Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
One of the most damaging insects to coniferous trees, especially pine and fir, is Pityokteines curvidens. It may cause significant tree death and alter the composition and structure of the forest. Early detection of forest stands vulnerable to bark beetle infestations and can help reduce the harmful effects of possible infestations. The main objective of the study was to compare the susceptibility maps of P. curvidens, utilizing the analytical hierarchy process and maximum entropy approaches. The study was conducted in the fir forests of the Küre Forest Enterprise in Türkiye. P. curvidens susceptibility maps were created using the stand structure, forest stand form, site index, slope, aspect, elevation, NDVI, and solar radiation as the eight key criteria. The accuracy of the maps based on the analytical hierarchy process and maximum entropy approaches was assessed using the receiver operating characteristic curves. The most crucial variables in both models’ predictions of P. curvidens susceptibility were found to be stand structure, NDVI, and elevation. Comparing the maps produced by the two models, the MaxEnt model gave more precise predictions than AHP. This study may make it easier for decision-makers to create susceptibility maps for fir engraver bark beetles.
Faculty of Forestry, Department of Forestry Engineering, Kastamonu University, Kastamonu, Turkey
Gonca Ece Özcan
Faculty of Forestry, Department of Forestry Engineering, Kastamonu University, Kastamonu, Turkey
Korhan Enez
Faculty of Forestry, Department of Forestry Engineering, Kastamonu University, Kastamonu, Turkey
*Address all correspondence to: fsivrikaya@kastamonu.edu.tr
1. Introduction
Bark beetles (Coleoptera: Curculionidae, Scolytinae), one of the most significant forest pests [1], are natural disturbances that have severe epidemics in the last century [2]. While only 2% of these species can cause epidemics in forests [3], endemic populations in small tree groups, and epidemic populations in large forest stand areas cause tree deaths [4, 5]. Host tree resistance prevents increasing of bark beetle populations [6]. However, attacks are easier on trees weakened by biotic and abiotic factors [7]. Epidemics significantly affects forest ecosystem values and causes deterioration of the functioning and characteristics of natural ecosystems [8, 9].
Abies (Miller) genus, which has two species and four subspecies in Türkiye [10], covers a total area of 511.703 ha [11]. Kazdağı fir (Abies nordmanniana ssp. equi-trojani (Asc. & Sint. ex Boiss.) Coode & Cullen), one of the endemic fir taxa for Türkiye, is economically and ecologically important and is very sensitive to climate variables [12]. Due to its declining population size and limited geographic range, the species has been given endangered status by the International Union for Conservation of Nature [13].
One of the most important of the bark beetles that damage fir is the fir engraver beetle (Pityokteines curvidens (Germar, 1824) Coleoptera: Curculionidae: Scolytinae) [14]. Although the main host of the species is Abies alba, it can also cause significant infestations in other Abies species (A. equi-trojani, A. sibirica, A. cephalonica, A. balsamea, A. bornmülleriana, A. borisii-regis, A. cilicica, A. nordmanniana, A. firme, A. sachalinensis, and A. fraseri). In addition, Pinus silvestris, P. brutia, P. strobus, Pseudotsuga menziesii, Picea orientalis, P.abies, Larix decidua, L. kaempferi, Cedrus libani, are other hosts [15, 16]. Abies ssp., Cedrus ssp., Pinus nigra, P. brutia and Picea pungens are hosts in Türkiye [17, 18, 19, 20].
P. curvidens, one of the important pests of fir forests in the Western Black Sea region of Türkiye [21, 22], prefers forests and monoculture stands where suitable habitat conditions are not available [23]. Although it is generally a secondary pest, it also becomes a primary pest depending on the increase in its population [24, 25]. It has been recorded that bark beetles caused a total damage of 2,353,057 m3 in 1987–2004 in this region [22].
Although it is not possible to prevent natural disasters, risks can be reduced by taking the necessary precautions [26]. Conducting risk assessment and modeling of the damages caused by bark beetles will help decision-makers in managing forest ecosystem [27]. In addition, there is a need for accurate and reliable estimation of pest distributions in forest ecosystems in today’s forestry [28]. Many different modeling techniques are used to estimate the distributions of bark beetle. Models that predict the geographical distribution of species are important for various applications in the field of ecology and conservation [29, 30]. Maximum Entropy (MaxEnt), which is one of the machine learning methods, is widely preferred in modeling species distributions and habitat suitability [31]. Spatial distribution of Dendroctonus mexicanus [28], potential distribution of D. ponderosae, D. brevicomis [32], Ips calligraphus, I. grandicollis [33], I. mannsfeldi [34] and I. aminutus [35], according to current and future climate scenarios and tree death events caused by bark beetles were estimated by MaxEnt [36].
The analytic hierarchy process (AHP), one of the multicriteria decision analysis methods (MCDA), is a widely used method in solving complex decision-making problems in many different disciplines [37]. While AHP is used effectively in forestry [38, 39, 40, 41, 42, 43], studies on bark beetles are limited [44, 45, 46].
