide. These breeds differ from each other in their phenotypic characteristics and were adapted to different climatic conditions and extreme environments [20]. After domestication, goats were spread globally very quickly through commercial trades and human migrations [21, 22]. When comparing goats with other species, such as cattle, sheep and pigs, they have undergone an increase (+34%) in population since the year 2000 larger than cattle (+14%), sheep (+14%) and pigs (+15%) [20]. Goat production is one of the key elements contributing to the economy of farmers living in arid and semi-arid regions [23]. They play a necessary role in the livelihood of a large proportion of small and marginal holders. Moreover, goats are easy to manage and house, so that they can be raised by children, women and small families. Simply goats provide a reliable source of food for billions of people [10, 20].
Furthermore, the genetic diversity of goat is indispensable to meat, milk and hair current production needs under various environments, allowing sustained genetic improvement and facilitating rapid adaptation to changing breeding objectives [24]. Goats show reasonable production performance and reproductive behaviour. However, it is important to improve the productive and reproductive efficiency of goats to become more competitive, with other species [25, 26]. In many countries, goat breeds have been selected for special production traits, for example meat (Boer and Landrace), milk (Alpine and Saanen) and fibre (Cashmere and Angora) [20].
3.1.1 Scientific classification (Taxonomic Position) for goat
Classification
Scientific name
Common terminology
Kingdom
Animalia
Animal
Phylum
Chordata
Vertebrates
Class
Mammalia
Suckle Young
Order
Ungulata
Hoofed Mammals
Sup order
Artiodactyla
Even-Toed Ungulata
Section
Pecora
Typical Ruminants
Family
Bovidae
Hollow Horned Ruminants
Subfamily
Caprinae
Sheep And Goat
Genus
Capra
Goat
Species
C. aegagrus
Domesticated Goat
Subspecies
C. a. hircus
3.2 The history and immigration of goats following domestication
Historically, goats accompanied people in migrations and dispersing across the globe. They migrated to Europe and arrived at the far west and north edges of the continent about 5000 years before the present (YBP) [27]. Expansion eastwards to Asia and southwards to Africa occurred at the same time [28]. Goats were present in North Africa around 6000–7000 YBP [29], and in Ethiopia and the Sahara around 5000 YBP [30, 31]. The goats arrived in South-Saharan Africa, around 2000 YBP. In Asia, the evidence referred that, the goats were present in most areas of China around 4500 YBP [32] and moved further east and south thousands of years later. The goats arrived in Oceania and the Americas approximately during the fifteenth and eighteenth centuries jointly with European migrations to Americans [33].
There were waves of early migrations out of the domestication centre (DC) or Fertile Crescent (a crescent-form area of popular fertile land that is located in the middle east, extends from the Persian Gulf to the eastern Mediterranean through the Valleys of Tigris and Euphrates rivers) (Figure 1). This area was the centre of the Neolithic developing of agriculture since 7000 years BC, and the cradle of the Babylonian, Sumerian and Assyrian civilizations in the area that covers Northwest Iran and East Anatolia, ca; 10,500: 9900 (YBP) [16, 34], where, early domesticated goat followed the spread of farming and agriculture by radiating from the Fertile Crescent to Asia, Africa and Europe. Logically, the breeds from the regions near DC are expected to have retained partial ancestral diversity; thus, the Turkish goat breeds and Iranian goat breeds are similar to the wild ancestor (Bezoar breed) [35]. Moreover, one thousand generations ago, those original populations had larger similarities than recent breeds.
After a long time of so-known soft selection around two-hundred years ago, the case changed dramatically with the appearance of the concept of the breed [36]. The selection increased intensively in local breeds, followed by reproductive breeding between populations, thus, standardization of trait performance, all of these led to the fragmentation of the Initial Gene Pools (IGPo) [37]. A long period later, the pressure of selection has increased again through the utilization of artificial insemination (ArI), resulting in limited artificial populations with decreased effectiveness of population size, high trait performance and profound new phenotypic characteristics, for example trait-driven breeding for meat, dairy and cashmere [38]. As indicated by the diversity of ancient DNA (aDNA), the distinct Neolithic goat breeds from Southwest Asia owned a remarkable genetic structure [35]. These breeds characterized the populations in different areas surrounding DC, with early domestic goats from the east, west and southwest sides of the Fertile Crescent. The genomic analysis confirmed the relationship between those breeds and the populations from Asia, Africa and Europe [39].
The migration waves included sources of distinct breeds from that left their traces in the partitioning of diversity between countries and continents. Colli et al. [39] reported that regional gene pools were further promoted through the high levels of gene flow that characterize the breeds in large regions within Europe and Africa. In Europe, goat breeds are partitioned locally by regions corresponding to the central Mediterranean, eastern Mediterranean and eastern Alps together with continental Ireland and France and North Europe [35]. In Africa, population clusters correspond to East, West, Southeast, Northeast Africa and Madagascar. The geographical distribution of the African gene pools overlaps with those of the populations that share similar morphological characteristics; among them, African dwarf populations in central-Africa have short-eared Trypanotolerant goats, Northeast African populations have lop-eared goats, Small East African populations have short-eared Trypanotolerant goats scattered throughout the southeast and the far south has lop-eared goats [39]. Subsequently, the trading and migration led to an increase in populations and breed exchanges and movements, generating a reduction of genetic partitioning and resulting admixture [39].
A recent study by Colli et al. [35] confirmed that South American goat breeds are occupying an intermediate position between breeds from South Europe (Spain) and Northwest Africa. On the other hand, there is a rapprochement between some breeds from Australia, namely Cashmere and Rangeland and Turkish breeds, whereas the Pakistani breeds were separated from the rest of the populations of West Asia. In Africa, there are many sub-groups corresponded to East Central, Southeast, Northwest Africa and the Canary Islands. In Europe, clusters of populations consistent with North Europe (Norway, Netherlands, Iceland and Finland), South Europe (Central, South Italy and Corsica) and Western Europe (France, Ireland, Sardinia, Alpine breeds and Spain) were revealed. Romania Balkan breed had an intermediate position between West Asian populations and South European (Central Italy). Also, several Pakistani and European breeds were individually assigned to distinct clusters. There is a strong introgression originally in Africa with South America, Spain and Southern Italy breeds. However, previous genetic investigations uncovered the variety of sheep breeds worldwide revealing a low degree of differentiation of 2.98% among varieties between continents and sharing high levels of haplotypes [40], while cattle breeds (Bos taurus, Bos indicus and Bos javanicus) possessed 13% of all variation, but were 3.2% for taurine cattle, between African and European breeds [38, 41].
Three large-scale studies, two of them depended on microsatellite markers, and the third was based on the high-density SNPs [20] confirmed the occurrence of many regional gene pools, jointly with a clinical reduction in variability from DC in Southwest Asia towards northern Europe, Indonesia and China [42, 43].
Another investigation utilizing the mitochondrial DNA technique (mt-DNA) [44] confirmed that the high frequency (> 90%) and worldwide distribution of haplogroup (A), associated with geographical differentiation between continents, was uncovered by chromosome (Y) haplotype analyses [28, 45]. Furthermore, the widely distributed haplotypes (Y1B) and (Y1A and 2A) have been confirmed in Near East, North Africa and Europe, haplotype (Y2C) in Turkey and haplotype (Y2B) in Asia [46]. A recent investigation utilizing aDNA data succeeded to structure strongly, the nuclear molecular and the mitochondrial variation of Neolithic goat flocks [39]. This presented direct support to the hypothesis that several wild origins existed for early domesticated goat populations as was already found from mtDNA data tests [22, 44] and further refers to that recruitment from different local Bezoar breeds was extensive [47].
3.2.1 Distribution of goats to the different regions of the world
Since domestication, goats have established a large geographic range because of their adaptability to hard climatic conditions and poor diets [48]. There are more than 1000 goat breeds have been surveyed in the world in 2012 [49]. In 2020, the Asia-Pacific region possesses the greatest share of the goat population followed by the African region (Table 1). Both regions harbour more than 90% of the world’s population [50], followed by Europe, the Americas and Oceania [9, 51]. Worth mentioning, the main hotspots of global goat diversity and differentiation are in Africa, Asia and some European countries [9, 19]. Skapetas and Bampidis [51] confirmed that about 95 per cent of the goat breeds throughout the world are found in developing countries, especially in Asia and Africa. Asia holds first place with 579,347,344 heads of different goat breeds (Table 1), which forms 59 per cent of the world breeds. Asian countries contribute a considerable amount of goat products to the world economy. Per cent contribution of the goat products produced in Asia to the world production is 70.7% for meat, 58.3% for milk and 76.5% for skin. Goat production is considered to be very important for its contribution to the development of rural area in many countries. They have performed agricultural, economic, cultural and even religious roles and form important meat, fibre and milk resource [52, 53].
Goat breeds are globally distributed to every ecological area, though concentrated in the tropical developing countries and dry zones [54, 55]. Diversity in the climate and geography jointly with factors of traditions and history led to the development of a large variety of goat populations, which were developed later to the modern goat breeds [19].
The genetic structure of small ruminants especially goats reflects their domestication formation into different breeds [15]. The history of genetics for goat and sheep has been investigated utilizing three major sources of variation at the level of genomics: mitochondrial genome, Y chromosome and autosomes. The mitochondrial genome for goats has proven highly informative for studies into domestication, with results of haplogroups for several breeds [56, 57]. Recent investigations have tested collections of small ruminants from northern, southern Europe, and the Middle East, where, the facilitated tests of genetic partitioning through the globe. Interestingly, the breeds of southern European displayed much genetic differentiation compared with those of northern European [58, 59].
3.2.2 Modern domestic goat
Man domesticated goats along with many other species of animals since ancient centuries. The newest genetic analysis (GA) confirmed that there is archaeological evidence that the wild Bezoar ibex Capra Aegagrus of the Mountains of Zagros is likely the origin of almost all modern domestic goats today [60]. However, the morphological and behavioural characteristics of modern domestic goats have changed when compared with the progenitor of wild Bezoar goats [61]. The modern goat exhibit a more docile demeanour, variation in coat colour, reduction in body size and the ability to adapt after domestication and breed formation, which have left detectable selection signatures inside the genomes [62, 63].
3.2.3 Natural selection (NS) and artificial selection (AS) in goat
Natural selection (NS) has a necessary role in selecting the species that have high adaptability to changes in environmental conditions. Side by side, both NS and artificial selection (AS) have been applied widely to many livestock species to achieve more target phenotype traits [64]. Goats have been selected for domestication, since 10,000 years ago [22, 61]. The process of selection resulted in the differentiation of breeds that are characterized for the production of meat, milk or fibre, or multi-purpose breeds in many different regions of the world [64].
The strategies of NS and AS imposed pressure on a region of genomes that control some traits such as meat, milk and fibre, in addition to many important characteristics, such as reproduction, adaptation to extreme environments, behaviour, body conformation (BCF), resistance to parasites and diseases [64].
3.3 The vital role played by goat
The domestication and adaptation of wild goat to different environments and subsequent intensive trait-driven selection, inbreeding and crossing have led to intermediate breeds and phenotypic purification for the high-quality production of meat, dairy, cashmere,… etc. [65].
3.3.1 The meat of goat
Total goat meat production reached 6,142,140 tonnes in 2020 throughout the world. Based on a comparison of more than one hundred-sixty countries in 2020, China ranked the highest in the production of goat meat with 4,825,000 tonnes followed by India and Australia. On the other end of the scale was Singapore with 32 tonnes, Seychelles with 22 tonnes and Suriname with 20 tonnes. This total is 1.51% more than that in the last year and 15.2% more than ten years ago. Historically, total goat meat production reached an all-time high of 6,142,140 tonnes in 2020 (Table 1) and an all-time low of 6,032,000 tonnes in 1961. The average annual growth amounted to 1.69% increased since the 1960s. In 2020, goat meat was the world’s 381st most traded product, with a total trade of 7,420,000,000$. Between 2019 and 2020 the exports of sheep and goat meat decreased by −7.21%, from 7,990,000,000$ to 7,420,000,000$. Trade in goat meat represents 0.044% of total world trade.
Goat meat is characterized as lean red meat with favourable nutritional elements [66, 67]. It has a coarser texture, somewhat darker colour and distinctively different flavour and aroma than mutton [68, 69]. The smell and taste of goat meat are similar to the meat of springer sheep lambs. The results of sensory investigations marked goat meat to be different from mutton but absolutely not inferior to lamb [67]. Goat meat tends to be less juicy than lamb predominantly due to its reduced fat content [70]. Its special flavour is related to the presence of 4-methylnonanoic acid and 4-methyloctanoic acid [71, 72]. Also, branched-chain fatty acids (FA) may contribute to the typical goat meat flavour [73, 74].
3.3.2 The milk of goat
Over one billion goats live worldwide and due to expanding demand for milk, goats raised primarily for milk production are increasing in number. The total number of dairy goat reached 220,921,370 heads, while the total goat milk production reached 20,629,610 tonnes in 2020 throughout the world [75]. Most dairy goats are produced in Asia, 114,335,466 heads produced 12,219,732 tonnes of goat milk (Table 1). A global picture of the dairy goat sector sheds light on the lessons learned in building successful modern dairy goat industries. In Europe, especially in France, goat milk is the most organized market. Goats are primarily raised for milk production, mainly for industrial cheese making, but also for producing traditional cheeses on farms. Because of rising consumer demand, strong prices and climate change, there is an emerging market for goat milk in countries with no goat milk traditions, such as China, United States and New Zealand [76, 77].
The milk production of goats presents about 2% of the total annual supply of milk worldwide. This milk has very small well-emulsified fat globules that make it does not need to homogenization. The butter from the goat milk is white because of converting the yellow beta-carotene to the colourless vitamin A [78]. The goat milk composition includes water (88.9 g), protein (3.1 g), fat (3.5 g), carbohydrates (4.4 g), sugars ‘lactose’ (4.1 g), cholesterol (10 mg) calcium (100 IU), saturated fatty acids (2.3 g), monounsaturated FA (0.8 g) and polyunsaturated fatty acids (0.1 g) per 100 grams (g) [78, 79].
