Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
The connection between inflammation and cancer has been well recognized at the epidemiological, biological, and pharmacological levels. Unresolved chronic inflammation is implicated in most stages of cancer development and thus can induce certain solid tumors. The molecular regulators of these linkages are emerging and should be well-decorticated. Toll-like receptors (TLRs) recognize pathogen/microbe-associated molecular patterns (PAMPs/MAMPs) and death–associated molecular patterns (DAMPs) secreted from dying or damaged cells of the host. TLRs can be pro and anti-tumorigenic depending on the type of TLR signaling, cancer, and its stage. Therefore, comprehensive studies are required in this direction. The current chapter supplies a concise schematic concerning the biology and the characteristics of TLRs and summarizes the major findings of the enigmatic role of TLRs and their associated signaling in the pathogenesis of human cancers. On one hand and in some neoplastic contexts, TLR activation mediates proliferation invasion, migration and correlates with poor prognosis and metastasis, and inhibits apoptosis, leading to cancer progression. On the other hand and depending on other neoplastic context, TLRs agonists enhance radiosensitivity and chemotherapy, apoptosis, immune cell infiltration, and raise the antitumor effect of T cells.
HPV Unit Research, Laboratory of Molecular Epidemiology and Experimental Pathology Applied to Infectious Diseases, Pasteur Institute of Tunis, Tunis, Tunisia
Department of Human and Experimental Pathology, Pasteur Institute of Tunis, Tunis, Tunisia
Emna Ennaifer
HPV Unit Research, Laboratory of Molecular Epidemiology and Experimental Pathology Applied to Infectious Diseases, Pasteur Institute of Tunis, Tunis, Tunisia
Department of Human and Experimental Pathology, Pasteur Institute of Tunis, Tunis, Tunisia
Monia Ardhaoui
HPV Unit Research, Laboratory of Molecular Epidemiology and Experimental Pathology Applied to Infectious Diseases, Pasteur Institute of Tunis, Tunis, Tunisia
Department of Human and Experimental Pathology, Pasteur Institute of Tunis, Tunis, Tunisia
Haifa Tounsi
HPV Unit Research, Laboratory of Molecular Epidemiology and Experimental Pathology Applied to Infectious Diseases, Pasteur Institute of Tunis, Tunis, Tunisia
Department of Human and Experimental Pathology, Pasteur Institute of Tunis, Tunis, Tunisia
Rahima Bel Haj Rhouma
HPV Unit Research, Laboratory of Molecular Epidemiology and Experimental Pathology Applied to Infectious Diseases, Pasteur Institute of Tunis, Tunis, Tunisia
Samir Boubaker
Department of Human and Experimental Pathology, Pasteur Institute of Tunis, Tunis, Tunisia
*Address all correspondence to: emna.fehri@pasteur.utm.tn, emnafehri78@gmail.com
1. Introduction
The innate immune mechanisms are the first distinct defense fighting pathogen attack. They implicated many immune cell types such as monocytes, macrophages, dendritic cells (DCs), neutrophils, and natural killer (NK) cells [1]. They also comprise innate humoral components produced by the innate immune cells such as complement system, cytokines, chemokines, and antimicrobial peptides (AMPs; LL37 and Bactericidal/permeability-increasing protein (BPI), etc.) [2, 3, 4, 5]. These innate immune cells contain various intracellular or membrane-associated pattern recognition receptors (PRRs). The PRR family germline-encoded receptors include several forms of recognition receptors: nucleotide-binding oligomerization domain-like receptors (NOD)-like receptors and retinoic acid inducible gene I (RIG-I)-like receptors (RLR), C-type lectin receptors (CLR), Aim2- like receptors (ALR) and Toll-like receptors (TLRs) and intracellular DNA sensors such as cyclic GPM-AMP synthase cGAS [6, 7]. These receptors could discriminate between self and non-self-molecules. The well-known are Toll-like receptors (TLRs) to acknowledge highly conserved molecules expressed by pathogens: pathogen-associated molecular pattern (PAMP) or microbial-associated molecular pattern MAMPs (expressed/released by pathogens) or endogenous ligands released from dying, stressed and damaged cells: damaged-associated molecular pattern (DAMP) molecules released from dying cells Hsp60, Hsp70, fibronectin Host DNA from dying cells, mitochondrial DNA etc. [8, 9]. They produce inflammatory cytokines and type I interferons (IFNs) to ascertain a highly effective defense system [10]. TLRs are expressed not only by immune cells but also by epithelial cells, for defense against pathogens invading the body either through skin or mucous membranes.
Despite the host’s protective role against foreign molecules and fighting diseases, including cancer, chronic inflammation has been referred to as one of the new hallmarks of cancer since 2011 [11], besides the above settled in 2000 as proposed by Hanahan and Weinberg [11]. Further proof of the link between chronic inflammation and carcinogenesis is revealed by the diminished cancer rates in patients receiving non-steroidal anti-inflammatory drugs and elevated rate in obese patients exhibiting high adipose tissue inflammation [6]. Chronic inflammation activates constitutively signaling pathways, such as Nuclear factor kappa B (NF-kB) or mitogen-activated protein kinase (MAPK) whose pro-tumoral effect is well documented [12]. In hepatocellular carcinoma (HCC) [13] and colitis-associated cancer, the activation of NF-kB prevented tumor apoptosis and stimulated the production of pro-inflammatory cytokines in the tumor microenvironment, thereby enhancing tumor progression. In this context, NF-kB activation is implicated in both tumor initiation and progression in liver cancer [14]. Therefore, the inflammatory response can lead to carcinogenesis after NF-kB activation, by the induction of anti-apoptotic molecules [15, 16]. TLR stimulation enhances NF-kB activation and thus leads by consequence to pro-inflammatory cytokines and chemokines, growth factors and anti-apoptotic proteins release leading to tumor progression and chemoresistance. TLR overstimulation may bring about strong inflammation since it plays a part in the recruitment of inflammatory cells in the tumor microenvironment.
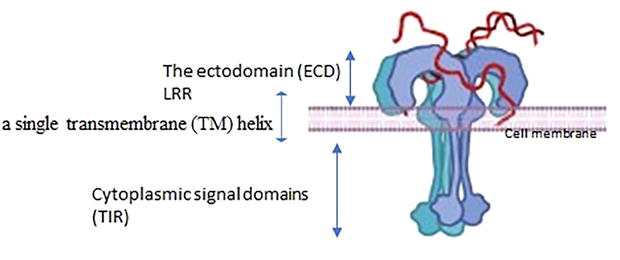
All TLRs, type I integral membrane receptors, have a characteristic structural organization with an extracellular recognition domain (ECD), a single transmembrane (TM) helix, and cytoplasmic signal domains (Figure 1) [17].
Figure 1.
Full-length TLR structure. The extracellular domain is composed of the N-terminal (LRRNT), LRRs and the C-terminal (LRR-CT) regions, a single transmembrane segment (TM), and an intracellular domain (TIR).
The ectodomain (ECD) is a terminal extracellular ligand binding domain made up of leucine-rich repeat (LRR), TLR-ECD shares a typically standard structural framework adopting horseshoe-shaped structures built from leucine-rich repeat (LRR) motif. The N-terminal extracellular ligand binding domain comprises varying numbers (19–26) hydrophobic leucine-rich repeat (LRR) modules (residues) every one of 20–30 Aa in length containing the consensus LxxLxLxx sequence in charge of ligand recognition and PAMPs and DAMPS recognition,” The LRR-NTs are disulfide linked β-hairpins, LRR-CTs are globular structures that have two α helices and are stabilized by two disulfide bonds [18, 19].
The intracellular C-terminal signaling domain of TLRs (150 Aa) is one of numerous evolutionarily conserved foundations of the immune system’ It’s named Toll IL-1 Receptor (TIR) domain since it shares homology with the signaling domain of IL-1R family members and is carboxyl-terminal to LRR [20]. TIR domain initiates downstream signaling cascades by interacting making use of its adaptor proteins such as myeloid differentiation response protein 88 (MyD88) that will be the absolute most commonly used adapter, MyD88 adaptor-like (MAL), TIR-domain-containing adapter-inducing interferon-β (TRIF), and TRIF-related adaptor molecule (TRAM). The signaling pathway activates transcription factors such as NF-κBs and interferon regulatory factors (IRFs), ultimately causing the release of proinflammatory cytokines and type I interferons, various anti-viral and anti-pathogen proteins, and initiation of the adaptive immune response [21].
Upon PAMPs or DAMPs sensing, TLRs undergo conformational changes, dimerize and reorient their TIR domains, to communicate with TIR-containing proteins, MyD88 and MAL, TRIF, and TRAM. This interaction results in undergoing a series of intracellular cascade signal transduction involving interleukin-1 receptor (IL-1R)-associated kinase 4 (IRAK4), IRAK2/1 phosphorylation resulting in the activation and dimerization of tumor necrosis factor receptor-associated factor 6 (TRAF6). TRIF interacts with TRAF3 and TRAF6 limited to TLR3 and TLR4. For endosomal TLR4, TRAM interplays with TRIF contributes to TRAF3-dependent stimulation of the kinase TBK1. These pathways promote inflammation and host defense and drive the IFN pathway activation with interferon-stimulated gene expression Table 1 [29, 30].
TLRs are classified into two subfamilies depending on their localization. TLRs localize on cell/surface membrane and/or reside within intracellular compartments such as endosomes, multivesicular bodied, lysosomes, and endolysosomes. TLR1, 2, 4, 5, and 6 are located in cell. However, TLR3, 7, 8, 9 are located on endosome membrane within the cell. Their different location is ultimately linked to the type of ligand they recognize. Thus, TLRs located on the cell membrane mainly bind microbial membrane lipids and proteins, and TLR4 binds bacterial lipopolysaccharides (LPS). TLR5 recognizes bacterial flagellin, TLR 2,1 and 6 recognize peptidoglycans and zymosan, whereas TLRs located on the endosomal membranes bind nucleic acids derived from virus and bacteria but also from self-nucleic acid. Thus, compartmentalization of nucleic acids sensing TLRs in the endolysosome is primordial to control their stimulation by self-derived nucleic acids and minimizes the risk of autoimmune reactions. TLR3 recognizes viral double stranded RNA (dsRNA), whereas TLR7 recognizes single-stranded RNA (ssRNA), TLR9 binds to bacterial and viral DNA (CpG-DNA motifs) [31, 32].