In this study, AHP and MaxEnt approaches were used to predict the susceptibility of P. curvidens in Kazdağı fir forests. The accuracy of the models was calculated by the receiver operating characteristic (ROC) analysis. In the theory part of this book chapter, the bark beetle’s infestation, their effects on the ecosystem, and the importance of bark beetle susceptibility maps, according to AHP and MaxEnt models were explained. In the material and methods section, the conceptual framework, dataset, model parameters, and models used were described. In the result and discussion section, the model results obtained according to two different models were evaluated and the results were compared with the literature. In the last section, current bottlenecks were mentioned and suggestions for models were presented.
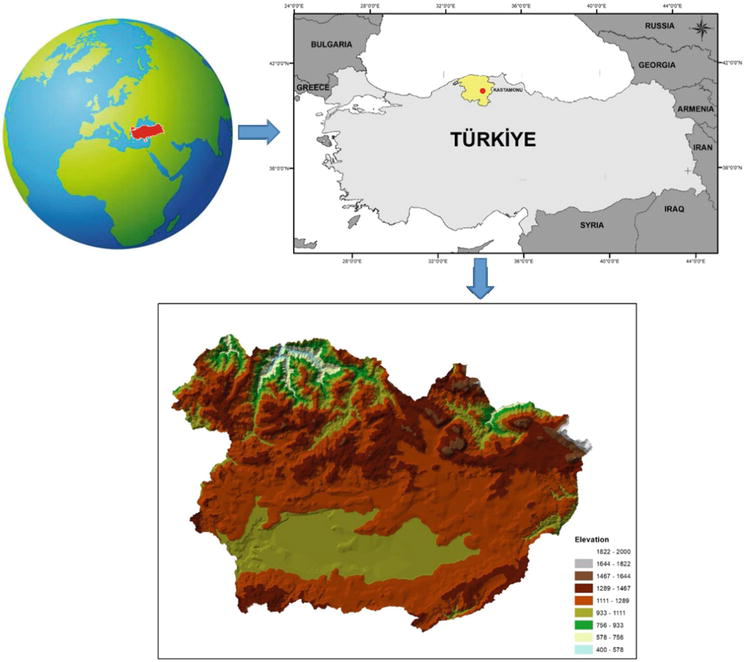
Küre Forest Enterprise (FE), a case study area, is situated on the İsfendiyar mountain in the Kastamonu Regional Directorate of Forestry in the Western Black Sea region at 41° 42′ N and 33° 42′ E (Figure 1). 52% of the Küre FE (73694.9 ha) is covered with forest areas. The dominant tree species in the study area are Abies ssp., Fagus ssp., Quercus ssp., Pinus ssp., Alnus ssp., Carpinus ssp., and Ulmus ssp. Pure, and mixed fir forest stands in the study area is 29911.6 ha and 21% of the fir forests (6217.1 ha) were under attack by Pityokteines curvidens. In the study area, fir stands were infested intensively by Pityokteines curvidens between 2008 and 2017. The average elevation of the study area above sea level is 960 meters. The study area is in the Black Sea climate zone. The relative humidity level is high in all seasons. The weather is cool in summer and not very cold in winter, but rainy.
Figure 1.
The study area.
2.2 Parameters selection and dataset
A review of the literature helped determine the criteria that should be used to assess a forest stand’s vulnerability to Pityokteines curvidens infestation. Eight parameters were thought to be essential indicators for forecasting P. curvidens epidemics: stand structure, forest stand form, site index (m), slope (%), aspect (degree), elevation (m), NDVI, and solar radiation (WH/m2) [27, 45].
A digital forest stand map was acquired from the Küre FE [47] and was processed using ArcGIS 10.6 to create the database. The forest stand form, stand structure, and site index layers were created using forest stand maps. To evaluate the damage caused by P. curvidens, a field study was conducted in Küre FE between 2008 and 2017, the most intense period of damage. The beginning of an attack by P. curvidens on a tree can be noticed in the bark by scum emission coming from the reproductive galleries’ entryways. The entrance holes have a 1.5 mm diameter. It usually takes about 2 months for the normal canopy color to change to pale yellow and then reddish. The trees then turn yellow and brown, and eventually die old or weakened trees [48, 49]. Tress that displayed these symptoms were regarded as infected and stands with infested trees were also considered as infested stands. The digital forest stand map was used to create the infested stand map once the P. curvidens-infested forest stands were added to the database.
The digital elevation model (DEM) and Landsat images (Landsat 7 ETM+ and Landsat 8 OLI between 2008 and 2017) were downloaded from the U.S. Geological Survey website [50]. DEM data were used to create the elevation, aspect, slope, and solar radiation maps, and Landsat images were used to compute Normalized Difference Vegetation Index (NDVI). The following formula was used to calculate the NDVI:
NDVI=(NIR−Red)/(NIR+Red)E1
where NIR is the near-infrared and red is the red band.
The resolutions of all layers have to match to provide results that are more reliable. Therefore, using the bilinear resampling method in Spatial Analyst, the NDVI map at 30 m resolution was transformed to 25 m resolution. Table 1 shows the spatial database for the case study area.
Data description
Source
Data type
Resolution
DEM
USGS
Raster
25 m
Landsat 7 ETM+ and Landsat 8 OLI
USGS
Raster
30 m
Forest Stand Map
Küre FE
Vector
25 m
Table 1.
The data and its sources used in the study.
