 Open access peer-reviewed chapter
Open access peer-reviewed chapter
Abstract
Fast-growing poplars have the potential to improve the biomass supply required for the transition to bio-based economies in the Nordic-Baltic region. As early successional trees, poplars are efficient biomass producers in relatively short rotations, when high-yielding, climate-adapted clones are available for commercial deployment. In Sweden, poplar breeding focused on adapting Populus trichocarpa to the Swedish climate by crossing parents from distant populations along latitudinal and maritime-continental clines on the Pacific coast of North America. Clonal trials with progeny from these crosses were established in the Nordic-Baltic region. Elite individuals in terms of stemwood production were used to identify candidate genes for adaptation to local photoperiod and climate in the region. The next breeding cycle utilized the elite individuals in the clonal trials to generate a training population. Genomic selection of the progeny in the training population will facilitate early selection of poplar clones for commercial deployment in the Nordic-Baltic region and reduce the time required for successive plant breeding cycles.
Keywords
- adaptive phenology
- alleles
- biomass
- bud burst
- bud set
- candidate genes
- forest industry
- genomic prediction model
- genomic selection
- heritability
- hybrid poplar
- genetic marker density
- nucleotides
- Populus trichocarpa
- stem wood
- training population
- woody biomass
- yield
1. Introduction
When breeding poplars for northern latitudes, significant attention is given to the adaptation of their phenological traits to regional climates [1, 2, 3, 4]. The most important phenological events that distinguish the period of dormancy and active growth are bud flushing, growth cessation, and bud set [5]. The timing of these events is crucial for an individual genotype to adjust its growth efficiently to the temperature and photoperiod in a given region [6]. Thus, different populations within the species’ geographic range are set under intense selection pressure to adapt to latitudinal and altitudinal gradients of daylight and temperature. Individuals with phenologies that do not match their environments are at a higher risk of being frost-injured, especially during the transition periods from autumn to winter and winter to spring. However, prolonged growth can also lead to a competitive advantage by allowing for greater utilization of the growing season [7, 8]. As a result, the phenology of each population is shaped by its adaptation to the local and regional environments but still contains significant genetic variation for phenological traits [9].
Adaptation of a species to new geographic regions relies on the variation within and between populations.
The Nordic-Baltic forest industry and energy sector have used native aspen (
For over 30 years, the Swedish University of Agricultural Sciences has been running a climate adaptation program for
2. Potential of hybrid poplars in the Nordic-Baltic wood market
Forests offer multiple ecosystem services and are a crucial resource in the transition to biobased economies. This is especially true for the Nordic-Baltic region, which has abundant forest resources and well-established wood industries. Consequently, the region is a major hub for education, research and development, testing, and production of innovative wood-based products, including textiles, biofuels, biochemicals, and engineered wood products for the construction sector [12]. However, the increasing demand for wood as a raw material is likely to be in conflict with the environmental role of forest ecosystems as carbon sinks and guardians of biodiversity [13]. Additionally, in a warming climate, forests may be more susceptible to severe droughts, fires, storms, diseases, and insect outbreaks, which could further reduce the availability of woody biomass [14].
Expanding the area of short-rotation poplar plantations is a viable option for increasing the stocks of industrial wood. Poplar plantations are usually designed for high biomass production enabling relatively frequent financial returns for the growers. This type of plantations increase the value of low-quality and marginal arable land, making them attractive investment targets for capital investors. Many EU countries have reported their anticipation of heightened private investments in fast-growing tree species [15].
In the Nordic-Baltic region, the current area of poplar plantations is approximately 5000 ha. Hybrid aspen is planted on an additional 12,000 ha. Consequently,
Among the Baltic countries, Lithuania has fewer forest resources than Estonia and Latvia but more set aside arable land suitable for conversion to poplar plantations. Lithuania and Latvia are now major producers of wood-based panels in the region and supply global companies such as IKEA (Figure 1). Estonia and Latvia also produce large quantities of pellets, mainly for export to Denmark, Great Britain, and the USA (Figure 2) [19]. Poplar wood is ideal for making products like oriented strand boards (OSB) and medium-density fiberboards (MDF). These products represent a significant market opportunity for poplar wood in the region. While poplars are not likely to be used for veneer in the Baltic countries, they can be utilized for inner layers in plywood, while birch veneer is used for the surface layers.
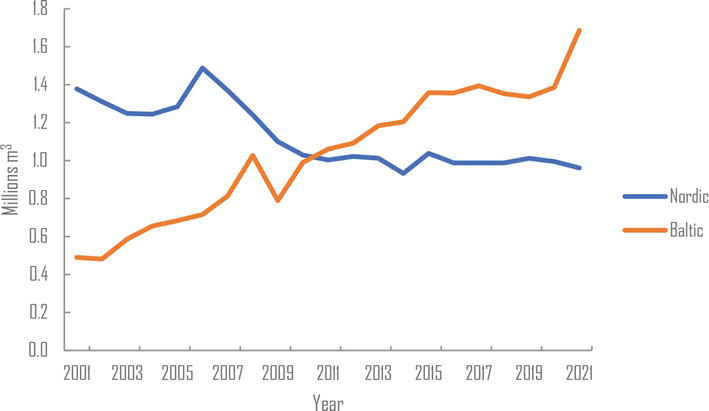
Figure 1.
Particle board production in the Nordic-Baltic countries since 2001. Nordic countries—Denmark, Finland, and Sweden. Baltic countries—Estonia, Latvia, and Lithuania [
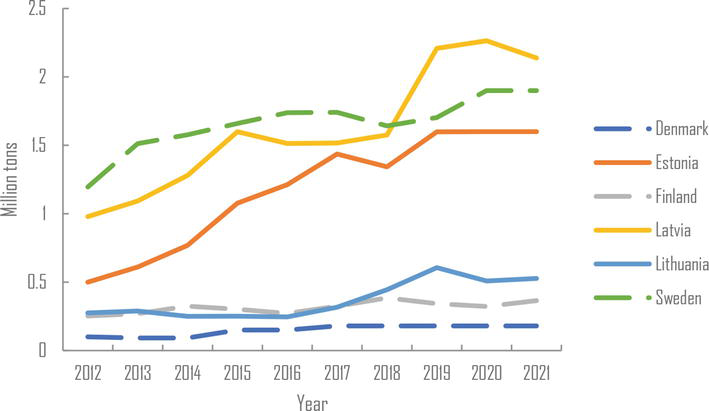
Figure 2.