TLRs are expressed in varied cell types. Mainly they are located in innate immune cells Such as monocytes/macrophages, mast cells (MCs), neutrophils, eosinophils, basophils, natural killer (NK) cells, γδ T cells, innate lymphoid cells, DCs, platelets; brain innate immune cells like microglia and astrocytes. They are also expressed in adaptive immunity such as T and B cells. Non-immune cells, such as endothelial cells/ECs could express TLRs. TLR1 is detected in DCs and B Cells [33]. TLR2 is expressed in peripheral mononuclear leucocytes, DCs, monocytes, and T Cells [34, 35, 36]. TLR3 is mainly expressed in DCs, NK cells, and T cells [37, 38]. TLR4 in macrophages, DCs, and T Cells [36, 37, 39]. TLR5 in monocytes, DCs, and NK cells [37, 40]. TLR6 is highly expressed in B cells and DCs, but low in monocytes and NK cells [37, 41]. TLR7 is expressed in B cells, DCs, monocytes, and T cells [38, 42]. TLR8 is expressed in monocytes, DCs, and low in NK and T cells [43, 44]. TLR9 is present in DCs, B cells, macrophages, NK, and microglial cells [33, 43].
TLRs are expressed in certain types of cancers. Their expression could be closely associated with cell proliferation invasion and the probability of metastasis promoting then cancer progression or enhancing tumor regression. In this regard, the conclusions from in vitro or mouse model studies differ significantly (Table 2).
Cervical cancer is caused essentially by High-risk human papillomavirus (HR-HPV) types. A study highlighted a down expression of TLR3 and an overexpression of TLR1 at transcriptional level in dysplastic and carcinoma epithelium. in stroma, TLR 1, 2, 5, 6, and 9 were overexpressed in association with disease severity. Thus, TLR3 and TLR1 are proposed as implicated in early and late stages of cervical carcinogenesis thus their usefulness for diagnostic and prognostic. Moreover, the stromal overexpression of TLRs may play a crucial role in cervical cancer progression [45].
TLR4 was over-expressed in cervical cancer, and its activation by LPS can accelerate the proliferation and promote anti-apoptosis in Hela cells in vitro in a dose-dependent manner. The mechanism causing this overexpression is regulated by the NF-kB/IL-6/TGF-β1 secretion [46]. Patients with persistent HR HPV exhibit over-expressed TLR9 for more than 1 year in comparison to women who cleared HPV infection and to those re-infected with low-risk LR HPV. These results speculate that overexpression of TLR9 in persistently infected women could lead to chronic inflammation thus contributing to cervical cancer risk [47]. Furthermore, TLR9 may play a role in progression of cervical neoplasia in Tunisian patients and could therefore be considered a convenient biomarker for malignant transformation of cervical cells [48]. TLR8 may be an interesting therapeutic target in cervical cancer. In this regard, overexpression of TLR8 in cervical cancer patients and Hela cells was observed. Furthermore, a strong correlation with increased expression of TLR8, VEGF, and Bcl-2 in cervical cancer patients was demonstrated. Continually, upon binding to its agonist CL075, TLR8 is able to remarkably increase the percentage of cells in G2/M + S of Hela cells, accompanied by increased COX-2, BCL-2, and VEGF mRNA levels [49]. TLR2 and TLR4 are strongly associated with TNF-α and TNF-β in cervical cancer with gradually increased expression from premalignant lesions to cervical cancer compared to normal controls [50], However, higher TLR expression is associated with HPV16 clearance revealing an important link between innate and adaptive immunity in the control of HPV infections after a persistent period [51]. It is speculated that high levels of TLR2, TLR7, and potentially TLR8 in cervical mucosa are important for CIN2 regression, suggesting their role in the clearance of HPV [52]. These studies highlight the important protective role of the innate immune system at different stages of HPV infection. A deeper understanding of host immune response toward virus factors would affect if a lesion progresses or regresses. This finding is crucial to drive the proper choice of today’s available immunotherapeutic for HPV-associated disease. Down-expression of TLR4 during the progression of cervical neoplasia is linked to P (16INK4A) expression, the crucial marker of HPV integration into host cells. These results demonstrate the crucial link between HPV infection and TLR signaling during the carcinogenesis of cervical cancer [114]. Remarkable diminution of mRNA level of TLRs 3, 4, and 5 and overexpression of TLR1 was noted in cervical squamous cell carcinoma (CSCC) as compared to controls. These results point toward TLRs 3, 4, and 5 agonists exploration as therapeutic targets to treat cervical cancer [53]. The chimeric molecule including the extracellular domain of CD200 and a murine IgG2a Fc region CD200Fc seemed to repress TLR4/NF-κB and NLRP3 inflammasome inflammatory effects in SiHa cells and Caski cells treated with LPS. It provided a novel mechanistic understanding toward the conceivable therapeutic usefulness of CD200Fc for cervical cancer [115].
Regarding TLR gene polymorphism and cervical cancer association, several studies in several ethnicities have been conducted, but results vary greatly. In like manner, Oliveira and al concluded there was not always a noteworthy association between TLR9 polymorphism and HPV clearance or persistence. Accordingly, polymorphism in the promoter region of the TLR9 gene does not appear to be primordial in the natural history of the HPV infection [116]. These authors concluded the usefulness of TLR 3 and 9 gene polymorphisms in cervical cancer susceptibility in North India [117].
In the same context, a Tunisian study searching for TLR2 (−196 to −174 del) and the TLR 9 (2848 G > A) polymorphisms and the susceptibility of cervical cancer among Tunisian women show no association. Whereas, in the same cohort study, TLR3 (c.1377 C > T) and (Asp299Gly) TLR4 polymorphism has been shown to be associated with a higher risk of cervical cancer [118]. TLR9 (−1486 T/C, rs187084)-but not TLR9 (2848G/A, rs352140)-may be a threat factor for cervical cancer [119]. Furthermore, Chinese Han patients with TLR9 rs352140-GA + AA genotype and infected with HPV have the highest cervical cancer risk, compared to no HPV-infected patients holding the rs352140-GG genotype. Furthermore, the minor alleles of TLR2-rs3775290, TLR4-rs7873784, and TLR9-rs352140, and interaction with HPV infection were linked to a high cervical cancer risk in Chinese Han populations [120]. In the Indian population, TLR4 haplotype ACAC is highly linked to multiple HR-HPV infections. In otherwise, TLR9 SNPs rs187084, rs352140, and rs352139 were linked to diminished risk of high HPV16 viral load [121]. The study gene polymorphisms in cervical cancer susceptibility in North Indian pointed toward the implication of TLR 2 (−196 to −174 del) and TLR 4 (Thr399Ile) in cervical cancer susceptibility. The TLR gene polymorphisms, may be a good tool in setting out mechanisms related to innate immunity in cervical cancer susceptibility [122]. The later Indian study points toward considering TLR4 haplotype GCAG and TLR9 haplotype GATC as threat to hrHPV infection. Further evaluation of a larger sample size covering diverse ethnic populations globally is warranted [123].
5.2 Ovarian cancer
Ovarian cancer (OC) is the deadliest in gynecological cancer. TLR4 expression was linked to ovarian cancer progression, treatment resistance, and poor prognosis. Kelly et al. demonstrated that TLR4 is overexpressed in various ovarian epithelial tumors. The high TLR4 expression correlates with increased tumor progression and leads to chemo-resistance to Paclitaxel. Thus, blocking TLR4 is proposed to be beneficial to the OC patient if targeted specifically in cancer cells that overexpress the molecule [54].
In several types of OC cells, TLR4 inhibition in synergy with standard chemotherapy may be therapeutically beneficial to decrease drug resistance. In this regard, osteopontin (OPN), brought by LPS activation, leads to the proliferation and metastasis of ovarian cancer cells. These results pave the way to decorticate novel mechanistic pathways promoting cancer progression, invasion, and metastasis. Furthermore, TLR4 combined with OPN may be considered a promising target for ovarian cancer therapy [55]. In the same issue, the TLR4/MyD88/NF-κB axis is associated with the survival of patients with ovarian epithelial cancers (OECs). MyD88 was demonstrated to be an independent prognostic predictor in patients with OECs. The TLR4/MyD88 axis may be a possible mechanism for poor prognosis in patients with clear cell types of OEC in association with drug resistance [56]. Moreover, overexpression of TLR9 is correlated with tumor-grade severity and within poorly differentiated tumors. OC patients with metastatic disease had in their serum an over level of TLR9 ligand (hypo-methylated DNA). Furthermore, the TLR/PI3K signaling axis modulated the invasion and metastasis through the production of galectin 1, suggesting that inhibition of the p110, the PI3K isoform, is a promising therapeutic approach against metastatic ovarian cancer [57]. Protein aggregate magnesium-ammonium phospholinoleate-palmitoleate anhydride (P-MAPA) upregulated TLR2 and TLR4 signaling pathway. Thus, the combination of (P-MAPA) with IL-12 improves the antitumor immunoresponse, paving the way to a novel therapeutic approach for fighting OC [58]. Another way, the association with (P-MAPA) and cisplatin (CIS) is thought to be a promising target therapy against OC cells through TLR4 signaling pathways activation [59]. Stimulation of the TLR8 abolishes glucose metabolism in CD4+ Tregs through mTOR signaling reduction, thus reverting the immunosuppressive function of the previous cells in an OC cell growth microenvironment [60].