2.3 Analytical hierarchy process (AHP)
Analytical hierarchy process (AHP) is one of the multicriteria decision analyses [51, 52]. AHP analyzes descriptive, qualitative, and quantitative parameters with pairwise comparison and weighting. In AHP, parameters are compared with each other in pairs, a pairwise comparison matrix is created, and then the weights of these parameters are determined [53]. The AHP allows the interpretation of experience and knowledge by modeling not only the basic parameters but also the relationship of the sub-parameters with each other in a certain hierarchical structure. These features make the method more flexible, and its systematic, hierarchical, and adaptive features make the AHP method very popular nowadays, where environmental problems are constantly increasing. For this reason, it is used effectively in various disciplines, such as landslide mapping [54], flood hazard mapping [53], forest fire risk mapping [55, 56], and bark beetle mapping [45, 57]. The knowledge of certain specialists is used to evaluate the comparative magnitudes of variables through pairwise comparisons at the first stage in AHP. Coefficients ranging from 1/9 to 9 are given to compare the relative importance of parameters for each pair in pairwise comparison (Table 2) [54, 58]. Consistency Ratio (CR) is an important validation parameter for the judgment matrix in AHP. The Random Index (RI) should be known before proceeding to the calculation of the CR. RI values were gathered from Saaty [52, 58] in this study. CR was calculated according to Eqs. (2) and (3) to determine the weight of the parameters and to demonstrate their consistency. If the CR is <0.10, it can be said that the generated judgment matrix is consistent. It is necessary to reevaluate the pairwise comparison matrix when the CR is >0.1.
Importance scale
Definitions of importance
Description
1
Equal
The objective is equally benefited by two actions.
3
Moderate
Judgment and experience are slightly preferred to one another.
5
Strong
Judgment and experience are strongly preferred to one another.
7
Very strong
Judgment and experience are very strongly preferred to one another.
9
Extreme
Knowledge and judgment are the strongest conceivable arguments.
2, 4, 6, 8
Intermediate
When a compromise is necessary.
Table 2.
The description of pairwise comparison in AHP.
CI=λmax−n/n−1E2
CR=CI/RIE3
where ƛmax is the biggest value, n is the number of criteria, and CI is the consistency index.
2.4 Maximum entropy modeling (MaxEnt)
Maximum entropy (MaxEnt) is one of the machine learning methods and estimates the probability distribution of species habitats using presence-only data [59]. MaxEnt uses environmental variables as the independent variable and the distribution points of the species as the independent variable. MaxEnt 3.4.4 program was used to determine the potential distribution of fir engraver bark beetle [30, 59, 60]. To create the MaxEnt model, first of all, data sets were created. In this context, stand structure, forest stand form, site index, slope, aspect, elevation, NDVI, and solar radiation were used as variables. These variables have been converted to ASCII format. Stand structure, forest stand form, and site index were considered categorical data, while the others were considered continuous data. The jackknife test was used to determine the contribution rates of the variables in the model to be developed, and the AUC was used to evaluate the model performance. Implementation of the cross-validation method included 10 repetitions, a maximum of 5000 iterations, 1 regularization multiplier, and retention of other parameters at their default values. The logistic output grid format was chosen.
2.5 Fir engraver beetle susceptibility mapping
The risk of a bark beetle infestation is estimated by the fir engraver beetle susceptibility index (FEBSI). Infestation risk is increased by a high FEBSI and vice versa. The significance of each parameter in the AHP model was added up to estimate the FEBSI. All parameter layers were overlaid and the FEBSI values were computed in the AHP model using Raster Calculator tool in ArcGIS 10.6. FEBSI was calculated automatically, according to the selected parameters in MaxEnt model. Based on AHP and MaxEnt models, the susceptibility maps for the fir engraver bark beetle were created. FEBSI were categorized into four groups as low, moderate, high, and extreme based on the natural breaks’ categorization technique in ArcGIS. Two fir engraver beetle susceptibility maps were created according to AHP and MaxEnt models.
2.6 Validation of fir engraver beetle susceptibility mappings
One of the most important stages of predictive models is assessing their performance. Therefore, the accuracy of the AHP and MaxEnt-based beetle susceptibility maps was assessed with the Receiver Operating Characteristic (ROC) method in this study. One of the most popular methods for verifying model-based forecasting is the ROC curve analysis, which ranks the precision of probabilistic and deterministic models, as well as compares them [42]. It is a graphic technique for investigating the trade-off between specificity and sensitivity with the x-axis and the y-axis, respectively. The area under the curve (AUC) is a commonly used indicator of prediction accuracy in ROC curve analysis. The best outcome is to have the highest AUC score, which varies between 0 and 1. An AUC score of 1 indicates a perfect prediction of the model. Five categories are used to categorize the AUC score: poor (0.5–0.6), medium (0.6–0.7), good (0.7–0.8), very good (0.8–0.9), and perfect (0.9–1.0) [33, 42, 45, 61, 62, 63, 64]. For the development of the model, 75% of the infested stands were utilized as training data, while the remaining 25% were used as validation data.
Fir engraver beetle susceptibility maps were produced in a GIS environment using AHP and MaxEnt models. Many different parameters that affect the sensitivity of the beetle have been taken into account to prepare susceptibility maps. The CR was calculated as 0.053 and all pairwise comparison matrices were found to be consistent and satisfactory in the AHP method (Table 3). Stand structure, NDVI, solar radiation, and elevation were found to be more important than other parameters for fir engraver beetle susceptibility with weights of 2.431, 2.354, 1.443, and 1.315, respectively.
