 Open access peer-reviewed chapter
Open access peer-reviewed chapter
Abstract
In the last decade, ostrich breeding in Chile has increased substantially, so it is necessary to increase studies to improve and understand the adaptation of this species to our ecology. Our objective is to contribute to the knowledge of the normal histology of the glands attached to the digestive system: liver, proventriculus and pancreas; digestive system: esophagus, proventriculus, muscular stomach, small intestine (duodenum, jejunum and ileum), large intestine (cecum, colon and rectum); respiratory system: larynx, trachea, syrinx, extrapulmonary primary bronchi, intrapulmonary primary bronchi or mesobronchi, secondary bronchi, parabronchi or tertiary bronchi, air capillaries and air sacs; and female reproductive system: ovary and oviduct (infundibulum, magnum, isthmus, uterus, and vagina) of Struthio camelus var. domesticus in a first stage. This research was carried out with clinically healthy ostriches, from which samples of the different systems were obtained for this study. Histological sections were made, which were stained and mounted for their comparative analysis under a light microscope, between ostrich and chicken. The comparative histological analysis allowed us to establish that the structure of the different apparatuses and systems is similar to that observed in the chicken. However, at the microscopic level some differences are observed.
Keywords
- ostrich
- histology
- digestive system glands
- digestive system
- respiratory system
- female reproductive system
1. Introduction
There are currently four subspecies of ostrich:
The digestive system presents glands such as the proventriculus (glandular stomach), liver, and pancreas that have been anatomically described. The true stomach of the ostrich corresponds to the proventriculus or glandular stomach, it is sac-shaped and anatomically occupies the cranial part of the abdomen in the left hypochondrium [4], where protein digestion begins through the secretion of pepsinogen and hydrochloric acid [3]. In relation to the general anatomical characteristics of the digestive tract, it is known that the ostrich presents similarities and differences, both with other birds, as well as with ruminants and other herbivores (equines) [3]. The comparative analysis with the hen presents certain anatomical differences, one of them is not having a crop, a food storage organ that exists in other birds. The proventriculus and the muscular stomach (gizzard), in the ostrich, can fulfill this function [5]. They do not have a gallbladder, so the emptying of bile is done directly into the small intestine. In addition, the large intestine of the ostrich, unlike other species, represents 50% of the total length of the digestive tract and the small intestine corresponds to only 35.5% [2].
Among the anatomical characteristics of the ostrich respiratory system, it is pointed out that the larynx is formed by two arytenoid cartilages and one cricoid cartilage, which together form a ring [1]. The trachea extends between the larynx and the syrinx, with the dorso-ventrally compressed lumen in the upper third. It is made up of approximately 200 complete cartilaginous or osseous rings [1]. The syrinx is simple, formed by the last rings of the trachea and the proximal half rings of the extrapulmonary bronchi. These extend into the lungs only a short distance [1]. Regarding the lungs, they are located in the dorsal third of the thoracoabdominal cavity, between the second and seventh pair of ribs [1]. Like flying birds, the ostrich has air sacs incorporated into its anatomical structure, in a number of five pairs (clavicular, interclavicular, pre-thoracic, post-thoracic, and abdominal). In birds, many of the bones have cavities that communicate with the air sacs, which is why they are called pneumatic bones. In the case of the ostrich (
The reproductive organs of the female ostrich are the ovary and left oviduct that empties into the urodeum, with the rudimentary right ovary being [1, 6]. In the ostrich, the ovary is suspended from the dorsal body wall, ventral to the kidney and dorsomedial to the ipsilateral abdominal air sac. The position, size, and shape of the ovary depend on seasonality and vary during the breeding season [2]. The oviduct carries the fertilized oocyte to the cloaca, and in the opposite direction allows sperm to reach the fertilization site [7]. The oviduct of birds has five regions each with their respective function, these are infundibulum, magnum, isthmus, uterus and vagina [8]. The infundibulum receives and encloses the yolk, and it is the site where fertilization occurs [9]. The albumin or white is secreted in the magnum and is the longest segment of the oviduct. In the isthmus, the testaceous membranes of the shell form around the egg [2]. Calcification of the shell occurs in the uterus (or shell gland) which is globose in shape. The vagina in birds allows the passage of the egg formed from the uterus to the cloaca during posture or oviposition and is also important in the selection, transport, and storage of spermatozoa [10]. The ostrich reaches sexual maturity in the third year of life, but with nutritional management, it can be brought forward to the second year of life. The ostrich has a seasonal breeding period of long days and can lay up to 50 eggs per season. The time it takes between the formation and laying of an egg is 48 hours compared to the hen which is approximately 24 hours [2].
The objective of this work is to identify, classify, and histologically characterize the glands associated with the digestive system: liver, proventriculus, and pancreas; the different segments of the digestive system: esophagus, proventriculus, muscular stomach, small intestine (duodenum, jejunum, and ileum), and large intestine (cecum, colon, and rectum); respiratory system: larynx, trachea, syrinx, extrapulmonary primary bronchi, intrapulmonary primary bronchi or mesobronchi, secondary bronchi, parabronchi or tertiary bronchi, air capillaries, and air sacs; and female genital apparatus: ovary and oviduct comprising: infundibulum, magnum, isthmus, uterus, and vagina of
2. Material and methods
The biological material used in this work corresponds to samples of glands associated with the digestive system and the different segments of the digestive, respiratory, and female reproductive systems, obtained from clinically healthy adult ostriches (
3. Glands attached to the digestive system
The glands attached to the ostrich digestive system correspond to: liver, which has 4 lobes and is located in the right hypochondrium; proventriculus, which presents a wide lumen and 4 layers recognized in its wall: mucosa, submucosa, muscular and serous; and pancreas, it is a mixed gland, exocrine and endocrine, very lobed, made up of parenchyma (pancreatic islets and glandular acini), and a stroma (capsule, septa, and hilum), [12].
Liver: The parenchyma is irregularly septated and made up of cords of hepatocytes arranged radiating from a central vein. Hepatocytes generally have one or two nuclei. Sinusoidal capillaries were found between the cords, where macrophages were observed (Figures 1 and 2). It should be noted that the hepatocytes located around the portal spaces are also arranged radially (Figure 3). From the capsule arise the partitions that divide the organ irregularly. The portal space corresponds to the loose connective tissue that contains blood vessels derived from the portal vein and the hepatic artery, lymphatic vessels, and bile ducts, the latter presenting simple cuboidal lining epithelium (Figures 1
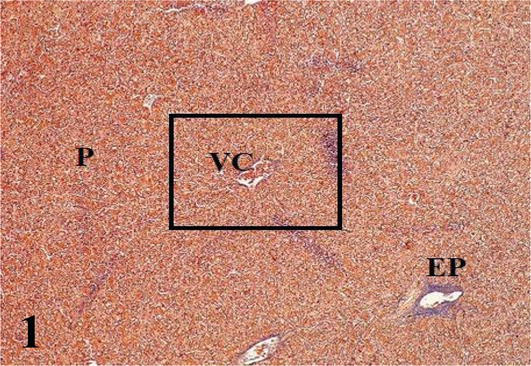
Figure 1.
Histological section of a liver lobule. VC: central vein; P: parenchyma and EP: portal space. Mallory’s trichrome. 100×.
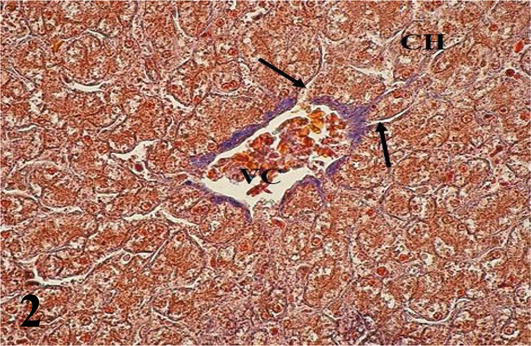
Figure 2.
Higher magnification of the inset of
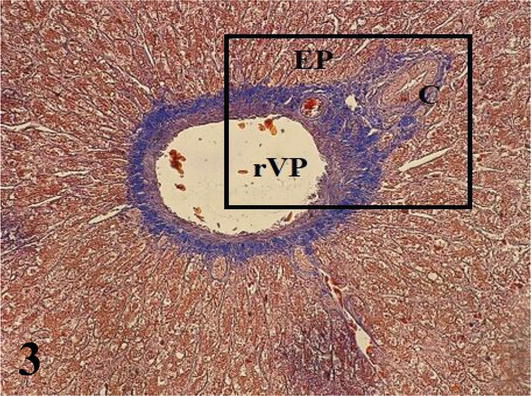
Figure 3.
Histological section of a portal space. EP: portal space; rVP: branch of the portal vein and C: bile duct. Note the ray arrangement of the hepatocytes around the portal space. Mallory’s trichrome. 200×.
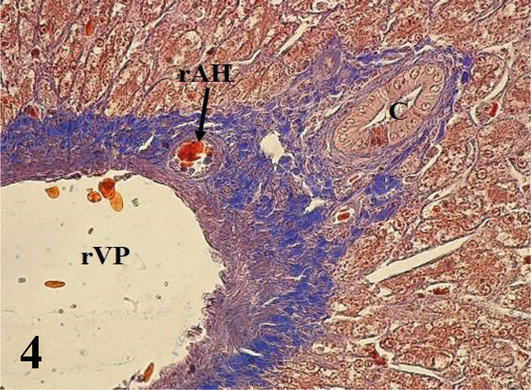
Figure 4.
Higher magnification of the inset of
Proventriculus (Mucosa and Submucosa): The mucosa with simple prismatic epithelium is covered with mucus (glycocalyx). With a large number of villi and between its bases crypts with simple cuboidal epithelium are observed. The lamina propria of loose connective tissue with simple or branched tubular glands secreting pepsinogen and hydrochloric acid, extensive lymphocyte infiltration (Figures 5 and 6). The muscularis mucosa of smooth muscle tissue, not well defined, emits septa into the submucosa. The submucosa is thick and made up of dense connective tissue that emits septa around the compound tubuloalveolar glands, constituting lobules formed by conglomerates of tubuloalveolar adenomeres around a central lumen (Figures 5 and 6). The glands discharge their secretion into the lumen of the proventriculus, through its excretory duct lined by simple columnar epithelium. Among the cubic and cylindrical secretory cells of the adenomeres, the parietal cells, which produce pepsinogen and hydrochloric acid, are abundant. Ganglion cells of the submucosal plexuses are also seen in the submucosa of the proventriculus.
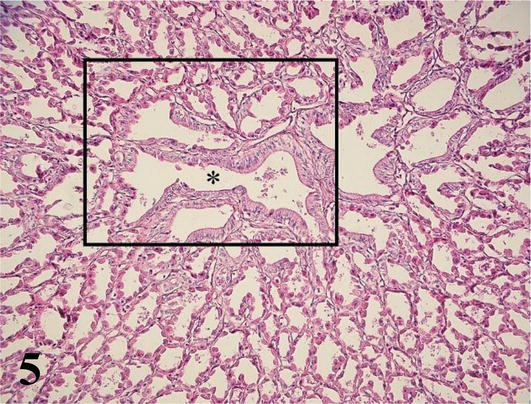
Figure 5.
Histological section of the submucosa of the proventriculus. (*): Excretory duct and tubuloalveolar adenomeres of the compound submucosal glands that empty into it. Staining: H-E. 200×.
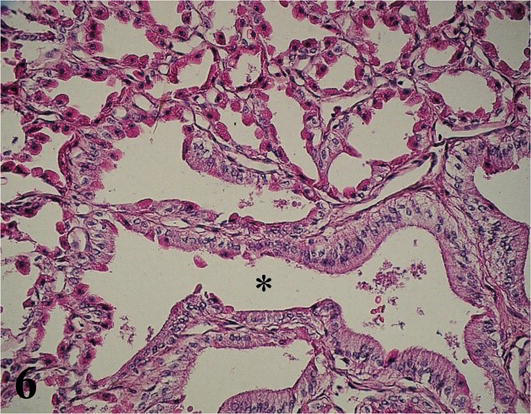
Figure 6.