5.3 Endometrial cancer
Uterine cancer is the second most common gynecologic cancer worldwide (globocan, 2020). Endometrial cancer (EC), originating in the epithelium is about 90% of uterine cancers. TLR3 and TLR4 protein expression level was studied during the menstrual cycle and in postmenopausal endometrium taking into account different grades of cancer from grade 1 to grade 3 (peritoneal endometriosis, hyperplasia, and endometrial adenocarcinoma specimens). The lowest TLR expression levels were shown in poorly differentiated carcinoma (grade 3). These findings propose the implication of TLR3 and TLR4 in endometrial diseases as shown by changed expression levels in endometriosis and endometrial cancer [61]. EC and endometrial hyperplasia cells overexpress TLR2 and TLR6. In addition, the expression of TLR6 marked an advanced stage of EC [62]. Wojcik-Krowiranda et al. study demonstrated a significant correlation between the expression of TLR1, TLR2, TLR3, and TLR4 and VEGFR1, VEGFR2, VEGF-A, and HIF1α on human neoplastic endometrial samples at the clinical stage and pathological grading of EC [63]. Ashton et al. reported that different alleles of TLR9 polymorphisms (The rs5743836 and rs187084 alleles of TLR9 polymorphisms in Caucasian women) were protective against EC [124]. In contrast to previous thoughts, the overexpression of TLR showed that the use of TLR agonists could be harmful in some types and stages of cancers. Indeed, TLR agonists are not only not useful in the late stages of the tumor but also may cause tumor progression. Considering this hypothesis, scientists lead various types of research using natural or synthetic inhibitors of TLRs. In the case of EC, the results showed that the use of TLR inhibitors, such as TLR4 inhibitors, could inhibit chronic inflammation. However, clinical trial research should be conducted to prove the usefulness of this therapeutic approach in comparison to other approaches in EC [125].
5.4 Melanoma
Cutaneous melanoma is one of the foremost aggressive tumors and a life-threatening skin cancer. Within the setting of rising incidence and mortality, there is a necessity to discover new prognostic markers and early diagnosis is the big challenge to improve its prognosis. When searching their transcriptional and protein expression level, TLR2, 3, 4, 7, and 9 exhibit a high amount of expression in tumor tissue and melanoma cell lines. The results are more disparate at the protein level. TLR3 and 8 ex vivo are down-expressed in invaded LNs and however over expressed by melanoma cell lines extracted from the same LNs. Moreover, TLR2 and 4 proteins were absent in melanoma cell lines, whereas they were expressed in invaded LNs. This study gives evidence for a broad range of TLR expression in melanoma cells both ex vivo and in vitro. On one side, TLR2 and TLR4 are highly expressed in melanoma cells ex vivo and down expressed in vitro. On the other side, TLR3 and 8 expression are weak or absent ex vivo in invaded LNs however overexpressed in cell lines. However, more research should be conducted to reveal the functional and mechanistic aspects of these receptors on melanoma cell lines and therefore discover the therapeutic potential of targeting these receptors to treat melanoma [126]. TLR7/8 agonists are Resiquimod (RSQ), an imidazoquinolinamine derivative. The immune checkpoint blockage, Anti-PD-1 in association with RSQ can significantly extend the survival of melanoma-challenged mice, compared to untreated mice and mice treated with only anti-PD-1 [64]. TLR2/6 agonists +IFNƳ treatment synergically enhances CXCL10 production from melanoma cells when compared to IFNƳ alone. CXCL10 promotes the migration of CD4+ and CD8 + T cells, thus suggesting new therapeutic way of enhancing immune cell infiltration through CXCL10 production [127].
More recently, vaccination with MelQbG10, an innovative vaccine that integrates three components essential for successful immunotherapy consisting of virus-like nanoparticle (VLP), A-type CpG-ODN, and a Melan-A/MART-1 peptide, was tolerated and used safely in melanoma patients by the induction of tumor-specific CD8+ T cells after MelQbG10 vaccination [128]. To further improve immune responses and increase MelQbG10 immunogenicity, additional immune stimulatory agents, such as imiquimod, a TLR7 agonist lead to enhance Melan-A-specific T-cell frequencies. Furthermore, MelQbG10 can be considered safe and well tolerated when given in combination with imiquimod [129].
Immunization against a genetically engineered tumor-specific antigen, ovalbumin, when adjuvanted with Diprovocim, which targets the innate immune receptor TLR1/TLR2 in mice and humans, inhibited the growth of B16 melanoma and prolonged survival in the presence of immune checkpoint blockade by anti–PD-L1; all mice responded to treatment. These data propose Diprovocim improving the benefit of anti–PD-L1 therapy by raising the number and activation of tumor-specific CTLs able to reply to this checkpoint inhibitor [65].
Activation of PPARγ by its agonist, pioglitazone, reduces tumor volume through LPS/TLR4/MyD-88/Nf-kb1/TNF-α axis in melanoma tumor. Moreover, treatment of melanoma cells with pioglitazone has a beneficial protective effect against melanoma through LPS-TLR4-dependent signaling pathways inhibition [66]. TLR3 polymorphism L412F was linked to an elevated mitotic index. Whereas TLR4 D299G and T399I polymorphisms were associated with melanoma severity, nodal metastases, and advanced stage III and could be then potential markers targeting the survival and prognostic of melanoma patients. TLR4 T399I polymorphism was highly correlated with worse survival [130]. However, no association between TLR7 single nucleotide G pp. ln11Leu polymorphism and susceptibility to develop melanoma was found. Further, other studies on TLR polymorphisms and their susceptibility to malignant melanoma are needed [131].
5.5 Hepatocellular carcinoma
In 2020, approximately, 830,200 people died from liver cancer globally. Despite some progress, hepatocellular carcinoma (HCC) remains a major cause of death often detected at inoperable stage. In 80%, HCC occurs often in the setting of chronic liver disease and cirrhosis [132]. New biomarkers to identify patients who could benefit from more aggressive treatment are needed. HCC cells exhibit cytoplasmic and nuclear TLR2 expression which are associated with proliferative index, Caspase-3 expression, and vascularization. Furthermore, HCC shows notably nuclear TLR2 expression; however, hepatitis and cirrhosis cell patients demonstrate a predominant cytoplasmic expression. At a functional level, upon TLR2 activation by its agonist, HUH7, HCC expressed cellular proliferation and vascularization markers CD34 and VEGF. These results propose a plausible role for TLR-2 in HCC pathogenesis [67]. The interaction of mtDNA-HMGB1 and TLR9 might contribute to tumor growth signaling pathways in response to hypoxia in vivo and in vitro in hepatocellular carcinoma [68]. TLR5 is suggested to be an independent predictor of poor prognosis in HCC. However, TLR8 was not associated with patient survival [69].
Jing et al. detected positive TLR4 expression in 86% of cases of HCC patients. Other studies confirmed that LPS/TLR4/NF-κB signaling is involved in the invasion and metastasis of HCC patients [70]. In the same context, another study demonstrated that HCC patients whose tumors expressed high levels of both TLR4 and TLR9 had a poor prognosis. TLR4–MyD88 signaling pathway appears to be primordial for hepatocarcinogenesis. LPS-induced TLR4 signaling also promotes cancer cell survival and proliferation in HCC [71].
Activation of TLR4 could enhance the proliferation of HCC cells. Furthermore, TLR4/COX-2/PGE2/STAT3 axis is activated in HCC cells. Abolition of TLR4 or COX-2/PGE2/STAT3 pathway reduces LPS-induced inflammation and proliferation of HCC cells and improves the perceptivity of HCC cells to chemotherapeutics in vitro. These findings shed light on the precise molecular mechanism involved in the TLR4 signaling pathway leading to HCC progression and then pointing toward TLR4 as a promising target for HCC treatment [72].
To further investigate the expression and function of TLR2 in hepatocarcinoma proliferation recombinant plasmids expressing one of three forms of TLR2 siRNA (sh-TLR2 RNAi (A, B, and C) were transfected into BLE-7402. sh-TLR2 RNAi (B) had the most important knockdown effect. TLR2 abolition with sh-TLR2 RNAi (B) reduced cell proliferation and secretion of IL-6 and IL-8. Also, mice treated with sh-TLR2 RNAi (B) demonstrate a severe reduction in tumor volume, suggesting that TLR2 knockdown inhibits proliferation of cultured hepatocarcinoma cells and decreases the secretion of cytokines. Thus, TLR2 silencing could be proposed for siRNA-based gene therapy to treat of hepatocarcinoma patients [133]. Abolition of TLR2 by siRNA exhibited a decrease in proliferation, invasion, migration, and NF-κB/P65 expression but an increase in apoptotic ratio. In other way, rHMGB1 led to proliferation, invasion, and migration, enhanced NF-κB/P65 expression, and abolished cells apoptosis. Moreover, TLR2 reduced the role of rHMGB1. This finding proposes TLR2 and HMGB1 potential therapeutic targets against HCC [73].
Overexpression of miR-122 significantly diminished TLR4 expression in hepatoma cells. However, knocking down miR-122 overexpressed TLR4. It was found a putative miR-122 target in TLR4 3’UTR. Over-expression/down-expression of miR-122 could impact the proliferation and the expression of natural immune factors [134]. NADPH oxidase 4 (Nox4) enhanced LPS/TLR4/NF-ĸB/AP-1 signaling pathways in hepatocytes. Moreover, the effect of Nox4 abolition was time-dependent as proved in the mice model. Therefore, these data suggest LPS/TLR4/Nox4 axis is a tumor-promoting pathway as demonstrated both in human hepatoma cells and murine hepatocytes [135].
Genetic polymorphisms of TLRs are suggested to influence susceptibility to HCV infection and progression to end-stage liver disease. One haplotype (GCCCTTAG) of TLR4 was associated significantly with a decrease in the occurrence of hepatocellular carcinoma [136]. In another study, a strong association between allele C of rs3804099 of TLR2 and C allele of rs10116253 TLR4 and the risk of HCC was observed. Furthermore, A powerful link between allele T of rs1816702 of TLR2 and allele A of rs5030728 of TLR4 and the non-responder group was found. Haplotypes CAGT of TLR4 and ATAC of TLR2 showed significant association with CH and HCC groups in comparison to haplotype TGAC for TLR4 and haplotype GCGT for TLR2 [137].