Factors
Weights
CR
Classes
Weights
CR
Stand structure
2.431
0.053
Pure
0.667
0.085
Mixed
0.333
Forest stand form
0.555
Even-aged
0.250
0.073
Uneven-aged
0.750
Site index (m)
0.689
>37.6
0.091
0.023
28.3–37.6
0.160
23.3–28.3
0.203
17.0–23.3
0.245
<17.0
0.301
Aspect
0.778
North
0.412
0.098
West
0.281
East
0.191
South
0.116
Slope (%)
0.435
0–20
0.312
0.046
20–40
0.490
>40
0.198
Elevation (m)
1.315
<1000
0.122
0.089
1000–1200
0.333
1200–1400
0.390
>1400
0.155
NDVI
2.354
<0.25
0.089
0.079
0.25–0.35
0.144
0.35–0.45
0.395
>0.45
0.372
Solar radiation (WH/m2)
1.443
<750,000
0.140
0.054
750,000–800,000
0.177
800,000–850,000
0.264
>850,000
0.419
Table 3.
The weights and consistency ratios for all variables in AHP.
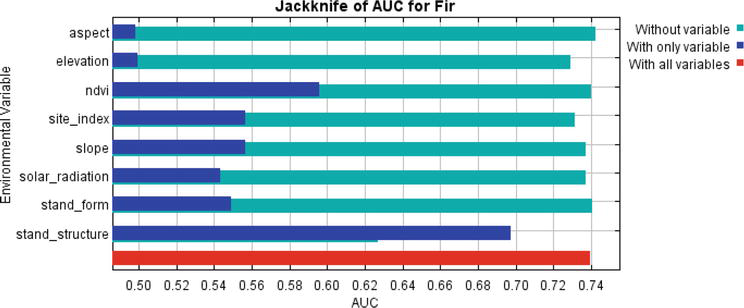
The percent contribution and permutation importance results of the estimation parameters based on the Jackknife test were given in Table 4 and Figure 2. Parameters were ranked based on percentage contribution as stand structure (74.4%), site index (10.9%), elevation (5.4%), and NDVI (2.6%). According to these results, stand structure, NDVI, and elevation were determined as the most important parameters in fir engraver beetle susceptibility in both models. This situation reveals that the stand structure, NDVI, and elevation should be taken into account while preparing the beetle risk map.
Variable
Percent contribution
Permutation importance
Stand structure
74.4
65.7
Site index
10.9
8.2
Elevation
5.4
7.6
NDVI
2.6
4.3
Solar radiation
2.5
6.9
Slope
2.3
4.8
Aspect
1.6
2.3
Forest stand form
0.3
0
Table 4.
Selected parameters and their percentage contribution.
Figure 2.
The Jackknife test of selected parameters for fir engraver beetle.
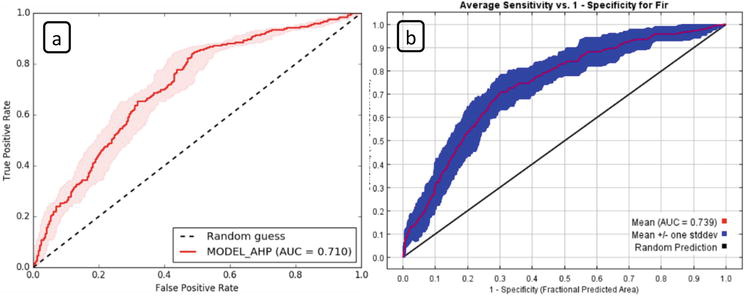
According to the ROC analysis for AHP model, the accuracy of the fir engraver beetle susceptibility mapping was calculated as 0.710, indicating a good prediction of the model. In the MaxEnt model, satisfactory results were also obtained for the fir engraver beetle susceptibility mapping with an AUC value of 0.739, which is higher than 0.5 of a random model (Figure 3). The maps produced according to both models gave reliable results. When both models were compared, it was revealed that the MaxEnt model produced more precise forecasts than AHP method. It can say that both AHP and MaxEnt give consistent results when preparing a beetle risk map.
Figure 3.
The ROC curve of the fir engraver beetle susceptibility maps created by (a) AHP and (b) MaxEnt models.
