 Open access peer-reviewed chapter
Open access peer-reviewed chapter
Abstract
Physical dormancy, also known as hardseededness or seed coat impermeability, is a condition that occurs when a seed’s coat becomes impermeable, preventing the entry of water, gases, and other external factors. This impermeability serves as a protective mechanism, delaying germination until suitable conditions are met. Factors influencing hard seed formation fall into two categories: internal and external. Internal factors pertain to plant-specific traits, such as species and seed morphology. Genetic variations and seed coat characteristics play a role in shaping hard seed formation. External factors, based on environmental conditions, also influence seed development. Soil nutrient availability, water supply, humidity, temperature, and light conditions impact seed coat permeability and germination. Additionally, the timing of seed maturity, drying, and storage conditions can contribute to hard seed formation. The interplay of these factors determines a plant’s tendency to produce hard seeds. Overcoming dormancy caused by seed coat impermeability involves various methods, including physical, chemical, and mechanical approaches. These methods enhance water and gas permeability, facilitating germination. The choice of method depends on seed characteristics and desired outcomes in breaking dormancy. This section emphasizes the impact of hardseededness on seed quality and the application of methods to enhance germination, underscoring its significance in seed science.
Keywords
- hardseededness
- seed coat
- germination
- breaking hardseededness
- factors responsible for physical dormancy
1. Introduction
Seeds, a remarkable wonder of nature, are the cornerstone of plant life. However, they are not just the beginning of a plant’s life; they also encompass many valuable lessons about the resilience of the natural world. This phenomenon, known as “Seed Resilience,” represents the fascinating story of how seeds possess the ability to protect themselves and adapt to environmental challenges.
Seed dormancy is a natural process in which seeds delay their germination, even when the environment is suitable for it. During the dormancy period, seeds typically remain in a dormant state, often protected by mechanisms that prevent early germination. Dormancy permits seeds to germinate when favorable conditions arise, and the diverse levels of dormancy within a population of seeds contribute to a gradual and staggered germination process over an extended period. This chronological variation in germination timing plays a critical role in ensuring the species’ survival, especially in demanding environmental circumstances [1].
While seed dormancy is generally viewed as an unfavorable trait in agriculture, where the primary objective is to promote rapid seed germination and growth, however, there are situations where seed dormancy can offer significant advantages, particularly during the seed development stage. This advantage is particularly noteworthy in the case of cereal crops. Cereals possess a dormancy mechanism that prevents germination while the grains are still attached to the parent plant’s ear. This mechanism acts as a crucial safeguard, especially when there is a period of rainfall during harvest (known as preharvest sprouting), as it prevents premature germination of cereal crops and helps avoid substantial losses in the agricultural sector [2].
Conversely, weed seeds often maintain their inherent dormancy mechanisms as they mature. This allows some weed seeds to persist in the soil for many years, patiently awaiting the right conditions for germination. This poses a threat to crop cultivation, as these seeds can rapidly multiply when favorable conditions finally occur. In summary, seed dormancy is a vital consideration in agriculture and crop cultivation, playing a significant role in the survival strategies of plant species [3].
Seed dormancy is not a single, straightforward trait; rather, it is a complex phenomenon influenced by various factors. Its complexity arises from the intricate interactions between these factors, and it is shaped by multiple elements. Among the numerous factors affecting seed dormancy, hardseededness stands out prominently [4]. Also known as physical dormancy or seed coat impermeability, hardseededness is a fascinating botanical phenomenon that has intrigued researchers and environmental scientists for a long time.
This distinctive feature, present in many plant species, serves as a crucial adaptation strategy. It allows seeds to postpone germination until favorable conditions emerge, bolstering the seed’s ability to withstand challenging environmental conditions [5]. It is a phenomenon that is an integral part of a seed’s life cycle and is a product of the natural selection process. Unlike seeds that readily sprout under suitable conditions, hard seeds boast a robust protective seed coat, serving as a formidable barrier that prevents premature germination, ensuring the seed’s survival until optimal conditions prevail.