5.6 Lung cancer
Lung cancer is a leading cause of cancer-related deaths with 18% of several deaths and 11.4% of new cases diagnosed in 2020 (Globocan, 2020). A better understanding of innate immunity and its related molecular mechanism in the lung could lead to the improvement and development of novel immunotherapy. TLR2 uses TLR1 and TLR6 [138]. TLR2 has been proposed to be a good target for lung cancer as the complex TLR1/2 promotes inhibition of tumor growth, decreases monocytic MDSC, and shifts macrophage toward M1 profile through JNK pathway [74]. However, the TLR2: TLR6 complex has been associated with promoting lung cancer metastasis [75]. TLR3 expression in lung cancer is a double-edged sword. Indeed, TLR3 activated by exosomal dsRN leads to chemokine liberation-inducing metastasis via SLIT2 gene [78]. However, an in vitro study demonstrates that activating TLR3 by its agonist could enhance apoptosis [76], and induce a protective immune response by activating DCs [76]. TLR4 activation in lung cancer exerts tumor progression by enhancing the immune escape through immunosuppressive cytokine liberation such as transforming growth factor beta (TGF-b), VEGF, and IL-8 [77]. and PDL1 [79]. TLR4 contributes to tumor regression through (calreticulin) CALR-TLR4-MyD88 axis promoting the maturation of DCs and boosting immunity [80]. TLR5 has a rather protective role in lung cancer, upon NSCLC cell treatment by flagellin (TLR5 agonists), inhibition of proliferation, migration, and invasion was observed [82]. TLR7/8 agonist, R848, combined with nanoemulsion (NE) exerts antitumor activity in lung cancer models through tumoral T cell activation [81]. In NSCLC cells, TLR7 induces inhibition of angiogenesis and survival [83]. However, on one side TLR9 activation by its agonist CpG, enhances lung tumor progression, proliferation, and metastasis [84]. On the other side, Vidutolimod (ODN-A), TLR9 agonist is used to treat patients with resistance to PD-1 blockade thus priming T-cell responses to fight lung cancer cells (clinic phase Ib trial) [85].
5.7 Breast cancer
Breast cancer remains a major cause of death in women in the developed world. Further to their expression on immune cells, TLRs are also expressed in tumor cells. Recent evidences hold up the connection between breast cancer and inflammation. TLR4 promotes breast cancer progression and metastasis. In vitro analysis demonstrates that MDA-MB-231 expresses TLR4 and abolishing this receptor induces a severe diminution of cell viability [86].
TLR2 and TLR9 activation in MDA-MB-231 cell lines promotes proinflammatory cytokines secretion leading to invasiveness and then metastasis. A high level of TLR9 was strongly associated with triple-negative subtypes. However, stimulation of TLR7 in combination with radiotherapy inhibits tumor growth, enhances T-cell memory, and is correlated with significant regression of spontaneous breast cancer in mice, suggesting a better prognosis [88]. TLR2 enhances breast cancer survival. Thus, preclinical in vivo breast cancer models demonstrate the efficiency of combining doxorubicin with CU-CPT22, the inhibitor of TLR2, to improve its anticancer effects and therefore attenuate breast cancer progression. This finding points toward considering TLR2 as a novel target therapy when associated with doxorubicin [87]. In vitro analysis showed that synthetic TLR3 agonist, polyI: C induces apoptosis either through caspase 3,8 and TRAIL pathway [89].
5.8 Colorectal cancer
Colorectal carcinoma (CC) is the third in cancer prevalence in both men and women and is a major leading cause of cancer morbidity and mortality worldwide. CC is among solid cancers in which inflammation is crucial in mechanistic oncogenesis. TLR4 is overexpressed in inflammatory-related colorectal cancer both in human and in vivo models. TLR4 induces colorectal through TLR4 high level of prostaglandin E2 and Cox-2 production. Inhibition of TLR4 by siRNA diminished tumor growth and abolished cell proliferation, migration, and invasion in colorectal cancer cell lines through suppressing ACAT1 expression [90]. A recent finding shows that TLR4 activity is inhibited through the direct binding of baicalein to TLR4 then abolishing HIF-1α and VEGF expressions and as a consequence inhibits CRC growth, angiogenesis, and metastasis [91]. Moreover, an in vitro study showed that Curcumae longae Rhizoma extract (CLRE) could oppose 5-Fu resistance in colon cancer by inhibiting TLR4/PI3K/AKT/mTORC1 pathway [92]. A synthetic agonist of TLR9, immune modulatory oligonucleotide (IMO), acts by impairing epidermal growth factor receptor (EGFR) expression and function and exhibited a synergistic antitumor effect with anti-EGFR antibody cetuximab or the tyrosin kinase inhibitor gefitinib in GEO human colon cancer xenografts, whereas it is ineffective in VEGF-overexpressing cetuximab-resistant GEO cetuximab-resistant (GEO-CR) tumors [93]. MGN1703, other synthetic TLR9 agonist show antitumor activity and III trial in colorectal cancer patients is conducted [94]. However, circulating cell-free DNA (cfDNA), when recognized by TLR9, could enhance colorectal cancer cell proliferation through TLR9/MyD88/IL8 signaling pathway [95]. Moreover, in vitro study shows that cellular migration and invasion is mediated through TLR9/MMP13suggesting inhibition of MMP13 as a therapeutic target to treat colorectal cancer [96]. In addition, chloroquine, an antagonist of TLR9, inhibited cell viability, proliferation, and migration of the CRC cell line HT29, the mechanism through this inhibition is downregulation of NF-κB, and Bcl-xl [97]. TLR2 agonist, Pam3Cys (P3C), exhibited cellular proliferation signaling in the human CRC cell line upon PI3K/Akt and NF-κB associated anti-apoptotic genes BCL2A1, WISP1, and BIRC3 suggesting a primordial role of TLR2 in colorectal carcinogenesis [99]. putative tumor suppressor, miR-143regulates negatively TLR2 pathway in colorectal cancer [139]. However, other studies using mice model of colitis cancer demonstrate a protective role of TLR2 in colitis against developing colorectal cancer [98]. TLR3 agonist polyI: C is able to boost colorectal cancer cell resistance to chemotherapy by promoting IFN-β production, which induces apoptosis and impairs proliferation in some cancer cells [100]. Furthermore, overexpression of TLR5 in colorectal tumor cells is linked to better prognosis in CRC patients [101].
5.9 Glioma
Gliomas are the deadliest type of brain tumor with a median survival time of 15 months. The current treatments are complex, offensive, and far from satisfactory. Thus, the urgent need for alternative efficient therapy. Chronic inflammation is largely implicated in glioma carcinogenesis and progression. TLR2 was found to be overexpressed in Glioma-associated microglia (GAM). This expression leads to tumor immune evasion through abolishing MCHII transcription via activation of MAPK/ERK1/2 signaling pathway thus altering the efficiency of T-cell–dependent tumor elimination in in vivo and in vitro models [103]. Moreover, in human and in vitro studies show that TLR2 promotes glioma carcinogenesis through autophagy [104]. Invasion of glioma stem cells (GSCs) is promoted via TLR2/MMP axis [105]. However previous studies focus on the crucial role of HMGB1/TLR2 signaling to enhance tumor regression [106]. TLR4 is implicated in the proliferation of glioma cells and inhibition of TLR4 by short hairpin (sh) RNA-induced abolition of cell proliferation and induction of apoptosis [107]. In vitro experiments suggested that the neurotrophic factor, prosaposin (PSAP) Highly expressed in glioma could lead to tumor progression through TLR4/NF-κB pathway [108]. Another study showed that impaired TLR4 function is associated with immune invasion at the site of glioma [109]. Parenteral treatment with the TLR7/8 agonist, resiquimod (R848), eventually induced complete tumor regression of CNS-1 glioblastoma tumors in Lewis rats [110]. The most crucial findings on TLR9-signaling in the pathogenesis of glioma are largely detailed in previous studies. TLR9 promotes brain cancer progression by inducing invasion-induced hypoxia via the matrix metalloproteinases (2, 9, and 13) [111]. However when combined with radiotherapy (RT), TLR9 agonist could enhance tumor regression through inhibition of angiogenesis [112], shift immune response to CD8 cytotoxic immunity or macrophage M1 [113, 140].
TLRs are involved in several carcinogenic processes to promote cancer progression in some kinds of tumors and some individuals by inducing cell proliferation, invasion, and metastasis. The mechanisms behind this implicate many hallmarks in the setting of tumor cells such as inhibition of apoptosis, and stimulation of angiogenesis. On the other side, TLRs are expressed mainly on macrophages and dendritic cells (DCs). Stimulation of TLRs fundamentally starts not only innate immune response but can also induce adaptive immune response boosting the immune system to fight cancer. In this respect, TLR ligands are used as immunostimulatory molecules as anticancer treatment in numerous preclinical and clinical studies. From both points of view, one should be aware when choosing for adjuvants to treat cancer patients. In related TLR clinical trials, the dual role of TLR should be considered and further mechanistic investigations on the dual roles of TLR in tumor biology are needed [140, 141, 142]. Thus, conducting a deeper understanding of each TLR on each cancer is of crucial need. It is worth knowing whether to design a TLR inducer or inhibitor when considering TLR-targeted therapeutic. Furthermore, with the era of novel pharmacology and personalized medicine, the use of nanotechnology in combination with conventional chemotherapy and radiotherapy is of great interest as it impacts efficiently cancer treatment trials.
We thank all the members of the human and experimental pathology department at the Institut Pasteur de Tunis. This work was supported by the Tunisian Ministry of Public Health and the Tunisian Ministry of Higher Education and Scientific Research (LR16IPT04 and CIC2016IPT02).