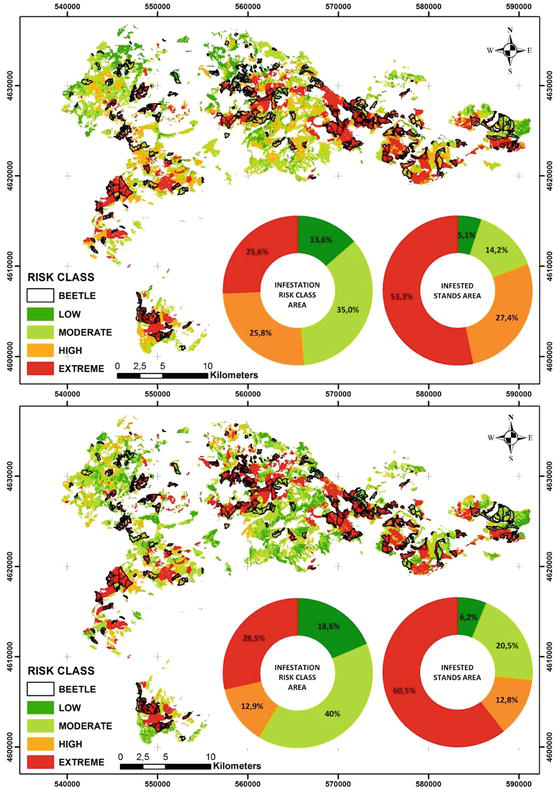
Fir engraver beetle susceptibility maps were created according to AHP and MaxEnt models (Figure 4). According to the AHP method, 5.1%, 14.2%, 27.4%, and 53.3% of the study area are low, moderate, high, and extreme susceptibility classes (Table 5). To verify the results, the maps of infested forest stands and fir engraver beetle susceptibility overlapped. It showed that 80.7% of the infested stands were high and extreme susceptibility classes. According to the MaxEnt model, 40.0% of the study area was in moderate susceptibility classes, while 28.5% is extreme susceptibility classes. 60.5% and 12.8% of infested stands were in extreme and high susceptibility classes, respectively. According to both models, fir forest stands are susceptible to fir engraver beetle.
Figure 4.
Pityokteines curvidens susceptibility maps produced by (a) AHP and (b) MaxEnt models.
ISC
AHP
MaxEnt
SA
IS
SA
IS
Area (ha)
%
Area (ha)
%
Area (ha)
%
Area (ha)
%
Low
4054.5
13.6
315.9
5.1
5576.9
18.6
384.3
6.2
Moderate
10473.5
35.0
882.5
14.2
11952.8
40.0
1272.7
20.5
High
7729.7
25.8
1706.6
27.4
3852.1
12.9
796.6
12.8
Extreme
7653.9
25.6
3312.1
53.3
8529.8
28.5
3763.5
60.5
Total
29911.6
100
6217.1
100
29911.6
100
6217.1
100
Table 5.
The class areas in susceptibility maps based on the AHP and MaxEnt models.
ISC: infestation susceptibility class, SA: study area, and IS: infested stands.
Our results are compatible with other studies. González-Hernández et al., [28] estimated the geographic distribution of Mexican pine bark beetle (Dendroctonus mexicanus) in conifer forests in Mexico using the MaxEnt model. They used bioclimatic (temperature, precipitation, etc.), and topographic variables (altitude, slope, and aspect) for bark beetle mapping. The potential distribution of the bark beetle was reliable, according to the AUC (0.93). The key component in identifying suitable bark beetle habitats was found to be stand structure. Sarıkaya et al. [34] tried to model the current and future habitats of Ips mannsfeldi in Türkiye using MaxEnt. RCP4.5 and RCP8.5 climate change scenarios, as well as bioclimatic factors were used to estimate bark beetle habitats. They stated that elevation was the most important variable and estimated habitat distribution showed high accuracy with an AUC (0.88). Sivrikaya et al., [45] produced I. sexdentatus susceptibility map based on AHP, Frequency Ratio (FR), and Logistic Regression (LR) models. They stated that crown closure was the most important variable for three models. The accuracy of I. sexdentatus susceptibility maps for the LR, FR, and AHP models `was evaluated using the ROC curve, and the FR model provided a more precise estimate than the others.
The invasion of bark beetles is influenced by a variety of biotic and abiotic variables. Early detection of forest stands vulnerable to bark beetle infestations can help reduce the harmful effects of possible infestations. Thus, it is ensured that more accurate decisions are taken for future of forests in the understanding of sustainable management. In this study, susceptibility maps of Pityokteines curvidens were created using analytical hierarchy process (AHP) and maximum entropy (MaxEnt) approaches and their accuracy was compared. According to these results, stand structure, NDVI, and elevation were determined as the most important parameters in fir engraver beetle susceptibility in both models. Comparing the maps created from both models, the MaxEnt model produced more accurate predictions than the AHP. However, it was seen that susceptibility maps based on AHP have also acceptable accuracy. After all, according to both models, fir forest stands are susceptible to the fir engraver beetle and the key component in identifying suitable bark beetle habitats was found to be stand structure.
We would like to say thank you to the Küre Forest Enterprise for providing the forest stand map and related fir engraver beetle information.