In this chapter, we focus on defining seed resilience, understanding why it is important, and exploring the different types of hard-seededness. Additionally, examines the techniques used to overcome physical dormancy and discusses the factors responsible for the formation and maintenance of this dormancy, aiming to explore various aspects of seed resilience.
2. Seeds of resilience
2.1 Definition of physical dormancy
Physical dormancy trait is an inherited characteristic that originates from both the outer and inner structures of the seed during its development [6]. Physical dormancy is recognized in 18 plant families that span a wide range of taxonomic groups and geographical locations worldwide [7, 8, 9, 10]. Studies have suggested that physical dormancy is often inherited as a Mendelian trait, with specific genes controlling the development of impermeable seed coats. The heritability of this trait varies among plant species, indicating a complex genetic basis influenced by both dominant and recessive alleles [11, 12]. When it comes to the genetic basis of hardseededness, two types of alleles come into play: dominant and recessive. Dominant alleles are the ones responsible for the formation of impermeable seed coats. Even a single dominant allele for hardseededness is sufficient to yield seeds adorned with impermeable seed coats. These robust seed coats serve as formidable shields, thwarting the intrusion of external elements [13]. On the other hand, recessive alleles lead to the formation of permeable seed coats. However, for a plant to produce permeable seeds, it must carry two recessive alleles, one from each parent. In other words, if both parent plants contribute a recessive allele for hardseededness, the resulting seeds will have permeable seed coats. This genetic interplay between dominant and recessive alleles determines whether a seed’s coat will be impermeable or permeable, ultimately influencing its ability to withstand external environmental conditions [14].
The degree of physical dormancy is influenced not only by genetics but also by several environmental factors. The primary factor influencing hardseededness in seeds is their moisture content. The level of moisture within seeds plays a critical role in determining their ability to germinate. Seeds with low moisture content frequently display hardseededness because the limited availability of moisture impedes the essential biochemical processes required for germination. In simpler terms, when seeds lack sufficient moisture, they tend to become hard and resistant to germination. It is widely accepted that hardseededness is typically a permanent trait. Once a seed coat becomes impermeable, it is considered unlikely to return to a permeable state unless there is damage to the seed coat or a specific structure within it, known as the “water gap” opens to enable water to reach the internal structures and rehydrate them [5, 15]. Many studies on seed development have demonstrated that the transition from a permeable to an impermeable seed coat coincides with the decrease in moisture content during the maturation drying phase of seed development [16, 17, 18].
Temperature is also undeniably the primary environmental factor influencing physical dormancy in seeds as a key element in synchronizing plant growth with changes in climate conditions. Many studies have investigated the relationship between temperature during seed maturation and the permeability of the resulting seeds [19, 20, 21]. These studies have consistently shown that seeds that mature at higher temperatures tend to produce a higher proportion of impermeable seeds compared to those that mature at lower temperatures. It is important to note that measuring temperature throughout the seed maturation period can be challenging, and seasonal or location-specific temperature variations around the parent plant may also play a role in determining the permeability of seeds. For example, seeds maturing at higher temperatures are likely to experience more water loss, leading to a higher proportion of seeds with physical dormancy.
2.2 Physical dormancy and germination
Determining when genetic and physiological differences emerge is a challenging endeavor, primarily because seed dormancy is intricately regulated at different stages of development, often influenced by environmental factors [22]. A dormant seed is essentially incapable of initiating germination, even when presented with favorable environmental conditions and a suitable habitat. This incapacity can be attributed to various factors, such as non-viability, the absence of an embryo, or the presence of dormancy. The impermeability of seed coats in physically dormant seeds is the result of complex structural and chemical adaptations (Figure 1).