3.3.3 The reproductive traits of goat
3.3.3.1 Female fertility
It has been known since domestication that goats are seasonally polyestrous which means they reproduce naturally at certain times of the year. There are several factors that affect this characteristic, including daylight hours (photoperiod), altitude and nutrition [80].
The breeding season of goats will be shorter at latitudes farther from the equator. The availability of nutrients and the environment will determine the presence of oestrus in latitudes closer to the equator. Oestrus cycles in goats last on average for 16 days, with a high frequency of short cycles occurring during reproductive seasons and in young animals [81]. Oestrus lasts on average 36 hours but can vary between 24 and 48 hours depending on factors such as age, season, breed and presence of a male [82].
Understanding goat reproduction is crucial to increasing productivity, which is largely a function of pregnancy rate, the number of offspring born and weaned and the frequency of kids reproducing. Goat reproductive management produces a high level of fertility (90% and more) and optimum litter size (twins and triple) with a high rate of survival to weaning. All of the above processes will help producers to manage their herd more efficiently and breed their does to produce kids that will fit a specific market niche to command a maximum price. Worth mentioning, the major constraints to the reproductive management of goats are lack of data on the reproductive performance, especially of domestic goat breeds and the seasonal nature of breeding [82].
3.3.3.2 Male fertility (Semen production and quality)
There is a seasonal effect on male fertility (semen production and quality) in goat. Significant seasonal effects were reported in the Murciano-granadina, Alpine, Saanen and Damascus, Payoya, Damascus, Rayini, Zairi, Jakhrana, Blanca Andaluza, Anglo Nubian, Brazilian, Spanish breeds such as ‘Payoya goat’ and Peshawar dairy goats [82]. However, semen quality for goat has a high-quality level in different seasons probably due to the high reproductive ability of males [83].
3.3.4 The fibre (cashmere/ mohair/ hair/ cashgora) of goat
There are many inhospitable and marginal agricultural areas in the world where goats are kept and fibre is an important product in those areas. Cashmere, mohair and hair are three notable goat fibre products. In addition, a hybrid type called cashgora has characteristics between cashmere and mohair. The quality and yield of mohair (produced by Angora goats) are influenced by nutrition and are better suited to more staple environments that do not experience dramatic fluctuations in food availability. Cashmere growth, on the other hand, is remarkably insensitive to the nutritional influence, and may successfully be produced in harsh, continental climates. Goat hair is used, particularly in arid areas, for the manufacture of coarse cloth, tent fabrics and ropes. Goats are able to utilize vegetation dominated by woody species, such as brush range or veld, which may be unpalatable to other grazing livestock. Their diet selection behaviour, when present at an appropriate stocking rate, can be used to achieve vegetation management objectives, for example sustainable brush or veld management, weed control or pasture improvement. Cashmere goats are produced primarily in highly extensive systems and by subsistence-level pastoralists and nomads. The major producing countries are China, Mongolia, Iran and Afghanistan. Systems have been developed since 1970 for cashmere production in Australia, New Zealand, United Kingdom and United States which together represent some 9% of total world production. Primary producers of mohair are South Africa, United States, Turkey, Lesotho and Argentina. Smaller numbers of mohair goats are kept in Australia, New Zealand, France, Armenia, Belarus, Kazakhstan, Kyrgyzstan, Moldova, Russia, Tajikistan, Turkmenistan, Uzbekistan, Denmark, Spain and United Kingdom [77, 84].
Most goat breeds possess insulating hair covering the skin, a desirable fibre for the textile industry named cashmere or pashmina (a Persian word that means fine wool) [84]. The cashmere goat breed produces a large quantity of high-quality hair which is one of the most expensive fibres commercially produced [84]. The fibres of goats and sheep were used in many desert areas around the world to make houses built from fibres, especially nomadic caravans (Figures 2 and 3).
Figure 2.
The houses of goat’s fibre in many desert areas in the middle east including (‘El-Hammam city, Matrouh, Egypt’, ‘Moroccan desert, Morocco’, ‘Zulfi City, Saudi Arabia’) (Developed by the authors).
Figure 3.
The various types of fibres/wool are most commonly used for applied purposes (Developed by the authors).
3.3.5 Using goat for land clearing
Goats have been used to clear vegetative residues for many centuries. They have been described as ‘biological control agents’ and ‘eating machines’ [85, 86]. In North America, goats were used in a conservation grazing programme (1991) to clear dry bushes from California hillsides to avoid being endangered by wildfires. Since goat herds are being hired by public and private agencies to perform conservation grazing such practice has become popular in the Pacific Northwest. Goats succeeded to remove invasive plant species that cannot be easily removed by people, for example thorned blackberry vines and poisonous oak [86, 87, 88].
3.3.6 Using goat for medical training
Because goat physiology and general anatomy are not too different from that of humans, many institutes use goats for training combat medics in some countries. In the United States, goats have become the main species used for this purpose after the Pentagon phased-out using dogs in the 1980s [89]. Moreover, modern mannequins are utilized in the training of medical scholars on simulating the behaviour of the body. The trainees feel that when goats exercise they provide a sense of urgency that only true life trauma can provide [90].
3.4 Biodiversity of Goat Genetic Resources (GGR)
In view of the declining diversity in animals, awareness has arisen for the study of variation among and within goat genetic resources (GGR). In livestock, animal genetic resource diversity is expressed among and within breeds. Such diversity is of great importance in planning and implementing genetic improvement programmes [1, 2]. Genetic variation cannot be quantified only by studying morphology but also by using molecular techniques which makes it more reliable. These techniques are now widely used to study the biodiversity between populations. Variation is the substrates that natural or artificial selection can act on, therefore, genetic variability and then biodiversity is fundamental to the long-term survival of natural or domesticated populations [3].
Molecular genetics can be used to enhance the understanding of how genetic variation is portioned within and between breeds of livestock and this can play a necessary role in animal breeding programmes and genetic improvement strategies [2, 4]. A breeding goal, especially in small populations, should also sustain genetic variability which is important for selection and breeding strategies. At the molecular level, clarification of the population structure can be achieved by the highly variable loci that provide a large amount of information on individual genotypes [5].
According to one measurement of biodiversity, the goat has the highest degree (0.9) in comparison with some livestock species, that is 0.8 for cattle and 0.5 for buffalo [6]. This index of biodiversity depends on the number of breeds per million of a population of the species. Most of the breeds in developing countries are named ‘local’ because they are not characterized due to the lack of resources needed for characterization or measuring biodiversity [7]. For many years, breeds have been studied according to their phenotype as an indicator of biodiversity between and within populations. Several studies used phenotypic expressions such as blood protein polymorphism, isozyme variability and blood plasma to assess genetic variation, genetic distances, heterozygosity and genetic structure. This brought in some help for animal breeders to perform genetic improvement programmes and selection [8].
3.4.1 The importance of studying genetic diversity in goat
Goats can withstand hard conditions and survive primarily through scavenging for nourishment, while requiring small investment for maintenance. In addition, in the case of investment capital availability, goats are able to yield handsome returns. Thus, goats are present in a wide variety of production systems under extreme environments. Also, limited formal crossbreeding and few cosmopolitan goat breeds are available compared to other species. For these reasons the best ways for studying the adaptation and genetic diversity of goats are similar [10, 35].
3.5 Modern versus old strategies to enhance goat genetic performance
3.5.1 Old strategies to enhance goat genetic performance
Since domestication, farmers and breeders have been attempting to define target traits which may pass to the next generations and depend on the information on pedigree or on measuring of phenotype in order to make decisions on small ruminants to be kept or mated in the farms. These decisions have been important and effective to make necessary changes in the characteristics of the animals, making these species more economically efficient [13, 91].
The selection programmes which depended on the classic approach (classic breeding/ phenotype characteristics only) have been very useful and successful in their old-time, but they face many limitations and known defects because of the use of phenotypes only in selection: 1) the high cost and extra time to make records on phenotypic traits value. 2) There are many target traits that appear on female-only such as milk yield, 3) some traits take a long time to measure such as longevity. 4) Other traits need the slaughtering of individuals such as meat quality. 5) Resistance traits require the animal to get sick when exposed to the disease in order to measure disease resistance [92, 93, 94, 95]. All these and many other obstacles such as phenotyping constraints limit genetic progress.
On the other side, there is often little focus on breeding goats by most development organizations. Smallholder farmers’ indiscriminate breeding practices often result in negative genetic effects due to the lack of continuous, structural and established genetic improvement programmes. The result is decreased genetic merit of goats leading to a decline in productivity. This results in an inadequate availability of improved breeds for multiplication. Therefore, a systematic scientific approach for breed improvement aiming to increase the productivity of goats through genetic improvement is indispensable for increasing the impact [96, 97].
The most important criteria that were utilized to identify some individuals from males or females to be utilized for breeding and production are the assessment of their breeding value (BV) to focus on target traits in the next step. The BV is defined as all of the additive effects in all loci that contribute to the QTL deviating from the mean of the population [98].
In the last few decades, genetic techniques and sophisticated statistical analysis have allowed breeders to obtain estimates for the genetic values (GV) of their herds, depending on pedigree information and phenotype [99, 100], utilizing the mixed model procedures, to obtain the Best-Linear Unbiased Prediction (BLUP) of Breeding Values (BVs). This way is globally utilized for the evaluation of GV in all commercial livestock species industries. In several investigations, using estimated breeding values (EBVs) to arrange matings and making decisions has contributed significantly to genetic progress and consequently to the profitability of goat and sheep raising [101, 102].
The population mean is what breeders aim to improve. The second factor is that progeny receive 50% of their alleles from mother and 50% from father, the records of phenotypes, especially for target traits and traits that are genetically correlated with them have been used as the only source of information to estimate BV for selected candidates. To do this, the statistical BLUP methodology of the mixed linear model can be successfully utilized. That method depends on the information from phenotypic records, which were not only collected from the individual itself but also from its relatives to maximize the accuracy of the resulting EBV [103, 104] (Figure 4). Selection theory and statistical models utilized in breeding programmes depend on the known infinitesimal model of quantitative genetics [98]. This model assumes that the traits are related to an infinite number of genes that have additive and minor effects [92].
Figure 4.
Estimation of breeding value (EBV) utilizing traditional methods (traditional selection) (Developed by the authors).
However, animal breeders built up a big deal of experience, investigating opportunities and making conclusions to obtain early measurements for target traits on selection candidates, which may be utilized to improve the accuracy of EBV estimation at an early age. One of the important early applications was utilizing the success in blood groups as a marker for disease resistance in chicken selection [93].
A substantial rate of genetic progress has been achieved for many quantitative traits by selection in livestock populations including goat breeds utilizing EBV derived from genotypes and phenotypes [105, 106]. This approach did not require information on genes, loci or signatures of selection (SS) that influence the target traits. Meanwhile, the application of molecular genetics tools for quantitative trait selection processes in the past decade was too expensive.
On the other side, the availability of new molecular techniques such as high-density SNP panels/chips and Whole-Genome Sequencing (WGS), and thus the genomic selection (GS) approach may have a good impact on livestock genetic improvement including goats. Additionally, the methods of genomic evaluation are able to improve substantially the accuracy and effectiveness of Genome Enhanced Breeding Values (GEBV) estimation in goats and thus accelerate the response for selection. The accuracy of the methodology that utilizes genomic prediction such as genomic best linear unbiased prediction (GBLUP) and best linear unbiased prediction-SNP (BLUP-SNP) is limited when the target family or population is small [107, 108], though the application of GBLUP increased accuracy of EBV than pedigree-based BLUP [109].
3.5.2 Modern strategies to enhance goat genetic performance
The development of next-generation sequencing (NGS)/ WGS since 2007 has allowed for obtaining the genome sequencing [109] of goat [110], sheep [111], cows [112] and buffaloes [113]. Thus, it became feasible to develop high-density SNP chips, for example the (Goat SNP K50 Bead-Chip), which contains 53,347 SNPs [114] and the (Ovine 600 K SNP chip), which contains 606,006 SNPs. The availability of such high technologies has allowed the utilization of genome-wide results for animal breeding improvement plans [40, 115].
In this regard, molecular tools had revolutionized animal species breeding around the world through increasing genetic gain compared to old or traditional methods. The improvement was achieved for many livestock products such as beef and milk from cattle utilizing both traditional and high technology methods; however, there is a lack of knowledge concerning small ruminants [9, 95, 116]. Additionally, getting Genome Enhanced Breeding Values (GEBV) at early age to apply selection on young individuals prior to getting the data of extensive progeny had crucial effects on breeding programmes in many species [117].
GS had revolutionized animal species breeding including goats, around the globe. Therefore, this work aimed to discuss the evolution and vital role of advanced molecular tools to develop livestock populations, especially goat breeds worldwide. Also, to scan topics that are necessary for the successful application of genomic results and review the candidate genes, SS and quantitative trait loci (QTL) influence important traits in livestock populations worldwide and present the role of GS which depends on knowledge of the behaviour of the phenotypic traits along with information about the genome, also taking the spotlight on the relationship between economic traits and genetic differentiation in goat.
3.5.2.1 Assess the genetic resources of indigenous goat populations
Several indigenous goat populations are threatened with extinction because of taking their places by cosmopolitan breeds; however, these breeds might represent unique valuable resources of genotyping. Recent studies estimated that 18% of indigenous goat breeds worldwide are threatened or extinct. Thus, the characterization and determination of genetic differences between and within indigenous goat populations is a potential tool to help the rapid improvement of economically important traits. Side by side, the characterization and determination of WGS variation in livestock species are possible by utilizing new technologies of sequencing [62].
Therefore, it is important to assess the genetic resources of indigenous goat populations, and, thus, manage those breeds sustainably in addition to present zoo-technical ways that take into consideration the preservation of goat genetic resources. This is probably useful at least under the current changes in the global environment [118]. To accurately determine and characterize resources of goat genetics, it is important to access the variety of data in WGS. This would help the identification of genes and mutations of alleles related to different environmental conditions especially, those potentially representing a necessary adaptive role [119].
