Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
Use of UHPH to Sterilize Grape Juices and to Facilitate the Implantation of Saccharomyces and Other Emerging Fermentation Biotechnologies in Wines
Written By
Antonio Morata, Carlos Escott, Juan Manuel del Fresno, Buenaventura Guamis, Iris Loira, María Antonia Bañuelos, Carmen López, Felipe Palomero and Carmen González
Submitted: 19 October 2023Reviewed: 28 November 2023Published: 07 February 2024
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
Ultra-High Pressure Homogenization (UHPH) is an emerging non-thermal technology that uses continuous pumping at 200–600 MPa (generally 300), and then the fluid is depressurized against a special valve made with highly resistant alleys and coated with diamond powder. The UHPH process is able to sterilize grape juice or whatever food fluid and at the same time is extremely efficient in inactivating oxidative enzymes. The total elimination of wild microorganisms in grape juice allows the perfect implantation of the selected Saccharomyces cerevisiae starter facilitating the expression of particular metabolic profiles or the expression of specific extracellular activities. Furthermore, the use of complex emerging biotechnologies such as the use of co-inoculations with S. cerevisiae and lactic acid bacteria to perform simultaneously alcoholic and malolactic fermentations works much better if the initial juice has a very low load of wild microorganisms. Additionally, the development of spoilage processes by wild microorganisms that can remain in the wine after fermentation is practically impossible when the initial juice is treated by UHPH. Thus, UHPH helps to obtain wines more stable from a microbiological perspective and therefore safer and more hygienic.
MALTA CONSOLIDER TEAM Research Network (RED2022-134388-T), enotecUPM, ETSIAAB, Universidad Politécnica de Madrid, Madrid, Spain
Carlos Escott
MALTA CONSOLIDER TEAM Research Network (RED2022-134388-T), enotecUPM, ETSIAAB, Universidad Politécnica de Madrid, Madrid, Spain
Juan Manuel del Fresno
MALTA CONSOLIDER TEAM Research Network (RED2022-134388-T), enotecUPM, ETSIAAB, Universidad Politécnica de Madrid, Madrid, Spain
Buenaventura Guamis
Departament de Ciència Animal I Dels Aliments, Facultat de Veterinària, Centre d’Innovació, Recerca I Transferència en Tecnologia Dels Aliments (CIRTTA), TECNIO, XaRTA, Universitat Autònoma de Barcelona, Bellaterra, Spain
Iris Loira
MALTA CONSOLIDER TEAM Research Network (RED2022-134388-T), enotecUPM, ETSIAAB, Universidad Politécnica de Madrid, Madrid, Spain
María Antonia Bañuelos
MALTA CONSOLIDER TEAM Research Network (RED2022-134388-T), enotecUPM, ETSIAAB, Universidad Politécnica de Madrid, Madrid, Spain
Carmen López
MALTA CONSOLIDER TEAM Research Network (RED2022-134388-T), enotecUPM, ETSIAAB, Universidad Politécnica de Madrid, Madrid, Spain
Felipe Palomero
MALTA CONSOLIDER TEAM Research Network (RED2022-134388-T), enotecUPM, ETSIAAB, Universidad Politécnica de Madrid, Madrid, Spain
Carmen González
MALTA CONSOLIDER TEAM Research Network (RED2022-134388-T), enotecUPM, ETSIAAB, Universidad Politécnica de Madrid, Madrid, Spain
*Address all correspondence to: antonio.morata@upm.es
1. Introduction
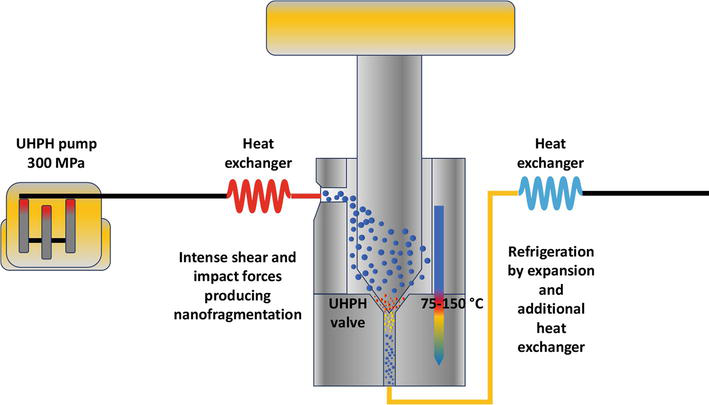
Ultra-High Pressure Homogenization (UHPH) is an emerging non-thermal technology with many interesting advantages in food processing, including: powerful antimicrobial effect, high inactivation of oxidative enzymes, nanofragmentation, formation of stable colloidal dispersions, nanocoating, and nanoencapsulation [1, 2, 3, 4]. The liquid must be pumped at ultra-high pressure (200–600 MPa, normally 300 MPa), reaching a speed of Mach 3 in the capillary pipeline upstream of the valve and a very high intense kinetic energy that is released against the valve with intense impact forces and shear stresses. The temperature at the valve can reach 70–80°C (Figure 1) when the fluid is at room temperature or refrigerated (4–20°C), or even much higher if the fluid is preheated. Downstream of the valve, subsequent depressurization leads to cooling of the liquid that can be also enhanced by the use of heat exchangers (Figure 1).