References
1.Raffa KF, Grégoire J-C, Lindgren BS. Natural history and ecology of bark beetles. In: Vega FE, Hofstetter RW, editors. Bark Beetles- Biology and Ecology of Native and Invasive Species. Academic Press: Elsevier; 2015. pp. 1-40
2.Negrón JF, Huckaby L. Reconstructing historical outbreaks of mountain pine beetle in lodgepole pine forests in the Colorado Front Range. Forest Ecology and Management. 2020;473:118270. DOI: 10.1016/j.foreco.2020.118270
3.Bentz BJ, Jönsson AM. Modeling bark beetle responses to climate change. In: Vega FE, Hofstetter RW, editors. Bark Beetles- Biology and Ecology of Native and Invasive Species. Academic Press: Elsevier; 2015. pp. 533-553
4.Sibold JS, Veblen TT, Chipko K, Lawson L, Mathis E, Scott J. Influences of secondary disturbances on lodgepole pine stand development in Rocky Mountain National Park. Ecological Applications. 2007;17:1638-1655. DOI: 10.1890/06-0907.1
5.Raffa KF, Aukema BH, Bentz BJ, Carroll AL, Hicke JA, Turner MG, et al. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. Bioscience. 2008;58:501-517. DOI: 10.1641/B580607
6.Boone CK, Aukema BH, Bohlmann J, Carroll AL, Raffa KF. Efficacy of tree defense physiology varies with bark beetle population density: A basis for positive feedback in eruptive species. Canadian Journal of Forest Research. 2011;41:1174-1188. DOI: 10.1139/x11-041
7.Faccoli M, Battisti A, Masutti L. Phenology of Tomicus destruens (Wollaston) in northern Italian pine stands. In: Lieutier F, Ghaioule D, editors. Entomological Research in Mediterranean Forest Ecosystems. Morocco: INRA; 2005. pp. 185-193
8.Wermelinger B. Ecology and management of the spruce bark beetle Ips typographus—A review of recent research. Forest Ecology and Management. 2004;202:67-82. DOI: 10.1016/j.foreco.2004.07.018
9.Kapil R, Marvasti-Zadeh SM, Goodsman D, Ray N, Erbilgin N. Classification of Bark Beetle-Induced Forest Tree Mortality using Deep Learning. 2022. arXiv preprint arXiv:2207.07241
10.Akkemik Ü, Oral D. Türkiye’nin doğal gymnospermleri (Açık Tohumlular). OGM Yayınları. 2011;2011:214
11.GDF. Türkiye Orman Varlığı. [Internet]. 2020 . Available from: https://www.ogm.gov.tr/tr/ormanlarimiz-sitesi/TurkiyeOrmanVarligi/Yayinlar/2020%20T%C3%BCrkiye%20Orman%20Varl%C4%B1%C4%9F%C4%B1.pdf [Accessed: November 26, 2022]
12.Kara F, Lhotka JM. Climate and silvicultural implications in modifying stand composition in mixed fir-pine stands. Journal of Sustainable Forestry. 2020;39:511-525. DOI: 10.1080/10549811.2019.1686030
13.IUCN. International Union for Conservation of Nature. [Internet]. 2022. Available from: https://www.iucnredlist.org/species/31325/9626365. [Accessed: October 15, 2022]
14.Sarikaya O, Avci M. Predators of scolytinae (Coleoptera: Curculionidae) species of the coniferous forests in the Western Mediterranean Region, Turkey. Turkish Journal of Entomology. 2009;33:253-264
15.Wood SL, Bright DE Jr. A catalog of Scolytidae and Platypodidae (Coleoptera), part I: Bibliography. Great Basin Naturalist Memoirs. 1987;1987:11
16.Pennacchio F, Gatti E, Roversi PF. Attacchi di Pityokteines spp. (Coleoptera Scolytidae) su Abies alba Miller in abetine dell’Appennino settentrionale e dell’Aspromonte. In: Proceedings of the Atti XIX Congresso nazionale italiano di Entomologia. Catania; 2002. pp. 921-925
17.Schimitschek E. Forest Insects of Turkey and their Environment. Basics of Turkish Forest Entomology. Prag: Volk and Reich Verlag; 1944. p. 371
18.Tosun İ. Insect pests of the coniferous forests of the Mediterranean region and researches on parasites and predators of their important species. Forestry General Directorate Publications. 1975;612:201
19.Sekendiz OA. Abies nordmanniana Sıev (Spach.ı'ın Doğu Karadeniz bölümü ormanlarındaki zararlı böcekleri ile koruma ve savaş). Trabzon; 1982. p. 201
20.Yüksel B, Öztürk N. First record of the silver fir bark beetle Pityokteines curvidens (Germ.) on blue spruce (Picea pungens Engelm.) in Turkey. Journal of Bartin Faculty of Forestry. 2022;24:25-31. DOI: 10.24011/barofd.1010243
21.Toper KA. The Biology of Pityokteines Curvidens (Germ.) (Coleoptera, Scolytidae) damaged on fir in Bartın and Karabük forests. Zonguldak: Zonguldak Karaelmas University; 1999
22.Serin M, Erdem M, Yüksel B, Akbulut S. Determination of the life cycle of effective bark beetles at Bolu and Aladağ Fir (Abies bornmülleriana Mattf.) stands and their possible preventive measurements. The Western Black Sea Forestry Research Institute. 2005;12:84
23.Chararas C. The biology of P. curvidens and its behaviour with oleoresin extractives. Annales de l'Institut National Agronomique, Alencon. 1958;44:83-131