3.5.2.2 The concept of modern genetic improvement
The genetic improvement in livestock domesticated populations utilized for multi-propose production includes mainly the selection of females and males that are mated to produce next generation that shows performance better than the average of the current parents. The genetic improvement that uses the fundamental foundations of old breeding strategies, such as BV side by side with modern genetic techniques/tools is a promising approach for improvement [92].
3.5.2.3 Quantitative traits
By using special statistical methods, researchers and breeders can locate chromosomal regions that contain many genes contributing to variation in a quantitative trait of interest in a population by starting with its phenotype (phenotypic) and moving forward to its genotype (genotypic). Since phenotypic performances partially reflect the genetic values of individuals in the breeding process, many traits of interest in animal breeding show quantitative inheritance. QTLs and environment interactions define a quantitative trait’s genetic variation. On the other hand, it is common for quantitative traits to be controlled by several genes and in some cases, hundreds, even thousands of genes, all in conjunction with the environment. Two of the most popular methods for mapping QTLs and exploitation of molecular markers in animal breeding are linkage analysis and association mapping. A subset of markers is found associated with one or more QTLs that regulate the expression of complex traits. There is currently an optimistic assessment of the possibilities of marker-assisted selection (MAS) based on the identification of QTLs that explain a significant proportion of phenotypic variance by QTL mapping, which may push strongly towards achieving the goal of genetic improvement [98].
3.6 Molecular (Modern) tools for measuring biodiversity
The development of molecular techniques in the last five decades facilitated studying biodiversity in goat genetics. There are three generations of molecular tools: 1) the first generation includes random amplification of polymorphic DNA (RAPD), amplified fragment length polymorphism (AFLP), restriction fragment length polymorphism (RFLP), single-strand conformation polymorphism (SSCP) and microsatellites (MST) / simple sequence repeats (SSRs). 2) Second-generation includes mitochondrial DNA (mtDNA), DNA microarray (biochip or DNA chip), the low and high-density of SNP chips and WGS and 3) third-generation single-molecule real-time (SMRT) [82, 120].
Worth mentioning, the new concepts in goat genetics and the breeding sector highlight the necessity and seriousness of characterizing and determining novel polymorphisms, which are associated with important traits. Detailed studies have been done and others are still underway to re-design genomic maps to understand the effect of allelic variants on the analysis of performing linkage and quantitative phenotypes to accelerate genetic improvement utilizing different molecular tools in goat breeding programmes [9, 95, 121].
In order to identify strategies of conservation and monitor genetic diversity, many investigations on molecular markers such as SNPs panels [122, 123] were carried out. Nowadays, the availability of panels for SNPs eased the testing of animal genomic diversity [124, 125]. These panels have replaced microsatellites in detecting genetic diversity and parentage assignment (PA) in many species [126]. The SNPs approach is one of the most typically utilized classes of a genetic marker. Through the second phase of the Hap-Map project, 4.4 million SNPs have been genotyped in the human genome [127]. In dogs, approximately 2.5 million SNPs have been found [128], in chickens, approximately 2.8 million [129], in mice, approximately 8.2 million [130], in cows, approximately 60,000 [131] and in goats >41 million [132]. In humans, genome-wide association studies (GWAS) have used these markers to discover genomic regions or sequence variants associated with 40 complex diseases [133].
In different species, the developing of the NGS permitted the check of the sequencing of the goat [110, 134], sheep [111], cattle [135, 136], buffaloes [137], deers [138], chickens [139] and pigs [140]. The first SNP chip 50 k Bovine having more than 50,000 SNPs was available in 2011. It was built utilizing ten geographically and biologically different breeds. On the other side, the International Goat Genome Consortium (IGGC) for goat genome affairs was established in 2012 [141], with primary goals of improving and increasing the tools for extracting genomic information on goat genetics and supporting the international efforts to develop the 52 K SNP chip known as commercial Illumina [142], in 2013 [114] by combining genome-libraries and WGS from eight populations (breeds) from Asia and Europe [141]. The first assembly of the genome of a goat was released by Dong et al. [110]. The identification of 12 million SNPs allowed the design of 53 K SNP and 54 K SNP chips that are extensively utilized globally [114]. A 54 k cattle SNP chip was applied to characterize the genome of cattle and buffaloes [143]. More recently, there are 60 K SNP and 62 K SNP chips [144]. Nowadays, there are chips of 40,000 to 65,000 SNP are available for most livestock species, including cattle, buffalo, sheep, goat, horse, poultry and pigs, and in other species, these chips are still under development [92]. It may provide powerful means for the direct discovery and identification of traits associated with sequence variations underlying the molecular mechanisms of adaptation and domestication by using suitable statistical methods [145].
3.7 Outcomes of utilizing advanced molecular tools in goat breeding
3.7.1 The high-throughput SNP (SNP panels)
The recent investigations proposed a new method of selecting SNP even when utilizing a limited number of individuals or breeds [146, 147]. The AdaptMap project initiative has collated 53 K genotypes from more than 140 breeds from 17 countries, providing the international dataset for goats. The main goal was to select a panel of SNPs that has high performance for the parentage assessment and assignment for 91 populations. Interestingly, two approaches were utilized for this aim: the first one is an approach that depends on the detection of SNPs for proving the relationship between individuals, and the second is an approach that depends on selecting the SNPs that maximize the minor allele frequencies (MAF) in the largest possible number of breeds as represented by Talenti et al. [126] who selected a panel of SNPs suitable and readily applicable for PA and assessment for a large number of goat breeds around the globe. Therefore, these SNPs were validated on a large data set involving Alpine and French Saanen goat populations.
The Caprine-SNP50k Bead-Chip was used to investigate and compare several Swiss goat breeds, and SS were identified in the regions that affect variation in milk composition, growth and coat colour [148]. More recent advances such as Goat 50 K SNP Bead-Chip offered the opportunity to identify regions in genomics that have undergone selection. There are few investigations utilizing the arrays of SNP focused on local goat breeds such as Moroccan and Italian [19, 62]. Lashmar et al. [66] reported that the Goat SNP 50 Bead-Chip has a marked positive change at the molecular level for small ruminant species. Now, the commercial 50 K SNP chips have been available for about 6 years [114] and have already been utilized on the national, regional and continental levels to explore goat diversity [20, 149]. Similar advances in sheep [150, 151], cattle [152, 153], pigs [154, 155] and chickens [153] have defined genes related to undergoing positive selection and contributed to phenotypic variation.
Brito et al. [64] reported that 1151 individuals belonging to nine breeds genotyped by the 50 K Bead-chip were identified for many genes related to important economic traits. Others were identified and found related to many traits such as milk protein and somatic cell count score, FAM13A [156, 157], reproduction traits, MEF2BNB [158], CACNB2 [159] and CYP19A1 [64], the efficiency of feed conversion, KIAA1211 and VAV-3 [160], adult body mass, GPR-61 [161], conformation traits, RNF-157 [162], abdominal fat deposition, PRPSAP-1 [163], metabolism of liver fat, TM6SF-2 [164], the fatty acids in milk, CDH-12 [165], heat-tolerance, GNAI-3 [166] and ear morphogenesis WNT5A gene [167].
3.7.2 Scanning the Mitochondrial DNA (mtDNA) sequence
NGS was used to detect the mtDNA variants [168, 169]. The mt-DNA has represented the most informative genomic element to investigate diversity in all closely related livestock and individuals within many species [170], because it displays the maternal inheritance relatively rapid against evolution rate and without recombination [18, 57]. Also, mt-DNA is one of the most useful tools/ approaches in molecular phylogenetics and population genetics [171].
Practically, mt-DNA variation or mutations create differentiation in the efficiency of the oxidative phosphorylation pathway and consequently cellular energy production thus variation in the performance [172]. The molecular tools allowed to discover these variations in mt-DNA and built an association with the performance of different traits in many livestock species [173].
Concerning mt-DNA of goats, the phylogeny of domestic goats elucidates six credible maternal haplogroups of domestic goats worldwide, namely; A, B, C, D, F and G. However, the classification and ownership of some low-frequency mt-DNA haplogroups in several small geographic regions remain controversial. [174]. The probable origin of mt-DNA haplogroup (A) could be Eastern-Anatolia, while haplotypes A and C have been detected in ancient goat samples (from an early Neolithic site) in Southern France. Meanwhile, haplotypes B, D and G have been found in Eastern Anatolian and Northern Iranian bezoars. Also, there was domestication marked by haplogroup C in the easternmost [175].
mt-DNA is an extensively exploited tool for the evaluation of evolutionary relationships and genetic diversity in goats. In this regard, the popularity of mt-DNA for phylogeographic investigations has been attributed to its remarkable properties, such as maternal inheritance, high copy number, higher mutation rate without any recombination and the clock-like nature of its substitution rate in goats and other species [175]. Also, the Displacement-region (D-loop) in mt-DNA is characterized by highly polymorphic regions (Hypervariable-region I and II) and has been particularly informative in explaining the origins of several livestock species including goat [174, 176].
3.7.3 Identification of candidate genes related to important traits
A big number of candidate genes have been discovered and identified in different species utilizing molecular tools [177]. Numerous studies reported the discovery of hundreds of genes in different animal species since the release of the genome sequence reference (Table 2). There is a relationship between candidate genes and economic traits, such as reproduction, production traits and disease included dozens of genes [165, 183], and the effects of those candidates vary as follows: 1) several candidates have associations with the physiological and metabolism pathways, for example IGFBP-3 [116], GH [184], GHR [185], IGF-I [186], CAST, CAPN-1 [187], POU1F1 [188], LEP [189] and MSTN [190, 191] genes that are crucial for birth weight, weaning weight, growth traits, bone formation, muscle growth, body size and meat quality. 2) Other candidates have an association with fertility, infertility and reproduction, proliferation and sex-determination, for example MTNR-1A [192], FOXL-2 [193], AMEL [194, 195], SRY [193, 196] BMPR-1B, GDF-9 and BMP-15 [95] genes. 3) The third part of genes is candidates for milk composition and milk yield traits, such as the family of casein genes [197, 198, 199]. Worth mentioning, genes related to casein formation in milk protein are already utilized in breeding programmes [200]. 4) The fourth part is related to fibre (hair/cashmere) traits, such as FGF-5 [77], IGFBP-7, MC1R [201] and KAP [202] genes, which are vital to several characteristics of fibre, for example hair length (short, medium or long), hair colour (black, white or brown, …), hair colour pattern (plain, patchy or spotted), hair type (straight or curly) and hair appearance (dull or glossy). 5) Also, part of these genes is related to the immune system and disease resistance, as MHC-DRB3 and MHC-DQA2 genes [203], as well as Tmem-154 gene is related to resistance to (MAEDI-VISNA) [204], Prp gene is related to scrapie resistance in goat [204] and sheep [205], and Socs-2 gene is related to the susceptibility to mastitis [206]. Worth mentioning, Fec-L and Prp genes in worldwide sheep, and α-s1-casein gene in French goat are especially used to pre-select candidates for progeny testing [165].
Number of QTLs for many species based on animal QTL database updated to 2022.
Also, utilization of advanced molecular tools resulted in mapping many useful individual genes in small ruminants [207, 208], dairy and beef cattle [209, 210]. Also, veterinary tests utilized advanced molecular tools for the diagnosis of genetic diseases, such as Deficiency of Uridine Monophosphate Synthase (DUMPS), Complex Vertebral Malformation (CVM) and Bovine Leukocyte Adhesion Deficiency (BLAD) in cattle which are tested to find out if the seed stock bulls are either carriers or non-carriers of these autosomal recessive mutations [211, 212]. Small ruminants individual testing for Prp gene associated with scrapie have been identified by molecular tools [213, 214].
3.7.4 Detection of Quantitative Trait Loci (QTL)
In the above molecular tools, the gene related to the trait of concern could have been either a single gene with a large effect which is known as a major gene [95], such as genes affecting, skin-tone and coat colour ‘KIT gene’, polledness and double-muscling ‘MSTN gene’, or could be one of several genes associated with a quantitative trait known as minor gene, such as genes related to growth, milk yield and wool quality [116]. The loci that affect a quantitative trait are termed QTL or Economic-Trait-Loci (ETL). By another meaning, a QTL is hypothesized as a specific region on a chromosome containing several genes that make a significant contribution to the expression of a quantitative/complex trait. In populations that had effective improvement programmes for many generations, MAS most likely will be for QTL rather than for major genes, since major genes with large favourable effects are likely to have been fixed in this population already [215]. QTL analysis started in the nineties and now a number of QTLs for many animal species are available (Table 2) followed by many sequences as described in Table 3, with a help of different molecular tools.
The first sequenced genomes for many animal species.
It was estimated that using markers linked to QTLs in goat, dairy and other livestock breeding programmes could increase animal response by up to 30% [234]. There are probable benefits from MAS and the scale of such potential benefits will depend on the QTL effect, the strength of the linkage between the marker and the QTL and the rates of possible changes by conventional means [235]. Utilization of modern molecular tools led to the discovery of thousands of QTLs associated with economic traits in livestock species including goats.
3.7.5 Obtaining whole-genome sequencing (WGS)
The progress in sequencing technology has made new perspectives towards the magnitude of the genome analysis; sequencing time and costs have decreased dramatically and now WGS can be obtained easily [119] (Table 3 and Figures 5 and 6). Obtaining the genome sequencing of goat and other domestic animals is extremely beneficial to detect the candidate genes, MAS, SS and QTLs, and their association with reproductive and production traits, animal health and welfare, also, is very beneficial to most animal production practices, besides understanding the genetic basis of the diseases, as well as GWAS applications [236, 237, 238].
Figure 5.
The steps of implementing the genomic selection (GS).
* DGV: Direct Genomic Values and * EBV: Estimating Breeding Values (Developed by the authors).
Figure 6.
The methodology of genomic selection in goat (Developed by the authors).
More recently, several whole-genomes have been investigated for many livestock species with huge data about the history of these species and their domestication. Also, GWAS has been investigated in most of the economic traits in farm animals, for example, cattle [239], sheep [240], goat [241], pigs [242], chickens [243] and rabbits [244], but others still underway (Table 4).