Production of wood pellets in six Nordic-Baltic countries for the period 2012–2021 [
In the Nordic-Baltic countries, poplar plantations were initially established as a part of land conversion programs and for bioenergy purposes. Sweden has seen an increase in the area of hybrid poplar and hybrid aspen plantations in the past few decades, reaching approximately 4000 ha [20]. At the same time, the area of willow coppice has decreased from 14,000 ha in 2001 to just over 7000 h in 2016 [21]. One of the main reasons for the decrease in willow coppice is the significant increase in cereal prices since 2007. Similarly, the decline in Italian poplar cultivation has also been related to the profitability of alternative land use [22]. The potential of poplars as a raw-material resource in the region depends directly on the expansion of plantations on arable or other non-forested land. Fertile agricultural land is more likely to be utilized for cultivation of food and feed crops, while arable land and grasslands of poor quality, or fields that are difficult to access with agricultural machinery can be available for poplar plantations. However, grasslands can be a valuable form of land use, particularly on organic soils, providing high biodiversity and significant carbon storage potential [23]. Poplar plantations can also expand by replacing forest tree species planted on arable land during the early conversions in the mid-twentieth century. This replacement is more of a resilience issue in a changing climate rather than an opportunity to increase biomass production significantly. Despite these limitations, it has been suggested that between 1.8 and 4.6 million ha of land in the Nordic-Baltic region could be available for conversion to poplar plantations [24, 25].
3. Biology and ecology of Populus
The
The taxonomy of
Black cottonwood (
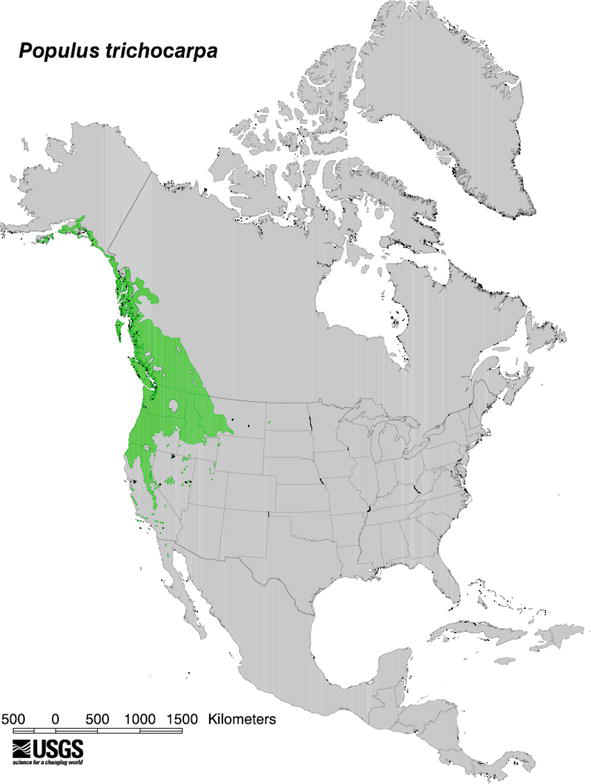
Figure 3.
Natural distribution of
4. Nordic-Baltic poplar cultivation practices
Operative poplar cultivation in the Nordic-Baltic region was first introduced during the early 1990s. Typically, plantations were established at a 3 × 3 m spacing (≈ 1000 trees ha−1), aiming at a rotation of 20 years (Figure 4). In forest-dominated areas of central Sweden, planting densities ranging from 1100 to 1700 trees per hectare are also used, with higher densities serving as a buffer against losses due to the browsing pressure. The great majority of Swedish poplar plantations were established with clone ‘OP42’ (

Figure 4.
A 19-year-old trial with climate-adapted black cottonwood (

Figure 5.
Seven-year-old poplar plantation in Lithuania established at 3 × 2 m spacing. The clone used was the Italian AF7, which suffered from repeated frost injuries on this site. This stand was established using 1.5 m long poles and harvested a couple of months after the photo was taken. Lithuanian poplar plantations are established without the application of herbicides and have been certified with FSC standards since August 2023 [
The intensity of work to prepare land for planting varies depending on its previous use but typically involves using herbicides and cultivating the soil. Various types of planting materials are used, ranging from rooted plants (either bare root or containerized) to cuttings of varying lengths, including unrooted 2-year-old poles. In some cases, cuttings are planted through degradable plastic mulch that can be replaced by biodegradable mulch paper. However, most Nordic and Baltic entrepreneurs rely on using robust planting material and mechanical weed control after planting.
Poplar growers in the Nordic-Baltic region face specific challenges due to the relatively short growing season and cold winters. Poplar clones with better frost hardiness are required compared to the material available from central/southern European breeding programs [3]. Moreover, the market for poplar wood is limited, which makes it difficult to justify cost-intensive tending. To overcome these obstacles, poplar growers need to optimize management by combining denser initial spacing, more robust plant material, and less frequent mechanical weeding. Additional challenges are present due to high browsing and rodent populations, particularly in small, isolated poplar plantations in forest-dominated landscapes.
Site and clone selection are the two most important decisions in poplar silviculture. Poplars perform best on deep, moist, well-drained, and light-textured soils. Dry sandy soils and waterlogged soils are unsuitable as they can result in poor growth and establishment. However, the productivity of dry soils can be improved by fertilization and irrigation with wastewater, while waterlogged soils can be ditched. Wet sites are typically located in lowland terrain on organic soils with abundant ground vegetation. There is a relative abundance of these sites in the Nordic-Baltic region, so farmers prioritize these areas when converting land into tree plantations. However, there is a high likelihood of frequent frost, high browsing pressure, vole populations, and serious competition from herbaceous vegetation occurring on these sites. Combined, these factors represent a severe challenge to poplar growers who must employ simultaneous measures to be successful, including heavy soil preparation (mounding, for example), herbicide treatments, frequent mechanical weeding, and sometimes plant protection.