Higher magnification of the inset of
Pancreas: The exocrine parenchyma is formed by tubuloacinar adenomeres composed of serous secretion and highly irrigated cell groups, which correspond to pancreatic islets that fulfill the endocrine function. The cells that constitute the adenomeres are prismatic with a round nucleus located in the basal third. The staining affinity of this zone is basophilic and the apical one is acidophilic (Figures 7 and 8). In the lumen of the acini, centro-acinous cells are observed that correspond to the beginning of the excretory ducts. The stroma is made up of the capsule that surrounds the organ, the septa, and the hilum. The capsule is relatively thin, made up of loose connective tissue that forms partitions toward the interior of the parenchyma, dividing it into small lobes and lobules.
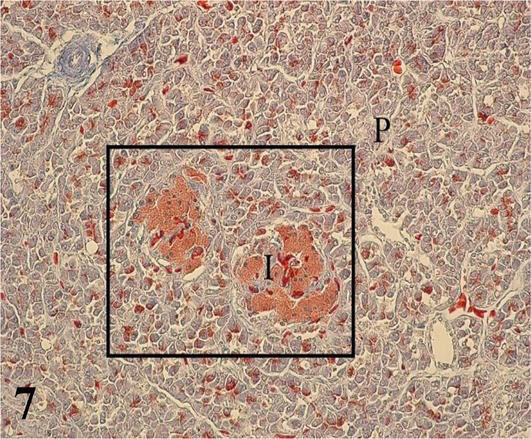
Figure 7.
Histological section of the pancreatic parenchyma. I: pancreatic islets and P: parenchyma. Mallory’s trichrome. 200×.
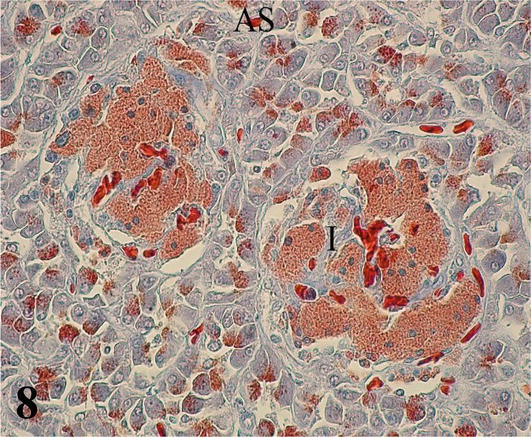
Figure 8.
Higher magnification of the inset of
4. Digestive system
The segments that make up the digestive system correspond to the esophagus, proventriculus (glandular stomach), muscular stomach (ventricle or gizzard), small intestine, and large intestine. The wall of the digestive system presents four concentric layers or tunics: mucosa; submucosa, muscular and serosa, and/or adventitia. Throughout the digestive system, there are histological differences in each layer, mainly in the tunica mucosa, which are manifested by the functional characteristics of each segment [13].
Esophagus: A mucosa with a non-keratinized stratified squamous epithelium. A lamina propria or chorion of dense connective tissue with abundant simple and branched tubuloalveolar mucous secretion glands throughout its length. A large number of lymph nodes are observed. The muscularis mucosa presents only a thick layer of longitudinal smooth muscle fibers and determines the boundary between the lamina propria and the submucosa (Figures 9 and 10). The submucosa is poorly developed, made up of loose connective tissue with submucosal nerve plexuses along its structure. The muscularis is made up of a thick internal circular layer of smooth muscle and a more irregular and thin external longitudinal layer. Between both muscle layers, myenteric nerve plexuses can be seen. The serosa is made up of the mesothelium and loose connective tissue.
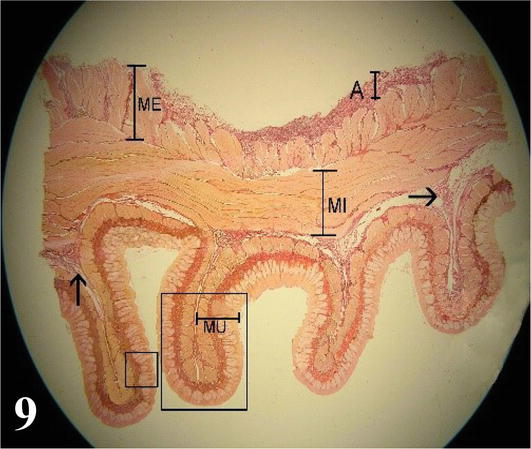
Figure 9.
Panoramic cross-section of the esophagus. MU. mucous membrane; submucosa (arrow); MI: internal muscle; ME: external muscle; A: adventitia. Note the longitudinal folds. Staining: Van Gieson 12.5×.
Figure 10.
Higher magnification of the large box in
Proventriculus (glandular stomach): The mucosa with the simple prismatic epithelium of mucus secretion, with numerous villi and covered by a large amount of mucus (glycocalyx). The simple and branched tubular fundic glands, with principal and parietal cells, empty their secretion into the crypts. A lamina propria of loose connective tissue. A muscularis mucosa of smooth muscle tissue. Compound tubuloalveolar-type glands with parietal cells and excretory ducts of simple prismatic epithelium are observed in the submucosa. Submucosal plexuses are observed. The muscularis formed by a thick internal circular layer and a thin external longitudinal layer of smooth muscle. Between both layers myenteric plexuses are observed. The serosa is constituted by the mesothelium and loose connective tissue. Between the muscular and serous layers, we find a thick sheet of compact connective tissue with parallel bundles; this tendon layer surrounds the entire organ (Figures 11 and 12).
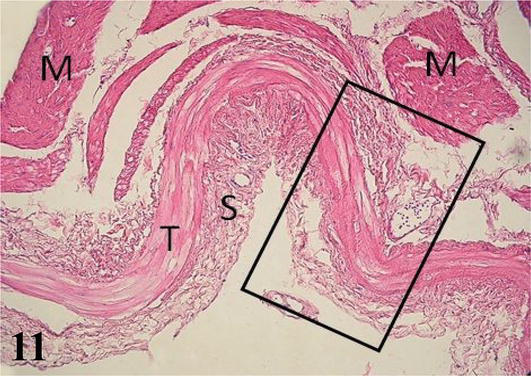
Figure 11.
Histological section of the proventriculus. M: muscular; T: tendon layer and S: serosa. Staining: H-E 200×.
Figure 12.
Higher magnification of the inset of
Muscular stomach (gizzard): The mucosa with simple prismatic epithelium is covered by a thick horny-looking layer, which corresponds to the cuticle (glycocalyx). It has a large number of wrinkles on its surface. A lamina propria of loose connective tissue with numerous simple and branching tubuloalveolar glands of mucous secretion. The muscularis mucosa is seen as a thin layer of smooth muscle fibers. These muscle fibers run parallel to the longitudinal axis and, in some portions, merge with the muscularis tunica. The submucosa of irregular dense connective tissue with septa branching into the muscular layer (Figure 13). The muscularis is thick, showing three layers of smooth muscle that correspond to an inner plexiform layer, a middle transverse layer, and a thick outer longitudinal layer (Figure 13). Numerous myenteric plexuses are observed between the muscle layers (Figure 14). The serosa is constituted by the mesothelium and loose connective tissue.
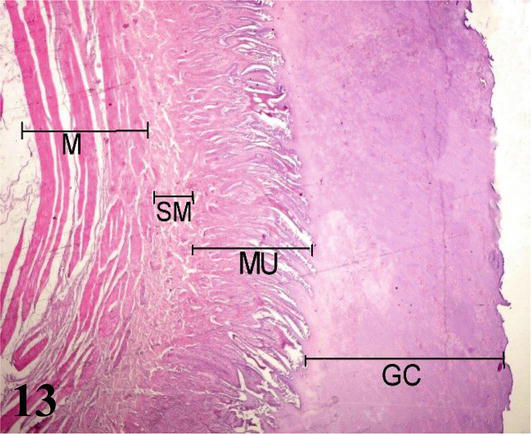
Figure 13.
Panoramic histological section of the muscular stomach wall. GC: glycocalyx; MU: mucosa; SM: submucosa and M: muscular. Staining: H-E 12.5×.
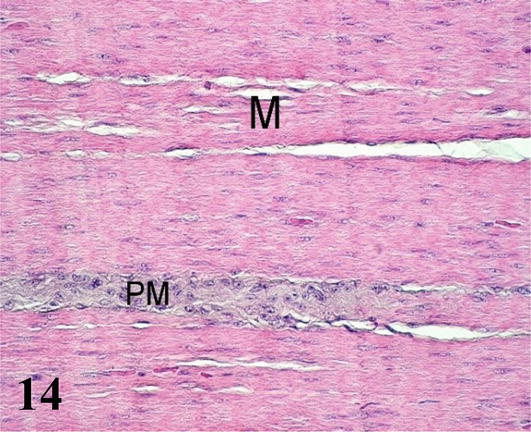
Figure 14.
Histological section of the muscular stomach. M: muscular and PM: myenteric nervous plexus. Staining: H-E 400×.
4.1 Small intestine
It consists of three segments called the duodenum, jejunum, and ileum. There are no limits defined between them. Its basic structure is similar to its entire length, being able to find different characteristics according to its function [13].
Duodenum: The mucosa is thick with a simple prismatic epithelium with striated plate and goblet cells forming numerous villi. Numerous intestinal crypts are seen at the base of the villi. Lamina propria of loose connective tissue with lymph nodes. The muscularis mucosa is thin and consists of an internal circular layer and an external longitudinal layer. The inner circular layer branches into the lamina propria to the apex of the villi (Figures 15 and 16). The poorly developed submucosa corresponds to loose connective tissue with nerve plexuses. Mucosal and submucosal projections corresponding to connivent valves are observed (Figures 15 and 16). The muscularis shows an internal circular layer and a finer external longitudinal layer. Numerous myenteric nerve plexuses are seen between the two layers. The serosa is constituted by the mesothelium and loose connective tissue.
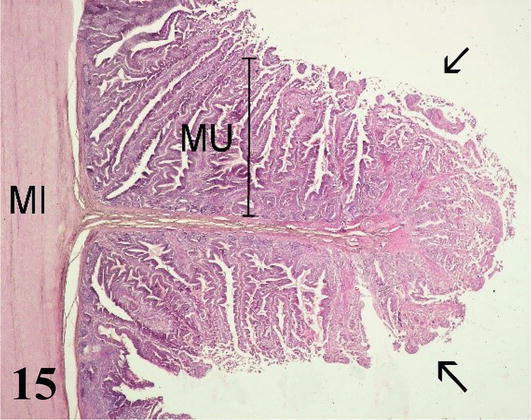
Figure 15.
Histological section of a connivent valve of the duodenum. MU: mucosa; MI: internal muscle and connivent valve (arrows). Staining: H-E 100×.
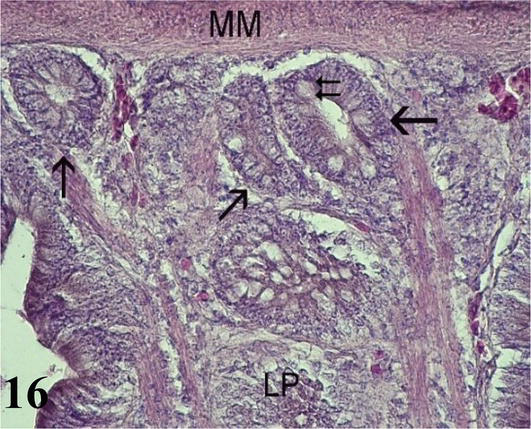
Figure 16.
Histological section of the duodenum. LP: lamina propria; MM: muscular mucosa; intestinal crypts (arrows) and goblet cells (double arrow). Staining: H-E 400×.