Summary of genome projects for several animal species.
In a recent investigation by Fu et al. [132] based on the results of modern molecular techniques genomes of 24 bezoars,46 wild ibexes, 82 ancient goats and 208 modern domestic goats (∼360 WGS) were used to produce a comprehensive genome variation database. That database hosts ∼41,440,000 SNPs, ∼5,140,000 indels, 6193 selected loci and 112 introgression regions which can be widely used in the future genetic comparison between different goat breeds worldwide.
3.7.5.1 The Genome-Wide Studies (GWS)
An essential goal in animal breeding is to select individuals that possess effective BVs for target traits and allow them to be parents of the new generations. The success of GWS in finding variation sequences linked to important complex traits led to increased interest in SNP genotyping approaches in animal species, with the main goal of detecting QTL, candidate genes and then GS [245].
The appearance of SNP genotyping in combination with new advanced statistical methods to analyse the available data on the prediction of BVs had benefitted the extensive application related to WGS and genomic studies in livestock species. The scientific community had to implement GS in many animal species [92, 246].
3.7.5.2 Genome-Wide Association Studies (GWAS)
A GWAS is an approach that includes rapidly scanning genetic markers (GM) across the whole sets of genomes of several individuals to find the genetic differentiation associated with a specific trait [247]. Once new associations are discovered, investigators can utilize this knowledge to create and develop better strategies to improve the trait in animal populations [247, 248]. Also, identifying associations between genetic markers and important economic traits will provide practical benefits for the goat industry, enabling genomic prediction of BV of individuals and facilitating the discovery of the underlying candidate genes and mutations [75].
3.7.6 Signatures of selection (SS)
The high-throughput SNP genotyping/ NGS contributed significantly and strongly to uncover the signatures of selection (SS) in different livestock populations [249, 250]. Where the unique patterns of genetic left behind in the genome under NS or AS is defined as SS also, known as the change, elimination or reduction of genetic variation in the regions of genomic, neighbouring the causative variants in response to the pressure of NS or AS, it is also very important for GS [20, 251]. These signatures are often important regions of the genome that have unique sequence variants. This access is necessary and relevant because it has the potential to elucidate the identities of mutations and genes associated with traits related to phenotypes but with no need to measure them [251]. The availability of high-throughput SNPs and genomic tools able to increase exploring and identifying SS and other genomic diversity resulting from adaptation to selective pressure and environment [20] are successfully used in circumscribed datasets. The detection of SS helps to elucidate the effectiveness and identify mutations and genes associated with economic traits in livestock species. In addition, they are important to find out the levels of polymorphism and genetic diversity in a population, as genetic differentiation in animal breeding represents the raw materials crucial for the implementation of GS [252].
3.8 Genomic selection (GS)
Applying the information of wide-genetic markers in animal breeding was originally suggested by Meuwissen et al. [253]. The traditional principles of MAS were involved in a relatively limited number of genetic markers (GM) [254]. Those markers were recognized from research results of the primary analyses of controlled tests [255] (Tables 3 and 4).
Nowadays, GS which is essentially a big-scale version of MAS involves a considerably big number of GM. The ‘effects and roles’ per each marker are simultaneously estimated during the process of GS. The number of MM involved in the genomic evaluations depends on the utilized procedure. GS assumes that all the trait’s genetic differences must be explained through markers. However, the polygenic effects involved in the model to account for genetic variation might be un-explainable by GM [256]. GS should ultimately lead to utilizing genotypes defined by the polymorphisms to select target phenotypes [257].
GS may play a substantial role in improving traits which achieved genetic gains by traditional methods. Some important traits are expensive or difficult to measure such as carcass traits and resistance to diseases, and others are measured on one-sex or at the end of the animal life such as milk composition and production, and carcass characteristics, respectively. Available modern highly technical approaches such as GS can overcome the constraints in genetic improvement [25].
The GS has the ability to promote the resilience of animal species, especially small ruminants such as goats, for increasing production, adaptation and resistance to deadly diseases [258]. This includes breeding to resistance to diseases, parasites, fly strike and facial eczema [259]. Moreover, GS has ethical benefits in reducing the number of individuals susceptible to diseases and those who might be suffering in the coming generations. Investigations are now underway to allow the prediction of genomics for examination of farms important and effective biological traits such as methane emissions and feed efficiency. The examination of the last two traits involves high costs and cannot be widely disseminated on farms; thus, likely GS might a promising alternative [259].
GS depends on genotypic, phenotypic and pedigree data which might open a window for adopting breeding programmes in farm animals to improve meat, milk and fibre/cashmere production and the traits that are difficult to handle using traditional ways such as reproduction, breeding seasonality, longevity, meat quality and carcass composition [260]. The feasibility of GS to deal with small ruminants has been evaluated in French dairy goats [261, 262], Australian mutton breed [263] and French dairy sheep [264].
The methods of GS has been successfully applied in breeding programmes of dairy cattle and succeeded to reduce the generation interval. In goats, though the generation interval is shorter than in cattle and buffaloes, it still should be reduced. This increases the intensity of selection of the genetic gains per year and consequently reduces costs and increases productivity [25].
GS includes testing SNPs and their high-density effects utilizing a model fitted simultaneously to each SNP and treating these effects as random variables. Many Bayesian models have been updated to implement statistical estimation utilizing the methodology of the Monte Carlo Markov Chain (MCMC) [265, 266].
3.8.1 The methodology of genomic selection (GS)
To implement GS in any animal population source requirements should be fulfilled: 1) large number of goats per genotype, 2) availability of information on phenotype specific to each genotype, 3) statistical methods suitable for accurate and effective genetic prediction, assuming that the breeding programme is optimal, to achieve that: a) an accurate system of genetic evaluation to relevant phenotypes, b) the breeding objective should be related to target traits, c) the scheme of breeding ensures long-term sustainable genetic gain [267].
In general, 1) the obtained data will be used as a reference to develop the new statistical models for estimating the effects of SNPs on the target traits. 2) The results are predictive equation to estimate GEBV [268]. 3) In the absence of accurate phenotypes, the genomic breeding value (GBV) of new individuals can be computed from the prediction eq. (PE), depending on the genotypes from the SNP arrays. 4) The accuracy of GEBV depends on the heritability of traits of interest and the size of the population [117, 269] (Figures 5 and 6).
3.8.2 Advantages of Genomic selection (GS)
There are many advantages for GS: 1) provides an easy way to improve hard traits such as sex-limited, low heritable and those measured late in the animal life, 2) applied early in the animal life, 3) increases the accuracy of selection, 4) ability to be extended can be to the traits that are recorded in the reference population, 5) increases the genetic gain through reducing generation interval, 6) not limited to a specific population, 7) possesses high intensity and 8) explains the differentiation at the DNA level more than MAS [268].
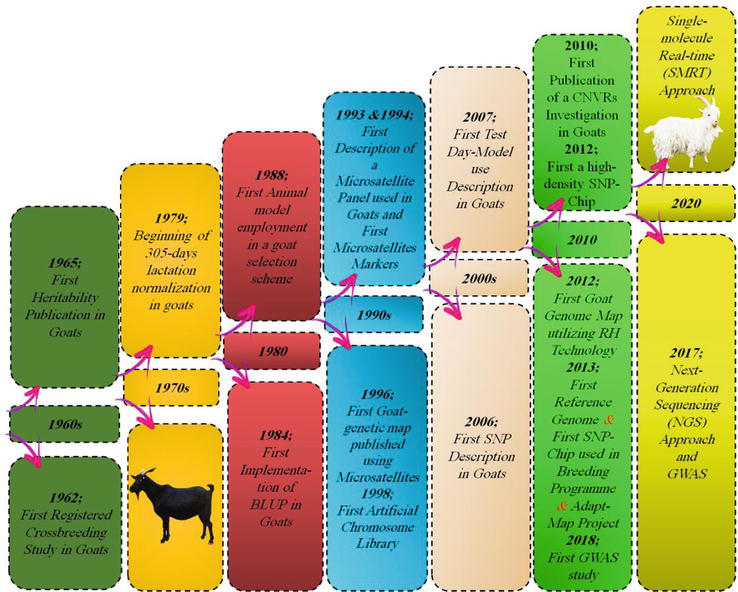
In the last six decades, a lot of genetic attempts have been made to improve the goat production sector, starting in the 1960s until now (Figure 7). In recent years, many studies have been analysing the genetics of goats, as one of the common domestic species adapted to extreme environments, and have phenotypic diversity. These studies have succeeded to identify the whole goat genome as well as some genes with key roles in domestication, adaptation to harsh environments or prominent economic traits [27, 110, 270].
Figure 7.
Timeline of goat genetic improvement (Developed by the authors).
Supakorn [106] and Saleh et al. [270] reported that more than 271 candidate genes have been identified and detected in goat breeds. This number will be doubled or tripled over the next few years with more focus on the association with economic traits [271, 272]. In addition, comprehensive studies have been done on the differentiation and polymorphism of several genes along with their roles in the control and management of economic traits. Moreover, some of these genes have either antagonistic or synergistic effects on the expression of phenotypic characteristics in nature. It is necessary to identify these candidate genes, along with their different impacts on economic and non-economic traits, which could help in goat breeding programmes. Also, it will be utilized as an aid in GS early in animal life [106, 272].
This chapter systematically introduces the basic information of the history, domestication and immigration of goats and modern versus old strategies to enhance goat genetic performance. It provides knowledge on the distribution of goats to the different regions of the world, the natural and artificial selection of goats, the production throughout the world, the vital role played by goats (fertility, milk, meat, fibres, land clearing, medical training, …) and the biodiversity of goat genetic resources, also, special characteristics of goats under different conditions. On the other hand, this chapter focused on modern strategies to enhance goat genetic performance, including different molecular tools, besides mentioning the outcomes of utilizing advanced molecular tools in goat breeding, as well as identification of candidate genes related to important economic traits, detection of SS and QTL, applied GWAS and the methodology of genomic selection.
In this chapter, the genetic investigations conducted on goats throughout the past three decades have been reviewed, along with a summary of the findings of those investigations in a focused manner with a discussion of the most significant QTL, candidate genomic regions and candidate genes that affect different body functions such as integrated course of adaptation, coat colour, skin sensitivity, body measurements, milk production, fibre production, meat production and quality, and goat diseases.
Thence, we believe this chapter is very crucial and will be a useful reference for readers and researchers to start answering the questions and solving the problems about goat production and industry. Also, we believe that this chapter opens a new window for researchers who are interested in the field of goat production development, especially genetic improvement, whether at the genome level or candidate genes. It also preserves the effort and time required to search dozens perhaps hundreds of investigations and studies related to this field which requires more investigations in the near future to cover every single gap in the previous studies.
3.Lawson L. Characterisation of diversity and evolution in European sheep breeds and their wild relatives. PhD thesis. University of East Anglia. 2000. Available from: http://worldcat.org/identities/nc-university%20of%20east%20anglia%20norwich%20gb.