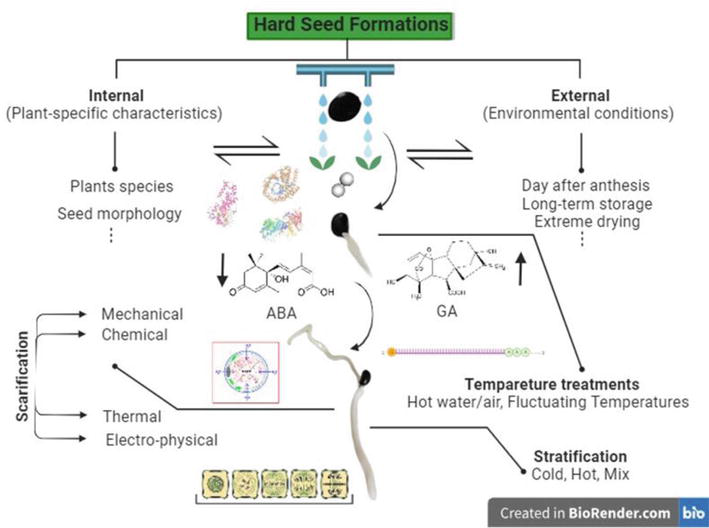
Figure 1.
Physical dormancy in hardseed formulations.
Germination encompasses a series of events that start with the absorption of water by a resting, desiccated seed and concludes with the elongation of the embryonic axis, ultimately resulting in the emergence of the radicle [23]. Water uptake by a seed occurs in three distinct phases. The first phase, known as Phase I, is characterized by a rapid and initial uptake of water. This phase typically occurs within 1–8 h. Water uptake occurs in all seeds, whether dormant or non-dormant, and even in non-viable seeds. In fact, non-viable seeds tend to absorb more water compared to viable seeds because the turgor pressure in the cells of viable seeds restricts excessive water uptake. Phase II, also referred to as the lag phase, marks a stage where water uptake becomes limited. However, during this phase, essential enzymes like amylase, endoxylanase, and phytase are produced. These enzymes are responsible for synthesizing new proteins required for germination. Additionally, the conversion of stored materials for germination, mRNA synthesis, and the initiation of energy production through sugar metabolism degradation also begin during this phase. Consequently, this phase facilitates the transfer of nutrients from storage areas in the endosperm to the growth points. Phase II can span from several hours to a few days and typically concludes when the radicle emerges from the seed coat. The third phase, Phase III, signifies the first visible sign of germination, with the emergence of the radicle. This emergence is primarily a result of cell enlargement rather than cell division. Shortly thereafter, cell division occurs at the tip of the radicle, initiating its elongation [24, 25, 26, 27, 28].
Physical germination is an event that occurs through a two-stage process involving the rupturing of the seed coat. The first stage involves the rupture of the seed’s outer covering, known as the testa. This protective layer shields the embryo within the seed, and its removal is necessary to initiate germination. The second stage involves the rupture of an inner layer called the endosperm. This inner layer serves as a storage site for nutrients within the seed and provides the essential nutrients for the initial growth of the embryo [22, 29].
Germination is completed, specifically the emergence of the radicle (the embryonic root) becomes visible. This marks a clear indicator of seed vitality and the beginning of germination. However, in seeds with physical dormancy, the seed coat is tough, which can limit water uptake. As a result, germination may either not occur at all or be restricted when water uptake is insufficient. Physical dormancy is an evolutionary strategy that prevents seeds from germinating until environmental conditions are favorable, and water uptake is a crucial component of this process. Most of the time, the seed coat limits germination by either preventing water and/or oxygen from passing through or by resisting the emergence of the radicle with its mechanical properties [30]. In this process, the seed coat serves as a vital organ involved in nourishing the embryo and subsequently safeguarding it against harmful environmental factors [31, 32].
Dormancy caused by seed coat impermeability can be overcome through various methods, including physical, chemical, and mechanical approaches. These methods aim to enhance water and gas permeability in hard seeds, facilitating moisture absorption and initiating germination. The choice of method depends on specific seed characteristics and desired outcomes in breaking dormancy.
3. Methods of breaking physical dormancy
Breaking physical dormancy involves various methods to weaken or remove this barrier, allowing water and oxygen to penetrate the seed and initiate germination. Physical dormancy may require different scarification methods involving either physical or chemical agents to overcome seed dormancy.
3.1 Effect of seed scarification
Seed scarification is a crucial technique used in agriculture and horticulture to promote germination by weakening the seed coat. This process involves various methods to facilitate water absorption and oxygen penetration into the seed, ultimately kickstarting the germination process. Various methods can be employed for seed scarification, including chemical, thermal, electro-physical, and mechanical approaches [33].