Deploying highly productive clones is the most efficient measure to increase the yields of poplar plantations. However, even for a small set of productive clones, there can be substantial variations in clonal performance [8, 30]. Some clones, such as ‘OP42’, are generalists and can grow well on many different soils. ‘OP42’ is outperformed by better-adapted clones on some sites or towards the edges of its deployment range. Planting clone mixtures is sometimes recommended to resolve the uncertainties related to clonal performance in different environments. This approach has both advantages and disadvantages, and the optimization of mixtures regarding the number and growth pattern of the clones in a mixture is necessary [31].
The Nordic-Baltic poplar plantations established with 1000 trees ha−1 yield total biomass at an average of 10 tons ha−1 year−1 dry weight over a 20-year rotation. This corresponds to 8 tons ha−1 year−1 of stem biomass or 20–25 m3 ha−1 year−1 [25]. During the second half of a rotation, individual trees face intense competition, which can impact their vitality, especially during periods of summer drought [32]. Therefore, thinning is commonly utilized to promote diameter growth and vitality in the remaining trees. The recommended final stocking of Nordic-Baltic poplar stands is 650–800 trees per hectare. The timing of thinning will depend on the initial density and site productivity, usually occurring between ages 8–14 when the mean height is 15–18 m. However, thinning poplar stands pose a risk as heavy machines can damage their shallow root system. The final harvest should be performed between December and April to ensure successful resprouting from stumps and roots. Harvesting during May–September may result in the failure of the coppice regeneration [33].
5. Current genetic resources and past breeding activities
In the 1970s, interest in poplars and other fast-growing species increased due to energy crises. Several trials were established in southern Sweden, mainly with material imported via Germany and Holland. Karlsson et al. [36] evaluated some of these trials, suggesting that hybrids with species from the
In the late 1980s, Ilstedt [3] conducted a test on a collection of Belgian
Recognizing the potential of
6. Adaptive phenology is one of the breeding goals
Flowering and bud flushing of poplar trees co-occur early in spring and are regulated mainly by temperature (Figure 6) [45]. In contrast, the timing of growth cessation is primarily governed by changes in day length and is known to have a significant genetic component (Figure 7) [46]. Bud formation is initiated after growth cessation and is also affected by the temperature [5]. Due to a rapid temperature cline along increasing latitudes in the Nordic-Baltic region, one of the primary goals of poplar climate adaptation is to identify genotypes with optimal timing of active growth initiation and termination.
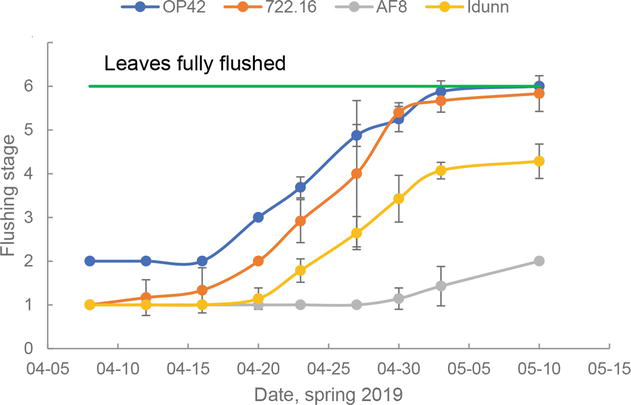
Figure 6.
Stages of bud burst (1—buds swelled but no green leaves visible; 6—leaves fully flushed with an initial shoot increment >10 mm) for four clones grown in a common garden in Uppsala, central Sweden (59°N). Clone ‘OP42’ (
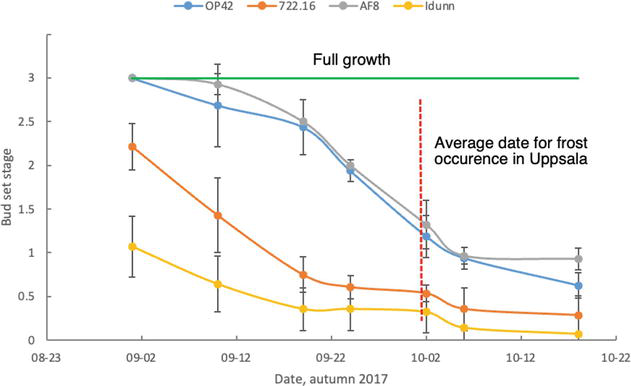
Figure 7.
Stages of bud set (3—fully growing shoots; 1.5—initiation of bud formation; 0.5—bud formed but not matured; 0—matured bud) for four clones grown in a common garden in Uppsala, Central Sweden (59°N). The growth cessation and bud set were inventoried in the year of planting. The first-year growth can be prolonged compared to the already established plants. Data from the second growing season (2018) was not useful due to an extremely dry summer causing atypic bud set in many clones, and the inventory was not possible in 2019 due to height of trees (up to 8 m). For example, the Icelandic clone ‘Idunn’ would normally initiate a bud set already in July. Clones ‘OP42’ and ‘AF8’ are late to set bud and will be at risk of frost damage at this latitude. Early frosts can induce frost damage to the top shoot. Cold temperatures also inhibit bud formation leading to incomplete dormancy and the risk of frost damage during the winter. The trial included 40 clones planted in 10 single-tree plots (blocks), and the number of observations for the four presented clones was ‘OP42’ = 4, ‘722.16’ = 6, ‘AF8’ = 7, and ‘Idunn’ = 7. Error bars are standard deviations.
These traits are controlled by many genes with minor effects, e.g., they are quantitatively inherited. New progeny genotypes obtained by cross-pollination of individuals from distant heterotic groups will display a spectrum of phenological characteristics. Consequently, breeding for climate adaptation involves such crosses to achieve hybrid vigor, which is a common strategy in European poplar breeding programs [47, 48]. In the Nordic-Baltic region, an example of a specific breeding goal for autumn phenology is to select clones that exhibit active growth throughout August but also have a rapid bud formation and maturation in September. This allows the new poplar clones to take advantage of favorable growing conditions with optimum temperature and rainfall in August while ensuring frost hardiness. A good benchmark for the timing of bud set in poplar clones can be developed from the Swedish horticultural growing zones (Figure 8), where bud formation should be accomplished approximately 1 week earlier within each successive growing zone. However, it is important to keep in mind that the timing of frost occurrence within each growing zone can vary significantly, so local conditions and premises for clone deployment should be considered. Nonetheless, having a reasonable estimate of bud set for a particular clone and zone is essential, as the timing for a clone is likely to differ across different growing zones.