Figure 1.
Main components of the UHPH system (not at scale), including the design of the UHPH valve, the nanofragmentation process, and the temperature change of the colloidal particles.
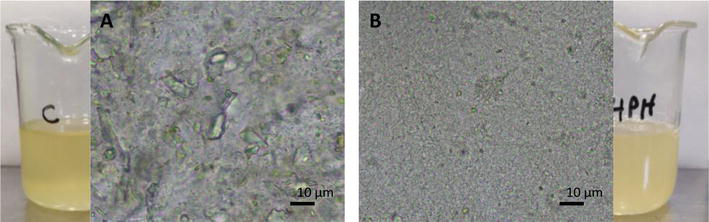
After passing through the valve, the colloidal particles in the fluid and the microorganisms are nanofragmented, resulting in inactivation of the microorganisms. It can be observed how the initial juice contains a lot of colloidal particles from the cell walls of the pulp but also tartrate crystals and wild microbial cells (Figure 2a). From optical microscopic observations, it can be seen that after UHPH treatment, the colloidal structure is thinner in the juice, which makes it highly stable and difficult to settle, giving the appearance of a haze in the beaker and of a small granulation under optical microscopy (Figure 2b). The colloidal particle size of a pressed juice is usually in the range of a few micrometers and after UHPH processing, is reduced to nanometric scale. We have measured by AFM and laser light scattering a particle size range of 0.8–3.2 μm which is decreased by nanofragmentation after UHPH to 235–744 nm [5, 6].
Figure 2.
Macroscopical appearance and optical microscopy of the colloids in a white grape juice Vitis vinifera L Verdejo variety (A) after pressing and settling, (B) processed by UHPH at 300 MPa.
Yeast counts of 6-log cfu/mL can be easily reached within a few hours in fresh grape must after grape pressing due to the growth and development of wild microorganisms and that population can be completely eliminated with a single pass through UHPH at 300 MPa [7]. Under the same conditions, 4-log bacteria and even spores can also be inactivated depending on the in-valve temperature [5, 7].
At the same time, the UHPH process is very gentle and protective of sensitive molecules with high impact on sensory quality, such as anthocyanins [6], terpenes [5], or thiols [8]. Even when high temperatures (75–150°C) can be reached in the valve, the amount of thermal markers as HMF [5] or furosine [9] is very low or not detectable. Furthermore, the formation of the carcinogenic molecule ethyl carbamate in sake after pasteurization and its absence after UHPH processing has recently been observed [10].
2. Antimicrobial effect and implantation of Saccharomyces
UHPH is a highly effective technology to eliminate vegetative cells of yeasts and bacteria working at 300 MPa. Yeasts are particularly sensitive to the effect of UHPH and can be easily controlled (Table 1), even considering very high populations that are abnormal in healthy grapes. The same degree of inactivation can be reached with vegetative cells of aerobic bacteria.
Effect of UHPH on the control of wild yeast and bacteria.
nd: not detected.
The degree of inactivation can be comparable to HTST (High Temperature Short Time) pasteurization when processed without thermal assistance at the valve, and similar to UHT (Ultra-High Temperature) sterilization, but with less thermal damage, when the temperature at the valve reaches 140–160°C to inactivate sporulated bacteria [1, 3]. A sporulated bacterium that causes significant problems in juices and especially in concentrated juices is Alicyclobacillus acidoterrestris, but it can be easily controlled by UHPH at 300 MPa with mild inlet temperatures of 80–90°C. This bacterium is aerobic, acidophilic, thermoresistant, capable of sporulating, and responsible for the production of off-flavors in juices, such as 2,6-dibromophenol and guaiacol.