4.Pearson WR, Lipman DJ. Improved tools for biological sequence comparison. Proceedings of the National Academy of Sciences. 1988;85(8):2444-2448
5.Balloux F, Lugon-Moulin N. The estimation of population differentiation with microsatellite markers. Molecular Ecology. 2002;11(2):155-165
6.Iniguez L. Characterization of Small Ruminant Breeds in West Asia and North Africa: International Center for Agricultural Research in the Dry Areas. Rome, Italy: Aleppo (Syria) eng; International Plant Genetic Resources Inst.; 2005
7.Galal S. Biodiversity in goats. Small Ruminant Research. 2005;60(1–2):75-81
8.Tapio M, Grigaliunaite I, Holm L-E, Jeppson S, Kantanen J, Miceikiene I, et al., editors. Mitochondrial differentiation in Northern European sheep. Proceedings of the 7th World Congress on Genetics Applied to Livestock Production (WCGALP); Montpellier, France: Management of genetic diversity; 2002
9.Ahmed A. Saleh AR, Nada N. Hassanine AM, Sharaby MA. Modern strategies to enhance goat genetic performance. LAP LAMBERT Academic Publishing; 2019. https://www.orellfuessli.ch/shop/home/artikeldetails/A1062043435. DOI; 10.6084/m9.figshare.11859945/ ISBN: 978-620-0-47020-1/ EAN: 9786200470201
10.Scherf BD, Pilling D. The Second Report on the State of the World’s Animal Genetic Resources for Food and Agriculture. 2015
11.Stanford K, Stitt J, Kellar J, McAllister T. Traceability in cattle and small ruminants in Canada. Revue Scientifique et Technique-Office International des Epizooties. 2001;20(2):510-522
12.Pirtle TR. History of the Dairy Industry. Mojonnier Bros. Co; 1926
13.Bruford MW, Ginja C, Hoffmann I, Joost S, Orozco-terWengel P, Alberto FJ, et al. Prospects and challenges for the conservation of farm animal genomic resources, 2015–2025. Frontiers in Genetics. 2015;6:314
14.Legge T. The beginning of caprine domestication in Southwest Asia. The origins and spread of agriculture and pastoralism in Eurasia. The University of Chicago Press on behalf of Wenner-Gren Foundation for Anthropological Research. 1996:238-62. https://www.jstor.org/stable/10.1086/659307
15.Kijas JW, Townley D, Dalrymple BP, Heaton MP, Maddox JF, McGrath A, et al. A genome wide survey of SNP variation reveals the genetic structure of sheep breeds. PLoS One. 2009;4(3):e4668
16.Naderi S, Rezaei H-R, Pompanon F, Blum MG, Negrini R, Naghash H-R, et al. The goat domestication process inferred from large-scale mitochondrial DNA analysis of wild and domestic individuals. Proceedings of the National Academy of Sciences. 2008;105(46):17659-17664
17.Saleh AA, Rashad AM, Hassanine NN, Sharaby MA, Zhao Y. Evaluation of morphological traits and physiological variables of several Chinese goat breeds and their crosses. Tropical Animal Health and Production. 2021;53(1):1-15
18.MacHugh DE, Bradley DG. Livestock genetic origins: Goats buck the trend. Proceedings of the National Academy of Sciences. 2001;98(10):5382-5384
19.Nicoloso L, Bomba L, Colli L, Negrini R, Milanesi M, Mazza R, et al. Genetic diversity of Italian goat breeds assessed with a medium-density SNP chip. Genetics, Selection, Evolution. 2015;47(1):62
20.Bertolini F, Servin B, Talenti A, Rochat E, Kim ES, Oget C, et al. Signatures of selection and environmental adaptation across the goat genome post-domestication. Genetics, Selection, Evolution. 2018;50(1):57
21.Clutton-Brock J. A natural history of domesticated mammals. Cambridge University Press; Nature; 1999. 238. Available from: https://books.google.com.eg/books/about/A_Natural_History_of_Domesticated_Mammal.html?id=cgL-EbbB8a0C&redir_esc=yClutton-Brock J. A Natural History of Domesticated Mammals. Cambridge University Press; 1999
22.Naderi S, Rezaei H-R, Pompanon F, Blum MG, Negrini R, Naghash H-R, et al. The goat domestication process inferred from large-scale mitochondrial DNA analysis of wild and domestic individuals. Proceedings of the National Academy of Sciences. 2008;2008:0804782105
23.Mohammadi A, Nassiry M, Mosafer J, Mohammadabadi M, Sulimova G. Distribution of BoLA-DRB3 allelic frequencies and identification of a new allele in the Iranian cattle breed Sistani (Bos indicus). Russian Journal of Genetics. 2009;45(2):198-202
24.Seilacher A, Bose PK, Pflüger F. Triploblastic animals more than 1 billion years ago: Trace fossil evidence from India. Science. 1998;282(5386):80-83
25.Brito LF, Jafarikia M, Grossi DA, Kijas JW, Porto-Neto LR, Ventura RV, et al. Characterization of linkage disequilibrium, consistency of gametic phase and admixture in Australian and Canadian goats. BMC Genetics. 2015;16(1):67
26.Saleh AA. Utilization of Molecular Markers to Detect some Genes and Mutations Affecting Economic Traits in Egyptian Sheep Breeds. MSc thesis, Egypt: Alexandria University; 2016
27.Gkiasta M, Russell T, Shennan S, Steele J. Neolithic transition in Europe: The radiocarbon record revisited. Antiquity. 2003;77(295):45-62
28.Pereira F, Queirós S, Gusmão L, Nijman IJ, Cuppen E, Lenstra JA, et al. Tracing the history of goat pastoralism: New clues from mitochondrial and Y chromosome DNA in North Africa. Molecular Biology and Evolution. 2009;26(12):2765-2773
29.Hassan FA. Climate and cattle in North Africa: a first approximation. The origins and development of African livestock: Archaeology, genetics, linguistics and ethnography. Taylor and Francies Group. 2006. p. 61. https://www.taylorfrancis.com/chapters/mono/10.4324/9780203984239-16
30.Newman JL. The peopling of Africa: a geographic interpretation. Yale University Press; 1995. Available from: https://www.amazon.com/Peopling-Africa-Geographic-Interpretation/dp/0300072805
31.Blench R, MacDonald K. The origins and development of African livestock: archaeology, genetics, linguistics and ethnography. Routledge, Taylor & Francis Group; 2006. DOI: 10.4324/9780203984239
32.Sanchez-Mazas A, Blench R, Ross MD, Peiros I, Lin M. Past human migrations in East Asia: Matching archaeology, linguistics and genetics. Routledge, Taylor & Francis Group; 2008. DOI: 10.4324/9780203926789
33.Porter V. Goats of the world. Farming Press; CAB Direct (CABI); 1996. pp.xii + 179 pp. ref.80
34.Zeder MA. Domestication and early agriculture in the Mediterranean Basin: Origins, diffusion, and impact. Proceedings of the National Academy of Sciences. 2008;105(33):11597-11604
35.Colli L, Milanesi M, Talenti A, Bertolini F, Chen M, Crisà A, et al. Genome-wide SNP profiling of worldwide goat populations reveals strong partitioning of diversity and highlights post-domestication migration routes. Genetics, Selection, Evolution. 2018;50(1):58
36.Taberlet P, Valentini A, Rezaei H, Naderi S, Pompanon F, Negrini R, et al. Are cattle, sheep, and goats endangered species? Molecular Ecology. 2008;17(1):275-284
37.Taberlet P, Coissac E, Pansu J, Pompanon F. Conservation genetics of cattle, sheep, and goats. Comptes Rendus Biologies. 2011;334(3):247-254
38.Decker JE, McKay SD, Rolf MM, Kim J, Alcalá AM, Sonstegard TS, et al. Worldwide patterns of ancestry, divergence, and admixture in domesticated cattle. PLoS Genetics. 2014;10(3):e1004254
39.Daly KG, Delser PM, Mullin VE, Scheu A, Mattiangeli V, Teasdale MD, et al. Ancient goat genomes reveal mosaic domestication in the Fertile Crescent. Science. 2018;361(6397):85-88
40.Kijas JW, Lenstra JA, Hayes B, Boitard S, Neto LRP, San Cristobal M, et al. Genome-wide analysis of the world's sheep breeds reveals high levels of historic mixture and strong recent selection. PLoS Biology. 2012;10(2):e1001258
41.McTavish EJ, Decker JE, Schnabel RD, Taylor JF, Hillis DM. New World cattle show ancestry from multiple independent domestication events. Proceedings of the National Academy of Sciences. 2013;110(15):E1398-EE406
42.Canon J, García D, García-Atance M, Obexer-Ruff G, Lenstra J, Ajmone-Marsan P, et al. Geographical partitioning of goat diversity in Europe and the Middle East. Animal Genetics. 2006;37(4):327-334
43.Periasamy K, Vahidi S, Silva P, Faruque M, Naqvi A, Basar M, et al. Mapping molecular diversity of indigenous goat genetic resources of Asia. Small Ruminant Research. 2017;148:2-10
44.Luikart G, Gielly L, Excoffier L, Vigne J-D, Bouvet J, Taberlet P. Multiple maternal origins and weak phylogeographic structure in domestic goats. Proceedings of the National Academy of Sciences. 2001;98(10):5927-5932
45.Kul BC, Bilgen N, Lenstra J, Agaoglu OK, Akyuz B, Ertugrul O. Y-chromosomal variation of local goat breeds of T urkey close to the domestication Centre. Journal of Animal Breeding and Genetics. 2015;6(132):449-453
46.Waki A, Sasazaki S, Kobayashi E, Mannen H. Paternal phylogeography and genetic diversity of East Asian goats. Animal Genetics. 2015;46(3):337-339
47.Colli L, Lancioni H, Cardinali I, Olivieri A, Capodiferro MR, Pellecchia M, et al. Whole mitochondrial genomes unveil the impact of domestication on goat matrilineal variability. BMC Genomics. 2015;16(1):1115
48.Scherf BD. World watch list for domestic animal diversity: Food and Agriculture Organization (FAO). Animal Production and Health Div.; 2000
49.Faostat F. Disponível em:< http://faostat. fao. org>. Acesso em. 2012;14
50.Zhao Y, Zhao R, Zhao Z, Xu H, Zhao E, Zhang J. Genetic diversity and molecular phylogeography of Chinese domestic goats by large-scale mitochondrial DNA analysis. Molecular Biology Reports. 2014;41(6):3695-3704
51.Skapetas B, Bampidis V. Goat production in the world: Present situation and trends. Livestock Research for Rural Development. 2016;28(11):200
52.Chen S-Y, Su Y-H, Wu S-F, Sha T, Zhang Y-P. Mitochondrial diversity and phylogeographic structure of Chinese domestic goats. Molecular Phylogenetics and Evolution. 2005;37(3):804-814
53.Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, et al. The sequence alignment/map format and SAMtools. Bioinformatics. 2009;25(16):2078-2079
54.Seré C, Steinfeld H, Groenewold J. World Livestock Production Systems: Food and Agriculture Organization of the United Nations; 1996
55.Hoffmann I. Climate change and the characterization, breeding and conservation of animal genetic resources. Animal Genetics. 2010;41:32-46
56.Sultana S, Mannen H, Tsuji S. Mitochondrial DNA diversity of Pakistani goats. Animal Genetics. 2003;34(6):417-421
57.Zhao Y, Zhang J, Zhao E, Zhang X, Liu X, Zhang N. Mitochondrial DNA diversity and origins of domestic goats in Southwest China (excluding Tibet). Small Ruminant Research. 2011;95(1):40-47
58.Álvarez I, Royo L, Fernandez I, Gutiérrez J, Gómez E, Goyache F. Genetic relationships and admixture among sheep breeds from northern Spain assessed using microsatellites. Journal of Animal Science. 2004;82(8):2246-2252
59.Mukesh M, Sodhi M, Bhatia S. Microsatellite-based diversity analysis and genetic relationships of three Indian sheep breeds. Journal of Animal Breeding and Genetics. 2006;123(4):258-264
60.Guan D, Luo N, Tan X, Zhao Z, Huang Y, Na R, et al. Scanning of selection signature provides a glimpse into important economic traits in goats (Capra hircus). Scientific Reports. 2016;6:36372
61.Zeder MA, Hesse B. The initial domestication of goats (Capra hircus) in the Zagros Mountains 10,000 years ago. Science. 2000;287(5461):2254-2257
62.Benjelloun B, Alberto FJ, Streeter I, Boyer F, Coissac E, Stucki S, et al. Characterizing neutral genomic diversity and selection signatures in indigenous populations of Moroccan goats (Capra hircus) using WGS data. Frontiers in Genetics. 2015;6:107
63.Dong Y, Zhang X, Xie M, Arefnezhad B, Wang Z, Wang W, et al. Reference genome of wild goat (capra aegagrus) and sequencing of goat breeds provide insight into genic basis of goat domestication. BMC Genomics. 2015;16(1):431
64.Brito LF, Kijas JW, Ventura RV, Sargolzaei M, Porto-Neto LR, Cánovas A, et al. Genetic diversity and signatures of selection in various goat breeds revealed by genome-wide SNP markers. BMC Genomics. 2017;18(1):229
65.Zhang B, Chang L, Lan X, Asif N, Guan F, Fu D, et al. Genome-wide definition of selective sweeps reveals molecular evidence of trait-driven domestication among elite goat (Capra species) breeds for the production of dairy, cashmere, and meat. GigaScience. 2018;7(12):giy105
66.Babiker S, El Khider I, Shafie S. Chemical composition and quality attributes of goat meat and lamb. Meat Science. 1990;28(4):273-277
67.Webb E, Casey N, Simela L. Goat meat quality. Small Ruminant Research. 2005;60(1–2):153-166
68.Schönfeldt H, Naude R, Bok W, Van Heerden S, Smit R, Boshoff E. Flavour-and tenderness-related quality characteristics of goat and sheep meat. Meat Science. 1993;34(3):363-379
69.Casey NH, Webb EC. Managing goat production for meat quality. Small Ruminant Research. 2010;89(2–3):218-224
70.Tshabalala P, Strydom P, Webb E, De Kock H. Meat quality of designated south African indigenous goat and sheep breeds. Meat Science. 2003;65(1):563-570
71.Cramer D. Chemical-compounds implicated IN lamb flavor. Food Technology. 1983;37(5):249-257
72.Mallatou H, Pappas C, Voutsinas L. Manufacture of feta cheese from sheep's milk, goats' milk or mixtures of these milks. International Dairy Journal. 1994;4(7):641-664