Jejunum: This segment does not present the connivent valves described in the duodenum. However, the basic histological structure of this segment is similar to that of the duodenum. The mucosa with simple prismatic lining epithelium with striated plate and numerous goblet cells show long villi with smooth muscle fibers from the muscularis mucosa (Figures 17 and 18). The lamina propria with lymph nodes. The muscularis mucosa is thin and corresponds to two layers of smooth muscle fibers, one internal with a circular arrangement and the other external longitudinal. The submucosa is poorly developed with submucosal nerve plexuses. The muscularis is thicker than seen in the anterior segment, with an inner circular layer and a thinner outer longitudinal layer. The serosa is constituted by the mesothelium and loose connective tissue.
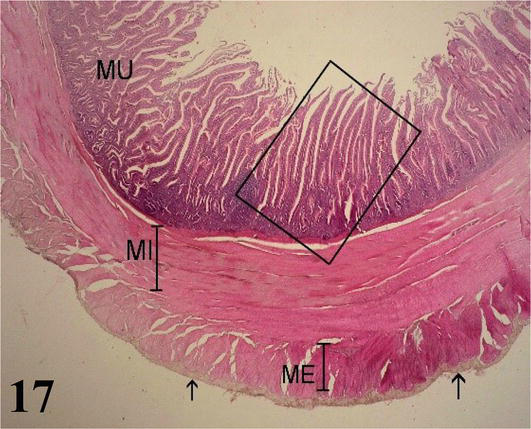
Figure 17.
Histological section of the jejunum. MU: mucosa; MI: internal muscle; ME: external muscle and serosa (arrows). Staining: H-E 40×.
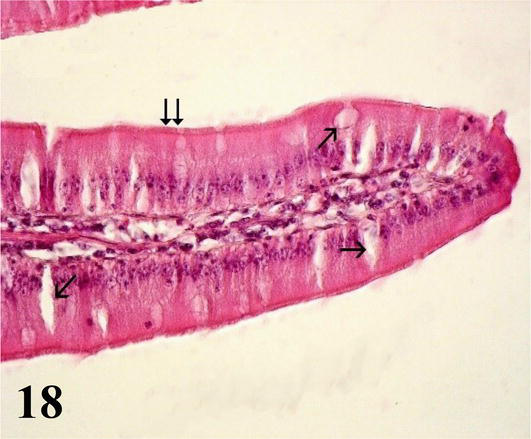
Figure 18.
Higher magnification of the inset of
Ileum: The mucosa with a simple prismatic epithelium, with a striated plate and goblet cells, shows fewer, shorter, and thicker villi than in the anterior segments. Numerous intestinal crypts are observed. The intestinal villi are made up of a central axis of loose connective tissue and smooth muscle fibers from branches of the muscularis mucosa (Figure 19). The lamina propria shows lymph nodes. The muscularis mucosa presents three layers of smooth muscle, an internal circular, an intermediate longitudinal, and an external circular (Figure 20). The submucosa is poorly developed with submucosal nerve plexuses. The muscularis is arranged in a thin internal circular layer and a thicker external longitudinal layer, between the two myenteric plexuses are observed. The serosa is made up of the mesothelium and loose connective tissue.
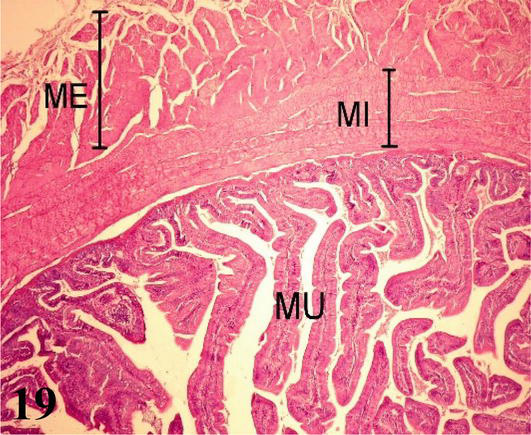
Figure 19.
Histological section of the ileum. MU: mucosa; MI: internal muscle and ME: external muscle. Staining: H-E 100×.
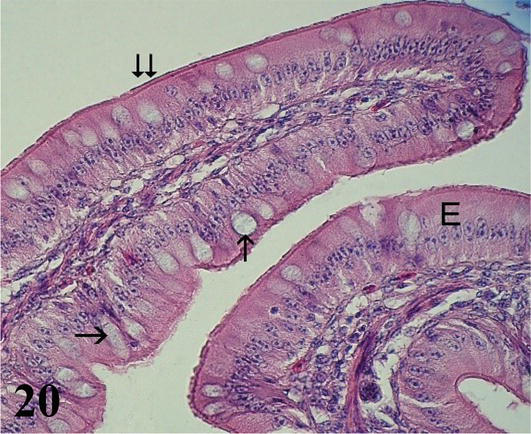
Figure 20.
Histological section of an ileum villus. E: epithelium; Goblet cells (arrows) and striated plate (double arrow). Staining: H-E 400×.
4.2 Large intestine
The large intestine of the adult ostrich measures approximately 16 m in length and represents the last portion of the alimentary canal. It has three segments: cecum, colon, and rectum. The general histological structure is similar throughout its length, showing differences in its segments according to its function [13].
Cecum: The mucosa with a simple prismatic epithelium without intestinal villi. With numerous intestinal crypts and numerous goblet cells, more abundant than in the small intestine. The lamina propria with isolated lymph nodes. The muscularis mucosa corresponds to two layers of smooth muscle fibers, one internal circular and one external longitudinal (Figure 21). The submucosa consists of a thin layer of loose connective tissue with submucosal nerve plexuses and a large amount of adipose tissue (Figure 22). The muscularis is less developed than in the small intestine, it is formed by a thin internal circular layer and an external longitudinal layer with myenteric nerve plexuses. Adipose tissue is observed between both muscle layers. The serosa is constituted by the mesothelium and loose connective tissue.
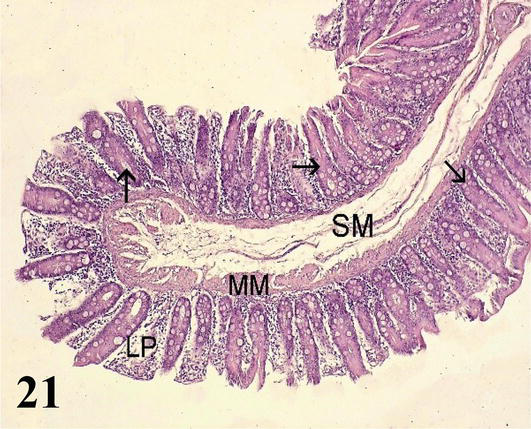
Figure 21.
Histological section of the cecum. LP: lamina propria; MM: muscular mucosa; SM: Submucosa and intestinal crypts (arrows). Staining: H-E 200X.
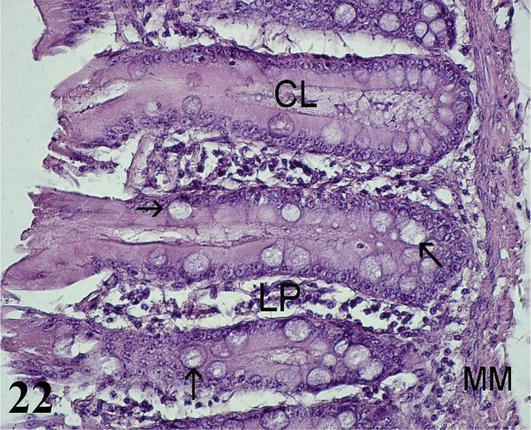
Figure 22.
Histological section of the cecum mucosa. LP: lamina propria; MM: muscular mucosa; CL: intestinal crypts and goblet cells (arrows). Staining: H-E 400×.
Colon: The mucosa with simple prismatic epithelium and fewer intestinal crypts with numerous goblet cells and no intestinal villi. The lamina propria shows isolated lymph nodes (Figures 23 and 24). The muscularis mucosa is thin and shows an internal circular layer and an external longitudinal layer. The internal circular layer has ramifications for the lamina propria (Figure 23). The submucosa is made up of loose connective tissue with adipose tissue and submucosal nerve plexuses. Connivent valves are observed toward the lumen, which are formed by nuclei of mucosa and submucosa (Figures 23 and 24). The muscularis formed by two layers of smooth muscle, an internal one with a circular arrangement and a thinner external one with a longitudinal arrangement with myenteric nerve plexuses. The serous layer of loose connective tissue forms sacs, which follow the direction of the sacs that form in the muscular layer.
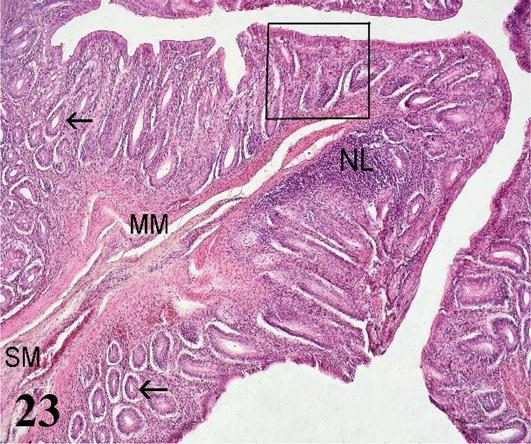
Figure 23.
Histological section of the colon. H-E 200×. MM: muscular mucosa; SM: submucosa; LN: lymph node and intestinal crypts (arrows). Staining: H-E 200×.
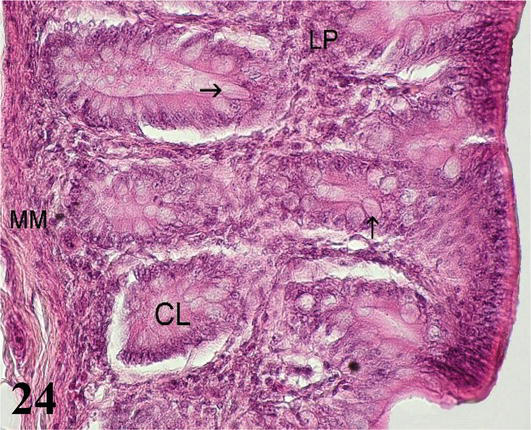
Figure 24.
Higher magnification of the inset of
Rectum: Long folds are visible toward the lumen longitudinal in the form of columns, which have a core mucosa and submucosa (Figure 25). The mucosa is smooth, devoid of villi, with a simple columnar epithelium. Intestinal crypts are deeper than in the small intestine, with a greater number of cells goblet than in the anterior segments. The muscular of the mucosa is thin and is formed by a layer of smooth muscle fibers in a circular arrangement (Figure 26). The submucosa is made up of loose connective tissue. Its structure is similar to that of the other segments (Figure 25). The muscularis is thick, and is formed by smooth muscles, one internal with a circular arrangement, and the other external longitudinal arrangement. The serosa does not show any differentiation from the usual.
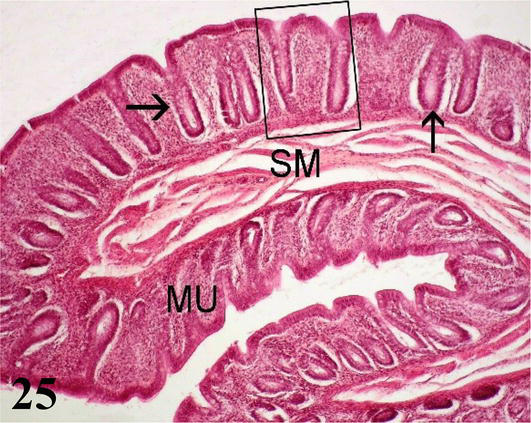
Figure 25.
Histological section of the rectum. MU: mucosa; SM: submucosa and intestinal crypts (arrows). Staining: H-E 200×.
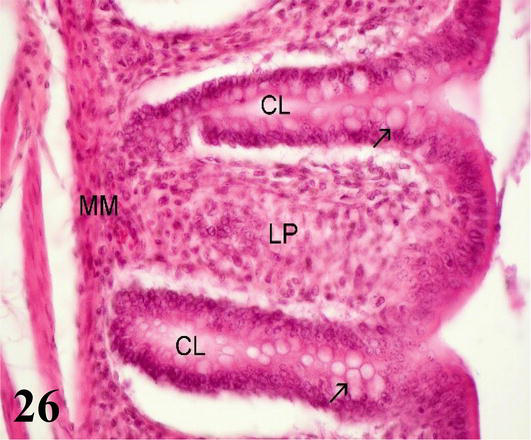
Figure 26.