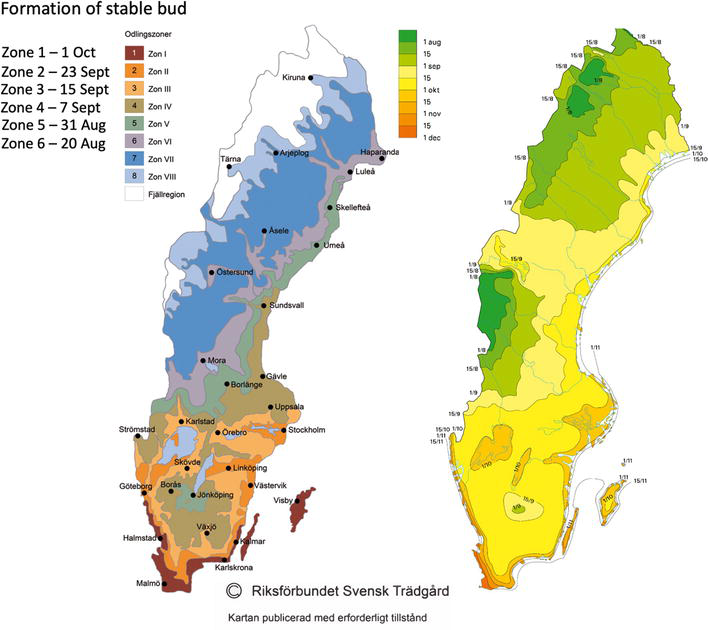
Figure 8.
The latest date for forming a stable bud is autumn within the six growing zones in Sweden. The horticultural growing zones (left) are used as guidance for clone deployment based on the timing of bud set (map from the Swedish Garden Association, with permission). The range of average dates for frost occurrence (right) is an essential benchmark for evaluating clone hardiness (map from the SMHI [
7. Genetic architecture of photoperiodic traits
The genetic architecture of photoperiodic traits is complex, involving many genes with minor effects. The alleles either decrease or increase the additive effect of the different genes. Many loci in the poplar genome have been identified through QTL mapping that can explain variation in bud set traits [50, 51, 52, 53, 54, 55, 56, 57, 58, 59]. The identified genes regulate the active growth and dormancy cycle based on the perception of photoperiodic and dormancy signals [60]. The circadian rhythm regulates the active growth cycle, a biological process that displays an endogenous adjustable oscillation of approximately 24 h. External cues such as daylight and temperature determine the circadian rhythm in a specific environment. The interplay between various photoreceptors and the circadian clock is important for induced growth cessation and bud set in deciduous trees [61, 62]. The large number of candidate genes involved in adaptation to photoperiod and climate highlights the quantitative nature of these traits (Figure 9).
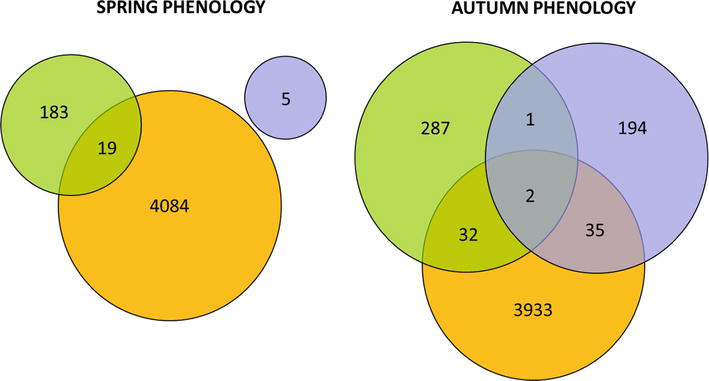
Figure 9.
Overlap between candidate genes of spring and autumn phenology in populations of Populus trichocarpa. Orange: wild accessions from Northwest coast of North America [
7.1 Circadian clock
Circadian clock genes are genes whose protein products generate and regulate circadian rhythms in different organisms. In poplar, several circadian clock genes have been identified:
7.2 Heritability of critical daylength
Typically, a day shorter than the specific length required for growth evokes growth cessation and bud set. The critical day length is when about 50% of individuals of a particular clone are setting buds [60]. Fabbrini et al. [46] and Rohde et al. [5] have demonstrated that traits related to the timing of growth cessation in black poplars (
7.3 Molecular markers for selection of operational varieties
Thus, abundant genomic information is available for many species in the genus
8. Timeline for the implementation of molecular markers in selection
Genomic selection (GS), initially proposed by Meuwissen et al. [85], is a method that uses genetic markers with genome-wide coverage and phenotypic information from a training population to develop a model that can be used to predict the breeding values for individuals from a breeding population with only genotype information available. GS has many advantages that increase the method’s utility in modern plant breeding. First, GS can be used to perform early selection, often even at the seedling stage, without the need for extensive field testing and screenings. This facilitates early selection, which reduces the time required for successive plant breeding cycles and thus contributes to an increased genetic gain per unit of time. Second, the estimated marker effects from GS are precise, and unbiased prior marker selection is avoided [86, 87]. GS is particularly suitable for trees due to their long generation times and traits characterized by a genetic architecture consisting of many underlying genes, each with relatively small effects. Furthermore, traits of interest for tree breeding are often expensive to phenotype or are displayed late in the life cycle, further increasing the utility of GS.
Implementing a genomic selection program for tree breeding encompasses two stages; the first stage relies on a “training population” consisting of individuals that are both phenotyped and genotyped. The combined genotype and phenotype data are used to develop a predictive model that links variation in genetic markers to variation in phenotypes of interest. Training populations are usually derived from existing progeny trials or breeding populations, where previously selected ‘elite’ parents have been crossed, and their progenies have been extensively tested in field trials. Breeding populations in forest trees usually contain 1000–2000 individuals and have effective population sizes (Ne) in the range of 30–100. The larger the training population, while keeping Ne in the appropriate range, the more precisely marker effects are estimated, ultimately resulting in a more accurate predictive model [88].