73.Wong E, Nixon LN, Johnson CB. Volatile medium chain fatty acids and mutton flavor. Journal of Agricultural and Food Chemistry. 1975;23(3):495-498
74.Ha JK, Lindsay R. Distribution of volatile branched-chain fatty acids in perinephric fats of various red meat species. Lebensmittel-Wissenschaft und Technologie. 1990;23(5):433-440
75.Scholtens M, Jiang A, Smith A, Littlejohn M, Lehnert K, Snell R, et al. Genome-wide association studies of lactation yields of milk, fat, protein and somatic cell score in New Zealand dairy goats. Journal of Animal Science and Biotechnology. 2020;11(1):1-14
76.Vargas-Bello-Pérez E, Tajonar K, Foggi G, Mele M, Simitzis P, Mavrommatis A, et al. Consumer attitudes toward dairy products from sheep and goats: A cross-continental perspective. Journal of Dairy Science. 2022;105(11):8718-8733
77.Saleh AA, Rashad A, Hassanine N, Sharaby MA, Zhao Y. Assessment of hair and cashmere properties and their genetic background of several goat breeds in Southwest China. Scientific Reports. 2022;12(1):1-15
78.Arora R, Bhojak N, Joshi R. Comparative aspects of goat and cow milk. International Journal of Engineering Science Invention. 2013;2(1):07-10
79.Alvinerie M, Sutra J, Galtier P. Ivermectin in goat plasma and milk after subcutaneous injection. Veterinary Research. 1993;24(5):417-421
80.Delgadillo JA, Vielma J, Hernandez H, Flores JA, Duarte G, Fernández IG, et al. Male goat vocalizations stimulate the estrous behavior and LH secretion in anestrous goats that have been previously exposed to bucks. Hormones and Behavior. 2012;62(4):525-530
81.Delgadillo JA, Flores J, Hernández H, Poindron P, Keller M, Fitz-Rodríguez G, et al. Sexually active males prevent the display of seasonal anestrus in female goats. Hormones and Behavior. 2015;69:8-15
83.Zhao Z, Yang T, Qiao L, He Q, Dai Z. Reproductive characteristics of Dazu black goats, a newly discovered Chinese indigenous breed resource with high litter sizes. Pakistan Journal of Zoology. 2019;51(2)
84.Yuan C, Wang X, Geng R, He X, Qu L, Chen Y. Discovery of cashmere goat (Capra hircus) microRNAs in skin and hair follicles by Solexa sequencing. BMC Genomics. 2013;14(1):511
85.McDonald C. Rent-a-goats gain footholds. Seattle Post-Intelligencer. 2007;2007:13
86.Luginbuhl J, Green J, Mueller J, Poore M, editors. Meat goats in land and forage management. Proceedings if the Southeast Regional Meat Goat Production Symposium “Meat Goat Production in the Southeast–Today and Tomorrow”. February 21-24, 1996. Florida A&M University, Tallahassee.http://www.cals.ncsu.edu/an_sci/extension/animal/meatgoat/MGLand.htm
87.Porr C, Childs C, Downing AK. Options for Clearing Land: Pasture Establishment for Horses. Publications, Virginia Cooperative Extension (VCE); 2009. p. 8181. http://hdl.handle.net/10919/48543
88.Eibl-Eibesfeldt I. The fighting behavior of animals. Scientific American. 1961;205(6):112-123
89.Kennedy BW. Training military medics with animal models. Laboratory Animals. 2013;42(7):259-260
90.Martinic G. The use of animals in live-tissue trauma training and military medical research. Laboratory Animals. 2011;40(10):319
91.Thornton PK. Livestock production: Recent trends, future prospects. Philosophical Transactions of the Royal Society, B: Biological Sciences. 2010;365(1554):2853-2867
92.Dekkers CM. Application of genomics tools to animal breeding. Current Genomics. 2012;13(3):207-212
93.Hansen M. Differences in susceotibility to Marek's disease in chickens carring two different B locus blood group alleles. Poultry Science. 1967;46:1268
94.Ahmed Saleh MHH, Dabour NA, Hafez EE, Sharaby M. IGFBP-3 gene diversity among Egyptian sheep breeds. Alexandria Journal of Agricultural Science. 2017;6(7250):87-92
95.Saleh AA, Hammoud M, Dabour NA, Hafez E, Sharaby MA. BMPR-1B, BMP-15 and GDF-9 genes structure and their relationship with litter size in six sheep breeds reared in Egypt. BMC Research Notes. 2020;13:1-7
96.Pophiwa P, Webb EC, Frylinck L. A review of factors affecting goat meat quality and mitigating strategies. Small Ruminant Research. 2020;183:106035
97.Tezera BT, Engidashet DH. Traditional sheep breeding practices under agroforestry system of Gedeo zone, southern Ethiopia. PLoS One. 2022;17(6):e0269263
98.Falconer D, Mackay T. Introduction to Quantitative Genetics. Harlow, UK: Longman Group Ltd.; 1996
99.Taylor JF, McKay SD, Rolf MM, Ramey HR, Decker JE, Schnabel RD. Genomic selection in beef cattle. Bovine Genomics. 2012;2012:211-233
100.Watson P, Holt WV. Cryobanking the genetic resource: Wildlife conservation for the future?. CRC Press; 2001. Available from: https://www.amazon.com/Cryobanking-Genetic-Resource-Wildlife-Conservation/dp/0748408142
101.Camara M, Symonds J. Genetic improvement of New Zealand aquaculture species: Programmes, progress and prospects. New Zealand Journal of Marine and Freshwater Research. 2014;48(3):466-491
102.Samore AB, Fontanesi L. Genomic selection in pigs: State of the art and perspectives. Italian Journal of Animal Science. 2016;15(2):211-232
103.Henderson CR. Applications of linear models in animal breeding. University of Guelph Guelph; CAB Direct (CABI). 1984. pp.xxiii + 462 pp. ref.8 pp. Available from: https://www.cabdirect.org/cabdirect/abstract/19860193912
104.Lynch M, Walsh B. Genetics and Analysis of Quantitative Traits: Sinauer. Sunderland, MA; 1998
105.Tambasco D, Paz C, Tambasco-Studart M, Pereira A, Alencar M, Freitas A, et al. Candidate genes for growth traits in beef cattle crosses Bos taurus× Bos indicus. Journal of Animal Breeding and Genetics. 2003;120(1):51-56
106.Supakorn C. The important candidate genes in goats—A review. Walailak Journal of Science and Technology. 2009;6(1):17-36
107.Legarra A, Aguilar I, Misztal I. A relationship matrix including full pedigree and genomic information. Journal of Dairy Science. 2009;92(9):4656-4663
108.Munoz PR, Resende MF, Huber DA, Quesada T, Resende MD, Neale DB, et al. Genomic relationship matrix for correcting pedigree errors in breeding populations: Impact on genetic parameters and genomic selection accuracy. Crop Science. 2014;54(3):1115-1123
109.Rupp R, Mucha S, Larroque H, McEwan J, Conington J. Genomic application in sheep and goat breeding. Animal Frontiers. 2016;6(1):39-44
110.Dong Y, Xie M, Jiang Y, Xiao N, Du X, Zhang W, et al. Sequencing and automated whole-genome optical mapping of the genome of a domestic goat (Capra hircus). Nature Biotechnology. 2013;31(2):135
111.Jiang Y, Xie M, Chen W, Talbot R, Maddox JF, Faraut T, et al. The sheep genome illuminates biology of the rumen and lipid metabolism. Science. 2014;344(6188):1168-1173
112.Sharma A, Park J-E, Chai H-H, Jang G-W, Lee S-H, Lim D. Next generation sequencing in livestock species-a review. Journal of Animal Breeding and Genomics JABG. 2017;1:23-30
113.Patel SM, Koringa PG, Nathani NM, Patel NV, Shah TM, Joshi CG. Exploring genetic polymorphism in innate immune genes in Indian cattle (Bos indicus) and buffalo (Bubalus bubalis) using next generation sequencing technology. Meta Gene. 2015;3:50-58
114.Tosser-Klopp G, Bardou P, Bouchez O, Cabau C, Crooijmans R, Dong Y, et al. Design and characterization of a 52K SNP chip for goats. PLoS One. 2014;9(1):e86227
115.Fan B, Du Z-Q, Gorbach DM, Rothschild MF. Development and application of high-density SNP arrays in genomic studies of domestic animals. Asian-Australasian Journal of Animal Sciences. 2010;23(7):833-847
116.Saleh AA, Rashad AM, Hassanine NN, Sharaby MA, Zhao Y. Comparative analysis of IGFBP-3 gene sequence in Egyptian sheep, cattle, and buffalo. BMC Research Notes. 2019;12(1):623
117.Schaeffer L. Strategy for applying genome-wide selection in dairy cattle. Journal of Animal Breeding and Genetics. 2006;123(4):218-223
118.Altermann W, Kazmierczak J. Archean microfossils: A reappraisal of early life on earth. Research in Microbiology. 2003;154(9):611-617
119.Snyder M, Du J, Gerstein M. Personal genome sequencing: Current approaches and challenges. Genes & Development. 2010;24(5):423-431
120.Ahmed Saleh NN. Hassanine. Egyptian Sheep Breeds and genetic tools to improve. LAP Lambert Academic Publishing. Available from: https://www.lap-publishing.com/catalog/details//store/gb/book/978-620-0-47168-0/egyptian-sheep-breeds-and-genetic-tools-to-improve
121.Qureshi MI, Sabir J, Mutawakil M, El Hanafy A, Ashmaoui H, Ramadan H, et al. Review of modern strategies to enhance livestock genetic performance: From molecular markers to next-generation sequencing technologies in goats. Journal of Food, Agriculture and Environment. 2014;12(7):5
122.Pariset L, Cappuccio I, Marsan PA, Dunner S, Luikart G, England P, et al. Assessment of population structure by single nucleotide polymorphisms (SNPs) in goat breeds. Journal of Chromatography B. 2006;833(1):117-120
123.Pariset L, Cuteri A, Ligda C, Ajmone-Marsan P, Valentini A. Geographical patterning of sixteen goat breeds from Italy, Albania and Greece assessed by single nucleotide polymorphisms. BMC Ecology. 2009;9(1):20
124.Consortium BH. Genome-wide survey of SNP variation uncovers the genetic structure of cattle breeds. Science. 2009;324(5926):528-532
125.Muir WM, Wong GK-S, Zhang Y, Wang J, Groenen MA, Crooijmans RP, et al. Genome-wide assessment of worldwide chicken SNP genetic diversity indicates significant absence of rare alleles in commercial breeds. Proceedings of the National Academy of Sciences. 2008;105(45):17312-17317
126.Talenti A, Palhière I, Tortereau F, Pagnacco G, Stella A, Nicolazzi EL, et al. Functional SNP panel for parentage assessment and assignment in worldwide goat breeds. Genetics, Selection, Evolution. 2018;50(1):55
127.Consortium IH. A second generation human haplotype map of over 3.1 million SNPs. Nature. 2007;449(7164):851
128.Consortium ICPM. A genetic variation map for chicken with 2.8 million single-nucleotide polymorphisms. Nature. 2004;432(7018):717
129.Lindblad-Toh K, Wade CM, Mikkelsen TS, Karlsson EK, Jaffe DB, Kamal M, et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. Nature. 2005;438(7069):803
130.Frazer KA, Eskin E, Kang HM, Bogue MA, Hinds DA, Beilharz EJ, et al. A sequence-based variation map of 8.27 million SNPs in inbred mouse strains. Nature. 2007;448(7157):1050
131.Van Tassell CP, Smith TP, Matukumalli LK, Taylor JF, Schnabel RD, Lawley CT, et al. SNP discovery and allele frequency estimation by deep sequencing of reduced representation libraries. Nature Methods. 2008;5(3):247
132.Fu W, Wang R, Yu J, Hu D, Cai Y, Shao J, et al. GGVD: A goat genome variation database for tracking the dynamic evolutionary process of selective signatures and ancient introgressions. Journal of Genetics and Genomics. 2021;48(3):248-256
133.Charlier C, Coppieters W, Rollin F, Desmecht D, Agerholm JS, Cambisano N, et al. Highly effective SNP-based association mapping and management of recessive defects in livestock. Nature Genetics. 2008;40(4):449
134.Saleh A, Rashad A, Hassanine N, Sharaby MA, Zhao Y. Morphological body measurements, body indices, and their genetic background for several Chinese goat breeds. Tropical Animal Health and Production. 2022;54(4):1-16
135.Bordbar F, Jensen J, Zhu B, Wang Z, Xu L, Chang T, et al. Identification of muscle-specific candidate genes in Simmental beef cattle using imputed next generation sequencing. PLoS One. 2019;14(10):e0223671
136.Yang L, Niu Q, Zhang T, Zhao G, Zhu B, Chen Y, et al. Genomic sequencing analysis reveals copy number variations and their association with economically important traits in beef cattle. 2021. Genomics. 2021;113(1, Part 2):812-820. DOI: 10.1016/j.ygeno.2020.10.012
137.El-Khishin DA, Ageez A, Saad ME, Ibrahim A, Shokrof M, Hassan LR, et al. Sequencing and assembly of the Egyptian buffalo genome. PLoS One. 2020;15(8):e0237087
138.Castañeda-Rico S, León-Paniagua L, Edwards CW, Maldonado JE. Ancient DNA from museum specimens and next generation sequencing help resolve the controversial evolutionary history of the critically endangered Puebla deer mouse. Frontiers in Ecology and Evolution. 2020;8:94
139.Gu J, Li S. Next-generation sequencing of the complete mitochondrial genome of the Nixi chicken (Gallus gallus). Mitochondrial DNA Part B. 2020;5(3):3271-3272
140.Wu F, Sun H, Lu S, Gou X, Yan D, Xu Z, et al. Genetic diversity and selection signatures within Diannan small-ear pigs revealed by next-generation sequencing. Frontiers in Genetics. 2020;11:733
141.Stella A, Nicolazzi EL, Van Tassell CP, Rothschild MF, Colli L, Rosen BD, et al. AdaptMap: Exploring goat diversity and adaptation. BioMed Central. 2018;2018
142.Tosser-Klopp G, Bardou P, Bouchez O, Cabau C, Crooijmans R, Dong Y, et al. Correction: Design and characterization of a 52K SNP Chip for goats. PLoS One. 2016;11(3):e0152632
143.Michelizzi VNDM, Pan Z, Amaral MEJ, Michal JJ, McLean DJ, Womack JE, et al. Water Buffalo genome science comes of age. International Journal of Biological Sciences. 2010;6(4):333-349
144.Groenen MA, Megens H-J, Zare Y, Warren WC, Hillier LW, Crooijmans RP, et al. The development and characterization of a 60K SNP chip for chicken. BMC Genomics. 2011;12(1):274