1.Janeway CA Jr, Medzhitov R. Innate immune recognition. Annual Review of Immunology. 2002;20:197-216. DOI: 10.1146/annurev.immunol.20.083001.084359
2.Morgan BP, Harris CL. Complement, a target for therapy in inflammatory and degenerative diseases. Nature Reviews. Drug Discovery. 2015;14(12):857-877. DOI: 10.1038/nrd4657. Epub 2015 Oct 23
3.Pasupuleti M, Schmidtchen A, Malmsten M. Antimicrobial peptides: Key components of the innate immune system. Critical Reviews in Biotechnology. 2012;32(2):143-171. DOI: 10.3109/07388551.2011.594423. Epub 2011 Nov 11
4.Wiesner J, Vilcinskas A. Antimicrobial peptides: The ancient arm of the human immune system. Virulence. 2010;1(5):440-464. DOI: 10.4161/viru.1.5.12983
5.Wagner E, Frank M. Therapeutic potential of complement modulation. Nature Reviews. Drug Discovery. 2010;9:43-56. DOI: 10.1038/nrd3011
6.Kawai T, Akira S. The roles of TLRs, RLRs and NLRs in pathogen recognition. International Immunology. 2009;21(4):317-337. DOI: 10.1093/intimm/dxp017. Epub 2009 Feb 26
7.Kumar V. The trinity of cGAS, TLR9, and ALRs guardians of the cellular galaxy against host-derived self-DNA. Frontiers in Immunology. 2021;11:624597. DOI: 10.3389/fimmu.2020.624597
8.Land WG. Prologue: About DAMPs, PAMPs, and MAMPs. In: Damage-Associated Molecular Patterns in Human Diseases. Cham: Springer; 2018. DOI: 10.1007/978-3-319-78655-1_11
9.Newton K, Dixit VM. Signaling in innate immunity and inflammation. Cold Spring Harbor Perspectives in Biology. 2012;4(3):a006049. DOI: 10.1101/
10.Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124(4):783-801. DOI: 10.1016/j.cell.2006.02.015
11.Hanahan D, Weinberg RA. Hallmarks of cancer: The next generation. Cell. 2011;144(5):646-674. DOI: 10.1016/j.cell.2011.02.013
12.Trinchieri G. Cancer and inflammation: An old intuition with rapidly evolving new concepts. Annual Review of Immunology. 2012;30:677-706. DOI: 10.1146/annurev-immunol-020711-075008
14.Pikarsky E, Porat RM, Stein I, Abramovitch R, Amit S, Kasem S, et al. NF-kappaB functions as a tumour promoter in inflammation-associated cancer. Nature. 2004;431(7007):461-466. DOI: 10.1038/nature02924
15.Coussens LM, Werb Z. Inflammation and cancer. Nature. 2002;420(6917):860-867. DOI: 10.1038/nature01322
16.Karin M. Nuclear factor-kappaB in cancer development and progression. Nature. 2006;441(7092):431-436. DOI: 10.1038/nature04870
17.Asami J, Shimizu T. Structural and functional understanding of the toll-like receptors. Protein Science. 2021;30(4):761-772. DOI: 10.1002/pro.4043
18.Botos I, Segal DM, Davies DR. The structural biology of toll-like receptors. Structure. 2011;19(4):447-459. DOI: 10.1016/j.str.2011.02.004
19.He XL, Bazan JF, McDermott G, Park JB, Wang K, Tessier-Lavigne M, et al. Structure of the Nogo receptor ectodomain: A recognition module implicated in myelin inhibition. Neuron. 2003;38:177-185
20.O’Neill LA, Bowie AG. The family of five: TIR-domain-containing adaptors in toll-like receptor signalling. Nature Reviews. Immunology. 2007;7(5):353-364. DOI: 10.1038/nri2079
21.Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: Update on toll-like receptors. Nature Immunology. 2010;11(5):373-384. DOI: 10.1038/ni.1863. Epub 2010 Apr 20
22.Fleer A, Krediet TG. Innate immunity: Toll-like receptors and some more. A brief history, basic organization and relevance for the human newborn. Neonatology. 2007;92(3):145-157. DOI: 10.1159/000102054. Epub 2007 Apr 27
24.Tartey S, Takeuchi O. Pathogen recognition and toll-like receptor targeted therapeutics in innate immune cells. International Reviews of Immunology. 2017;36(2):57-73. DOI: 10.1080/08830185.2016.1261318. Epub 2017 Jan 6
25.De Leo MG, Staiano L, Vicinanza M, Luciani A, Carissimo A, Mutarelli M, et al. Autophagosome-lysosome fusion triggers a lysosomal response mediated by TLR9 and controlled by OCRL. Nature Cell Biology. 2016;18(8):839-850. DOI: 10.1038/ncb3386. Epub 2016 Jul 11
26.Henrick BM, Yao XD, Zahoor MA, Abimiku A, Osawe S, Rosenthal KL. TLR10 senses HIV-1 proteins and significantly enhances HIV-1 infection. Frontiers in Immunology. 2019;10:482. DOI: 10.3389/fimmu.2019.00482
27.Fore F, Indriputri C, Mamutse J, Nugraha J. TLR10 and its unique anti-inflammatory properties and potential use as a target in therapeutics. Immune Network. 2020;20(3):e21. DOI: 10.4110/in.2020.20.e21
28.Kumar V. Toll-like receptors in sepsis-associated cytokine storm and their endogenous negative regulators as future immunomodulatory targets. International Immunopharmacology. 2020;89(Pt B):107087. DOI: 10.1016/j.intimp.2020.107087. Epub 2020 Oct 12
29.Takeda K, Akira S. TLR signaling pathways. Seminars in Immunology. 2004;16(1):3-9. DOI: 10.1016/j.smim.2003.10.003
30.Kawai T, Akira S. TLR signaling. Seminars in Immunology. 2007;19(1):24-32. DOI: 10.1016/j.smim.2006.12.004. Epub 2007 Feb 1
31.O’Neill LA, Golenbock D, Bowie AG. The history of toll-like receptors - redefining innate immunity. Nature Reviews. Immunology. 2013;13(6):453-460. DOI: 10.1038/nri3446. Epub 2013 May 17
32.Hamonic G, Pasternak JA, Wilson HL. Recognizing conserved non-canonical localization patterns of toll-like receptors in tissues and across species. Cell and Tissue Research. 2018;372(1):1-11. DOI: 10.1007/s00441-017-2767-9
33.Kawai T, Akira S. TLR signaling. Cell Death and Differentiation. 2006;13(5):816-825. DOI: 10.1038/sj.cdd.4401850
34.Sutmuller RP, den Brok MH, Kramer M, Bennink EJ, Toonen LW, Kullberg BJ, et al. Toll-like receptor 2 controls expansion and function of regulatory T cells. The Journal of Clinical Investigation. 2006;116(2):485-494. DOI: 10.1172/JCI25439
35.Patel M, Xu D, Kewin P, Choo-Kang B, McSharry C, Thomson NC, et al. TLR2 agonist ameliorates established allergic airway inflammation by promoting Th1 response and not via regulatory T cells. Journal of Immunology. 2005;174(12):7558-7563. DOI: 10.4049/jimmunol.174.12.7558
36.Murphy TJ, Ni Choileain N, Zang Y, Mannick JA, Lederer JA. CD4+CD25+ regulatory T cells control innate immune reactivity after injury. Journal of Immunology. 2005;174(5):2957-2963. DOI: 10.4049/jimmunol.174.5.2957
37.Bellocchio S, Moretti S, Perruccio K, Fallarino F, Bozza S, Montagnoli C, et al. TLRs govern neutrophil activity in aspergillosis. Journal of Immunology. 2004;173(12):7406-7415. DOI: 10.4049/jimmunol.173.12.7406
38.Nishiya T, Kajita E, Miwa S, Defranco AL. TLR3 and TLR7 are targeted to the same intracellular compartments by distinct regulatory elements. The Journal of Biological Chemistry. 2005;280(44):37107-37117. DOI: 10.1074/jbc.M504951200
39.Fitzgerald KA, McWhirter SM, Faia KL, Rowe DC, Latz E, Golenbock DT, et al. IKKepsilon and TBK1 are essential components of the IRF3 signaling pathway. Nature Immunology. 2003;4(5):491-496. DOI: 10.1038/ni921
40.Crellin NK, Garcia RV, Hadisfar O, Allan SE, Steiner TS, Levings MK. Human CD4+ T cells express TLR5 and its ligand flagellin enhances the suppressive capacity and expression of FOXP3 in CD4+CD25+ T regulatory cells. Journal of Immunology. 2005;175(12):8051-8059. DOI: 10.4049/jimmunol.175.12.8051
41.Wang RF. Regulatory T cells and toll-like receptors in cancer therapy. Cancer Research. 2006;66(10):4987-4990. DOI: 10.1158/0008-5472.CAN-05-4676
42.Lee J, Wu CC, Lee KJ, Chuang TH, Katakura K, Liu YT, et al. Activation of anti-hepatitis C virus responses via toll-like receptor 7. Proceedings of the National Academy of Sciences of the United States of America. 2006;103(6):1828-1833. DOI: 10.1073/pnas.0510801103
43.Sun CM, Deriaud E, Leclerc C, Lo-Man R. Upon TLR9 signaling, CD5+ B cells control the IL-12-dependent Th1-priming capacity of neonatal DCs. Immunity. 2005;22(4):467-477. DOI: 10.1016/j.immuni.2005.02.008
44.Peng G, Guo Z, Kiniwa Y, Voo KS, Peng W, Fu T, et al. Toll-like receptor 8-mediated reversal of CD4+ regulatory T cell function. Science. 2005;309(5739):1380-1384. DOI: 10.1126/science.1113401
45.DeCarlo CA, Rosa B, Jackson R, Niccoli S, Escott NG, Zehbe I. Toll-like receptor transcriptome in the HPV-positive cervical cancer microenvironment. Clinical & Developmental Immunology. 2012;2012:785825. DOI: 10.1155/2012/785825. Epub 2011 Oct 13
46.Cheng YX, Qi XY, Huang JL, Hu M, Zhou LM, Li BS, et al. Toll-like receptor 4 signaling promotes the immunosuppressive cytokine production of human cervical cancer. European Journal of Gynaecological Oncology. 2012;33(3):291-294