Higher magnification of the inset of
5. Respiratory system
5.1 General description of the respiratory system of the ostrich
The segments analyzed correspond to the larynx, trachea, syrinx, extrapulmonary bronchus, intrapulmonary bronchus or mesobronchus, secondary bronchus, parabronchus or tertiary bronchus, and air capillaries. The wall of the respiratory system presents three concentric layers or tunics: mucosa, submucosa, and adventitia. The greatest difference in the wall of these pathways was observed at the level of the mucosal layer, which expresses the functional characteristics of each segment [14].
Larynx: The mucosa of the larynx, from cranial to caudal, undergoes a change from noncornified squamous stratified epithelium to ciliated prismatic pseudostratified with goblet cells with shallow crypts (Figures 27 and 28). The lamina propria of dense connective tissue with simple tubuloalveolar glands of mucus secretion. Between the glands is diffuse dense lymphoid tissue with isolated lymphoid nodules, which are also frequently seen at the onset and end points of epithelial changes (Figures 27 and 28). The submucosa is very thin and has loose connective tissue. The adventitia has hyaline cartilage that constitutes the craniolateral portion of the arytenoid cartilage (Figures 27 and 28). Toward the caudal region, the laryngeal mucosa is histologically similar and its adventitia shows a hyaline cartilage corresponding to the cricoid cartilage. More externally, it presents a layer of skeletal musculature that surrounds the larynx. Caudally, the larynx is continuous with numerous cartilaginous rings that constitute the beginning of the trachea.
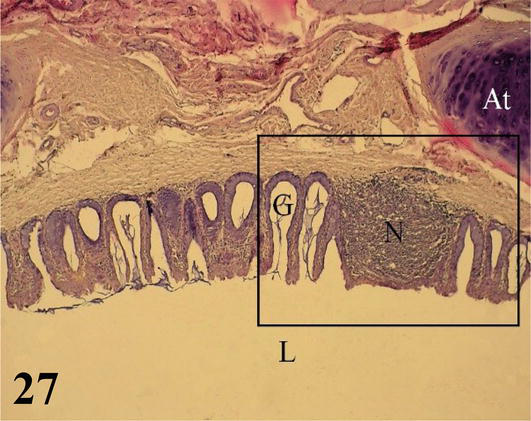
Figure 27.
Ciliated prismatic pseudostratified epithelium with goblet cells, with shallow crypts and simple mucus-secreting tubulo-alveolar glands (G); lymphoid nodule (N) and tracheal cartilage ring (At). Lumen of the larynx (L). Staining: H-E, 100×.
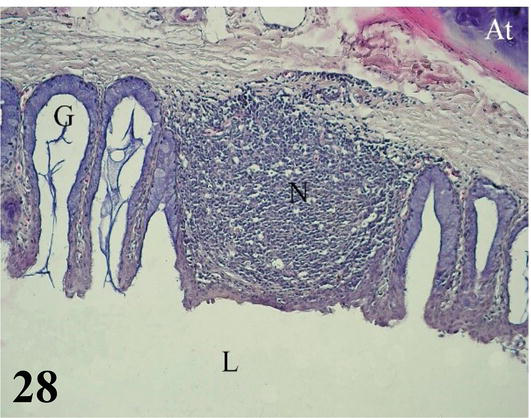
Figure 28.
Higher magnification of the inset of
Trachea: The mucosa with ciliated prismatic pseudostratified epithelium with goblet cells with shallow crypts (Figures 29–31). The lamina propria of dense connective tissue with simple mucus-secreting tubuloalveolar glands and isolated lymphoid nodules (Figures 30 and 31). The loose connective tissue submucosa is sparse. The adventitia is represented by rings of hyaline cartilage. On each side of the trachea and external to the cartilage, fascicles of skeletal muscle can be seen running longitudinally from the larynx to the syrinx, the lateral muscle of the trachea. The tracheal adventitia at the level of the syrinx is limited on its ventral or anterior face by mesothelium, which constitutes part of the interclavicular air sac.
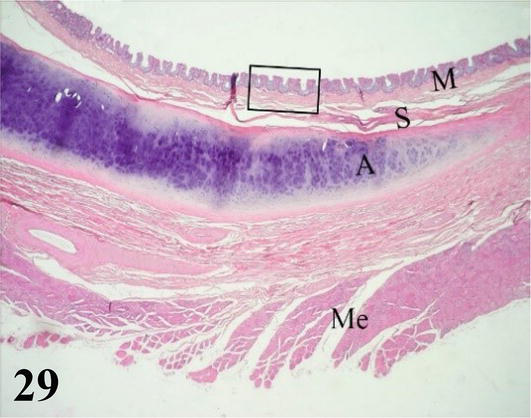
Figure 29.
This figure shows the mucosa (M), submucosa (S), and adventitia A, represented by a tracheal ring. Externally, the skeletal musculature (Me) can be seen. Staining: H-E, 40×.
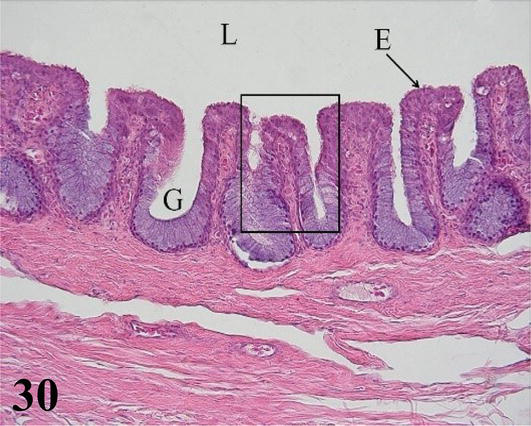
Figure 30.
Higher magnification of the inset of
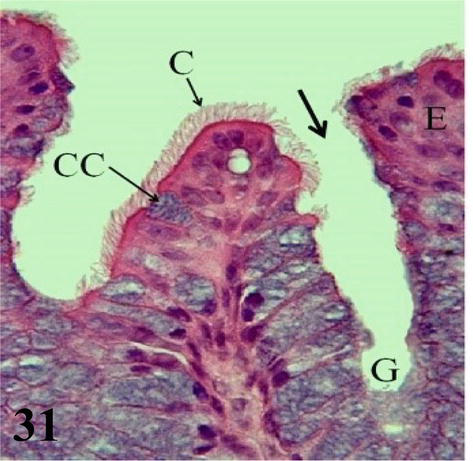
Figure 31.
Higher magnification of the inset of
Syrinx: Microscopically, the syrinx in a coronal section shows the syringeal (tracheosyringeal) rings that are morphologically indistinguishable from the tracheal rings (Figures 32 and 33). In the widest area of the syrinx, a group of cartilaginous rings is observed accompanied by abundant loose connective tissue (Figure 33). Mucosa with ciliated prismatic pseudostratified epithelium with goblet cells and shallow crypts. The lamina propria of dense connective tissue with simple tubuloalveolar glands of mucus secretion, and isolated lymphoid nodules are observed (Figures 34 and 35). In the caudal direction, the adventitia is represented by cartilaginous rings that become semirings (bronchosyringeals), which are continuous with those that constitute the extrapulmonary primary bronchi. On the floor of the bifurcation of the syrinx, a double thickening of a medial and dorsoventral arrangement is observed, which prominence toward the syringeal cavity, the pessulus (Figure 32), with a ciliated prismatic pseudostratified epithelium with goblet cells and shallow crypts and lamina propria of connective tissue dense with simple tubuloalveolar glands of mucus secretion with isolated lymphoid nodules and deeper a thick layer of adipose tissue (Figures 33–35). The loose connective tissue submucosa is sparse. The hyaline cartilage adventitia in the syrinx is limited by the pleural mesothelium, which becomes the wall of the thoracoabdominal air sacs, the latter present a ciliated prismatic pseudostratified epithelium with goblet cells, which in some sectors can be seen simple tubuloalveolar glands of mucous secretion (Figures 36–38).
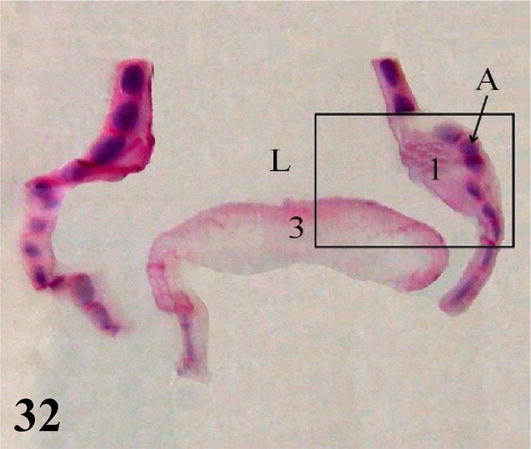
Figure 32.
Coronal panoramic histological section of the syrinx. 1: lateral tympaniform membrane; 3: central membrane or pessulus; A: hyaline cartilage of the tracheosyringeal ring; L: lumen of the syrinx. Staining: H-E, 12.5×.
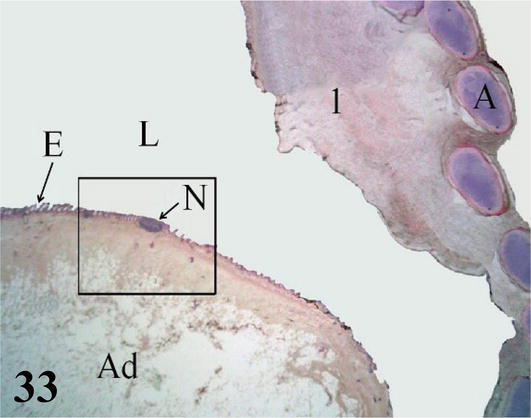
Figure 33.
Higher magnification of the inset of
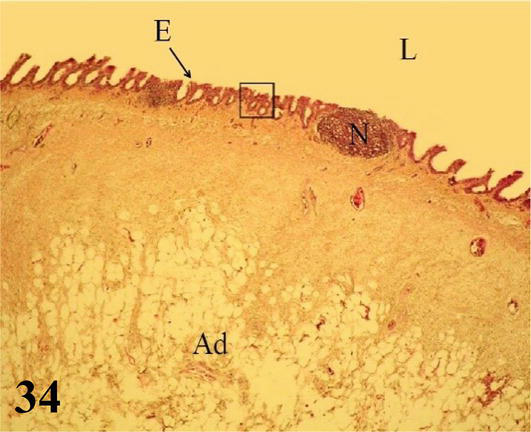
Figure 34.
Higher magnification of the inset of
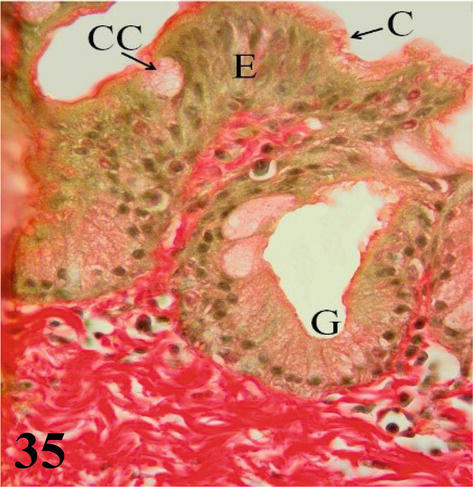
Figure 35.
Higher magnification of the inset of
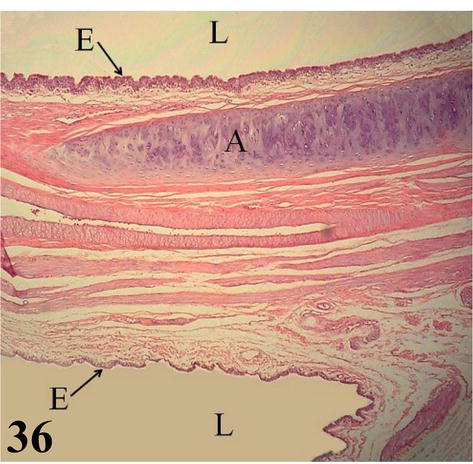
Figure 36.