The predominant form of scarification is mechanical scarification, wherein the testa, or seed coat, undergoes physical manipulation to enable the ingress of moisture and air. Various approaches are utilized in this process, encompassing the abrasion of seed coats with tools such as metal files, sandpaper, knives for nicking, gentle hammering for cracking, or any method that weakens or opens the seed coat [34, 35, 36, 37, 38, 39]. This technique finds extensive application within the realms of horticulture and agriculture, serving as a fundamental method to facilitate the germination of seeds encased in rigid or impermeable seed coats. Its primary function lies in enabling seeds to surmount physical dormancy, thereby instigating the pivotal processes of water absorption and germination. Mechanical scarification stands as a pragmatic and efficacious method, widely adopted to unlock the growth potential of numerous plant species reliant on this mechanism for breaking their dormancy and flourishing within diverse ecological contexts.
Chemical scarification is a seed treatment method that employs various chemical substances, including potent acids such as H2SO4 (sulfuric acid), HCl (hydrochloric acid), NaHClO3 (sodium hypochlorite), and hydrogen peroxide (H2O2). Also, organic solvents such as alcohol and acetone are used to eliminate seed dormancy. These chemicals have been the subject of extensive research, revealing their capability to effectively disrupt and overcome the dormancy mechanisms present in specific types of seeds. Through the application of these chemicals, the hard or impermeable seed coats of certain plant species can be altered or weakened, allowing for enhanced water penetration and subsequent germination. In recent studies, chemical scarification with H2SO4 has been particularly effective in enhancing seed germination and seedling growth in various plant species. For instance, in okra [40, 41], strelitzia [42], muscari [43], and raspberry seeds [44], H2SO4 treatment has been found to break physical dormancy barriers and promote successful germination. Breaking physical dormancy can also take place as seeds pass through an animal’s digestive system. Unlike conventional approaches to dormancy relief, exposure to hydrochloric acid (HCl) within the digestive tract gradually erodes the impermeable seed layers [45, 46]. As well as the scarification treatment involving HCl acid is recognized for its high effectiveness in promoting germination for species with hard seed coats. This method involves the use of HCl to weaken or modify the tough outer seed coat, allowing water to penetrate and initiate germination more easily. Various methods have been employed to overcome physical dormancy, and they have proven effective in breaking seed dormancy and enhancing germination success in a range of plants, including guava [47], sunflower seeds [48], Indian siris [49], as well as certain forest tree seeds [50]. Sodium hypochlorite scarification is one such method that has proven highly efficient in breaking physical seed dormancy. Recent studies [51, 52] have reported significant improvements in seed germination, notably in
3.2 Effect of temperature treatment
The impacts of wet heat, dry heat, and the alternating use of wet heat and ice water on the alleviation of seed dormancy play crucial roles in the process of seed germination and propagation.
3.2.1 Hot water/air treatment
The process of hot water or air treatment entails immersing seeds in hot water or exposing them to high-temperature air. This procedure serves to soften the seed coat, eliminate any wax or grease present, and facilitate the penetration of water into the seed under natural conditions. These treatments hold the potential to physically disrupt or soften the macrosclerid layer, with a specific focus on the strophiolar plug. This may lead to the creation of cracks, a softer plug, or alterations that enhance water absorption and other germination-related processes [59, 60].
The presence of cracks or fissures in the seed coat plays a critical role in initiating the germination process. These openings act as conduits through which water can enter the seed. Once water infiltrates the seed, it sets off a series of vital biochemical processes necessary for germination. This mechanism has been extensively documented in the research conducted by Baskin and Baskin in [61]. Furthermore, the use of hot air or water as a treatment for seed germination can have additional consequences beyond facilitating water entry. It may expose the embryo to thermal shock, characterized by a sudden temperature change that can affect its metabolic processes. The impact of this thermal shock can vary among different species, potentially either promoting or inhibiting germination, contingent on a species’ adaptability to temperature fluctuations [62]. The efficacy of dry heat treatment, on the other hand, is contingent on the extent and duration of exposure. Therefore, determining the optimal combination of temperature and duration that yields a high germination rate necessitates empirical research tailored to each specific species [63].