47.Cannella F, Pierangeli A, Scagnolari C, Cacciotti G, Tranquilli G, Stentella P, et al. TLR9 is expressed in human papillomavirus-positive cervical cells and is overexpressed in persistent infections. Immunobiology. 2015;220(3):363-368. DOI: 10.1016/j.imbio.2014.10.012. Epub 2014 Oct 23
48.Fehri E, Ennaifer E, Ardhaoui M, Ouerhani K, Laassili T, Bel Haj Rhouma R, et al. Expression of toll-like receptor 9 increases with progression of cervical neoplasia in Tunisian women--a comparative analysis of condyloma, cervical intraepithelial neoplasia and invasive carcinoma. Asian Pacific Journal of Cancer Prevention. 2014;15(15):6145-6150. DOI: 10.7314/apjcp.2014.15.15.6145
49.Zhang Y, Yang H, Barnie PA, Yang P, Su Z, Chen J, et al. The expression of toll-like receptor 8 and its relationship with VEGF and Bcl-2 in cervical cancer. International Journal of Medical Sciences. 2014;11(6):608-613. DOI: 10.7150/ijms.8428
50.de Matos LG, Cândido EB, Vidigal PV, Bordoni PH, Lamaita RM, Carneiro MM, et al. Association between toll-like receptor and tumor necrosis factor immunological pathways in uterine cervical neoplasms. Tumori. 2017;103(1):81-86. DOI: 10.5301/tj.5000576. Epub 2016 Nov 15
51.Scott ME, Ma Y, Farhat S, Moscicki AB. Expression of nucleic acid-sensing toll-like receptors predicts HPV16 clearance associated with an E6-directed cell-mediated response. International Journal of Cancer. 2015;136(10):2402-2408. DOI: 10.1002/ijc.29283. Epub 2014 Oct 30
52.Halec G, Scott ME, Farhat S, Darragh TM, Moscicki AB. Toll-like receptors: Important immune checkpoints in the regression of cervical intra-epithelial neoplasia 2. International Journal of Cancer. 2018;143(11):2884-2891. DOI: 10.1002/ijc.31814. Epub 2018 Oct 9
53.Aggarwal R, Misra S, Guleria C, Suri V, Mangat N, Sharma M, et al. Characterization of toll-like receptor transcriptome in squamous cell carcinoma of cervix: A case-control study. Gynecologic Oncology. 2015;138(2):358-362. DOI: 10.1016/j.ygyno.2015.05.029. Epub 2015 May 27
54.Kelly MG, Alvero AB, Chen R, Silasi DA, Abrahams VM, Chan S, et al. TLR-4 signaling promotes tumor growth and paclitaxel chemoresistance in ovarian cancer. Cancer Research. 2006;66(7):3859-3868. DOI: 10.1158/0008-5472.CAN-05-3948
55.Xu C, Li H, Yin M, Yang T, An L, Yang G. Osteopontin is involved in TLR4 pathway contributing to ovarian cancer cell proliferation and metastasis. Oncotarget. 2017;8(58):98394-98404. DOI: 10.18632/oncotarget.21844
56.Kim KH, Jo MS, Suh DS, Yoon MS, Shin DH, Lee JH, et al. Expression and significance of the TLR4/MyD88 signaling pathway in ovarian epithelial cancers. World Journal of Surgical Oncology. 2012;10:193. DOI: 10.1186/1477-7819-10-193
57.Park GB, Chung YH, Kim D. Induction of galectin-1 by TLR-dependent PI3K activation enhances epithelial-mesenchymal transition of metastatic ovarian cancer cells. Oncology Reports. 2017;37(5):3137-3145. DOI: 10.3892/or.2017.5533
58.Silveira HS, Lupi LA, Romagnoli GG, Kaneno R, da Silva NI, Fávaro WJ, et al. P-MAPA activates TLR2 and TLR4 signaling while its combination with IL-12 stimulates CD4+ and CD8+ effector T cells in ovarian cancer. Life Sciences. 2020;254:117786. DOI: 10.1016/j.lfs.2020.117786
59.de Almeida Chuffa LG, de Moura FG, Lupi LA, da Silva NI, Fávaro WJ. P-MAPA immunotherapy potentiates the effect of cisplatin on serous ovarian carcinoma through targeting TLR4 signaling. Journal of Ovarian Research. 2018;11(1):8. DOI: 10.1186/s13048-018-0380-5
60.Wu M, Fu X, Xu R, Liu S, Li R, Xu J, et al. Glucose metabolism and function of CD4+ Tregs are regulated by the TLR8/mTOR signal in an environment of SKOV3 cell growth. Cancer Medicine. 2023;12(15):16310-16322. DOI: 10.1002/cam4.6247
61.Allhorn S, Böing C, Koch AA, Kimmig R, Gashaw I. TLR3 and TLR4 expression in healthy and diseased human endometrium. Reproductive Biology and Endocrinology. 2008;6:40. DOI: 10.1186/1477-7827-6-40
62.Gençoğlu Bakbak BB, Ilhan TT, Pekin A, Kerimoğlu ÖS, Yılmaz SA, Kebapçılar A, et al. Evaluation of toll-like receptor expression with clinicopathologic variables in endometrium cancer. Sisli Etfal Hastan Tip Bulletin. 2018;52(3):196-200
63.Wojcik-Krowiranda KM, Forma E, Bienkiewicz A, Cwonda L, Wronska-Stefaniak J, Brys M. TLR family gene expression in relation to the HIF1α and the VEGFR pathway activation in endometrial cancer. Ginekologia Polska. 2020;91(8):439-446
64.Tambunlertchai S, Geary SM, Naguib YW, Salem AK. Anti-melanoma effects of resiquimod (RSQ) in vitro and in combination with immune checkpoint blockade in vivo. The AAPS Journal. 2023;25(4):57. DOI: 10.1208/s12248-023-00824-3
65.Wang Y, Su L, Morin MD, Jones BT, Mifune Y, Shi H, et al. Adjuvant effect of the novel TLR1/TLR2 agonist diprovocim synergizes with anti-PD-L1 to eliminate melanoma in mice. Proceedings of the National Academy of Sciences of the United States of America. 2018;115(37):E8698-E8706. DOI: 10.1073/pnas.1809232115
66.Dana N, Vaseghi G, Haghjooy JS. PPAR γ agonist, pioglitazone, suppresses melanoma cancer in mice by inhibiting TLR4 signaling. Journal of Pharmaceutical Sciences. 2019;22(1):418-423. DOI: 10.18433/jpps30626
67.Mohamed FEA, Hammad S, Luong TV, Dewidar B, Al-Jehani R, Davies N, et al. Expression of TLR-2 in hepatocellular carcinoma is associated with tumour proliferation, angiogenesis and Caspase-3 expression. Pathology, Research and Practice. 2020;216(8):152980. DOI: 10.1016/j.prp.2020.152980
68.Liu Y, Yan W, Tohme S, Chen M, Fu Y, Tian D, et al. Hypoxia induced HMGB1 and mitochondrial DNA interactions mediate tumor growth in hepatocellular carcinoma through toll-like receptor 9. Journal of Hepatology. 2015;63(1):114-121. DOI: 10.1016/j.jhep.2015.02.009
69.Kairaluoma V, Kemi N, Huhta H, Pohjanen VM, Helminen O. Toll-like receptor 5 and 8 in hepatocellular carcinoma. APMIS. 2021;129(8):470-479. DOI: 10.1111/apm.13142
70.Jing YY, Han ZP, Sun K, et al. Toll-like receptor 4 signaling promotes epithelial-mesenchymal transition in human hepatocellular carcinoma induced by lipopolysaccharide. BMC Medicine. 2012;10:98
71.Wang L, Zhu R, Huang Z, Li H, Zhu H. Lipopolysaccharide induced toll-like receptor 4 signaling in cancer cells promotes cell survival and proliferation in hepatocellular carcinoma. Digestive Diseases and Sciences. 2013;58(8):2223-2236
72.Lin A, Wang G, Zhao H, Zhang Y, Han Q , Zhang C, et al. TLR4 signaling promotes a COX-2/PGE2/STAT3 positive feedback loop in hepatocellular carcinoma (HCC) cells. Oncoimmunology. 2015;5(2):e1074376. DOI: 10.1080/2162402X.2015.1074376
73.Shi W, Su L, Li Q , Sun L, Lv J, Li J, et al. Suppression of toll-like receptor 2 expression inhibits the bioactivity of human hepatocellular carcinoma. Tumour Biology. 2014;35(10):9627-9637. DOI: 10.1007/s13277-014-2268-3
74.Deng Y, Yang J, Qian J, Liu R, Huang E, Wang Y, et al. TLR1/TLR2 signaling blocks the suppression of monocytic myeloid-derived suppressor cell by promoting its differentiation into M1-type macrophage. Molecular Immunology. 2019;112:266-273. DOI: 10.1016/j.molimm.2019.06.006. Epub 2019 Jun 15
75.Kim S, Takahashi H, Lin WW, Descargues P, Grivennikov S, Kim Y, et al. Carcinoma-produced factors activate myeloid cells through TLR2 to stimulate metastasis. Nature. 2009;457(7225):102-106. DOI: 10.1038/nature07623
76.Bianchi F, Alexiadis S, Camisaschi C, Truini M, Centonze G, Milione M, et al. TLR3 expression induces apoptosis in human non-small-cell lung cancer. International Journal of Molecular Sciences. 2020;21(4):1-14. DOI: 10.3390/ijms2104144
77.He W, Liu Q , Wang L, Chen W, Li N, Cao X. TLR4 signaling promotes immune escape of human lung cancer cells by inducing immunosuppressive cytokines and apoptosis resistance. Molecular Immunology. 2007;44(11):2850-2859. DOI: 10.1016/j.molimm.2007.01.022
78.Tavora B, Mederer T, Wessel KJ, Ruffing S, Sadjadi M, Missmahl M, et al. Tumoural activation of TLR3-SLIT2 axis in endothelium drives metastasis. Nature. 2020;586(7828):299-304. DOI: 10.1038/s41586-020-2774
79.Berthon C, Driss V, Liu J, Kuranda K, Leleu X, Jouy N, et al. In acute myeloid leukemia, B7-H1 (PD-L1) protection of blasts from cytotoxic T cells is induced by TLR ligands and interferon-gamma and can be reversed using MEK inhibitors. Cancer Immunology, Immunotherapy. 2010;59(12):1839-1849. DOI: 10.1007/s00262-010-0909-y
80.Chen R, Huang M, Yang X, Chen XH, Shi MY, Li ZF, et al. CALR-TLR4 complex inhibits non-small cell lung cancer progression by regulating the migration and maturation of dendritic cells. Frontiers in Oncology. 2021;11:743050. DOI: 10.3389/fonc.2021.743050