GS is then employed by genotyping a large number of individuals that form ‘selection candidates’, usually consisting of full- or half-sib families derived from individuals that are part of the training population. The genotype information is used with the prediction model to estimate genomics-based genotypic values (GEGVs) for all selection candidate individuals. Top-ranking individuals from the selection candidates, based on the GEGVs, are then selected and used to establish the next generation of the breeding program. To further enhance testing, top-ranked selection candidates can also be clonally propagated and tested in clonal trials, where elite clones are eventually selected for operational plantation, especially for tree species that rely on clonal deployment, such as
Large training population sizes have thus far characterized studies of GS in forest trees, and large numbers of genetic markers have also typically been used, especially when compared to GS studies in crops. Recent studies in forest trees include a diverse array of species, including eucalypts [89, 90], spruces [91, 92], pines [93], and
GS’s utility depends on fundamental population and quantitative genetics aspects as well as more practical and logistical aspects relating to resource allocation and cost-benefit considerations. The accuracy of a genomic prediction model is the most important aspect of the success of GS, and many factors immediately impact GS model accuracy. The first important aspect is the effective size of the training population (Ne), which directly influences the extent of linkage disequilibrium (LD), which, in turn, dictates the necessary genetic marker density needed for successful model building. The following essential aspect is phenotyping accuracy in the training population, which sets an upper limit to how much variation can be explained by marker effects. Similarly, the heritability and genetic architecture of the traits of interest are essential, and traits with high heritabilities generally have higher prediction accuracies. Finally, the statistical methods used for model building and deployment are also relevant, and this is currently an area of active research. Simulation studies have partially assessed all of these factors and have provided some general guidelines for implementing GS in forest trees [86, 89].
The extent of LD is perhaps the most important aspect influencing the accuracy of GS, and LD, in turn, depends on the Ne of the training population. The extent of LD directly affects the marker density needed for successful implementation of GS, and marker density has been shown to tightly scale with Ne of the training population, where larger populations need more markers. The level of LD between markers and the unknown quantitative trait loci (QTLs) controlling traits of interest can be increased by reducing Ne. Previous studies have shown that to maintain reasonable levels of LD while maintaining sufficient genetic diversity to sustain long-term genetic gains in breeding, Ne in the range of 40–100 is typically recommended. These Nes typically correspond to census sizes of 100–200 related individuals; for example, most advanced GS populations in
The design of the training population depends on the actual breeding strategy that is adopted. Training populations are established using trees from existing progeny trials based on crosses among elite parents. The relatedness between the training population and selection of candidates is another factor that has great importance for the success of GS. Increasing the genetic relationships between the training population and selection candidates also leads to higher prediction accuracies in a manner similar to the effects of reducing Ne [98]. Similarly, the genetic architecture underlying trait variation has significant consequences for the accuracy of GS. A small number of loci controlling a large proportion of the phenotypic variation allows for more variation to be captured compared to more complex genetic architectures involving more loci, each with a smaller effect size [99]. GS accuracy drops with the increasing number of QTLs contributing to a trait, and this effect is more pronounced when marker density is low or in populations with large Ne [86]. Traits heritability only has minor impacts on GS accuracy as long as the training population size is large enough to estimate marker effects adequately.
Based on the issues outlined in the preceding paragraphs, the prospects for implementing GS in
9. Conclusions
Extensive collaboration between forestry sector and academia in the Nordic-Baltic region over the past decades has resulted in readiness for successful hybrid poplar breeding programs in the region. The existing collections of
Acknowledgments
The authors will greatly acknowledge our partners from the forestry sector who have been driven in innovations in the procurement of woody raw material in the Nordic-Baltic region: Anders Ekstrand from Södra Skog AB, Lars-Georg Hedlund from Södra Latvia SIA and Mindaugas Šilininkas from Euromediena UAB. This chapter has been written within three following projects during the last decade: (1) an EU project within the EUREKA program Eurostars E! 8443—SnowTiger during 2014–2016, (2) grant number 942-2016-20001 from Swedish Research Council FORMAS “Climate-adapted poplar through more efficient breeding and better tools for matching genotype and site—developing the poplar bio-economy market in Sweden and the Baltic Region” during 2016–2021. Finally, (3) this chapter was finalized within the “NutriBiomass4LIFE” project (LIFE17/ENV/LV000310) co-financed by the Swedish Energy Agency (P-45082-1).
References
- 1.
Jonsson TH, Oskarsson U. Shoot growth strategy of 29 black cottonwood ( Populus trichocarpa ) clones. Icelandic Agricultural Sciences. 2007;20 :25-36 - 2.
Sverrisson H. A Review of the Icelandic Poplar Breeding Program. Iceland: Iceland Forest Service; 2017 - 3.
Ilstedt B. Genetics and performance of Belgian poplar clones tested in Sweden. Forest Genetics. 1996; 3 (4):183-195 - 4.
Ilstedt B. Anpassning av Populus trichocarpa, jättepoppel, till svenskt klimat. In: Proceedings from a Poplar Seminar at the Department of Short Rotation Forestry. Uppsala: Swedish University of Agricultural Sciences; 2005 - 5.
Rohde A et al. Bud set in poplar—Genetic dissection of a complex trait in natural and hybrid populations. New Phytologist. 2011; 189 (1):106-121 - 6.
Ronnberg-Wastljung AC et al. Phenotypic plasticity in Populus trichocarpa clones across environments in the Nordic-Baltic region. Scandinavian Journal of Forest Research. 2022; 37 (1):1-5 - 7.
Adler A et al. Variation of growth and phenology traits in poplars planted in clonal trials in Northern Europe-implications for breeding. Bioenergy Research. 2021; 14 (2):426-444 - 8.
Karacic A et al. An analysis of poplar growth and quality traits to facilitate identification of climate-adapted plant material for Sweden. Bioenergy Research. 2021; 14 (2):409-425 - 9.
Farmer REJ. The genecology of Populus. In: Stettler R, Bradshaw HD Jr, Heilman PE, Hinckley TM, editors. Biology of Populus and Its Implications for Management and Conservation. NRC Research Press, National Research Council of Canada: Ottawa, ON, Canada; 1996. pp. 33-55 - 10.