24.Schwerdtfeger F. Die Waldkrankheiten: Lehrbuch der Forstpathologie und des Forstschutzes. Waldkrankheiten. 4th ed. Berlin: Hamburg; 1981
25.Urban J. Diagnostics of bark beetles of the genus Pityokteines Fuchs important in forestry. Journal of Forest Science. 2002;48:329-341. DOI: 10.17221/11925-JFS
26.Javidan N, Kavian A, Pourghasemi HR, Conoscenti C, Jafarian Z, Rodrigo-Comino J. Evaluation of multi-hazard map produced using MaxEnt machine learning technique. Scientific Reports. 2021;11:1-20. DOI: 10.1038/s41598-021-85862-7
27.Özcan GE, Sivrikaya F, Sakici OE, Enez K. Determination of some factors leading to the infestation of Ips sexdentatus in crimean pine stands. Forest Ecology and Management. 2022;519:120316. DOI: 10.1016/j.foreco.2022.120316
28.González-Hernández A, Morales-Villafaña R, Romero-Sánchez ME, Islas-Trejo B, Pérez-Miranda R. Modelling potential distribution of a pine bark beetle in Mexican temperate forests using forecast data and spatial analysis tools. Journal of Forestry Research. 2020;31:649-659. DOI: 10.1007/s11676-018-0858-4
29.Graham CH, Ferrier S, Huettman F, Moritz C, Peterson T. New developments in museum-based informatics and applications in biodiversity analysis. Trends in Ecology & Evolution. 2004;19:497-503. DOI: 10.1016/j.tree.2004.07.006
30.Phillips SJ, Dudík M. Modeling of species distributions with maxent: New extensions and a comprehensive evaluation. Ecography. 2008;31:161-175. DOI: 10.1111/j.0906-7590.2008.5203.x
31.Cunze S, Tackenberg O. Decomposition of the maximum entropy niche function–a step beyond modelling species distribution. Environmental Modelling & Software. 2015;72:250-260. DOI: 10.1016/j.envsoft.2015.05.004
32.Evangelista PH, Kumar S, Stohlgren TJ, Young NE. Assessing forest vulnerability and the potential distribution of pine beetles under current and future climate scenarios in the Interior West of the US. Forest Ecology and Management. 2011;262:307-316. DOI: 10.1016/j.foreco.2011.03.036
33.Li Y, Johnson AJ, Gao L, Wu C, Hulcr J. Two new invasive Ips bark beetles (Coleoptera: Curculionidae) in mainland China and their potential distribution in Asia. Pest Management Science. 2021;77:4000-4008. DOI: 10.1002/ps.6423
34.Sarikaya O, Karaceylan IB, Sen I. Maximum entropy modeling (maxent) of current and future distributions of Ips mannsfeldi (Wachtl, 1879) (Curculionidae: Scolytinae) in Turkey. Applied Ecology and Environmental Research. ;2018:2527-2535. DOI: 10.15666/aeer/1603_25272535
35.Økland B, Flø D, Schroeder M, Zach P, Cocos D, Martikainen P, et al. Range expansion of the small spruce bark beetle Ips amitinus: A newcomer in northern Europe. Agricultural and Forest Entomology. 2019;21:286-298. DOI: 10.1111/afe.12331
36.Jaime L, Batllori E, Margalef-Marrase J, Navarro MÁP, Lloret F. Scots pine (Pinus sylvestris L.) mortality is explained by the climatic suitability of both host tree and bark beetle populations. Forest Ecology and Management. 2019;448:119-129. DOI: 10.1016/j.foreco.2019.05.070
37.Wong JK, Li H. Application of the analytic hierarchy process (AHP) in multi-criteria analysis of the selection of intelligent building systems. Building and Environment. 2008;43:108-125. DOI: 10.1016/j.buildenv.2006.11.019
38.Nasırı M. Determining the priority of effective factors on forest fire from analytical hierarchy process. Journal of Applied Biological Sciences. 2013;7:52-55
39.Akay AO, Demir M, Akgul M. Assessment of risk factors in forest road design and construction activities with fuzzy analytic hierarchy process approach in Turkey. Environmental Monitoring and Assessment. 2018;190(9):1-12. DOI: 10.1007/s10661-018-6948-0
40.Tiwari A, Shoab M, Dixit A. GIS-based forest fire susceptibility modeling in Pauri Garhwal, India: A comparative assessment of frequency ratio, analytic hierarchy process and fuzzy modeling techniques. Natural Hazards. 2021;105:1189-1230. DOI: 10.1007/s11069-020-04351-8
41.Kumari B. Pandey AC geo-informatics based multi-criteria decision analysis (MCDA) through analytic hierarchy process (AHP) for forest fire risk mapping in Palamau Tiger Reserve, Jharkhand state, India. Journal of Earth System Science. 2020;129:1-16. DOI: 10.1007/s12040-020-01461-6
42.Sivrikaya F, Küçük Ö. Modeling forest fire risk based on GIS-based analytical hierarchy process and statistical analysis in Mediterranean region. Ecological Informatics. 2022;68:101537. DOI: 10.1016/j.ecoinf.2021.101537
43.Vijith H, Dodge-Wan D. Modelling terrain erosion susceptibility of logged and regenerated forested region in northern Borneo through the Analytical Hierarchy Process (AHP) and GIS techniques. Geoenvironmental Disasters. 2019;6:1-18. DOI: 10.1186/s40677-019-0124-x