81.Liotti F, Marotta M, Sorriento D, Pone E, Morra F, Melillo RM, et al. Toll like receptor 7 mediates inflammation resolution and inhibition of angiogenesis in non-small cell lung cancer. Cancers (Basel). 2021;13(4):1-18. DOI: 10.3390/cancers13040740
82.Zhou H, Chen JH, Hu J, Luo YZ, Li F, Xiao L, et al. High expression of toll like receptor 5 correlates with better prognosis in non-small-cell lung cancer: An anti-tumor effect of TLR5 signaling in non-small cell lung cancer. Journal of Cancer Research and Clinical Oncology. 2014;140(4):633-643. DOI: 10.1007/s00432-014-1616-4
83.Koh J, Kim S, Lee SN, Kim SY, Kim JE, Lee KY, et al. Therapeutic efficacy of cancer vaccine adjuvanted with nanoemulsion loaded with TLR7/8 agonist in lung cancer model. Nanomedicine. 2021;37:102415. DOI: 10.1016/j.nano.2021.102415
84.Ren T, Xu L, Jiao S, Wang Y, Cai Y, Liang Y, et al. TLR9 signaling promotes tumor progression of human lung cancer cell in vivo. Pathology Oncology Research. 2009;15(4):623-630. DOI: 10.1007/s12253-009-9162-0
85.Negrao MV, Papadimitrakopoulou VA, Price AC, Tam AL, Furqan M, Laroia ST, et al. Vidutolimod in combination with Atezolizumab with and without radiation therapy in patients with programmed cell death protein 1 or programmed death-ligand 1 blockade-resistant advanced NSCLC. JTO Clinical and Research Reports. 2022;4(3):100423. DOI: 10.1016/j.jtocrr.2022.100423
86.Yang H, Zhou H, Feng P, Zhou X, Wen H, Xie X, et al. Reduced expression of toll-like receptor 4 inhibits human breast cancer cells proliferation and inflammatory cytokines secretion. Journal of Experimental & Clinical Cancer Research. 2010;29:92. DOI: 10.1186/1756-9966-29-92
87.Di Lorenzo A, Bolli E, Ruiu R, Ferrauto G, Di Gregorio E, Avalle L, et al. Toll-like receptor 2 promotes breast cancer progression and resistance to chemotherapy. Oncoimmunology. 2022;11(1):2086752. DOI: 10.1080/2162402X.2022.2086752
88.Dewan MZ, Vanpouille-Box C, Kawashima N, DiNapoli S, Babb JS, Formenti SC, et al. Synergy of topical toll-like receptor 7 agonist with radiation and low-dose cyclophosphamide in a mouse model of cutaneous breast cancer. Clinical Cancer Research. 2012;18(24):6668-6678. DOI: 10.1158/1078-0432.CCR-12-0984. Epub 2012 Oct 9
89.Bernardo AR, Cosgaya JM, Aranda A, Jiménez-Lara AM. Synergy between RA and TLR3 promotes type I IFN-dependent apoptosis through upregulation of TRAIL pathway in breast cancer cells. Cell Death & Disease. 2013;4(1):e479. DOI: 10.1038/cddis.2013.5
90.Ye K, Wu Y, Sun Y, Lin J, Xu J. TLR4 siRNA inhibits proliferation and invasion in colorectal cancer cells by downregulating ACAT1 expression. Life Sciences. 2016;155:133-139. DOI: 10.1016/j.lfs.2016.05.012. Epub 2016 May 10
91.Chen M, Zhong K, Tan J, Meng M, Liu CM, Chen B, et al. Baicalein is a novel TLR4-targeting therapeutics agent that inhibits TLR4/HIF-1α/VEGF signaling pathway in colorectal cancer. Clinical and Translational Medicine. 2021;11(11):e564. DOI: 10.1002/ctm2.564
92.Teng Z, Sun X, Guo Y, Zhang M, Liu Y, Xu M. Curcumae longae Rhizoma (Jianghuang) extract reverses the 5-Fluoruracil resistance in colorectal cancer cells via TLR4/PI3K/Akt/mTOR pathway. Clinics and Research in Hepatology and Gastroenterology. 2022;46(9):101976. DOI: 10.1016/j.clinre.2022.101976. Epub 2022 Jun 13
93.Damiano V, Rosa R, Formisano L, Nappi L, Gelardi T, Marciano R, et al. Toll-like receptor 9 agonist IMO cooperates with everolimus in renal cell carcinoma by interfering with tumour growth and angiogenesis. British Journal of Cancer. 2013;108(8):1616-1623. DOI: 10.1038/bjc.2013.153. Epub 2013 Apr 9
94.Weihrauch MR, Richly H, von Bergwelt-Baildon MS, Becker HJ, Schmidt M, Hacker UT, et al. Phase I clinical study of the toll-like receptor 9 agonist MGN1703 in patients with metastatic solid tumours. European Journal of Cancer. 2015;51(2):146-156. DOI: 10.1016/j.ejca.2014.11.002. Epub 2014 Dec 2
95.Niu Z, Tang W, Liu T, Xu P, Zhu D, Ji M, et al. Cell-free DNA derived from cancer cells facilitates tumor malignancy through toll-like receptor 9 signaling-triggered interleukin-8 secretion in colorectal cancer. Acta Biochimica et Biophysica Sinica (Shanghai). 2018;50(10):1007-1017. DOI: 10.1093/abbs/gmy104
96.Rath T, Stöckle J, Roderfeld M, Tschuschner A, Graf J, Roeb E. Matrix metalloproteinase-13 is regulated by toll-like receptor-9 in colorectal cancer cells and mediates cellular migration. Oncology Letters. 2011;2(3):483-488. DOI: 10.3892/ol.2011.276. Epub 2011 Mar 21
97.Luo Q , Zeng L, Tang C, Zhang Z, Chen Y, Zeng C. TLR9 induces colitis-associated colorectal carcinogenesis by regulating NF-κB expression levels. Oncology Letters. 2020;20(4):110. DOI: 10.3892/ol.2020.11971. Epub 2020 Aug 12
98.Lowe EL, Crother TR, Rabizadeh S, Hu B, Wang H, Chen S, et al. Toll-like receptor 2 signaling protects mice from tumor development in a mouse model of colitis-induced cancer. PLoS One. 2010;5(9):e13027. DOI: 10.1371/journal.pone.0013027
99.Liu YD, Ji CB, Li SB, Yan F, Gu QS, Balic JJ, et al. Toll-like receptor 2 stimulation promotes colorectal cancer cell growth via PI3K/Akt and NF-κB signaling pathways. International Immunopharmacology. 2018;59:375-383. DOI: 10.1016/j.intimp.2018.04.033. Epub 2018 Apr 24
100.Zhao J, Xue Y, Pan Y, Yao A, Wang G, Li D, et al. Toll-like receptor 3 agonist poly I:C reinforces the potency of cytotoxic chemotherapy via the TLR3-UNC93B1-IFN-β signaling axis in paclitaxel-resistant colon cancer. Journal of Cellular Physiology. 2019;234(5):7051-7061. DOI: 10.1002/jcp.27459. Epub 2018 Nov 1
101.Beilmann-Lehtonen I, Hagström J, Mustonen H, Koskensalo S, Haglund C, Böckelman C. High tissue TLR5 expression predicts better outcomes in colorectal cancer patients. Oncology. 2021;99(9):589-600. DOI: 10.1159/000516543. Epub 2021 Jun 17
102.Qian J, Luo F, Yang J, Liu J, Liu R, Wang L, et al. TLR2 promotes glioma immune evasion by downregulating MHC class II molecules in microglia. Cancer Immunology Research. 2018;6(10):1220-1233. DOI: 10.1158/2326-6066.CIR-18-0020. Epub 2018 Aug 21
103.Li C, Ma L, Liu Y, Li Z, Wang Q , Chen Z, et al. TLR2 promotes development and progression of human glioma via enhancing autophagy. Gene. 2019;700:52-59. DOI: 10.1016/j.gene.2019.02.084. Epub 2019 Mar 19
104.Wang F, Zhang P, Yang L, Yu X, Ye X, Yang J, et al. Activation of toll-like receptor 2 promotes invasion by upregulating MMPs in glioma stem cells. American Journal of Translational Research. 2015;7(3):607-615
105.Curtin JF, Liu N, Candolfi M, Xiong W, Assi H, Yagiz K, et al. HMGB1 mediates endogenous TLR2 activation and brain tumor regression. PLoS Medicine. 2009;6(1):e10. DOI: 10.1371/journal.pmed.1000010
106.Liu Y, Ju Y, Liu J, Chen Y, Huo X, Liu L. Inhibition of proliferation and migration and induction of apoptosis in glioma cells by silencing TLR4 expression levels via RNA interference. Oncology Letters. 2021;21(1):13. DOI: 10.3892/ol.2020.12274. Epub 2020 Nov 6
107.Jiang Y, Zhou J, Luo P, Gao H, Ma Y, Chen YS, et al. Prosaposin promotes the proliferation and tumorigenesis of glioma through toll-like receptor 4 (TLR4)-mediated NF-κB signaling pathway. eBioMedicine. 2018;37:78-90. DOI: 10.1016/j.ebiom.2018.10.053. Epub 2018 Oct 29
108.da Cruz LLP, de Souza PO, Dal Prá M, Falchetti M, de Abreu AM, Azambuja JH, et al. TLR4 expression and functionality are downregulated in glioblastoma cells and in tumor-associated macrophages: A new mechanism of immune evasion? Biochimica et Biophysica Acta - Molecular Basis of Disease. 2021;1867(8):166155. DOI: 10.1016/j.bbadis.2021.166155. Epub 2021 Apr 28
109.Stathopoulos A, Pretto C, Devillers L, Pierre D, Hofman FM, Kruse C, et al. Development of immune memory to glial brain tumors after tumor regression induced by immunotherapeutic toll-like receptor 7/8 activation. Oncoimmunology. 2012;1(3):298-305. DOI: 10.4161/onci.19068
110.Sandholm J, Tuomela J, Kauppila JH, Harris KW, Graves D, Selander KS. Hypoxia regulates toll-like receptor-9 expression and invasive function in human brain cancer cells in vitro. Oncology Letters. 2014;8(1):266-274. DOI: 10.3892/ol.2014.2095. Epub 2014 Apr 25
111.Liu D, Cao G, Cen Y, Liu T, Peng W, Sun J, et al. The radiosensitizing effect of CpG ODN107 on human glioma cells is tightly related to its antiangiogenic activity via suppression of HIF-1α/VEGF pathway. International Immunopharmacology. 2013;17(2):237-244. DOI: 10.1016/j.intimp.2013.06.002. Epub 2013 Jun 19