This method is also proficiently employed to break seed dormancy in rice [64], okra [65, 66], black mimosa (
Another method employed to overcome hard seed dormancy is a combination of temperature and moisture, often referred to as after-ripening. It is utilized to break hard seed dormancy, particularly in seeds with physical or physiological dormancy. It involves subjecting seeds to specific temperature and moisture conditions, which trigger a series of biochemical and physiological changes that ultimately promote germination [70]. After-ripening takes place in a broad spectrum of warm and dry environmental settings, and the particular combinations of temperature and relative humidity (RH) significantly impact the after-ripening rate of different species. Numerous investigations into after-ripening have been conducted, during which the moisture content (MC) of seeds was assessed, and the seeds were subsequently stored at defined temperatures while being observed until their dormancy was disrupted. Heat and moisture combination treatments were discovered to be effective in overcoming seed dormancy in Malvaceae species, as reported by Demir [66] and Ellis et al. [71]. Furthermore, physical dormancy in seeds of
The well-documented role of fire in aiding the germination of species naturally found in fire-prone regions is significant. The process of vegetation combustion, as seen in forest fires, releases a variety of factors that actively promote seed germination and facilitate the breaking of seed coats, allowing water to penetrate for germination. Due to the heat shock from the fire, the specialized, heat-sensitive tissues within the seed coat are either broken or displaced, leading to the creation of ‘water gaps’ [17, 75, 76]. This allows the seed to become capable of absorbing water and oxygen, which in turn aids in the ongoing process of germination. Several studies have presented compelling evidence that smoke and its components can initiate seed germination and promote growth after germination [77, 78, 79, 80]. The nature and extent of the temperature required to break seed dormancy and the response to temperature can vary depending on several factors. These factors encompass the characteristics of the seed coat, such as the thickness and structure of the cuticle, epidermis, palisade, parenchyma, and the type of water gap [81]. Seed size is also a significant consideration, as smaller seeds tend to exhibit higher temperature tolerance and lower temperature thresholds [82]. Additionally, it has been observed that the age of the seed can influence its response to temperature [83]. Finally, whether the temperature conditions are moist or dry can also shape the seed’s response [84].
3.2.2 Exposing seeds to fluctuating temperatures and lights
Physiological dormancy can be alleviated not only through the process of after-ripening in dry storage conditions but also by subjecting seeds to variable temperature conditions. The specific temperature exposure required for this process can vary depending on the needs and germination characteristics of different plant species. The choice of temperature ranges and the duration of exposure may vary based on the species of seeds and their sources. In 1916 [85], Harrington’s research demonstrated that seeds of Trifolium and Melilotus underwent a softening process when exposed to temperature fluctuations, alternating between 10°C or lower and 20°C or higher. This softening effect was significantly enhanced when the seeds had previously experienced cooler temperatures below 10°C. In another study, exposure to specific temperature conditions, such as warm stratification (WS) and cold stratification (CS), increased the germination rate to over 80% [86]. However, it has been shown that seed germination behavior is often associated with factors such as habitat, seed mass, and life cycle type. For instance, many plants in moist habitats like wetlands tend to have their seed germination promoted by temperature fluctuations, while forest plants typically do not respond positively to temperature fluctuations [87]. Fenner and Thompson [88] suggested that small-seeded plant species may be more stimulated by temperature fluctuations compared to larger-seeded species.