44.Falťan V, Petrovič F, Gábor M, Šagát V, Hruška M. Mountain landscape dynamics after large wind and bark beetle disasters and subsequent logging—Case studies from the Carpathians. Remote Sensing. 2021;13:3873. DOI: 10.3390/rs13193873
45.Sivrikaya F, Özcan GE, Enez K, Sakici OE. Comparative study of the analytical hierarchy process, frequency ratio, and logistic regression models for predicting the susceptibility to Ips sexdentatus in Crimean pine forests. Ecological Informatics. 2022;71:101811. DOI: 10.1016/j.ecoinf.2022.101811
46.Tahri M, Kašpar J, Madsen AL, Modlinger R, Zabihi K, Marušák R, et al. Comparative study of fuzzy-AHP and BBN for spatially-explicit prediction of bark beetle predisposition. Environmental Modelling. 2022;147:105233. DOI: 10.1016/j.envsoft.2021.105233
47.FMP. Forest Management Plans of Küre Forest Enterprise. Ankara: General Directorate of Forestry; 2011
48.Maksymov JK. Untersuchungen über den krummzähnigen Weißtannenborkenkäfer Ips curvidens Germ. während seiner Massenvermehrung 1947-49 in der Schweiz. Aus dem Entomologischen Institut der Eidg. Technischen Hochschule in Zürich. 1950;1950:499-581
50.USGS. Geological Survey. [Internet]. 2022 . Available from: https://earthexplorer.usgs.gov/ [Accessed: November 20, 2022]
51.Saaty TL. The Analytic Hierarchy Process: Planning, Priority Setting and Resource Allocation. New York: McGraw-Hill; 1980
52.Saaty TL. Theory and Applications of the Analytic Network Process. Decision Making with Benefits, Opportunities, Costs, and Risks. 3rd ed. RWS: Pittsburgh; 2005
53.Kazakis N, Kougias I, Patsialis T. Assessment of flood hazard areas at a regional scale sing an index-based approach and analytical hierarchy process: Application in Rhodope–Evros region, Greece. Science of the Total Environment. 2015;538:555-563. DOI: 10.1016/j.scitotenv.2015.08.055
54.Saha A, Mandal S, Saha S. Geo-spatial approach-based landslide susceptibility mapping using analytical hierarchical process, frequency ratio, logistic regression and their ensemble methods. SN Applied Sciences. 2020;2:1-21. DOI: 10.1007/s42452-020-03441-3
55.Babitha BG, Danumah JH, Pradeep GS, Costache R, Patel N, Prasad MK, et al. A framework employing the AHP and FR methods to assess the landslide susceptibility of the Western Ghats region in Kollam district. Safety in Extreme Environments. 2022;4:171-191. DOI: 10.1007/s42797-022-00061-
56.Bentekhici N, Bellal S, Zegrar A. Contribution of remote sensing and GIS to mapping the fire risk of Mediterranean forest case of the forest massif of Tlemcen (north-West Algeria). Natural Hazards. 2020;104:811-831. DOI: 10.1007/s11069- 020-04191-6
57.Rahaman H, Rehman S, Sajjad H. Indicator-based inherent forest vulnerability using multicriteria decision-making analysis in the Darjeeling District of West Bengal. In: Rani M, Chaudhary BS, Jamal S, Kumar P, editors. Towards Sustainable Natural Resources. Cham: Springer. pp. 51-67. DOI: 10.1007/978-3-031-06443-2_4
58.Saaty TL. Decision Making for Leaders: The Analytic Hierarchy Process for Decisions in a Complex World. 3rd ed. Pittsburgh: RWS Publications; 2012
59.Phillips SJ, Anderson RP, Schapire RE. Maximum entropy modeling of species geographic distributions. Ecological Modelling. 2006;190:231-259. DOI: 10.1016/j.ecolmodel.2005.03.026
60.Phillips SJ, Anderson RP, Dudík M, Schapire RE, Blair ME. Opening the black box: An open-source release of maxent. Ecography. 2017;40:887-893. DOI: 10.1111/ecog.03049
61.de Figueiredo M, Cordella CBY, Jouan-Rimbaud Bouveresse D, Archer X, Bégué JM, Rutledge DN. A variable selection method for multiclass classification problems using two-class ROC analysis. Chemometrics and Intelligent Laboratory Systems. 2018;177:35-46. DOI: 10.1016/j.chemolab.2018.04.005
62.Duračiová R, Muňko M, Ivan B, Koreň M, Resnerová K, Holuša J, et al. A bark beetle infestation predictive model based on satellite data in the frame of decision support system TANABBO. iForest. 2020;13:215-223. DOI: 10.3832/ifor3271-013
63.Gheshlaghi HA, Feizizadeh B, Blaschke B. GIS-based forest fire risk mapping using the analytical network process and fuzzy logic. Journal of Environmental Planning Management. 2020;63:481-499. DOI: 10.1080/09640568.2019.1594726
64.Pourghasemi HR, Beheshtirad M, Pradhan B. A comparative assessment of prediction capabilities of modified analytical hierarchy process (M-AHP) and Mamdani fuzzy logic models using Netcad-GIS for forest fire susceptibility mapping. Geomatics, Natural Hazards Risk. 2016;7:861-885. DOI: 10.1080/19475705.2014.984247
Written By
Fatih Sivrikaya, Gonca Ece Özcan and Korhan Enez
Submitted: 30 November 2022Reviewed: 09 December 2022Published: 10 February 2023
 Open access peer-reviewed chapter
Open access peer-reviewed chapter