112.El Andaloussi A, Sonabend AM, Han Y, Lesniak MS. Stimulation of TLR9 with CpG ODN enhances apoptosis of glioma and prolongs the survival of mice with experimental brain tumors. Glia. 2006;54(6):526-535. DOI: 10.1002/glia.20401
113.Tiwari RK, Singh S, Gupta CL, Pandey P, Singh VK, Sayyed U, et al. Repolarization of glioblastoma macrophage cells using non-agonistic dectin-1 ligand encapsulating TLR-9 agonist: Plausible role in regenerative medicine against brain tumor. The International Journal of Neuroscience. 2021;131(6):591-598. DOI: 10.1080/00207454.2020.1750393. Epub 2020 Apr 19
114.Yu L, Wang L, Li M, Zhong J, Wang Z, Chen S. Expression of toll-like receptor 4 is down-regulated during progression of cervical neoplasia. Cancer Immunology, Immunotherapy. 2010;59(7):1021-1028. DOI: 10.1007/s00262-010-0825-1. Epub 2010 Feb 23
115.He A, Shao J, Zhang Y, Lu H, Wu Z, Xu Y. CD200Fc reduces LPS-induced IL-1β activation in human cervical cancer cells by modulating TLR4-NF-κB and NLRP3 inflammasome pathway. Oncotarget. 2017;8(20):33214-33224. DOI: 10.18632/oncotarget.16596
116.Oliveira LB, Louvanto K, Ramanakumar AV, Franco EL, Villa LL. For the Ludwig-McGill cohort study. Polymorphism in the promoter region of the toll-like receptor 9 gene and cervical human papillomavirus infection. The Journal of General Virology. 2013;94(Pt 8):1858-1864. DOI: 10.1099/vir.0.052811-0. Epub 2013 May 15
117.Pandey S, Mittal B, Srivastava M, Singh S, Srivastava K, Lal P, et al. Evaluation of toll-like receptors 3 (c.1377C/T) and 9 (G2848A) gene polymorphisms in cervical cancer susceptibility. Molecular Biology Reports. 2011;38(7):4715-4721. DOI: 10.1007/s11033-010-0607-z. Epub 2010 Dec 5
118.Zidi S, Verdi H, Yilmaz-Yalcin Y, Yazici AC, Gazouani E, Mezlini A, et al. Involvement of toll-like receptors in cervical cancer susceptibility among Tunisian women. Bulletin du Cancer. 2014;101(10):E31-E35. DOI: 10.1684/bdc.2014.2037
119.Mu X, Zhao J, Yuan X, Zhao X, Yao K, Liu Y, et al. Gene polymorphisms of toll-like receptor 9-1486T/C and 2848G/a in cervical cancer risk. International Journal of Gynecological Cancer. 2015;25(7):1173-1178. DOI: 10.1097/IGC.0000000000000494
120.Jin Y, Qiu S, Shao N, Zheng J. Association of toll-like receptor gene polymorphisms and its interaction with HPV infection in determining the susceptibility of cervical cancer in Chinese Han population. Mammalian Genome. 2017;28(5-6):213-219. DOI: 10.1007/s00335-017-9691-x
121.Pandey N, Chauhan A, Raithatha N, Patel P, Khandelwal R, Desai A, et al. Influence of TLR4 and TLR9 polymorphisms and haplotypes on multiple hrHPV infections and HPV16 copy number in cervical cancer and cervicitis. Microbial Pathogenesis. 2021;159:105149. DOI: 10.1016/j.micpath.2021.105149
122.Pandey S, Mittal RD, Srivastava M, Srivastava K, Singh S, Srivastava S, et al. Impact of toll-like receptors [TLR] 2 (−196 to −174 del) and TLR 4 (Asp299Gly, Thr399Ile) in cervical cancer susceptibility in north Indian women. Gynecologic Oncology. 2009;114(3):501-505. DOI: 10.1016/j.ygyno.2009.05.032. Epub 2009 Jun 21
123.Pandey NO, Chauhan AV, Raithatha NS, Patel PK, Khandelwal R, Desai AN, et al. Association of TLR4 and TLR9 polymorphisms and haplotypes with cervical cancer susceptibility. Scientific Reports. 2019;9(1):9729. DOI: 10.1038/s41598-019-46077-z. Erratum in: Sci Rep.;9(1):18658
124.Ashton KA, Proietto A, Otton G, Symonds I, McEvoy M, Attia J, et al. Toll-like receptor (TLR) and nucleosome-binding oligomerization domain (NOD) gene polymorphisms and endometrial cancer risk. BMC Cancer. 2010;10:382. DOI: 10.1186/1471-2407-10-382
125.Lupi LA, Cucielo MS, Silveira HS, Gaiotte LB, Cesário RC, Seiva FRF, et al. The role of toll-like receptor 4 signaling pathway in ovarian, cervical, and endometrial cancers. Life Sciences. 2020;247:117435. DOI: 10.1016/j.lfs.2020.117435
126.Saint-Jean M, Knol AC, Nguyen JM, Khammari A, Dréno B. TLR expression in human melanoma cells. European Journal of Dermatology. 2011;21(6):899-905. DOI: 10.1684/ejd.2011.1526
127.Mauldin IS, Wang E, Deacon DH, Olson WC, Bao Y, Slingluff CL Jr. TLR2/6 agonists and interferon-gamma induce human melanoma cells to produce CXCL10. International Journal of Cancer. 2015;137(6):1386-1396. DOI: 10.1002/ijc.29515
128.Speiser DE, Schwarz K, Baumgaertner P, Manolova V, Devevre E, Sterry W, et al. Memory and effector CD8 T-cell responses after nanoparticle vaccination of melanoma patients. Journal of Immunotherapy. 2010;33(8):848-858. DOI: 10.1097/CJI.0b013e3181f1d614
129.Goldinger SM, Dummer R, Baumgaertner P, Mihic-Probst D, Schwarz K, Hammann-Haenni A, et al. Nano-particle vaccination combined with TLR-7 and -9 ligands triggers memory and effector CD8+ T-cell responses in melanoma patients. European Journal of Immunology. 2012;42(11):3049-3061. DOI: 10.1002/eji.201142361. Epub 2012 Aug 28. Erratum in: Eur J Immunol. 2016 Feb;46(2):493
130.Ostojic N, Radevic T, Kandolf Sekulovic L, Djordjevic B, Jaukovic L, Stepic N, et al. Polymorphisms in toll-like receptor 3 and 4 genes as prognostic and outcome biomarkers in melanoma patients. Melanoma Research. 2022;32(5):309-317. DOI: 10.1097/CMR.0000000000000836
131.Elefanti L, Sacco G, Stagni C, Rastrelli M, Menin C, Russo I, et al. TLR7 Gln11Leu single nucleotide polymorphism and susceptibility to cutaneous melanoma. Oncology Letters. 2016;12(1):275-280. DOI: 10.3892/ol.2016.4584
132.Suresh D, Srinivas AN, Kumar DP. Etiology of hepatocellular carcinoma: Special focus on fatty liver disease. Frontiers in Oncology. 2020;10:601710. DOI: 10.3389/fonc.2020.601710
133.Huang Y, Cai B, Xu M, Qiu Z, Tao Y, Zhang Y, et al. Gene silencing of toll-like receptor 2 inhibits proliferation of human liver cancer cells and secretion of inflammatory cytokines. PLoS One. 2012;7(7):e38890. DOI: 10.1371/journal.pone.0038890
134.Shi L, Zheng X, Fan Y, Yang X, Li A, Qian J. The contribution of miR-122 to the innate immunity by regulating toll-like receptor 4 in hepatoma cells. BMC Gastroenterology. 2019;19(1):130. DOI: 10.1186/s12876-019-1048-3
135.Singh A, Koduru B, Carlisle C, Akhter H, Liu RM, Schroder K, et al. NADPH oxidase 4 modulates hepatic responses to lipopolysaccharide mediated by toll-like receptor-4. Scientific Reports. 2017;7(1):14346. DOI: 10.1038/s41598-017-14574-8
136.Minmin S, Xiaoqian X, Hao C, et al. Single nucleotide polymorphisms of toll-like receptor 4 decrease the risk of development of hepatocellular carcinoma. PLoS One. 2011;6(4):e19466
137.Neamatallah M, El-Bendary M, Elalfy H, Besheer T, El-Maksoud MA, Elhammady D, et al. Impact of toll-like receptors 2(TLR2) and TLR 4 gene variations on HCV susceptibility, response to treatment and development of hepatocellular carcinoma in cirrhotic HCV patients. Immunological Investigations. 2020;49(4):462-476. DOI: 10.1080/08820139.2019.1673772. Epub 2019 Oct 15
138.Bas S, Neff L, Vuillet M, Spenato U, Seya T, Matsumoto M. Gabay C the proinflammatory cytokine response to chlamydia trachomatis elementary bodies in human macrophages is partly mediated by a lipoprotein, the macrophage infectivity potentiator, through TLR2/TLR1/TLR6 and CD14. Journal of Immunology. 2008;180(2):1158-1168. DOI: 10.4049/jimmunol.180.2.1158
139.Guo H, Chen Y, Hu X, Qian G, Ge S, Zhang J. The regulation of toll-like receptor 2 by miR-143 suppresses the invasion and migration of a subset of human colorectal carcinoma cells. Molecular Cancer. 2013;12:77. DOI: 10.1186/1476-4598-12-77
140.Dajon M, Iribarren K, Cremer I. Toll-like receptor stimulation in cancer: A pro- and anti-tumor double-edged sword. Immunobiology. 2017;222(1):89-100. DOI: 10.1016/j.imbio.2016.06.009
141.Fehri E, Ennaifer E, Bel Haj Rhouma R, Ardhaoui M, Boubaker S. TLR9 and glioma: Friends or foes? Cell. 2022;12(1):152. DOI: 10.3390/cells12010152
142.Fehri E, Ennaifer E, Bel Haj Rhouma R, Guizani-Tabbane L, Guizani I, Boubaker S. The role of toll-like receptor 9 in gynecologic cancer. Current Research in Translational Medicine. 2016;64(3):155-159. DOI: 10.1016/j.retram.2016.01.010
Written By
Emna Fehri, Emna Ennaifer, Monia Ardhaoui, Haifa Tounsi, Rahima Bel Haj Rhouma and Samir Boubaker
Submitted: 01 September 2023Reviewed: 04 September 2023Published: 20 March 2024
 Open access peer-reviewed chapter
Open access peer-reviewed chapter