145.Chou H-J. An algorithm for structural variant detection with third-generation sequencing. MSc thesis. Graduate School-Camden. Rutgers University (The State University of New Jersey) 2017. Available from: https://rucore.libraries.rutgers.edu/rutgers-lib/52726/PDF/1/play
146.Heaton MP, Leymaster KA, Kalbfleisch TS, Kijas JW, Clarke SM, McEwan J, et al. SNPs for parentage testing and traceability in globally diverse breeds of sheep. PLoS One. 2014;9(4):e94851
147.Talenti A, Nicolazzi E, Chessa S, Frattini S, Moretti R, Coizet B, et al. A method for single nucleotide polymorphism selection for parentage assessment in goats. Journal of Dairy Science. 2016;99(5):3646-3653
148.Burren A, Neuditschko M, Signer-Hasler H, Frischknecht M, Reber I, Menzi F, et al. Genetic diversity analyses reveal first insights into breed-specific selection signatures within Swiss goat breeds. Animal Genetics. 2016;47(6):727-739
149.Mdladla K, Dzomba E, Huson H, Muchadeyi F. Population genomic structure and linkage disequilibrium analysis of South African goat breeds using genome-wide SNP data. Animal Genetics. 2016;47(4):471-482
150.McRae KM, McEwan JC, Dodds KG, Gemmell NJ. Signatures of selection in sheep bred for resistance or susceptibility to gastrointestinal nematodes. BMC genomics. 2014;15(1):637. p: 1-13. DOI: 10.1186/1471-2164-15-637
151.Moradi MH, Nejati-Javaremi A, Moradi-Shahrbabak M, Dodds KG, McEwan JC. Genomic scan of selective sweeps in thin and fat tail sheep breeds for identifying of candidate regions associated with fat deposition. BMC Genetics. 2012;13(1):10
152.Makina SO, Muchadeyi FC, Marle-Köster E, Taylor JF, Makgahlela ML, Maiwashe A. Genome-wide scan for selection signatures in six cattle breeds in South Africa. Genetics, Selection, Evolution. 2015;47(1):92
153.Zhao F, McParland S, Kearney F, Du L, Berry DP. Detection of selection signatures in dairy and beef cattle using high-density genomic information. Genetics, Selection, Evolution. 2015;47(1):49
154.Ai H, Yang B, Li J, Xie X, Chen H, Ren J. Population history and genomic signatures for high-altitude adaptation in Tibetan pigs. BMC Genomics. 2014;15(1):834
155.Ai H, Huang L, Ren J. Genetic diversity, linkage disequilibrium and selection signatures in Chinese and Western pigs revealed by genome-wide SNP markers. PLoS One. 2013;8(2):e56001
156.Kowalewska-Łuczak I, Kulig H. Polymorphism of the FAM13A, ABCG2, OPN, LAP3, HCAP-G, PPARGC1A genes and somatic cell count of Jersey cows–preliminary study. Research in Veterinary Science. 2013;94(2):252-255
157.Cohen M, Reichenstein M, Everts-van der Wind A, Heon-Lee J, Shani M, Lewin HA, et al. Cloning and characterization of FAM13A1—A gene near a milk protein QTL on BTA6: Evidence for population-wide linkage disequilibrium in Israeli Holsteins. Genomics. 2004;84(2):374-383
158.Hatzirodos N, Hummitzsch K, Irving-Rodgers HF, Harland ML, Morris SE, Rodgers RJ. Transcriptome profiling of granulosa cells from bovine ovarian follicles during atresia. BMC Genomics. 2014;15(1):40
159.Sugimoto M, Sasaki S, Gotoh Y, Nakamura Y, Aoyagi Y, Kawahara T, et al. Genetic variants related to gap junctions and hormone secretion influence conception rates in cows. Proceedings of the National Academy of Sciences. 2013;110(48):19495-19500
160.Wang K, Liu D, Hernandez-Sanchez J, Chen J, Liu C, Wu Z, et al. Genome wide association analysis reveals new production trait genes in a male Duroc population. PLoS One. 2015;10(9):e0139207
161.Felix JF, Bradfield JP, Monnereau C, Van Der Valk RJ, Stergiakouli E, Chesi A, et al. Genome-wide association analysis identifies three new susceptibility loci for childhood body mass index. Human Molecular Genetics. 2015;25(2):389-403
162.Cole JB, Wiggans GR, Ma L, Sonstegard TS, Lawlor TJ, Crooker BA, et al. Genome-wide association analysis of thirty one production, health, reproduction and body conformation traits in contemporary US Holstein cows. BMC Genomics. 2011;12(1):408
163.Zhang H, Wang S-Z, Wang Z-P, Da Y, Wang N, Hu X-X, et al. A genome-wide scan of selective sweeps in two broiler chicken lines divergently selected for abdominal fat content. BMC Genomics. 2012;13(1):704
164.Mahdessian H, Taxiarchis A, Popov S, Silveira A, Franco-Cereceda A, Hamsten A, et al. TM6SF2 is a regulator of liver fat metabolism influencing triglyceride secretion and hepatic lipid droplet content. Proceedings of the National Academy of Sciences. 2014;2014:201323785
165.Martin P, Raoul J, Bodin L. Effects of the FecL major gene in the Lacaune meat sheep population. Genetics, Selection, Evolution. 2014;46(1):48
166.Kim E-S, Elbeltagy A, Aboul-Naga A, Rischkowsky B, Sayre B, Mwacharo JM, et al. Multiple genomic signatures of selection in goats and sheep indigenous to a hot arid environment. Heredity. 2016;116(3):255
167.Donaldson IJ, Amin S, Hensman JJ, Kutejova E, Rattray M, Lawrence N, et al. Genome-wide occupancy links Hoxa2 to Wnt–β-catenin signaling in mouse embryonic development. Nucleic Acids Research. 2012;40(9):3990-4001
168.Bris C, Goudenege D, Desquiret-Dumas V, Charif M, Colin E, Bonneau D, et al. Bioinformatics tools and databases to assess the pathogenicity of mitochondrial DNA variants in the field of next generation sequencing. Frontiers in Genetics. 2018;9:632
169.Goudenège D, Bris C, Hoffmann V, Desquiret-Dumas V, Jardel C, Rucheton B, et al. eKLIPse: A sensitive tool for the detection and quantification of mitochondrial DNA deletions from next-generation sequencing data. Genetics in Medicine. 2019;21(6):1407-1416
170.Ruo-Yu L, Gong-She Y, Chu-Zhao L. The genetic diversity of mtDNA D-loop and the origin of Chinese goats. Acta Genetica Sinica. 2006;33(5):420-428
171.Ladoukakis ED, Zouros E. Evolution and inheritance of animal mitochondrial DNA: Rules and exceptions. Journal of Biological Research (Thessaloniki). 2017;24(1):1-7
172.Smith S, Turbill C, Suchentrunk F. Introducing mother’s curse: Low male fertility associated with an imported mtDNA haplotype in a captive colony of brown hares. Molecular Ecology. 2010;19(1):36-43
173.Mannen H, Kojima T, Oyama K, Mukai F, Ishida T, Tsuji S. Effect of mitochondrial DNA variation on carcass traits of Japanese black cattle. Journal of Animal Science. 1998;76(1):36-41
174.Colli L, Lancioni H, Cardinali I, Olivieri A, Capodiferro MR, Pellecchia M, et al. Whole mitochondrial genomes unveil the impact of domestication on goat matrilineal variability. BMC Genomics. 2015;16(1):1-12
175.Baenyi Simon P, Owino Junga J, Tarekegn GM, Machuka E, Tiambo CK, Kabange D, et al. Haplotype analysis of the mitochondrial DNA d-loop region reveals the maternal origin and historical dynamics among the indigenous goat populations in east and west of the Democratic Republic of Congo. Ecology and Evolution. 2022;12(3):e8713
176.De AK, Muthiyan R, Sunder J, Sawhney S, Sujatha T, Bhattacharya D. The whole mitochondrial genome signature of Teressa goat, an indigenous goat germplasm of Andaman and Nicobar Islands, India. Small Ruminant Research. 2022;2022:106848
177.Zhang L, Liu J, Zhao F, Ren H, Xu L, Lu J, et al. Genome-wide association studies for growth and meat production traits in sheep. PLoS One. 2013;8(6):e66569
178.Database. AQ. Animal QTLdb. 2022. https://www.animalgenome.org/cgi-bin/QTLdb/index. Retrieved Aug 24, 2022
179.Hu Z-L, Park CA, Wu X-L, Reecy JM. Animal QTLdb: An improved database tool for livestock animal QTL/association data dissemination in the post-genome era. Nucleic Acids Research. 2013;41(D1):D871-D8D9
180.LB. Week 5. 2017. https://xmlpipedb.cs.lmu.edu/biodb/fall2017/index.php/Week_5. Retrieved September 28, 2017
181.Bioinformatics. USN. Genome Informatics Resources. 2017. https://www.animalgenome.org/bioinfo/.Retrieved October 1, 2017
182.Hu Z-L, Park CA, Reecy JM. Bringing the animal QTLdb and CorrDB into the future: Meeting new challenges and providing updated services. Nucleic Acids Research. 2022;50(D1):D956-DD61
183.Demars J, Fabre S, Sarry J, Rossetti R, Gilbert H, Persani L, et al. Genome-wide association studies identify two novel BMP15 mutations responsible for an atypical hyperprolificacy phenotype in sheep. PLoS Genetics. 2013;9(4):e1003482
184.Mahrous KF, Abdel-Aziem SH, Abdel-Hafez MA, Abdel-Mordy M, Rushdi HE. Polymorphism of growth hormone gene in three goat breeds in Egypt. Bulletin of the National Research Centre. 2018;42(1):35
185.Shareef M, Atia Basheer IZ, Anjum AA. Polymorphisms IN growth hormone (GH) and insulin-like growth FACTOR-1 (IGF-1) gene and their association with growth traits IN BEETAL goat. Pakistan Journal of Agricultural Sciences. 2018;55(3):713-720
186.Mikawa S, Yoshikawa G-I, Aoki H, Yamano Y, Sakai H, Komano T. Dynamic aspects in the expression of the goat insulin-like growth factor-I (IGF-I) gene: Diversity in transcription and post-transcription. Bioscience, Biotechnology, and Biochemistry. 1995;59(1):87-92
187.Tait RG Jr, Cushman RA, McNeel AK, Casas E, Smith TP, Freetly HC, et al. μ-Calpain (CAPN1), calpastatin (CAST), and growth hormone receptor (GHR) genetic effects on Angus beef heifer performance traits and reproduction. Theriogenology. 2018;113:1-7
188.Zhou F, Yang Q, Lei C, Chen H, Lan X. Relationship between genetic variants of POU1F1, PROP1, IGFBP3 genes and milk performance in Guanzhong dairy goats. Small Ruminant Research. 2016;140:40-45
189.BAKIRCIOĞLU Ö, ÖZTABAK KÖ. Characterization of exon 2 and intron 2 of leptin gene in native Anatolian goat breeds. İstanbul Üniversitesi Veteriner Fakültesi Dergisi. 2016;42(2):178-185
190.Wang X, Yu H, Lei A, Zhou J, Zeng W, Zhu H, et al. Generation of gene-modified goats targeting MSTN and FGF5 via zygote injection of CRISPR/Cas9 system. Scientific Reports. 2015;5:13878
191.Wang L, Cai B, Zhou S, Zhu H, Qu L, Wang X, et al. RNA-seq reveals transcriptome changes in goats following myostatin gene knockout. PLoS One. 2017;12(12):e0187966
192.Saeedabadi S, Abazari-Kia AH, Rajabi H, Parivar K, Salehi M. Melatonin improves the developmental competence of goat oocytes. International Journal of Fertility & Sterility. 2018;12(2):157
193.Fábián R, Kovács A, Stéger V, Frank K, Egerszegi I, Oláh J, et al. X-and Y-chromosome-specific variants of the amelogenin gene allow non-invasive sex diagnosis for the detection of pseudohermaphrodite goats. Acta Veterinaria Hungarica. 2017;65(4):500-504
194.Zhou M, Yu J, Xue R, Li B, Yang J. A reliable non-invasive PCR method for takin (Budorcas taxicolor) sex identification based on amelogenin gene. Conservation Genetics Resources. 2018:1-4
195.Tsai T, Wu S, Chen H, Tung Y, Cheng W, Huang J, et al. Identification of sex-specific polymorphic sequences in the goat amelogenin gene for embryo sexing. Journal of Animal Science. 2011;89(8):2407-2414
196.Payen EJ, Cotinot CY. Comparative HMG-box sequences of the SRY gene between sheep, cattle and goats. Nucleic Acids Research. 1993;21(11):2772
197.Sacchi P, Chessa S, Budelli E, Bolla P, Ceriotti G, Soglia D, et al. Casein haplotype structure in five Italian goat breeds. Journal of Dairy Science. 2005;88(4):1561-1568
198.Teissier M, Larroque H, Robert-Granié C. Weighted single-step genomic BLUP improves accuracy of genomic breeding values for protein content in French dairy goats: A quantitative trait influenced by a major gene. Genetics, Selection, Evolution. 2018;50(1):31
199.Perna A, Simonetti A, Grassi G, Gambacorta E. Effect of αS1-casein genotype on phenolic compounds and antioxidant activity in goat milk yogurt fortified with Rhus coriaria leaf powder. Journal of Dairy Science. 2018;101(9):7691-7701
200.Leroux C, Martin P, Mahé MF, Levéziel H, Mercier JC. Restriction fragment length polymorphism identification of goat αs1-casein alleles: A potential tool in selection of individuals carrying alleles associated with a high level protein synthesis. Animal Genetics. 1990;21(4):341-351
201.Li J, Chen W, Wu S, Ma T, Jiang H, Zhang Q. Differential expression of MC1R gene in Liaoning cashmere goats with different coat colors. Animal Biotechnology. 2018;2018:1-6
202.Guo J, Tao H, Li P, Li L, Zhong T, Wang L, et al. Whole-genome sequencing reveals selection signatures associated with important traits in six goat breeds. Scientific Reports. 2018;8(1):10405
203.Gowane G, Akram N, Misra S, Prakash V, Kumar A. Genetic diversity of Cahi DRB and DQB genes of caprine MHC class II in Sirohi goat. Journal of Genetics. 2018;97(2):483-492
204.Våge DI, Boman IA. A nonsense mutation in the beta-carotene oxygenase 2 (BCO2) gene is tightly associated with accumulation of carotenoids in adipose tissue in sheep (Ovis aries). BMC Genetics. 2010;11(1):10
205.Elsen J-M, Amigues Y, Schelcher F, Ducrocq V, Andreoletti O, Eychenne F, et al. Genetic susceptibility and transmission factors in scrapie: Detailed analysis of an epidemic in a closed flock of Romanov. Archives of Virology. 1999;144(3):431-445
206.Rupp R, Senin P, Sarry J, Allain C, Tasca C, Ligat L, et al. A point mutation in suppressor of cytokine signalling 2 (Socs2) increases the susceptibility to inflammation of the mammary gland while associated with higher body weight and size and higher milk production in a sheep model. PLoS Genetics. 2015;11(12):e1005629