While all seeds require water, oxygen, and appropriate temperature conditions for germination, some species also need light to germinate. It has been observed that light filters through the seed coat, and specific wavelengths of light can penetrate the embryo [89]. Generally, larger-seeded species do not require light during germination because they have enough nutrients to grow in the dark. On the other hand, smaller-seeded species need light to germinate. In fact, these seeds remain dormant even if they have absorbed water when planted at a depth where they cannot access light. Light requirement is a dormancy factor and may not prevent germination in vegetable species except under extreme conditions. However, wild species in nature and tree seeds in forests generally have a greater requirement for light. The effect of light on seeds varies depending on genotype and environmental conditions during seed maturation, dormancy breaking, and germination [23]. Light and gibberellins (GA) are also two key factors that can help overcome this type of dormancy. It has been suggested that as a photoreceptor, phytohormone can contribute to the regulation of light in initiating or terminating the processes of seed dormancy [90]. Some seeds have light-sensitive pigments in their seed coats, and exposure to specific wavelengths of light (usually red or far-red light) can trigger biochemical changes in the seed coat. This can lead to the degradation of inhibitory compounds and the softening of the seed coat, allowing water and oxygen to penetrate the seed, which is essential for germination [91, 92, 93]. However, the mechanism by which light promotes or inhibits seed germination has not yet been fully understood [94].
Gibberellins, a class of plant hormones, play a crucial role in regulating various growth and developmental processes within plants, including the germination of seeds. One of the primary functions of gibberellins in seed germination is their ability to stimulate the production of specific enzymes, such as α-amylase [95]. These enzymes are responsible for breaking down stored starches present in the endosperm of the seed, which provides the energy for the growth of roots and shoots [96]. During the early stages of seed germination, the embryo relies on these stored starches as a source of energy and essential nutrients. The breakdown of starches by alpha-amylase leads to the formation of soluble sugars, which serve as a vital energy source for the developing embryo [97]. This energy is necessary to support the growth and emergence of the embryonic shoot and root from within the seed. In addition to their role in providing energy, gibberellins also contribute to alleviating dormancy in seeds. Dormancy is a state of inhibited growth and development that some seeds enter to ensure their survival until favorable conditions for germination are met. Gibberellins promote dormancy release, effectively signaling to the seed that it is time to initiate the germination process. Moreover, gibberellins can influence the physical properties of the seed coat. They have the capacity to soften the seed coat, making it more permeable to water and gases. The function of gibberellins in dormancy alleviation is combined with the necessity for light [98]. This softening of the seed coat is particularly important for the emerging radicle, which is the embryonic root of the plant. The softening of the seed coat facilitates the penetration of the radicle through the seed coat, allowing it to emerge more easily and initiate the germination process.
The increase in GA levels occurs with the release of seed dormancy, for example, during imbibition and stratification. In some dormant seeds, treatment with exogenous GA can take advantage of this opportunity to break dormancy, thereby facilitating faster seedling production [99]. Many previous studies have utilized GA to facilitate the breakage of dormancy in various plant species, including
4. Bibliometric analysis
To gain a comprehensive insight into the research landscape concerning physical dormancy and hardseededness within the agricultural domain, a thorough bibliometric analysis was undertaken (Figure 2).

Figure 2.
Visual representation of keywords based on co-word (co-occurrence) using RStudio.
In the bibliometric analysis conducted based on the content indexed in Web of Science, all agricultural fields were selected with the keywords “Physical Dormancy” and “Hardseededness” 675 results were obtained in English documents. According to years, the oldest being 1990 and the newest being 2023, accessed 629 articles, 25 proceeding papers, 21 review articles, 7 early access, 4 editorial materials, 3 book chapters, 2 meeting abstracts and 1 research note from 31 different disciplines. As seen in Figure 2, it is divided into 6 different clusters according to the author’s keywords, and in the red cluster, the term physical dormancy is included in 167 works, germination is 136, seed dormancy is 113, dormancy is 103, seed dormancy is 60 works and hardseededness is 49 works. In physical dormancy studies, 1836 different authors were found, 24.74% of which were multinational studies. Considering the topicality of the research topic, it was observed that the document average age was 10.1 years, and the average number of citations per document was 17.07.
5. Conclusions
In summary, seeds represent a remarkable natural wonder, serving as the bedrock of plant life and imparting valuable lessons about the resilience of the natural world, a concept known as “Seed Resilience.” Seed dormancy, a natural process that delays germination even in optimal conditions, plays a vital role in ensuring the survival of plant species by enabling staggered germination to adapt to challenging environments. Physical dormancy, or hardseededness, is a specific type of dormancy characterized by an impermeable seed coat, acting as a protective shield to prevent premature germination and safeguard the seed until favorable germination conditions prevail. The factors influencing the formation of hard seeds can be categorized into internal (plant-specific) and external (environmental) factors, encompassing genetic differences among plant species, morphological seed features, and environmental conditions like soil quality, temperature, humidity, and light exposure.