Extrapulmonary primary bronchus. E: ciliated prismatic pseudostratified epithelium with goblet cells, both in the bronchial (upper) and saccular (lower) lumen (L); A: semi-ring of hyaline cartilage. Staining: H-E, 200×.
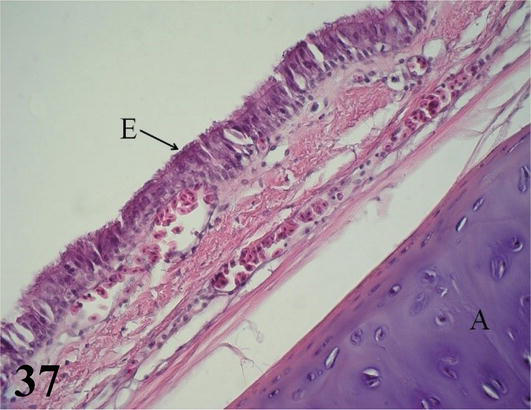
Figure 37.
Extrapulmonary primary bronchus. E: ciliated prismatic pseudostratified epithelium with goblet cells and A: hyaline cartilage of the bronchial semiring. Staining: H-E, 400×.
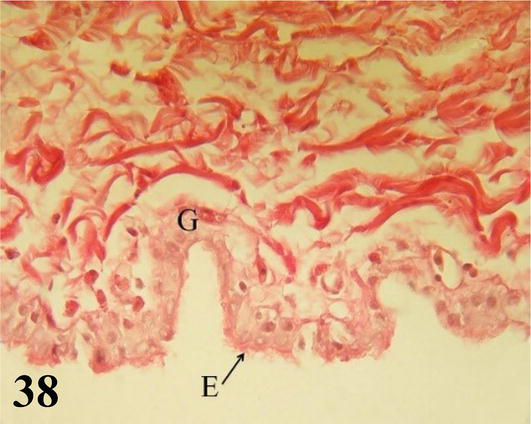
Figure 38.
Wall of the air sac. E: pseudostratified prismatic ciliated epithelium with goblet cells with simple tubuloalveolar glands that secrete mucus (G). Type I collagen tendon layer. Staining: Van Gieson, 1000×.
Extrapulmonary primary bronchi: They originate from the bifurcation of the syrinx. They present an extrapulmonary portion and another intrapulmonary or mesobronchus, they have a semi-ring of hyaline cartilage, whose ends are joined in the medial region by connective tissue and smooth muscle. Microscopically, the extrapulmonary primary bronchi maintain the histological characteristics of the tracheal wall (Figures 36 and 37). Mucosa with ciliated prismatic pseudostratified epithelium with goblet cells. Lamina propria of dense connective tissue with isolated lymphoid nodules. The thin submucosa of loose connective tissue. The adventitia is formed by semi-rings of hyaline cartilage, whose ends are joined in their medial region by smooth muscle and connective tissue. In this region, simple tubuloalveolar glands of mucous secretion are observed. Medially and at the bifurcation of the right and left bronchi, the mucosa constitutes the medial tympani-form membranes (Figures 32 and 33). The mucosa and submucosa at this level present numerous folds of different sizes made up of abundant connective tissue interspersed with smooth muscle tissue. Bundles of circularly arranged smooth muscle unite their ends, throughout their entire length. The extrapulmonary primary bronchi are limited lateroventrally by the pleural mesothelium that constitutes the thoracoabdominal air sacs (Figures 36 and 38). The portion seen from part of a thoracoabdominal air sac shows a ciliated prismatic pseudostratified lining epithelium with goblet cells (Figures 36 and 37).
Air sacks: The air sacs that are in contact with the extrapulmonary primary bronchi present a ciliated prismatic pseudostratified epithelium with cell goblet. However, in some sectors, it can be observed simple secretory tubuloalveolar glands have mucous membranes. Then a thin layer of loose connective tissue, followed by a thick tendon layer of collagen fibers type I (Figure 38).
5.2 Lung
In the lungs, we find primary intrapulmonary bronchi or mesobronchi, secondary bronchi distributed into three groups (dorsal, ventral, and lateral), parabronchi or tertiary bronchi, and the lung parenchyma made up of blood capillaries that surround the air capillaries that originate from the atria. This parenchyma is the place where hematosis occurs [14].
Primary intrapulmonary bronchi or mesobronchi: They run medially and craniocaudally throughout the lung until they contact the posterior air sacs through the lateral secondary bronchi. Microscopically: The mucosa with ciliated prismatic pseudostratified epithelium with goblet cells, presents numerous folds with shallow crypts. Lamina propria of dense connective tissue with simple tubuloalveolar glands of mucous secretion throughout its entire length. The submucosa of loose connective tissue with abundant bundles of smooth muscle fibers. The adventitia with hyaline cartilage is arranged in cartilaginous plates (not in semi-rings), a characteristic that gives its mucosa a scalloped appearance. These plates decrease in size until they disappear at the level of the secondary bronchi. This adventitia is surrounded by a layer of smooth muscle with a helical arrangement and its ventral region is surrounded by the pleural serosa (Figures 39 and 40).
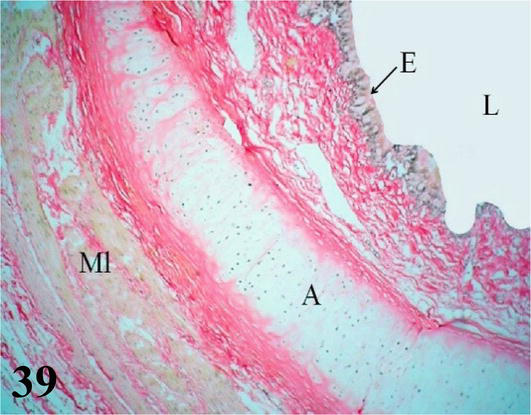
Figure 39.
Primary intrapulmonary bronchus or mesobronchus. E: Ciliated prismatic pseudostratified epithelium with goblet cells, with simple tubulo-alveolar glands and chorion of dense connective tissue; A: hyaline cartilage; ML: smooth muscle. Staining: Van Gieson, 200×.
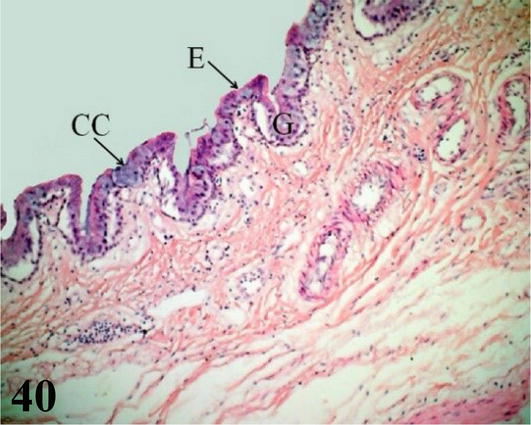
Figure 40.
Primary intrapulmonary bronchus or mesobronchus. E: ciliated prismatic pseudostratified epithelium with goblet cells (CC), deep crypts with simple tubulo-alveolar glands of mucus secretion (G). Staining: H-E, 200×.
Secondary bronchi: In each lung, the intrapulmonary primary bronchi or mesobronchi give rise to three groups of secondary bronchi: four dorsal secondaries, three ventral secondaries, and two lateral secondaries. These branch in the lung parenchyma and give rise to the parabronchi or tertiary bronchi (Figures 41 and 42). The mucosa of the secondary bronchi with a ciliated prismatic pseudostratified epithelium with goblet cells and shallow crypts. Lamina propria of dense connective tissue with simple mucus-secreting tubuloalveolar glands. Under the lamina propria, there is a thick layer of smooth muscle in a circular arrangement. In the adventitia of the secondary bronchi, small plates of hyaline cartilage are still observed that disappear before originating the parabronchi (Figures 41
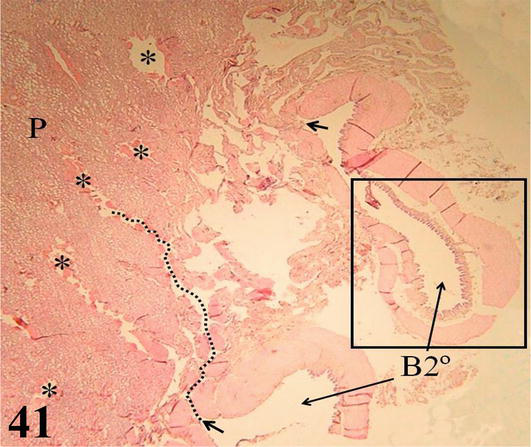
Figure 41.
Panoramic histological section showing the lung parenchyma (P), parabronchi (*); and its origin (arrows) from the secondary bronchi (B2°). The dashed line marks the luminal path. Stain: H-E, 12.5×.
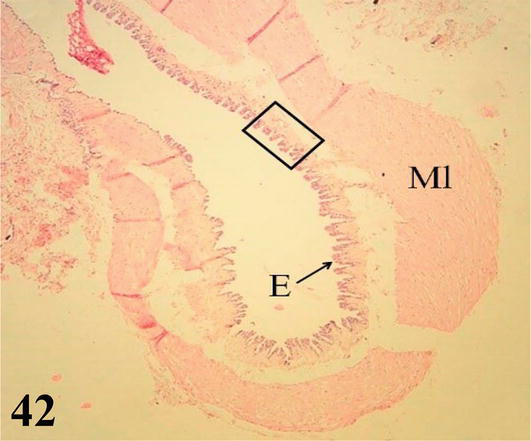
Figure 42.
Higher magnification of the inset of
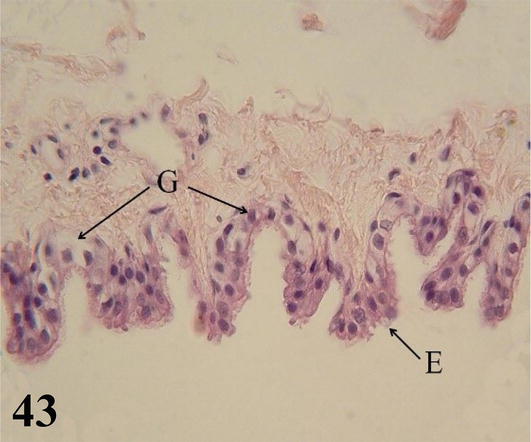
Figure 43.
Higher magnification of the inset of
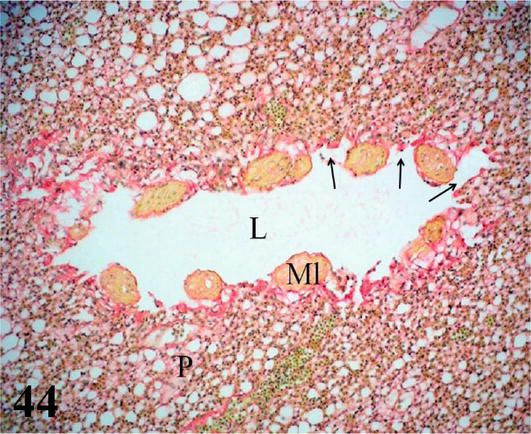
Figure 44.
Histological section of the lung parenchyma (P) and of a transversely arranged parabronchi or tertiary bronchus with its lumen (L); smooth muscle (Ml) and atria (arrows). Staining: Van Gieson, 200×.
Parabronchi or tertiary bronchi: The mucosa with a low cubic simple epithelium, lamina propria formed by a thin layer of connective tissue. The parabronchi are characterized by having bundles of smooth muscle with a helical arrangement, both clockwise and counterclockwise. Multiple cavities called atria originate from the parabronchi, which are continuous with the air capillaries, both lined by a simple flat epithelium (Figures 41 and 42). Surrounding the wall of the atrium and around the bundles of smooth muscle, there are accumulations of macrophages, among these structures are numerous lymphocytes, blood, and lymphatic capillaries.