Dickmann DI, Kuzovkina J. Poplars and Willows of the World, with Emphasis on Silviculturally Important Species. Rome, Italy: FAO/IPC; 2008. p. 134. Working Paper IPC/9-2, in FAO/IPC Poplars and Willows in the World - 11.
Soolanayakanahally RY et al. Timing of photoperiodic competency causes phenological mismatch in balsam poplar ( Populus balsamifera L.). Plant Cell and Environment. 2013;36 (1):116-127 - 12.
Hetemäki L. The outlook for Nordic-Baltic forest bioeconomy to 2030. In: Liuhto K, editor. The Forest Industry around the Baltic Sea Region:Future Challenges and Opportunities. Helsinki: Centrum Balticum Foundation; 2020. pp. 14-24 - 13.
Lundmark T. Skogen räcker inte till—hur ska vi prioritera? In Swedish. In: Future Forests Rapportserie 2020. Umeå: U.o.A. Sciences; 2020. p. 24 - 14.
Schlyter P et al. Assessment of the impacts of climate change and weather extremes on boreal forests in northern Europe, focusing on Norway spruce. Climate Research. 2006; 31 (1):75-84 - 15.
IPC. Synthesis of country progress reports. The International Commission on poplars and other fast-growing trees sustaining people and the environment (IPC). Twent-sixth session. FO:IPC/2021/Inf2. 2021: FAO, Rome. 2021 - 16.
Estonian Cell. 2023. About us. Available from: https://www.estoniancell.ee/en/about-us/facts-figures/ [Accessed: September 30, 2023] - 17.
Jensen V. Snabbväxande lövträd kan öka avkastning och sprida dina risker. In Swedish. SkogsAktuellt, Independent magazine on forestry in Sweden. 2023;3 - 18.
FAOSTAT. 2023. Available from: https://www.fao.org/faostat/en/#data/FO [Accessed: August 1, 2023] - 19.
Hedlund L.-G. CEO of Södra Latvia SIA, personal communication. 2023 - 20.
Adler A et al. Country Report 2016-2019 from National Commission of Fast-Growing Trees in Sweden. Uppsala: National Commission of Fast-Growing Trees in Sweden; 2020. p. 18 - 21.
Xu X, Mola-Yudego B. Where and when are plantations established? Land-use replacement patterns of fast-growing plantations on agricultural land. Biomass & Bioenergy. 2021; 144 :105921. pp. 1-10 - 22.
Pra A, Pettenella D. Investment returns from hybrid poplar plantations in northern Italy between 2001 and 2016: Are we losing a bio-based segment of the primary economy? Rivista di Economia Agraria. 2019; 74 (1):49-71 - 23.
Hungate BA et al. The economic value of grassland species for carbon storage. Science Advances. 2017; 3 (4):e1601880. p. 8 - 24.
Stener L-G, Rytter L, Beuker E, Tullus H, Lutter R. Hybrid Aspen and Poplars in the Baltic Sea Region and Iceland—Results from a Questionnaire and a Literature Review. Uppsala, Sweden: Skogforsk; 2019 - 25.
Adler A et al. Lignin-first biorefining of Nordic poplar to produce cellulose fibers could displace cotton production on agricultural lands. Joule. 2022; 6 (8):1845-1858 - 26.
Eckenwalder J. Systematics an evolution of Populus. In: Stettler R, Bradshaw HD Jr, Heilman PE, Hinckley TM, editors. Biology of Populus and its Implications for Management and Conservation. Ottawa, ON, Canada: NRC Research Press, National Research Council of Canada; 1996. pp. 7-32 - 27.
Little EL Jr. Atlas of United States Trees, Vol. 1, Conifers and Important Hardwoods: U.S. Department of Agriculture Miscellaneous Publication No. 1146, 200 maps. Public domain to the map via Wikimedia Commons. 1971. 9 p. Available from: https://upload.wikimedia.org/wikipedia/commons/d/da/Populus_trichocarpa_range_map_1.png - 28.
Dickmann DI. An overview of genus Populus. In: Dickmann DI, Isebrands JG, Eckenwalder JE, Richardson J, editors. Poplar Culture in North America. Ottawa, ON K1A 0R6, Canada: NCR Press, National Research Council of Canada; 2001. pp. 1-42 - 29.
Silininkas M. Personal communication with Mindaugas Silininkas, CEO Euromediena UAB. 2023 - 30.
Stener LG. Tillväxt, vitalitet och densitet för kloner av hybridasp och poppel i sydsvenska fältförsök, in Arbetsrapport från SkogForsk nr 717. Uppsala. 2010; 50 pp - 31.
Elferjani R, DesRochers A, Tremblay F. Effects of mixing clones on hybrid poplar productivity, photosynthesis and root development in northeastern Canadian plantations. Forest Ecology and Management. 2014; 327 :157-166 - 32.
Karacic A, Verwijst T, Weih M. Above-ground woody biomass production of short-rotation populus plantations on agricultural land in Sweden. Scandinavian Journal of Forest Research. 2003; 18 (5):427-437 - 33.
Jónsson B. Coppice silviculture of black cottonwood for production of wood chips—Effects of harvesting date on regeneration and yield. In: The WoodBio Conference. Reykjavik, Iceland; 2017 - 34.
Johnsson A. Föreningen för växtförädling av skogsträd—1936-1958. In Swedish, F.f.v.a. skogsträd, Editor. Uppsala, Sweden. 1959 - 35.
Eriksson H. Yield of aspen and poplars in Sweden. In: Perttu K, editor. Ecology and Management of Forest Biomass Production Systems. Swedish: Department of Ecology and Environmental Research, Swedish University of Agricultural Sciences; 1984. pp. 393-419 - 36.
Karlsson B, Werner M, Stener LG. Resultat från två klonförsök med poppel, in Arbetsrapport från SkogForsk nr 319. Uppsala. 1996. 16 p - 37.
Stener LG. Resultat från sydsvenska klontester med poppel. Skogforsk. 2004 - 38.
Stener LG, Westin J. Early growth and phenology of hybrid aspen and poplar in clonal field tests in Scandinavia. Silva Fennica. 2017; 51 (3):5656. p. 22 - 39.