The initial yeast population in wine grapes normally ranges between 3 and 4-log CFU/mL depending on maturity and environmental conditions. It consists mainly of low fermentative apiculate yeasts, such as Hanseniaspora spp., Metschnikowia spp., Candida spp., Pichia spp., Lachancea spp., Kluyveromyces, and Saccharomyces [11, 12, 13, 14], and other yeast-like saprophytic fungi, such as Aureobasidium pullulans. Saccharomyces cerevisiae is found in very low counts and is, therefore, hardly representative of the initial grape population, although, due to its peculiar physiology, it will predominate during alcoholic fermentation.
No clear resistance to UHPH processing has been observed among the different non-Saccharomyces species, all of them can be easily eliminated at populations ranging from 3 to 5-log CFU/mL (Table 1) [5, 6, 7]. Furthermore, we have observed an even higher inactivation in UHPH processing of yeast biomasses. Therefore, the use of UHPH is a powerful tool to eliminate initial wild yeast populations from grape juices and to facilitate the implantation of Saccharomyces starters inoculated at even lower doses that those recommended by yeast producers (often 20–40 g/Hl of active dry yeast). For those Saccharomyces strains with low fermentative power or slower fermentation kinetics, or non-Saccharomyces yeasts that are weak in terms of competition for nutrients and colonization of the medium, pre-treatment of the juice with UHPH greatly increases their chances of implantation, growth, and development of their metabolomic profile [15].
The selection of Saccharomyces cerevisiae strains looks for specific properties that can help to improve fermentation, to produce low volatile acidity and off-flavors (some sulfur compounds, volatile acidity, ethyl acetate, and others), to decrease alcoholic strength, or to develop suitable fermentations under stress conditions such as high sugar concentrations, low temperature, or limited yeast assimilable nitrogen [16, 17, 18]. Special features with impact on aroma, such as the production of acetate esters of higher alcohols or ethanol esters of medium chain fatty acids, the expression of beta-glucosidase enzymes to release volatile terpenes, or carbon lyase activities to release thiols, are also of interest [16, 17]. Similarly, specific properties with effect on color stability, such as the production of pyruvate and acetaldehyde to form stable vitisin pigments or the expression of hydroxy cinnamate decarboxylase activity to form vinylphenolic pyranoanthocyanins during fermentation or the formation of polymeric pigments, are also studied for the selection of Saccharomyces cerevisiae strains [17]. The influence of the yeast on the structure, body, and softening of tannins during fermentation is also considered by selecting appropriate strains that can produce more glycerol or release cell wall polysaccharides [16, 17, 18].
Currently, several non-Saccharomyces yeasts are widely used in enology to control pH by acidification with Lachancea thermotolerans [19] or deacidification with Schizosaccharomyces pombe [20] or japonicus [21], and to improve aroma, flavor, and structure with Torulaspora delbrueckii [22], Hanseniaspora spp. [23] or Metschnikowia pulcherrima [24]. All these non-Saccharomyces yeast species are usually less competitive than wild or inoculated Saccharomyces and their implantation and development are much better in juices processed by UHPH [15, 25]. Lachancea thermotolerans produces effective acidification during alcoholic fermentation by metabolizing sugars to lactic acid which can decrease the pH under enological conditions by up to 0.5 units and, at the same time, slightly reduce the alcohol content. Furthermore, it produces low volatile acidity and has positive effects on the formation of esters and the release of volatile thiols [19].