Air capillaries: These originate and branch from the atria, which in turn, arise from the parabronchi. Microscopically, together with the blood capillaries, they constitute the lung parenchyma (Figure 45). With simple epithelium, which presents flat cells (type I pneumocytes) and others cubic (type II pneumocytes). These air capillaries are closely related to numerous blood capillaries, linked by a common basement membrane. In the wall of the atria, which originate from the parabronchi and constitute the entrance to the air capillaries, accumulations of fixed and free-form macrophages are observed in their lumen (Figure 45).
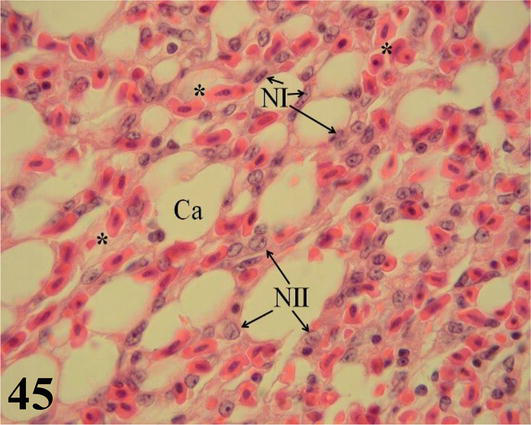
Figure 45.
Histological section of the lung parenchyma. Aerial capillaries (Ca) with flat cells (NI) and cuboidal cells (NII), surrounded by blood capillaries with nucleated erythrocytes (*). Staining: H-E, 1000×.
6. Female reproductive system
The female ostrich presents only the left ovary and oviduct. The ovary is large, cluster-shaped, which varies according to the seasons. It presents primordial, previtellogenic, vitellogenic, and atretic follicles. The oviduct has the following segments from the cephalad to the caudal: infundibulum, magnum, isthmus, uterus, and vagina, which empties into the cloaca. In them, there are folds of varying length, thickness, and number that compromise the mucosa and submucosa, with glands of mucous and serous secretion with the exception of the vagina, [15].
6.1 Ovary: ovarian follicles in different stages of maturation are arranged as a cluster
The ovarian cortex is covered in some areas by simple cuboidal epithelium, in others by ciliated simple prismatic epithelium and with pseudostratified prismatic epithelium (Figure 46). Beneath the epithelium is a thin layer of dense connective tissue, the albuginea (Figure 46). In the cortical stroma, follicles are observed in different stages of maturation, some of them, the largest, protrude on the surface. These are classified as primordial, previtellogenic, vitellogenic, and atresic. In primordial follicles, the oocyte with a vesicular nucleus is surrounded by a flat or low cuboidal epithelium of follicular cells that correspond to the granulosa layer. Externally to the basement membrane, some myofibroblasts are arranged (Figure 47). In previtellogenic follicles, the oocyte with a vesicular nucleus and the scant yolk is surrounded by a granulosa of cuboidal epithelium, a very conspicuous basement membrane and externally an undifferentiated theca vascularized with concentrically arranged myofibroblasts (Figures 46 and 47). In vitellogenic follicles, the oocyte with abundant yolk is surrounded by a granular layer of cuboidal epithelium, the theca interna vascularized, and the theca externa with abundant myofibroblasts arranged concentrically (Figures 48 and 49). Atretic follicles, which are smaller, have numerous lipid vacuoles.
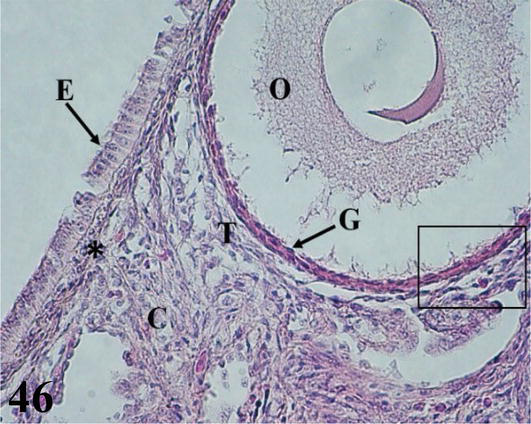
Figure 46.
Histological section of the ovary. E: simple ciliated prismatic ovarian epithelium; O: oocyte from a previtellogenic follicle; C: chorion of connective tissue; (*): albugínea; T: highly vascularized undifferentiated theca and G: granulosa. Staining: H-E, 400×.
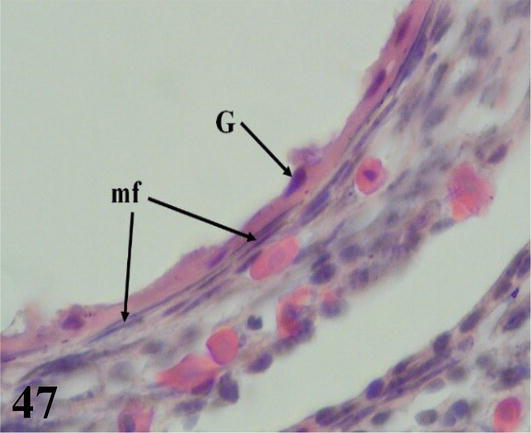
Figure 47.
Higher magnification of the inset of
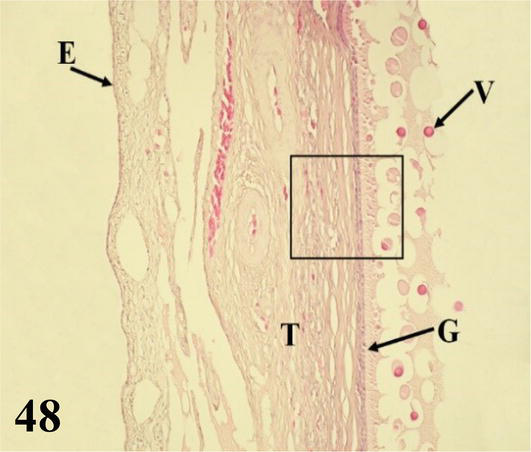
Figure 48.
Panoramic histological section of the wall of a vitellogenic follicle. E: ovarian epithelium; T: theca; G: granulosa and V: drops of yolk. Staining: H-E, 200×.
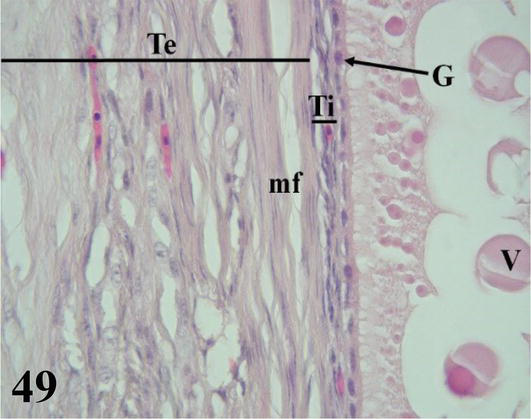
Figure 49.
Higher magnification of the inset of
6.2 Oviduct: it is a duct that has five segments
Infundibulum: it is the first segment of the oviduct, adjacent to the thin-walled ovary with long and numerous folds. The mucosa with ciliated pseudostratified prismatic epithelium with goblet cells and a lamina propria of dense connective tissue with abundant collagenous fibers. Interposed in the epithelium are simple tubuloalveolar (Figures 50 and 51) and simple alveolar glands of mucous secretion. No submucosa is seen. The muscularis, plexiform in arrangement, emits smooth muscle fibers that project through the mucosa of the folds. The serosa possesses dense connective tissue and is surrounded by mesothelium.
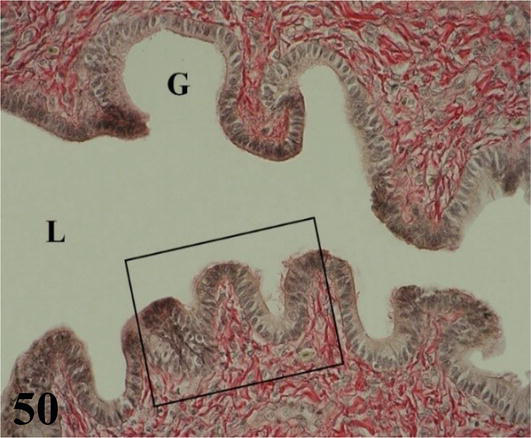
Figure 50.
Shows the mucosa of the infundibulum. L: lumen and G: simple alveolar glands of mucus secretion. Staining: Van Gieson, 400×.
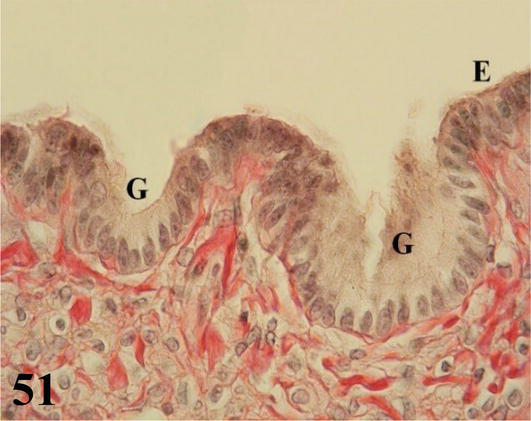
Figure 51.
Higher magnification of the inset of
Magnum: it is the longest segment of the oviduct, with the greatest diameter and numerous thick folds. Mucosa with ciliated prismatic pseudostratified epithelium with goblet cells. The lamina propria with abundant branched tubular glands of serous secretion (Figures 52 and 53). The loose connective tissue submucosa is highly irrigated and with smooth muscle fibers. The muscularis is arranged in an internal circular layer that is projected slightly by the wide submucosa of the folds and an external layer of plexiform arrangement. The serosa has loose connective tissue that is highly irrigated and is surrounded by mesothelium.
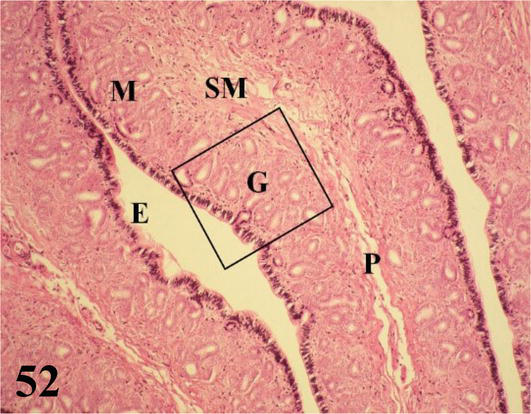
Figure 52.
The figure shows a fold of the magnum (P), the mucosa (M) with its epithelium (E), chorion of connective tissue with glands (G) and submucosa (SM). Staining: HE, 200×.
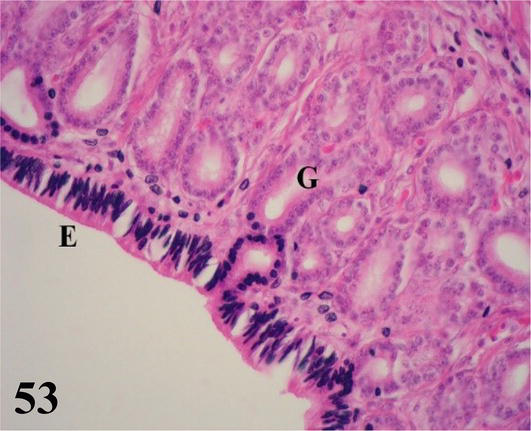
Figure 53.
The figure shows the inset of
Isthmus: it is the shortest segment of the oviduct, with the smallest diameter and thinnest wall, with numerous tortuous folds. Mucosa with ciliated pseudostratified prismatic epithelium with goblet cells. The lamina propria with abundant simple tubular glands of serous secretion (Figures 54 and 55). The submucosa, made of loose connective tissue, is well supplied with some smooth muscle fibers. The muscularis is arranged in an internal circular layer that is projected finely by the submucosa of the folds and an external layer of plexiform arrangement. The serosa has loose connective tissue with a high blood supply and is surrounded by mesothelium.