Christersson L. Biomass production of intensively grown poplars in the southernmost part of Sweden: Observations of characters, traits and growth potential. Biomass & Bioenergy. 2006; 30 (6):497-508 - 40.
Stanton BJ, Neale DB, Li S. Populus breeding: From the classical to the genomic approach. In: Jansson S, Bhalerao RP, Groover AT, editors. Genetics and Genomics of Populus. Vol. 8. 2010. pp. 309-348 - 41.
Steenackers MS, de Uyper C, Michiels B. Breeding and selection of poplars for durable resistance to Melampsora larici-Populina. In: First IUFRO Rusts of Forest Trees Working Party Conference, August 2-7 1998. Saariselkä, Finland: Finnish Forest Research Institute, Rovaniemi Research Station; 1998 - 42.
Ilstedt B. Breeding strategy for poplar Populus trichocarpa, jättepoppel, till svenskt klimat. Norwegian Journal of Agricultural Sciences. 1994:39-45 - 43.
Richards TJ et al. Quantitative genetic architecture of adaptive phenology traits in the deciduous tree, Populus trichocarpa (Torr. and Gray). Heredity. 2020;125 (6):449-458 - 44.
Vico G et al. Consistent poplar clone ranking based on leaf phenology and temperature along a latitudinal and climatic gradient in Northern Europe. Bioenergy Research. 2021; 14 (2):445-459 - 45.
Olson MS et al. The adaptive potential of Populus balsamifera L. to phenology requirements in a warmer global climate. Molecular Ecology. 2013;22 (5):1214-1230 - 46.
Fabbrini F et al. Phenotypic plasticity, QTL mapping and genomic characterization of bud set in black poplar. BMC Plant Biology. 2012; 12 :47. p. 16 - 47.
Ducros ET. Breeding strategies with poplars in EUROPE. Forest Ecology and Management. 1984; 8 (1):23-39 - 48.
Steenackers J et al. Poplar diseases, consequences on growth and wood quality. Biomass & Bioenergy. 1996; 10 (5-6):267-274 - 49.
Swedish Meteorological and Hydrological Institute (SMHI). 2017. Available from: https://www.smhi.se/data/meteorologi/temperatur/genomsnittliga-datum-for-den-forsta-hostfrosten-1.4074 [Accessed: August 5, 2023] - 50.
Frewen BE et al. Quantitative trait loci and candidate gene mapping of bud set and bud flush in Populus . Genetics. 2000;154 (2):837-845 - 51.
Rohde A et al. PtABI3 impinges on the growth and differentiation of embryonic leaves during bud set in poplar. Plant Cell. 2002; 14 (8):1885-1901 - 52.
Bohlenius H et al. CO/FT regulatory module controls timing of flowering and seasonal growth cessation in trees. Science. 2006; 312 (5776):1040-1043 - 53.
Ingvarsson PK et al. Nucleotide polymoirphism and phenotypic associations within and around the phytochrome B2 locus in European aspen ( Populus tremula , Salicaceae). Genetics. 2008;178 (4):2217-2226 - 54.
Ruonala R et al. CENL1 expression in the rib meristem affects stem elongation and the transition to dormancy in Populus . Plant Cell. 2008;20 (1):59-74 - 55.
Jimenez S et al. Phylogenetic analysis and molecular evolution of the dormancy associated MADS-box genes from peach. BMC Plant Biology. 2009; 9 :81. p. 12 - 56.
Li Z et al. Dormancy-associated MADS genes from the EVG locus of peach Prunus persica (L.) Batsch have distinct seasonal and photoperiodic expression patterns. Journal of Experimental Botany. 2009;60 (12):3521-3530 - 57.
Ibanez C et al. Circadian clock components regulate entry and affect exit of seasonal dormancy as well as winter hardiness in Populus trees. Plant Physiology. 2010;153 (4):1823-1833 - 58.
Rohde A, Bastien C, Boerjan W. Temperature signals contribute to the timing of photoperiodic growth cessation and bud set in poplar. Tree Physiology. 2011; 31 (5):472-482 - 59.
Ruttink T et al. A molecular timetable for apical bud formation and dormancy induction in poplar. Plant Cell. 2007; 19 (8):2370-2390 - 60.
Cooke JEK, Eriksson ME, Junttila O. The dynamic nature of bud dormancy in trees: Environmental control and molecular mechanisms. Plant Cell and Environment. 2012; 35 (10):1707-1728 - 61.
Savolainen O, Lascoux M, Merila J. Ecological genomics of local adaptation. Nature Reviews Genetics. 2013; 14 (11):807-820 - 62.
Olsen JE. Light and temperature sensing and signaling in induction of bud dormancy in woody plants. Plant Molecular Biology. 2010; 73 (1-2):37-47 - 63.
Evans LM et al. Population genomics of Populus trichocarpa identifies signatures of selection and adaptive trait associations. Nature Genetics. 2014;46 (10):1089-1096 - 64.
McKown AD et al. Genome-wide association implicates numerous genes underlying ecological trait variation in natural populations of Populus trichocarpa . New Phytologist. 2014;203 (2):535-553 - 65.
Apuli R-P et al. The genetic basis of adaptation in phenology in an introduced population of Black Cottonwood ( Populus trichocarpa, Torr. & Gray). BMC Plant Biology. 2021;21 (1):317 - 66.
McWatters HG, Devlin PF. Timing in plants—A rhythmic arrangement. FEBS Letters. 2011; 585 (10):1474-1484 - 67.
Howe GT et al. Evidence that the phytochrome gene family in black cottonwood has one PHYA locus and two PHYB loci but lacks members of the PHYC/F and PHYE subfamilies. Molecular Biology and Evolution. 1998; 15 (2):160-175 - 68.
Olsen JE et al. Ectopic expression of oat phytochrome A in hybrid aspen changes critical daylength for growth and prevents cold acclimatization. Plant Journal. 1997; 12 (6):1339-1350 - 69.
Hall D et al. Adaptive population differentiation in phenology across a latitudinal gradient in European Aspen ( Populus tremula , L.): A comparison of neutral markers, candidate genes and phenotypic traits. Evolution. 2007;61 (12):2849-2860 - 70.
Wang J et al. A major locus controls local adaptation and adaptive life history variation in a perennial plant. Genome Biology. 2018; 19 :72 - 71.