Schizosaccharomyces pombe and S. japonicus have been used for their ability to degrade malic acid to ethanol by the maloalcoholic fermentation and also have shown a good impact on wine palatability by releasing high amounts of cell wall polysaccharides [20, 21]. Some strains can reach 13–15% v/v ethanol and can be used for complete fermentation in the absence of S. cerevisiae [20]. Moreover, S. pombe has shown a high release of pyruvate, which influences the formation of vitisin A-type pigments [20]. Also, some strains have a positive effect on aroma, but the main problem can be the excessive production of volatile acidity [20]. Torulaspora delbrueckii was the first non-Saccharomyces produced at industrial level as dry yeast due to its positive impact on structure and aroma; juices fermented with this yeast species increase fruitiness and complexity [22]. Some specific markers of Torulaspora delbrueckii metabolism, such as 3-ethoxy propanol, have been reported. Hanseniaspora spp. are normally described as high producers of acetate esters and therefore with a significant impact on fruity and floral aroma perception. Some of them are especially potent in the formation of bencenoids, such as H. vineae which also has a high fermentative power and low volatile acidity formation [23]. This species also has a positive role in the release of free terpenes from non-volatile glycosides. Metschnikowia pulcherrima is another key non-Saccharomyces in wine biotechnology for its ability to improve the aromatic complexity, express enzymatic activities and its interesting effects on bioprotection [24]. Recently, its high compatibility with Lachancea thermotolerans and a synergistic effect on acidification have also been reported [26].
Most of these non-Saccharomyces species have low fermentative power ranging from 1 to 10% v/v ethanol [19, 22, 23, 24, 27], and are unable to finish alcoholic fermentation in still wines, with a probable alcohol ranging from 12 to 16% v/v ethanol. Therefore, they must be used together with S. cerevisiae to completely dry the musts and obtain wines without residual sugars. At industrial level, two typical strategies are used: sequential fermentations [28] or mixed fermentations (co-inoculations) [29, 30]. The first option facilitates the control of fermentation and metabolomic influence of the non-Saccharomyces species by just deciding the inoculation time of the Saccharomyces yeast strain. However, it requires two inoculations. The second one is easier to carry out at industrial level, because a single inoculation is enough, but the ratio of non-Saccharomyces/Saccharomyces populations in the inoculum must be carefully evaluated to obtain good implantation and suitable metabolomic expression of each yeast species. In all situations, the use of juice previously processed by UHPH increases the possibilities of achieving good implantation without the competitiveness of wild yeasts [5, 6, 7, 15].
Additionally, most of these non-Saccharomyces yeasts are sensitive or highly sensitive to SO2 [19, 22, 23, 24, 27]. Actually, in the past, and also now, sulfites are used to control the development of wild non-Saccharomyces yeasts, to facilitate fermentation by S. cerevisiae, and to control some parameters, such as volatile acidity or ethyl acetate that can decrease wine quality and can be promoted by some non-Saccharomyces yeasts. The use of UHPH avoids the deleterious effects of sulfites on non-Saccharomyces yeasts while improving their implantation during fermentation.
Currently, it is also frequent to develop alcoholic (AF) and malolactic (MLF) fermentations, at the same time to reduce the total fermentation time with safer fermentations [31, 32, 33, 34, 35] and often with higher contents of ethyl and acetate esters [31]. For this particular biotechnology, it is important to avoid unsuitable wild yeast and bacteria species and to achieve a good implantation of the inoculated bacteria-yeast starter. It is usually recommended to make yeast-bacteria co-inoculations on healthy grapes with pH equal or lower than 3.5 and with a low wild population, to avoid undesired developments of wild bacteria and the increase of volatile acidity and other metabolites that can behave as off-flavors [32].
Therefore, UHPH is a powerful tool to facilitate the use of emerging biotechnologies in wine fermentation. Nowadays, the use of more complex mixtures of microorganisms with special growth conditions and delicate fermentative metabolisms makes it more essential to eliminate competitive wild species to better implant these inoculants and favor the expression of their metabolisms to improve the sensory quality of the wine [26, 36, 37, 38, 39, 40, 41, 42]. In this framework, UHPH produces sterilized or pasteurized juices in which all the competitiveness can be avoided while preserving the sensory quality of the juice [5, 6, 7, 8].
4. Control of polyphenol oxidase enzymes, antioxidant capacity, and SO2 reduction
Oxidative enzymes are very detrimental to wine quality and rapidly affect color, reducing paleness in white wines and increasing browning in whites (Figure 3), rosés, and reds. The main oxidative enzymes in white wines are polyphenol oxidase enzymes (PPOs), among which the most important are tyrosinase in healthy grapes, and laccase in grapes affected by Botrytis cinerea. PPOs catalyze the oxidation of ortho-diphenols to quinones that polymerize to relatively insoluble brown melanoidins [43]. This browning decreases the color quality of most wines, usually related to evolution, oxidation, but also to the degradation of the fruitiness and varietal aroma.