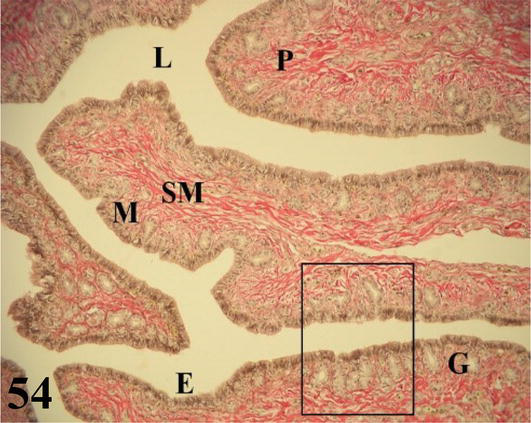
Figure 54.
The figure shows a fold of the isthmus (P), the mucosa (M) with its epithelium (E), chorion of dense connective tissue with glands (G), submucosa (SM) and lumen (L). Staining: Van Gieson, 200×.
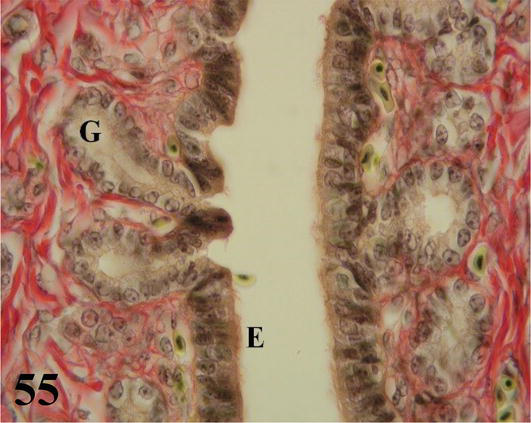
Figure 55.
Higher magnification of the inset of
Uterus: globose in shape with numerous long folds (Figure 56). Mucosa with ciliated pseudostratified prismatic epithelium with goblet cells. The lamina propria shows abundant branched tubular glands of serous secretion (Figures 56 and 57). The thin submucosa of loose connective tissue is well supplied with some smooth muscle fibers. The muscularis is arranged in an internal circular layer that emits fine projections toward the submucosa and an external layer with a plexiform arrangement. The serosa contains loose connective tissue with a high blood supply and is surrounded by mesothelium.
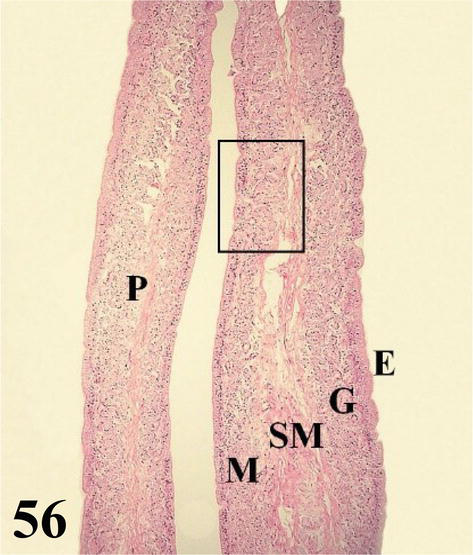
Figure 56.
The figure shows a fold of the uterus (P), the mucosa (M) with its epithelium (E), and chorion of connective tissue with numerous glands (G) and submucosa (SM). Staining: H-E, 200×.
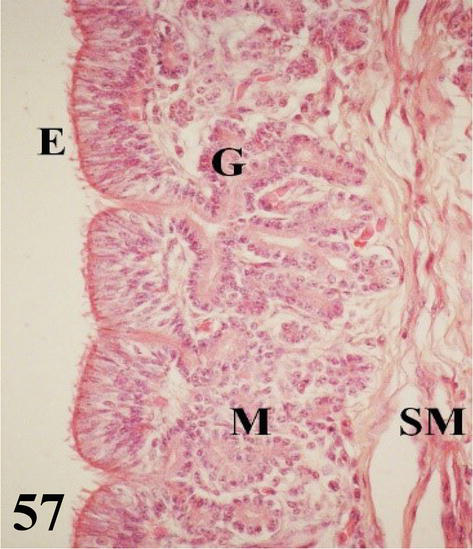
Figure 57.
Higher magnification of the inset of
Vagina: it is a short and narrow segment with a highly developed wall with numerous, long, and highly branched folds, with numerous villi forming a dense pattern (Figures 58 and 59). Mucosa presents ciliated pseudostratified prismatic epithelium with goblet cells (Figure 60). The dense connective tissue lamina propria is glandless. The arrangement of these folds gives rise to the formation of numerous spaces. The loose connective tissue submucosa is highly irrigated and bundled with smooth muscle fibers. The muscularis is arranged in a thick internal circular layer that projects notably through the submucosa of the folds and its ramifications, also reaching the villi and an external layer with a plexiform arrangement (Figures 58 and 59). An adventitia of loose connective tissue and a serosa that has loose connective tissue and is surrounded by mesothelium.
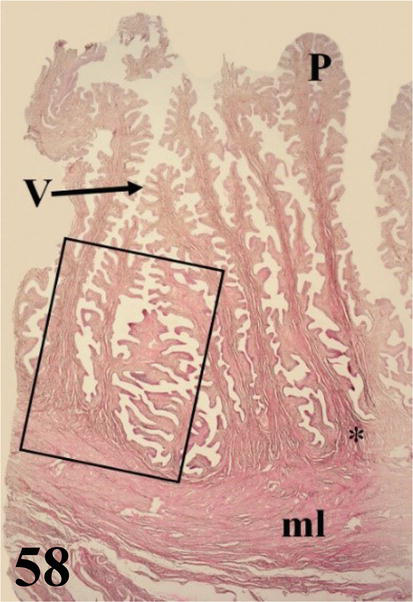
Figure 58.
Panoramic histological section of the vaginal wall with long, branched folds (P) and villi (V). Smooth muscle (ml) with the internal circular arrangement and external plexiform. The first is projected into the submucosa (*) of the folds and their ramifications. Staining: HE, 12.5×.
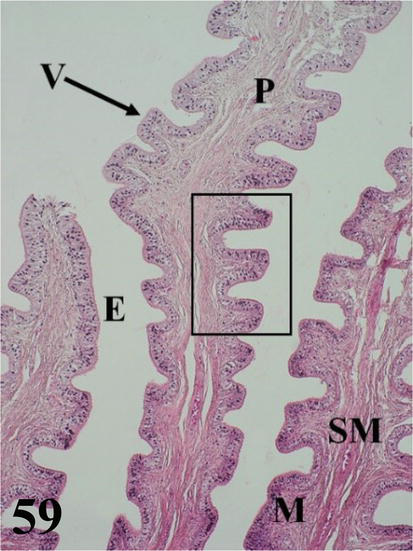
Figure 59.
Higher magnification of the inset of
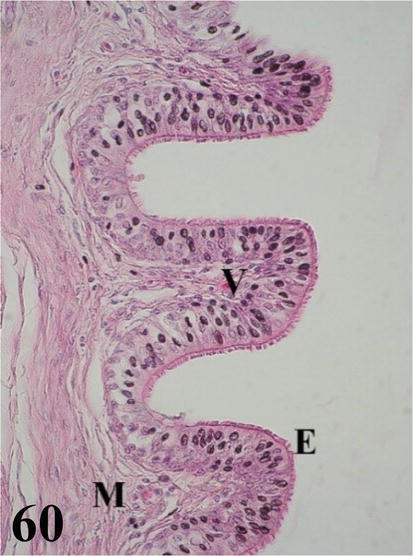
Figure 60.
Higher magnification of the inset of
7. Discussion and conclusions
At the level of the glands attached to the ostrich digestive system, certain histological differences are observed. The ostrich liver does not present significant differences from the liver of the chicken and other species [16]. However, the arrangement of the hepatocyte cords, both around the central vein and the portal spaces, is radiated, and a clear organization into lobules is not observed. In the ostrich proventriculus, we can point out that it does not present great differences from that of the chicken [16, 17]. Although the simple and branched tubular glands of the mucosa are poorly developed, they are very similar to the fundic glands of monogastric mammals (dogs) and non-true ruminants (camelids) [18, 19]. The ostrich and chicken submucosa are similar, presenting compound tubulo-alveolar glands [16, 17]. In the ostrich, it should be noted that these glands have a large number of parietal cells that secrete hydrochloric acid [12]. The pancreas of both the ostrich and the chicken present adenomeres of the tubulo-acinar type composed of serous secretion [16].
The epithelium of the esophageal mucosa in the ostrich is non-keratinized stratified squamous type, while in ruminants the mucosa presents a high degree of parakeratinization, which is slight in the horse [20]. The lamina propria in the ostrich shows numerous simple and branched tubuloalveolar glands of mucus secretion, arranged along its entire length [13]. The hen presents mucous secretion glands of the tubuloacinar type composed [21]. In other species, such as ruminants, horses and pigs, the mucosa lacks these glands [16]. The muscularis mucosa in the ostrich is observed as a thick layer of longitudinally arranged smooth muscle fibers. The hen has two layers, an internal one with a circular arrangement, formed by thin fascicles that are not always appreciable, and an external longitudinal one that is more developed [22]. The muscularis mucosa of domestic birds is formed only by a layer of longitudinally arranged smooth muscle fibers [21]. In the ostrich, the esophageal submucosa is very poorly developed, and as in the chicken, it lacks glands [21], while in the horse and ruminants, these glands are present, but they are only observed in the pharyngoesophageal junction [16, 20]. The muscularis of the ostrich esophagus, as in the chicken, has two layers of smooth muscle with an internal circular and external longitudinal arrangement [21, 22]. In the ruminant it is made up of striated musculature, which extends from the esophagus to the reticular groove; and in the case of the horse, this striated muscle comprises the proximal two-thirds of the esophagus, but gradually changes to smooth muscle in the distal third [20]. In the ostrich proventriculus, protein digestion begins through the secretion of pepsinogen and hydrochloric acid, given in part by the secretion of poorly developed simple and branched tubular fundic glands from the mucosa. The greatest contribution of hydrochloric acid, apparently, is the product of the tubuloalveolar glands composed of the submucosa [12]. In the ostrich, between the muscular layer and the serosa, we find a thick layer of compact connective tissue of parallel bundles of type I collagen; this tendinous layer surrounds the entire organ. Such a structure was also observed in the abomasum wall of an adult South American camelid, the llama (Lama glama) [18]. In the muscular ostrich stomach, the mucosa is covered by a thick layer with a horny appearance, which corresponds to the cuticle (glycocalyx), similar to that described in the adult rooster [22]. The muscle in the ostrich is similar to the chicken, it has three thick layers of smooth muscle fibers. However, in the chicken, in some sectors, only a layer of circularly arranged smooth muscle fibers can be seen [22].
Throughout the mucosa of the small intestine, long intestinal villi are observed, also described in the chicken. In ruminants, they are short and thick structures [16]. In ruminants and pigs, eosinophilic granule cells called Paneth cells [20] are observed at the bottom of the intestinal crypts. These cells were not observed in ostriches (Bezuidenhout) [23]. The muscularis mucosa in the ostrich duodenum and jejunum presents two layers of smooth muscle fibers, one internal circular and one external longitudinal, and three layers of muscle fibers are described in the ileum. On the other hand, in the hen, the muscularis mucosa consists of only one layer of smooth muscle fibers arranged longitudinally along the entire segment [22]. The submucosa in the ostrich, as in the chicken, is observed as a thin layer of loose connective tissue, it shows a greater thickness when it contains submucosal nerve plexuses or blood vessels and does not show glands [17, 21]. In contrast, in ruminants and horses, the submucosa shows tubuloalveolar glands, whose excretory ducts cross the muscularis mucosa and open at the bottom of the intestinal crypts. These glands are of the mucous type in ruminants and of the serous type in the horse, which are observed up to the jejunum [20].