The breaking of dormancy in hard seeds can be accomplished through various methods, including mechanical abrasion, chemical treatments, temperature fluctuations, and cycles of drying and rehydration, all aimed at improving water and gas permeability for successful germination. Understanding physical dormancy is of utmost importance for researchers, ecologists, and horticulturists, as it significantly impacts seed germination timing and success, with broad implications for plant populations, crop production, and ecosystem restoration efforts.
Furthermore, the role of light in regulating dormancy and germination remains a topic of ongoing debate. Light has been recognized both as a stimulant for germination and as a dormancy terminator, contingent upon the perspective taken. It is considered the final step in the dormancy-breaking process, allowing seeds to germinate even in darkness. Particularly, red light, mediated through phytochrome, can reverse dormancy and promote germination. In seeds with coat dormancy, both light and gibberellins (GA) are believed to release dormancy and facilitate germination.
In the field of bibliometrics, an extensive body of research in this area is evident, with numerous authors contributing to the understanding of physical dormancy, including a significant portion of multinational studies. The research is current and well-cited, underscoring the significance of this topic. In conclusion, the intricate interplay of factors that influence seed dormancy and germination is a complex subject under ongoing research and debate. A comprehensive understanding of these mechanisms is vital for effective seed management and ecosystem restoration practices.
References
- 1.
Bewley JK, Nonagaki H. Seed maturation and germination. In: Roitberg BD, editor. Book of Reference Module in Life Sciences. Matsudo, Japan: Elseiver; 2017. DOI: 10.1016/B978-0-12-809633-8.05092-5 - 2.
Bewley JD, Bradford KJ, Hilhorst HWM, Nonogaki H. Seeds: Physiology of Development, Germination and Dormancy. New York, NY, USA: Springer; 2013. p. 392. DOI: 10.1007/978-1-4614-4693-4 - 3.
Qasem JR. Weed seed dormancy: The ecophysiology and survival strategies. In: Carlos J-LC, editor. Book of Seed Dormancy and Germination. London, UK: IntechOpen; 2019. pp. 1-36. DOI: 10.5772/intechopen.88015 - 4.
Graeber KAI, Nakabayashi K, Miatton E, Leubner-Metzger G, Soppe WJ. Molecular mechanisms of seed dormancy. Plant, Cell and Environment. 2012; 35 (10):1769-1786. DOI: 10.1111/j.1365-3040.2012.02542.x - 5.
Baskin CC, Baskin JM, Li X. Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Species Biology. 2000; 155 :139-152. DOI: 10.1046/j.1442-1984.2000.00034.x - 6.
Hudson AR, Ayre DJ, Ooi MK. Physical dormancy in a changing climate. Seed Science Research. 2015; 25 (2):66-81. DOI: 10.1017/S0960258514000403 - 7.
Baskin CC. Breaking physical dormancy in seeds: Focusing on the lens. New Phytologist. 2003; 158 :229-232. DOI: 10.1046/j.1469-8137.2003.00751.x - 8.
Baskin JM, Baskin C, Dixon KW. Physical dormancy in the endemic Australian genus Stylobasium, a first report for the family Surianaceae (Fabales). Seed Science Research. 2006; 16 :229-232. DOI: 10.1079/SSR2006248 - 9.
Jayasuriya K, Baskin JM, Baskin CC. Sensitivity cycling and its ecological role in seeds with physical dormancy. Seed Science Research. 2009; 19 :3-13. DOI: 10.1017/S096025850818730X - 10.
Gama-Arachchige NS, Baskin JM, Geneve RL, Baskin CC. Identification and characterization of the water gap in physically dormant seeds of Geraniaceae, with special reference to Geranium carolinianum . Annals of Botany. 2010;105 :977-990. DOI: 10.1093/aob/mcq078 - 11.