Figure 3.
Effect on color of PPOs after a few hours of air exposition in the absence of antioxidants in a control (left) and in a juice processed by UHPH (right) at 300 MPa.
The protection of juice and wine from oxidation by PPOs is done by adding antioxidants such as SO2, or ascorbic acid. SO2 is highly effective, but currently under discussion due to its toxicity for the general public, but especially for sensitive people. SO2 has a double role: anti-oxidase by inhibiting the effect of PPOs and antioxidant by protecting ortho-diphenols from oxidation by PPOs [44]. Considering the potential toxicity of SO2, new molecules, such as glutathione (GSH), are being studied and used to control oxidation and browning in juices and wines [45, 46].
UHPH causes the fragmentation or unfold and denaturation of several enzymes by impact and shear efforts, including oxidative PPOs. In white varieties fermented in the absence of SO2, it can be observed how musts processed by UHPH remain pale and without browning even under high aeration conditions and with sensitive varieties, such as Muscat (Figure 3). We have also observed that oxidation continues after fermentation and if we clarify the wines with fining agents, the wines obtained from the non-processed juices continue to form brown compounds, but this does not occur with wines obtained from musts treated by UHPH. The effects on PPO inactivation, antioxidant activity, and anthocyanin contents are summarized (Table 2) for 3 grape varieties (2 whites and 1 red). UHPH control of PPO is very useful when it is desired to apply emerging biotechnologies, such as the use of non-Saccharomyces yeasts that are very sensitive to SO2. By using UHPH, it is possible to preserve the aromatic quality and prevent oxidation in musts of delicate aromatic white varieties, and at the same time, maintain suitable conditions for the implantation of non-Saccharomyces yeasts by the total elimination of wild yeasts and the absence of toxic sulfites.
Effect of UHPH on the control of PPOs and oxidation.
In addition to the improved sensory quality and stability of the juices processed by UHPH, higher antioxidant activity was observed, probably due to the inactivation of oxidative enzymes (Table 2). These effects made the UHPH juices healthier and with better nutraceutical properties. These characteristics are maintained in the subsequent wines after fermentation. The inactivation of enzymes by UHPH probably by unfold and denaturation could also have a positive effect on protein haze in white wines [5]. Therefore, UHPH musts can have fewer proteins able to produce turbidity or even the unfolding produced by the UHPH can facilitate the removal of unstable proteins by the use of proteases without the need for thermal pre-treatments.
UHPH is a powerful emerging non-thermal technology able to eliminate wild microorganisms facilitating the use of new biotechnologies with non-competitive non-Saccharomyces yeasts and non-Oenococcus lactic acid bacteria. At the same time, UHPH is very effective in controlling oxidative enzymes and can therefore be used to produce fermentations in the absence or with very low levels of SO2, what favors the development of sensitive yeasts such as many non-Saccharomyces species. Moreover, UHPH is a gentle technique with a protective effect on sensitive molecules, such as anthocyanins, terpenes, thiols, and others that are strongly affected by other conventional treatments. Additionally, UHPH can be considered an environmentally sustainable technique with low water and energy consumption.
This research was funded by MINISTERIO DE CIENCIA E INNOVACIÓN, project ENOINNOVAPRESS – PID2021-124250OB-I00 Ministerio de Ciencia e Innovación. This work was partially supported by the State Program to Promote Scientific-Technical Research and its Transfer of the Spanish Ministry of Science and Innovation through the MALTA CONSOLIDER TEAM Research Network (RED2022-134388-T).