At the level of the large intestine, in the cecum of the ostrich a spiral fold is described, formed by a nucleus of mucosa and submucosa, this plays an important role in the absorption of volatile fatty acids and other metabolites produced by the microbial fermentation of cellulose and hemicellulose (Bezuidenhout) [23]. The colon and rectum present longitudinal folds made up of mucosal and submucosal nuclei, as described for other domestic species [20]. The mucosa in the ostrich’s large intestine lacks villi and is formed by a simple columnar epithelium, similar to that described in the horse and in the ruminant [20]. The microscopic analysis carried out on the glands and the different segments of the ostrich digestive tract shows the general structure present in the wall of the digestive tract of vertebrates. However, certain differences are observed that may be the product of functional and adaptation changes. This histological analysis is a contribution not only to biology, but can also be very useful, especially to better understand the food and nutritional processes during ostrich breeding and management. In addition, they constitute a tool to face the diseases that are produced in them.
The conduction pathway from the larynx to the secondary bronchi is characterized by a mucosa lined by a ciliated prismatic pseudostratified epithelium with goblet cells, interrupted by crypts with simple mucus-secreting tubuloalveolar glands. With the exception of the extrapulmonary primary bronchi, which show this characteristic only in the mucosa of the medial region where the ends of the hyaline cartilage semi-rings of their adventitia converge. The remaining mucosa of these presents a ciliated prismatic pseudostratified epithelium with goblet cells without crypts or simple tubuloalveolar glands with interposed mucus secretion [14]. However, it should be noted that in the mucosa of the air sacs, crypts with simple tubuloalveolar glands of mucus secretion can be observed in some sectors. In these air sacs, the mucosa is reinforced with type I collagen fibers, constituting a tendinous layer [14]. This tendinous layer of type I collagen is observed in the ostrich proventriculus subserosa [13] and in the abomasum subserosa in the llama [18]. The ostrich syrinx is located at the bifurcation of the trachea, it presents three thickenings of the mucosa, the lateral tympaniform membranes, the medial tympaniform membranes that are analogous to the vocal folds of mammals [24], and the central membrane or pessulus, lined by ciliated prismatic pseudostratified epithelium with goblet cells, also observed and described by [25]. However, crypts with simple mucus-secreting tubuloalveolar glands interposed in their epithelium are also observed. In the ostrich pessulus, neither cartilage nor bone tissue is seen [14]. In the chicken, the pessulus presents a skeleton of cartilage or bone tissue [26, 27].
At the intrapulmonary level, the primary intrapulmonary bronchi or mesobronchi in the ostrich are lined by a ciliated prismatic pseudostratified epithelium with goblet cells, interrupted by crypts with simple mucus-secreting tubuloalveolar glands [14]. The hyaline cartilage of its adventitia is not arranged in semi-rings, but in cartilaginous plates that gradually decrease in size until they disappear at the level of the secondary bronchi. Unlike the ostrich, the intrapulmonary primary bronchi in the chicken have a simple ciliated prismatic epithelium, with numerous cells of mucus secretion [27]. The secondary bronchi in the ostrich present a ciliated prismatic pseudostratified epithelium with goblet cells, with deep crypts ending in simple mucus-secreting tubuloalveolar glands. They do not have hyaline cartilage; however, they show bundles of smooth muscle in a circular arrangement [14]. In the case of chickens, they present a simple ciliated prismatic epithelium with mucous cells [27]. The parabronchi in the ostrich present a low simple cuboidal epithelium over a thin layer of connective tissue. Then a layer of smooth muscle in a helical arrangement in both directions [14]. Various dilations or cavities called atria originate from this bronchial mucosa, which conduct air toward the air capillaries, also described by [28]. The parabronchi in the chicken present the same type of epithelium, with the abundant presence of atria [26, 27]. The air capillaries of the ostrich and the chicken are very similar, they present a simple epithelium, where flat cells (type I pneumocytes) and cuboidal cells (type II pneumocytes) are observed, similar to the epithelium of the lung alveoli in mammals [14, 27]. The mucosa of the conducting airways in the ostrich is adapted to generate large amounts of mucus. This mucus production capacity allows foreign particles to be retained, and temperature and humidity regulation of the inspired air. In this way, it protects this extensive mucosal surface from drying out, given the ecological characteristics where this species reproduces, develops, and lives. Through the comparative analysis, it was possible to establish similarities and differences with the chicken, with special emphasis on the mucous layer. Although many of its characteristics remind us of those observed in other bird species, some differences were observed.
The ovarian cortex in the ostrich is covered by a lining epithelium that changes from simple cuboidal to simple ciliated prismatic and in sectors to pseudostratified prismatic, whereas in the chicken it is simple cuboidal [8, 27]. In the ostrich, the follicles show a microscopic structure similar to those of the chicken [27, 29]. However, in the ostrich, follicles in different stages of development show myofibroblasts concentrically arranged in the theca surrounding the granulosa layer. Myofibroblasts are very noticeable in the theca interna of vitellogenic follicles [15].
In the ostrich oviduct, the mucosa of the folds of the infundibulum, magnum, isthmus, uterus, and vagina present a ciliated prismatic pseudostratified epithelium with goblet cells. In the hen, it is the same, except for the infundibulum which presents a simple ciliated prismatic epithelium [27]. In the chorion of the ostrich oviduct, at the level of the infundibulum, simple alveolar and tubuloalveolar glands of mucous secretion are observed. This type of gland is also found in abundance in the chorion of the esophageal and respiratory mucosa of these birds [14, 18]. The chorion of the magnum presents branched tubular glands of serous secretion. The chorion of the isthmus presents simple tubular glands of serous secretion. The chorion of the uterus has branched tubular glands of serous secretion, and the chorion of the vagina lacks glands. In the chicken, it is similar, however, the chorion of the infundibulum does not present glands and that of the vagina presents simple tubular glands of serous secretion [27]. These would contribute to forming the cuticle of the eggshell [30] and act as a sperm reservoir [10, 27]. In the ostrich, it is likely that the labyrinth of folds, ramifications, and villi present in the vagina acts as a reservoir for spermatozoa. The histological study of the reproductive system of the female will be one more tool that will allow for solving reproductive and management problems in ostrich breeding. In addition, it will be very useful to face the diseases that occur in it.
References
- 1.
Deeming DC. Introducción. In: El Avestruz: Biología, producción y sanidad. Zaragoza: Acribia; 2001. pp. 1-11 - 2.
Camiruaga, M. El Avestruz: Sistema de Producción en Chile. Fundación para la Innovación Agraria, Santiago, Chile, 2004 - 3.
Camiruaga M, Simoneti C. Avestruces, Sistema digestivo y su alimentación. 2003. Available from: http://www.faif.puc.cl - 4.
Bezuidenhout AJ. Anatomía. In: El Avestruz: Biología, producción y sanidad. Zaragoza: Acribia; 2001. pp. 13-50 - 5.
Angel CR. A review of ratites nutrition. Animal Feed Science and Technology. 1966; 60 :241-246 - 6.
Huchzermeyer F, W. Patología de Avestruces y otras Ratites. Madrid: Ediciones Mundi-Prensa; 2000 - 7.
Dyce KM, Sack WO, Wensing CJG. Anatomia Veterinaria. 2a ed. Ciudad de México: McGraw-Hill Interamericana; 2004 - 8.
Fudge MA. Laboratory Medicine Avian and Exotic Pets. California: Saunders; 2000 - 9.
Whittow GC. Sturkies Avian Physiology. 5a ed. Hawaii: Academic Press; 2000 - 10.
Hafez ESE. Reproducción e Inseminación Artificial en Animales. 7th ed. Ciudad de México: McGraw-Hill Interamericana; 2002 - 11.
Luna L. Manual of Histologic Staining Methods of the Armed Forces Institute of a Pathology. 3rd ed. New York: McGraw-Hill; 1968 - 12.
Illanes J, Fertilio B, Quijada M, Leyton V, Verdugo F. Histologic description of the annexed glands from the ostrich digestive system Struthio camelus var. domesticus. International Journal of Morphology. 2006; 24 (3):297-302 - 13.
Illanes J, Fertilio B, Chamblas M, Leyton V, Verdugo F. Histologic description of the different segments from the ostrich digestive system (Struthio camelus var. domesticus). International Journal of Morphology. 2006; 24 (2):205-214 - 14.
Illanes J, Leichtle J, Leyton V, Tapia LF, Fertilio B, Castro M. Histologic description of the different segments from the ostrich respiratory system (Struthio camelus var. domesticus). International Journal of Morphology. 2014; 32 (4):1325-1336 - 15.
Illanes J, Matus F, Leyton V, Venegas F, Fertilio B, Castro M. Histological description of the ostrich female reproductive system (Strutio camelus var. Domesticus). International Journal of Morphology. 2018; 36 (4):1252-1261 - 16.
Bacha WJ, Wood LM. Aparato Digestivo. In: Atlas color de Histología Veterinaria. Buenos Aires: Intermédica; 1991. pp. 111-150 - 17.
McLelland J. Atlas en color de Anatomía de las aves. En aparato digestivo. Buenos Aires: Interamericana; 1992. pp. 53-54 - 18.
Illanes J, Vigneaux IM, Adaro L, Olivares R, Mendoza J, Riveros V. Análisis morfológico comparado de la región fúndica entre un cánido (Canis familiaris) y un camélido suramericano (Lama glama). Revista chilena de anatomia. 1994; 12 (2):161-167 - 19.
Cummings J, Munnell J, Vallenas A. The mucigenous glandular mucosa in the complex stomach of two new-world camelids, the llama and guanaco. Journal of Morphages. 1972; 137 :71-110 - 20.
Stinson AW, Calhoun ML. Sistema Digestivo. En: Dieter Dellmann, H. Histología Veterinaria. Zaragoza, Acribia, 1993. pp. 239-256 - 21.
Dieter Dellmann H, Eurell J. Textbook of Veterinary Histology. 5th ed. Philadelphia: Lippincott Williams & Wilkins; 1998 - 22.
Delhon GA, Fernández-Surribas J, Gagna G, Lacolla D, von Lawzewitsch I. Aparato digestivo de las aves domésticas. En: Lecciones de histología veterinaria. Hemisferio Sur, Buenos Aires, 1984. pp. 45-111 - 23.
Cooper R, Mahroze K. Anatomy and physiology of the gastrointestinal tract and growth curves of the ostrich ( Struthio camelus ). Animal Science Journal. 2004;75 :491-498 - 24.
Nickel R, Schummer AY, Seiferle E. Anatomy of the Domestic Birds. Berlin: Verlag Paul Parey; 1977. p. 65 - 25.
Yildiz H, Bahadir A, Akkoç A. A study on the morphological structure of syrinx in ostriches (Struthio camelus). Anatomia, Histologia, Embryologia. 2003; 32 (3):187-191 - 26.
Sisson S, Grossman JD. Anatomía de los animales domésticos. Tomo II. 5a ed. México D. F: Masson; 2000 - 27.
Bacha WJ, Bacha LM. Atlas Color de Histología Veterinaria. 2a ed. Buenos Aires: Intermédica; 2001 - 28.
Maina JN, Nathaniel C. A qualitative and quantitative study of the lung of an ostrich, Struthio camelus . Journal of Experimental Biology. 2001;204 (Pt. 13):2313-2330 - 29.
Kimaro WH. An immunohistochemical and ultrastructural study of the ovary of the immature ostrich ( Struthio camelus ) [M.Sc. thesis]. Pretoria: Department of Anatomy and Physiology, Faculty of Veterinary Science, University of Pretoria; 2005. Available from:https://repository.up.ac.za/bitstream/handle/2263/23731/Complete.pdf?sequence=3 - 30.
Fujii S. Histological and histochemical studies on the oviduct of the domestic fowl with special reference to the region of uterovaginal juncture. Archivum Histologicum Japonicum. 1963; 23 :447-459