207.Montgomery G, Kinghorn B. Recent developments in gene mapping and progress towards marker-assisted selection in sheep this review is one of a series invited by the Journal’s advisory committee. Australian Journal of Agricultural Research. 1997;48(6):729-741
208.Davis G. Fecundity genes in sheep. Animal Reproduction Science. 2004;82:247-253
209.Van Laere A-S, Nguyen M, Braunschweig M, Nezer C, Collette C, Moreau L, et al. A regulatory mutation in IGF2 causes a major QTL effect on muscle growth in the pig. Nature. 2003;425(6960):832
210.Dekkers JC. Commercial application of marker-and gene-assisted selection in livestock: Strategies and lessons. Journal of Animal Science. 2004;82(suppl_13):E313-EE28
211.Robinson J, Dombrowski D, Harpestad G, Shanks R. Detection and prevalence of UMP synthase deficiency among dairy cattle. The Journal of Heredity. 1984;75(4):277-280
212.Shuster DE, Kehrli ME, Ackermann MR, Gilbert RO. Identification and prevalence of a genetic defect that causes leukocyte adhesion deficiency in Holstein cattle. Proceedings of the National Academy of Sciences. 1992;89(19):9225-9229
213.Thomsen B, Horn P, Panitz F, Bendixen E, Petersen AH, Holm L-E, et al. A missense mutation in the bovine SLC35A3 gene, encoding a UDP-N-acetylglucosamine transporter, causes complex vertebral malformation. Genome Research. 2006;16(1):97-105
214.Belt PB, Muileman IH, Schreuder BE, Bos-de Ruijter J, Gielkens AL, Smits MA. Identification of five allelic variants of the sheep PrP gene and their association with natural scrapie. The Journal of General Virology. 1995;76(3):509-517
215.Arias JA, Keehan M, Fisher P, Coppieters W, Spelman R. A high density linkage map of the bovine genome. BMC Genetics. 2009;10(1):18
216.Consortium MGS, Waterston R, Lindblad-Toh K. Initial sequencing and comparative analysis of the mouse genome. Nature. 2002;420(6915):520-562
217.Vila C, Sundqvist AK, Flagstad Ø, Seddon J, Rnerfeldt SB, Kojola I, et al. Rescue of a severely bottlenecked wolf (Canis lupus) population by a single immigrant. Proceedings of the Royal Society of London, Series B: Biological Sciences. 2003;270(1510):91-97
218.Wallis JW, Aerts J, Groenen MA, Crooijmans RP, Layman D, Graves TA, et al. A physical map of the chicken genome. Nature. 2004;432(7018):761
219.Venkatesh B, Kirkness EF, Loh Y-H, Halpern AL, Lee AP, Johnson J, et al. Survey sequencing and comparative analysis of the elephant shark (Callorhinchus milii) genome. PLoS Biology. 2007;5(4):e101. DOI: 10.1371/journal.pbio.0050101
220.Gibbs RA, Rogers J, Katze MG, Bumgarner R, Weinstock GM, Mardis ER, et al. Evolutionary and biomedical insights from the rhesus macaque genome. Science. 2007;316(5822):222-234
221.Warren WC, Hillier LW, Graves JAM, Birney E, Ponting CP, Grützner F, et al. Genome analysis of the platypus reveals unique signatures of evolution. Nature. 2008;453(7192):175
222.Pontius JU, Mullikin JC, Smith DR, Team AS, Lindblad-Toh K, Gnerre S, et al. Initial sequence and comparative analysis of the cat genome. Genome Research. 2007;17(11):1675-1689
223.Dávila-Rodríguez M, Cortés-Gutiérrez E, López-Fernández C, Pita M, Mezzanotte R, Gosálvez J. Whole-comparative genomic hybridization in domestic sheep (Ovis aries) breeds. Cytogenetic and Genome Research. 2009;124(1):19-26
224.Amaral AJ, Megens H-J, Kerstens HH, Heuven HC, Dibbits B, Crooijmans RP, et al. Application of massive parallel sequencing to whole genome SNP discovery in the porcine genome. BMC Genomics. 2009;10(1):374
225.Tellam R, the Bovine Genome Sequencing and Analysis Consortium. What does analysis of the bovine genome sequence say about innate immunity. In: Plant and Animal Genome XVII Conference W. 2009
226.Wade C, Giulotto E, Sigurdsson S, Zoli M, Gnerre S, Imsland F, et al. Genome sequence, comparative analysis, and population genetics of the domestic horse. Science. 2009;326(5954):865-867
227.Hellsten U, Harland RM, Gilchrist MJ, Hendrix D, Jurka J, Kapitonov V, et al. The genome of the Western clawed frog Xenopus tropicalis. Science. 2010;328(5978):633-636
228.Li J, Wang L, Zhan Q, Chen Y. Establishment of Sorghum bicolor expressed sequence tag-simple sequence repeat (EST-SSR) marker and its preliminary application to Sorghum sudanense. Pratacultural Science. 2010;27(3):112-117
229.Ghosh P, Hsu C, Alyamani EJ, Shehata MM, Al-Dubaib MA, Al-Naeem A, et al. Genome-wide analysis of the emerging infection with Mycobacterium avium subspecies paratuberculosis in the Arabian camels (Camelus dromedarius). PLoS One. 2012;7(2):e31947
230.Renfree MB, Papenfuss AT, Deakin JE, Lindsay J, Heider T, Belov K, et al. Genome sequence of an Australian kangaroo, Macropus eugenii, provides insight into the evolution of mammalian reproduction and development. Genome Biology. 2011;12(8):R81
231.Hassanin A, Bonillo C, Nguyen BX, Cruaud C. Comparisons between mitochondrial genomes of domestic goat (Capra hircus) reveal the presence of numts and multiple sequencing errors. Mitochondrial DNA. 2010;21(3–4):68-76
232.Lowe TM, Eddy SR. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Research. 1997;25(5):955-964
233.Jarvis ED, Mirarab S, Aberer AJ, Li B, Houde P, Li C, et al. Whole-genome analyses resolve early branches in the tree of life of modern birds. Science. 2014;346(6215):1320-1331
234.Kashi Y, Hallerman E, Soller M. Marker-assisted selection of candidate bulls for progeny testing programmes. Animal Science. 1990;51(1):63-74
235.Teneva A. Molecular markers in animal genome analysis. Biotechnology in Animal Husbandry. 2009;25(5):1267-1284
236.Bai X, Yang T, Putz A, Wang Z, Li C, Fortin F, et al. Investigating the Genetic Architecture of Disease Resilience in Pigs by Genome-Wide Association Studies of Complete Blood Count Traits Collected from a Natural Disease Challenge Model. 2020.
237.Tong X, Hou L, He W, Mei C, Huang B, Zhang C, et al. Whole genome sequence analysis reveals genetic structure and X-chromosome haplotype structure in indigenous Chinese pigs. Scientific Reports. 2020;10(1):1-10
238.Venkatesh B, Kirkness EF, Loh Y-H, Halpern AL, Lee AP, Johnson J, et al. Survey sequencing and comparative analysis of the elephant shark (Callorhinchus milii) genome. PLoS biology. 2007;5(4):e101. DOI: 10.1371/journal.pbio.0050101
239.Zhang F, Wang Y, Mukiibi R, Chen L, Vinsky M, Plastow G, et al. Genetic architecture of quantitative traits in beef cattle revealed by genome wide association studies of imputed whole genome sequence variants: I: Feed efficiency and component traits. BMC Genomics. 2020;21(1):36
240.Lu Z, Yue Y, Yuan C, Liu J, Chen Z, Niu C, et al. Genome-wide association study of body weight traits in Chinese fine-wool sheep. Animals. 2020;10(1):170
241.Wang K, Kang Z, Jiang E, Yan H, Zhu H, Liu J, et al. Genetic effects of DSCAML1 identified in genome-wide association study revealing strong associations with litter size and semen quality in goat (Capra hircus). Theriogenology. 2020;146:20-25
242.van den Berg S, Vandenplas J, van Eeuwijk FA, Bouwman AC, Lopes MS, Veerkamp RF. Imputation to whole-genome sequence using multiple pig populations and its use in genome-wide association studies. Genetics, Selection, Evolution. 2019;51(1):1-13
243.Rubin C-J, Zody MC, Eriksson J, Meadows JR, Sherwood E, Webster MT, et al. Whole-genome resequencing reveals loci under selection during chicken domestication. Nature. 2010;464(7288):587-591
244.El-Sabrout K, Aggag S, Souza Jr JBF. Some recent applications of rabbit biotechnology–a review. Animal Biotechnology. 2020;31(1):76-80
245.Fariello M-I, Servin B, Tosser-Klopp G, Rupp R, Moreno C, San Cristobal M, et al. Selection signatures in worldwide sheep populations. PLoS One. 2014;9(8):e103813
246.Lee JJ, Wedow R, Okbay A, Kong E, Maghzian O, Zacher M, et al. Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nature Genetics. 2018;50(8):1112
247.Khanzadeh H, Ghavi Hossein-Zadeh N, Ghovvati S. Genome wide association studies, next generation sequencing and their application in animal breeding and genetics: A review. Iranian Journal of Applied Animal Science. 2020;10(3):395-404
248.Raza SHA, Khan S, Amjadi M, Abdelnour SA, Ohran H, Alanazi KM, et al. Genome-wide association studies reveal novel loci associated with carcass and body measures in beef cattle. Archives of Biochemistry and Biophysics. 2020;2020:108543
249.Wang Y, Niu Z, Zeng Z, Jiang Y, Jiang Y, Ding Y, et al. Using high-density SNP Array to reveal selection signatures related to prolificacy in Chinese and Kazakhstan sheep breeds. Animals. 2020;10(9):1633
250.Zhang W, Yang M, Zhou M, Wang Y, Wu X, Zhang X, et al. Identification of signatures of selection by whole-genome resequencing of a Chinese native pig. Frontiers in Genetics. 2020;11
251.Qanbari S, Simianer H. Mapping signatures of positive selection in the genome of livestock. Livestock Science. 2014;166:133-143
252.Li X, Su R, Wan W, Zhang W, Jiang H, Qiao X, et al. Identification of selection signals by large-scale whole-genome resequencing of cashmere goats. Scientific Reports. 2017;7(1):15142
253.Meuwissen T, Goddard M. Accurate prediction of genetic values for complex traits by whole-genome resequencing. Genetics. 2010;185(2):623-631
254.Fernando R, Grossman M. Marker assisted selection using best linear unbiased prediction. Genetics, Selection, Evolution. 1989;21(4):467
255.Georges M, Nielsen D, Mackinnon M, Mishra A, Okimoto R, Pasquino AT, et al. Mapping quantitative trait loci controlling milk production in dairy cattle by exploiting progeny testing. Genetics. 1995;139(2):907-920
256.Hayes B, Goddard M. Genome-wide association and genomic selection in animal breeding. Genome. 2010;53(11):876-883
257.Seidel G. Brief introduction to whole-genome selection in cattle using single nucleotide polymorphisms. Reproduction, Fertility, and Development. 2009;22(1):138-144
258.Phua S, Hyndman D, Baird H, Auvray B, McEwan J, Lee M, et al. Towards genomic selection for facial eczema disease tolerance in the N ew Z ealand sheep industry. Animal Genetics. 2014;45(4):559-564
259.Pickering NK, Auvray B, Dodds KG, McEwan JC. Genomic prediction and genome-wide association study for dagginess and host internal parasite resistance in New Zealand sheep. BMC Genomics. 2015;16(1):958
260.Daetwyler HD, Swan AA, van der Werf JH, Hayes BJ. Accuracy of pedigree and genomic predictions of carcass and novel meat quality traits in multi-breed sheep data assessed by cross-validation. Genetics, Selection, Evolution. 2012;44(1):33
261.Carillier C, Larroque H, Palhiere I, Clément V, Rupp R, Robert-Granié C. A first step toward genomic selection in the multi-breed French dairy goat population. Journal of Dairy Science. 2013;96(11):7294-7305
262.Larroque H, Robert-Granié C. Comparison of joint versus purebred genomic evaluation in the French multi-breed dairy goat population. Genetics Selection Evolution. 2014;2014:46
263.Daetwyler H, Hickey J, Henshall J, Dominik S, Gredler B, Van Der Werf J, et al. Accuracy of estimated genomic breeding values for wool and meat traits in a multi-breed sheep population. Animal Production Science. 2010;50(12):1004-1010
264.Daetwyler H, Kemper K, Van der Werf J, Hayes B. Components of the accuracy of genomic prediction in a multi-breed sheep population. Journal of Animal Science. 2012;90(10):3375-3384
265.Gianola D, Gustavo A, Hill WG, Manfredi E, Fernando RL. Additive genetic variability and the Bayesian alphabet. Genetics. 2009
266.Habier D, Fernando RL, Kizilkaya K, Garrick DJ. Extension of the Bayesian alphabet for genomic selection. BMC Bioinformatics. 2011;12(1):186
267.Van Marle-Köster E, Visser C, Berry D. A review of genomic selection-implications for the south African beef and dairy cattle industries. South African Journal of Animal Science. 2013;43(1):1-17
268.Yadav V, Singh N, Sharma S, Lakhani N, Bhimte A, Khare A, et al. Genomic Selection and it’s Application in Livestock Improvement. 2018
269.González-Recio O, Gianola D, Rosa GJ, Weigel KA, Kranis A. Genome-assisted prediction of a quantitative trait measured in parents and progeny: Application to food conversion rate in chickens. Genetics, Selection, Evolution. 2009;41(1):3
270.Saleh AA, Rashad AM, Hassanine NN, Sharaby MA. Candidate genes and signature of selection associated with different biological aspects and general characteristics of goat. Emerging Animal Species. 2022:100013
271.Moaeen-ud-Din M, Danish Muner R, Khan MS. Genome wide association study identifies novel candidate genes for growth and body conformation traits in goats. Scientific Reports. 2022;12(1):1-12
272.Salgado Pardo JI, Delgado Bermejo JV, González Ariza A, León Jurado JM, Marín Navas C, Iglesias Pastrana C, et al. Candidate genes and their expressions involved in the regulation of Milk and meat production and quality in goats (Capra hircus). Animals. 2022;12(8):988
Written By
Ahmed A. Saleh, Amr M.A. Rashad, Nada N.A.M. Hassanine, Mahmoud A. Sharaby and Sobhy M.A. Sallam
Submitted: 09 December 2022Reviewed: 31 December 2022Published: 08 March 2023
 Open access peer-reviewed chapter
Open access peer-reviewed chapter