1.Zamora A, Guamis B. Opportunities for ultra-high-pressure homogenization (UHPH) for the food industry. Food Engineering Reviews. 2015;7:130-142. DOI: 10.1007/s12393-014-9097-4
2.Patrignani F, Lanciotti R. Applications of high and ultra high pressure homogenization for food safety. Frontiers in Microbiology. 2016;7:1132. DOI: 10.3389/fmicb.2016.01132
3.Morata A, Guamis B. Use of UHPH to obtain juices with better nutritional quality and healthier wines with low levels of SO2. Frontiers in Nutrition. 2020;7:598286. DOI: 10.3389/fnut.2020.598286
4.Morata A, Loira I, Guamis B, Raso J, et al. Emerging technologies to increase extraction, control microorganisms, and reduce SO2. In: Cosme F, Nunes FM, Filipe-Ribeiro L, editors. Chemistry and Biochemistry of Winemaking, Wine Stabilization and Aging. London, UK: IntechOpen; 2021. DOI: 10.5772/intechopen.92035
5.Bañuelos MA, Loira I, Guamis B, Escott C, Del Fresno JM, Codina-Torrella I, et al. White wine processing by UHPH without SO2. Elimination of microbial populations and effect in oxidative enzymes, colloidal stability and sensory quality. Food Chemistry. 2020;332:127417. DOI: 10.1016/j.foodchem.2020.127417
6.Vaquero C, Escott C, Loira I, Guamis B del Fresno JM, et al. Cabernet sauvignon red must processing by UHPH to produce wine without SO2: The colloidal structure, microbial and oxidation control, colour protection and sensory quality of the wine. Food and Bioprocess Technology. 2022;15:620-634. DOI: 10.1007/s11947-022-02766-8
7.Loira I, Morata A, Bañuelos MA, Puig-Pujol A, Guamis B, González C, et al. Use of ultra-high pressure homogenization processing in winemaking: Control of microbial populations in grape musts and effects in sensory quality. Innovative Food Science and Emerging Technologies. 2018;50:50-56. DOI: 10.1016/j.ifset.2018.10.005
8.Morata A, Escott C, Vaquero C, del Fresno JM, Guamis B, Loira I, et al. Improving the implantation of non-Saccharomyces yeasts in winemaking by UHPH processing. Oral communication. In: Proceedings of the 44th World Congress of Vine and Wine; 5-9 June 2023; Cadiz-Jerez, Spain. BIO Web of Conferences. Vol. 68. Dijon, France: OIV; 2023. p. 02001
9.Ferragut V, Hernández-Herrero M, Veciana-Nogués MT, Borras-Suarez M, González-Linares J, Vidal-Carou MC, et al. Ultra-high-pressure homogenization (UHPH) system for producing high-quality vegetable-based beverages: Physicochemical, microbiological, nutritional and toxicological characteristics. Journal of the Science of Food and Agriculture. 2015;95:953-961. DOI: 10.1002/jsfa.6769
10.Arakawa G, Yokoi K. Application of multiple ultra-high-pressure homogenization to the pasteurization process of Japanese rice wine, sake. Journal of Bioscience and Bioengineering. 2023;136:117-122. DOI: 10.1016/j.jbiosc.2023.05.003
11.Cocolin L, Bisson LF, Mills DA. Direct profiling of the yeast dynamics in wine fermentations. FEMS Microbiology Letters. 2000;189(1):81-87. DOI: 10.1111/j.1574-6968.2000.tb09210.x
12.Mills DA, Johannsen EA, Cocolin L. Yeast diversity and persistence in Botrytis-affected wine fermentations. Applied and Environmental Microbiology. 2002;68(10):4884-4893. DOI: 10.1128/AEM.68.10.4884-4893.2002
13.Fleet GH. Wine yeasts for the future. FEMS Yeast Research. Nov 2008;8(7):979-995. DOI: 10.1111/j.1567-1364.2008.00427.x
14.Anagnostopoulos D, Kamilari E, Tsaltas D. Contribution of the microbiome as a tool for estimating Wine’s fermentation output and authentication. In: Morata A, Loira I, editors. Advances in Grape and Wine Biotechnology. London, UK: IntechOpen; 2019. DOI: 10.5772/intechopen.85692
15.Loira I, Escott C, del Fresno JM, Bañuelos MA, González C, Guamis B, et al. Chapter 5 – White must preservation by ultra-high pressure homogenization without SO2. In: Morata A, editor. White Wine Technology. London, UK: Academic Press; 2022. pp. 49-59. DOI: 10.1016/B978-0-12-823497-6.00014-4