Zhang M, Suren H, Holliday JA. Phenotypic and genomic local adaptation across latitude and altitude in Populus trichocarpa . Genome Biology and Evolution. 2019;11 (8):2256-2272 - 72.
Jansson S, Douglas CJ. Populus : A model system for plant biology. Annual Review of Plant Biology. 2007;58 :435-458 - 73.
Bradshaw HD et al. Emerging model systems in plant biology: Poplar ( Populus ) as a model forest tree. Journal of Plant Growth Regulation. 2000;19 (3):306-313 - 74.
Tuskan GA et al. The Genome of Black Cottonwood, Populus trichocarpa (Torr. & Gray). Science. 2006;313 (5793):1596-1604 - 75.
Ingvarsson PK. Nucleotide polymorphism and linkage disequilbrium within and among natural populations of European Aspen ( Populus tremula L., Salicaceae). Genetics. 2005;169 (2):945-953 - 76.
Kelleher CT et al. A physical map of the highly heterozygous Populus genome: Integration with the genome sequence and genetic map and analysis of haplotype variation. Plant Journal. 2007;50 (6):1063-1078 - 77.
Olson MS et al. Nucleotide diversity and linkage disequilibrium in balsam poplar ( Populus balsamifera ). New Phytologist. 2010;186 (2):526-536 - 78.
Slavov GT et al. Genome resequencing reveals multiscale geographic structure and extensive linkage disequilibrium in the forest tree Populus trichocarpa . New Phytologist. 2012;196 (3):713-725 - 79.
Wang J et al. Natural selection and recombination rate variation shape nucleotide polymorphism across the genomes of three related Populus species. Genetics. 2016;202 (3):1185 - 80.
Geraldes A et al. A 34K SNP genotyping array for Populus trichocarpa : Design, application to the study of natural populations and transferability to otherPopulus species. Molecular Ecology Resources. 2013;13 (2):306-323 - 81.
Schilling MP et al. Genotyping-by-sequencing for Populus population genomics: An assessment of genome sampling patterns and filtering approaches. PLoS One. 2014;9 (4):e95292 - 82.
Faivre-Rampant P et al. New resources for genetic studies in Populus nigra : Genome-wide SNP discovery and development of a 12k Infinium array. Molecular Ecology Resources. 2016;16 (4):1023-1036 - 83.
Isabel N, Lamothe M, Thompson SL. A second-generation diagnostic single nucleotide polymorphism (SNP)-based assay, optimized to distinguish among eight poplar ( Populus L .) species and their early hybrids. Tree Genetics & Genomes. 2013;9 (2):621-626 - 84.
Chhetri HB et al. Multitrait genome-wide association analysis of Populus trichocarpa identifies key polymorphisms controlling morphological and physiological traits. New Phytologist. 2019;223 (1):293-309 - 85.
Meuwissen THE, Luan T, Woolliams JA. The unified approach to the use of genomic and pedigree information in genomic evaluations revisited. Journal of Animal Breeding and Genetics. 2011; 128 (6):429-439 - 86.
Grattapaglia D, Resende MDV. Genomic selection in forest tree breeding. Tree Genetics & Genomes. 2011; 7 (2):241-255 - 87.
Nakaya A, Isobe SN. Will genomic selection be a practical method for plant breeding? Annals of Botany. 2012; 110 (6):1303-1316 - 88.
Hayes BJ et al. Accuracy of genomic selection: Comparing theory and results. Matching genetics and environment: a new look at an old topic. In: Proceedings of the 18th Conference of the Association for the Advancement of Animal Breeding and Genetics, September 28–October 1, 2009. Barossa Valley, South Australia, Australia; 2009. pp. 34-37 - 89.
Resende MDV et al. Genomic selection for growth and wood quality in Eucalyptus: Capturing the missing heritability and accelerating breeding for complex traits in forest trees. New Phytologist. 2012; 194 (1):116-128 - 90.
Tan B et al. Evaluating the accuracy of genomic prediction of growth and wood traits in two Eucalyptus species and their F-1 hybrids. BMC Plant Biology. 2017; 17 :110 - 91.
El-Dien OG et al. Prediction accuracies for growth and wood attributes of interior spruce in space using genotyping-by-sequencing. BMC Genomics. 2015; 16 :370 - 92.
Ratcliffe B et al. A comparison of genomic selection models across time in interior spruce (Picea engelmannii x glauca) using unordered SNP imputation methods. Heredity. 2015; 115 (6):547-555 - 93.
de Almeida Filho JE et al. The contribution of dominance to phenotype prediction in a pine breeding and simulated population. Heredity. 2016; 117 (1):33-41 - 94.
Kirst M, Resende M, Munoz P, Neves L. Capturing and genotyping the genome-wide genetic diversity of trees for association mapping. In: BMC Proceedings, IUFRO Tree Biotechnology Conference 2011: From Genomes to Integration and Delivery, Arraial d’Ajuda, Bahia, Brazil. 2011. pp. 1-2 - 95.
Alves FC et al. Accelerating forest tree breeding by integrating genomic selection and greenhouse phenotyping. Plant Genome. 2020; 13 (3):e20048 - 96.
Kerr RJ et al. Simulation of hybrid forest tree breeding strategies. Canadian Journal of Forest Research. 2004; 34 (1):195-208 - 97.
Perez-Enciso M, Rincon JC, Legarra A. Sequence- vs. chip-assisted genomic selection: Accurate biological information is advised. Genetics Selection Evolution. 2015; 47 :43 - 98.
Beaulieu J et al. Genomic selection accuracies within and between environments and small breeding groups in white spruce. BMC Genomics. 2014; 15 :1048 - 99.
Grattapaglia D, do Amaral Diener PS, dos Santos GA. Performance of microsatellites for parentage assignment following mass controlled pollination in a clonal seed orchard of loblolly pine ( Pinus taeda L.). Tree Genetics & Genomes. 2014;10 (6):1631-1643 - 100.
Silva-Junior OB, Grattapaglia D. Genome-wide patterns of recombination, linkage disequilibrium and nucleotide diversity from pooled resequencing and single nucleotide polymorphism genotyping unlock the evolutionary history of Eucalyptus grandis. New Phytologist. 2015; 208 (3):830-845