16.Pretorius IS. Tailoring wine yeast for the new millennium: Novel approaches to the ancient art of winemaking. Yeast. 2000;16:675-729. DOI: 10.1002/1097-0061(20000615)16:8<675::AID-YEA585>3.0.CO;2-B
17.Suárez-Lepe JA, Morata A. New trends in yeast selection for winemaking. Trends in Food Science & Technology. 2012;23:39-50. DOI: 10.1016/j.tifs.2011.08.005
18.Loira I, Morata A, Bañuelos MA, Suárez-Lepe JA. Chapter 15 – Isolation, selection, and identification techniques for non-Saccharomyces yeasts of oenological interest. In: Grumezescu AM, Holban AM, editors. Biotechnological Progress and Beverage Consumption. London, UK: Academic Press; 2020. pp. 467-508. DOI: 10.1016/B978-0-12-816678-9.00015-1
19.Morata A, Loira I, Tesfaye W, Bañuelos MA, González C, Suárez Lepe JA. Lachancea thermotolerans applications in wine technology. Fermentation. 2018;4:53. DOI: 10.3390/fermentation4030053
20.Loira I, Morata A, Palomero F, González C, Suárez-Lepe JA. Schizosaccharomyces pombe: A promising biotechnology for modulating wine composition. Fermentation. 2018;4:70. DOI: 10.3390/fermentation4030070
21.Portaro L, Maioli F, Canuti V, Picchi M, Lencioni L, Mannazzu I, et al. Schizosaccharomyces japonicus/Saccharomyces cerevisiae mixed starter cultures: New perspectives for the improvement of Sangiovese aroma, taste, and color stability. LWT. 2022;156:113009. DOI: 10.1016/j.lwt.2021.113009
22.Ramírez M, Velázquez R. The yeast Torulaspora delbrueckii: An interesting but difficult-to-use tool for winemaking. Fermentation. 2018;4:94. DOI: 10.3390/fermentation4040094
23.Martin V, Valera MJ, Medina K, Boido E, Carrau F. Oenological impact of the Hanseniaspora/Kloeckera yeast genus on wines—A review. Fermentation. 2018;4:76. DOI: 10.3390/fermentation4030076
24.Morata A, Loira I, Escott C, del Fresno JM, Bañuelos MA, Suárez-Lepe JA. Applications of Metschnikowia pulcherrima in wine biotechnology. Fermentation. 2019;5:63. DOI: 10.3390/fermentation5030063
25.Morata A, Escott C, Bañuelos MA, Loira I, del Fresno JM, González C, et al. Contribution of non-Saccharomyces yeasts to wine freshness: A review. Biomolecules. 2020;10:34. DOI: 10.3390/biom10010034
26.Escott C, Vaquero C, Loira I, López C, González C, Morata A. Synergetic effect of Metschnikowia pulcherrima and Lachancea thermotolerans in acidification and aroma compounds in airén wines. Food. 2022;11:3734. DOI: 10.3390/foods11223734
27.Jolly NP, Varela C, Pretorius IS. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Research. 2014;14:215-237. DOI: 10.1111/1567-1364.12111
28.Gobbi M, Comitini F, Domizio P, Romani C, Lencioni L, Mannazzu I, et al. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiology. 2013;33:271-281. DOI: 10.1016/j.fm.2012.10.004
29.Ciani M, Comitini F, Mannazzu I, Domizio P. Controlled mixed culture fermentation: A new perspective on the use of non-Saccharomyces yeasts in winemaking. FEMS Yeast Research. 2010;10:123-133. DOI: 10.1111/j.1567-1364.2009.00579.x
30.Comitini F, Gobbi M, Domizio P, Romani C, Lencioni L, Mannazzu I, et al. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiology. 2011;28:873-882. DOI: 10.1016/j.fm.2010.12.001
31.Knoll C, Fritsch S, Schnell S, Grossmann M, Krieger-Weber S, du Toit M, et al. Impact of different malolactic fermentation inoculation scenarios on Riesling wine aroma. World Journal of Microbiology and Biotechnology. 2012;28:1143-1153. DOI: 10.1007/s11274-011-0917-x
32.Pardo I, Ferrer S. Chapter 7 - Yeast-bacteria coinoculation. In: Morata A, editor. Red Wine Technology. London, UK: Academic Press; 2019. pp. 99-114. DOI: 10.1016/B978-0-12-814399-5.00007-4
33.Krieger-Weber S. Application of yeast and bacteria as starter cultures. In: König H, Unden G, Fröhlich J, editors. Biology of Microorganisms on Grapes, in Must and in Wine. 2nd ed. Cham: Springer International Publishing; 2017. pp. 605-634. DOI: 10.1007/978-3-319-60021-5_25
34.Bartowsky EJ, Costello PJ, Chambers PJ. Emerging trends in the application of malolactic fermentation. Australian Journal of Grape and Wine Research. 2015;21:663-669. DOI: 10.1111/ajgw.12185
35.Krieger-Weber S, Heras JM, Suarez C. Lactobacillus plantarum, a new biological tool to control malolactic fermentation: A review and an outlook. Beverages. 2020;6:23. DOI: 10.3390/beverages6020023
36.Vaquero C, Loira I, Heras JM, Carrau F, González C, Morata A. Biocompatibility in ternary fermentations with Lachancea thermotolerans, other non-Saccharomyces and Saccharomyces cerevisiae to control pH and improve the sensory profile of wines from warm areas. Frontiers in Microbiology. 2021;12:656262. DOI: 10.3389/fmicb.2021.656262
37.Morata A, Arroyo T, Bañuelos MA, Blanco P, Briones A, Cantoral JM, et al. Wine yeast selection in the Iberian Peninsula: Saccharomyces and non-Saccharomyces as drivers of innovation in Spanish and Portuguese wine industries. Critical Reviews in Food Science and Nutrition. 2022;10:1-29. DOI: 10.1080/10408398.2022.2083574
38.Vaquero C, Escott C, Heras JM, Carrau F, Morata A. Co-inoculations of Lachancea thermotolerans with different Hanseniaspora spp.: Acidification, aroma, biocompatibility, and effects of nutrients in wine. Food Research International. 2022;161:111891. DOI: 10.1016/j.foodres.2022.111891
39.Del Fresno JM, Escott C, Loira I, Herbert-Pucheta JE, Schneider R, Carrau F, et al. Impact of Hanseniaspora vineae in alcoholic fermentation and ageing on lees of high-quality white wine. Fermentation. 2020;6:66. DOI: 10.3390/fermentation6030066
40.Vaquero C, Izquierdo-Cañas PM, Mena-Morales A, Marchante-Cuevas L, Heras JM, Morata A. Use of Lachancea thermotolerans for biological vs. chemical acidification at pilot-scale in white wines from warm areas. Fermentation. 2021;7:193. DOI: 10.3390/fermentation7030193
41.Del Fresno JM, Escott C, Loira I, Carrau F, Cuerda R, Schneider R, et al. The impact of Hanseniaspora vineae fermentation and ageing on lees on the terpenic aromatic profile of white wines of the Albillo variety. International Journal of Molecular Sciences. 2021;22:2195. DOI: 10.3390/ijms22042195
42.Vaquero C, Loira I, Bañuelos MA, Heras JM, Cuerda R, Morata A. Industrial performance of several Lachancea thermotolerans strains for pH control in white wines from warm areas. Microorganisms. 2020;8:830. DOI: 10.3390/microorganisms8060830
43.Martinez MV, Whitaker JR. The biochemistry and control of enzymatic browning. Trends in Food Science & Technology. 1995;6:195-200. DOI: 10.1016/S0924-2244(00)89054-8
44.Giacosa S, Río Segade S, Cagnasso E, Caudana A, Rolle L, Gerbi V. Chapter 21 – SO2 in wines: Rational use and possible alternatives. In: Morata A, editor. Red Wine Technology. London, UK: Academic Press; 2019. pp. 309-321. DOI: 10.1016/B978-0-12-814399-5.00021-9
45.Martínez J, González-Arenzana L. Chapter 3 – Use of glutathione in the winemaking of white grape varieties. In: Morata A, editor. White Wine Technology. London, UK: Academic Press; 2022. pp. 29-38. DOI: 10.1016/B978-0-12-823497-6.00021-1
46.Kilmartin P. Chapter 23 - Impacts of phenolics and prefermentation antioxidant additions on wine aroma. In: Morata A, editor. White Wine Technology. London, UK: Academic Press; 2022. pp. 301-312. DOI: 10.1016/B978-0-12-823497-6.00029-6
Written By
Antonio Morata, Carlos Escott, Juan Manuel del Fresno, Buenaventura Guamis, Iris Loira, María Antonia Bañuelos, Carmen López, Felipe Palomero and Carmen González
Submitted: 19 October 2023Reviewed: 28 November 2023Published: 07 February 2024
 Open access peer-reviewed chapter
Open access peer-reviewed chapter