Todd-Bockarie AH, Duryea ML. Seedpretreatment methods to improve germination of the multipurpose West African forest species Dialium guineense . Forest Ecological Management. 1993;57 :257-273. DOI: 10.1016/0378-1127(93)90176-N - 12.
Singh I, Gill MS, Bains TS, Singh P. Genetic analysis of Hardseededness in Mungbean ( Vigna radiata L. Wilzeck). Crop Improvement-India. 2005;32 (2):170-172 - 13.
Marjushkin VF, Sichkar VI, Michailov VG, Polivoda LV. Research notes: Inheritance of hardseededness in soybean. Soybean Genetics Newsletter. 1987; 14 :Article 74, 294-Article307 - 14.
Smýkal P, Vernoud V, Blair MW, Soukup A, Thompson RD. The role of the testa during development and in establishment of dormancy of the legume seed. Frontiers in Plant Science. 2014; 5 (351):1-19. DOI: 10.3389/fpls.2014.00351 - 15.
Paulsen TR, Colville L, Kranner I, Daws MI, Högstedt G, Vandvik V, et al. Physical dormancy in seeds: A game of hide and seek? The New Phytologist. 2013; 198 :496-503. DOI: 10.1111/nph.12191 - 16.
Baskin JM, Baskin CC, Li X. Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Species Biology. 2008; 15 :139-152. DOI: 10.1046/j.1442-1984.2000.00034.x - 17.
Gama-Arachchige NS, Baskin JM, Geneve RL, Baskin CC. Quantitative analysis of the thermal requirements for stepwise physical dormancy-break in seeds of the winter annual Geranium carolinianum (Geraniaceae). Annals of Botany. 2013;111 :849-858. DOI: 10.1093/aob/mct046 - 18.
Jaganathan GK. Influence of maternal environment in developing different levels of physical dormancy and its ecological significance. Plant Ecology. 2016; 217 :71-79. DOI: 10.1007/s11258-015-0560-y - 19.
MacGregor DR, Kendall SL, Florance H, Fedi F, Moore K, Paszkiewicz K, et al. Seed production temperature regulation of primary dormancy occurs through control of seed coat phenylpropanoid metabolism. New Phytologist. 2015; 205 (2):642-652. DOI: 10.1111/nph.13090 - 20.
El-Keblawy A, Shabana HA, Navarro T, Soliman S. Effect of maturation time on dormancy and germination of Citrullus colocynthis (Cucurbitaceae) seeds from the Arabian hyper-arid deserts. BMC Plant Biology. 2017;17 :1-10. DOI: 10.1186/s12870-017-1209-x - 21.
Lachabrouilli AS, Rigal K, Corbineau F, Bailly C. Effects of agroclimatic conditions on sunflower seed dormancy at harvest. European Journal of Agronomy. 2021; 124 :126209. DOI: 10.1016/j.eja.2020.126209 - 22.
Bentsink L, Koornneef M. Seed dormancy and germination. In: John Herlihy J, Ludwig RN, Ackerveken G, JM MD, editors. The Arabidopsis Book. Vol. 6. Washington US: American Society of Plant Biologists; 2008. pp. 1-18. DOI: 10.1199/tab.0119 - 23.
Mavi K, Ermiş S, Kenanoğlu BB, Demir İ. Fide yetiştiriciliğinde tohum kalitesi ve tohum uygulamalarının önemi. In: Elliatıoğlu S, Yetişir H, editors. Sebzelerde Fide Yetiştiriciliği. 1st ed. Ankara, Türkiye: Gece Publishing; 2022. pp. 69-134 (in Turkish) - 24.
Schopfer P, Plachy C. Control of seed germination by abscisic acid. II. Effect on embryo water uptake in Brassica napus L. Plant Physiology. 1984;766 (1):155-160. DOI: 10.1104/pp.77.3.676 - 25.
Bewley JD. Seed germination and dormancy. The Plant Cell. 1997; 9 (7):1055. DOI: 10.1105/tpc.9.7.1055 - 26.
Manz B, Muller K, Kucera B, Volke F, Leubner-Metzger G. Water uptake and distribution in germinating tobacco seeds investigated in vivo by nuclear magnetic resonance imaging. Plant Physiology. 2005; 1386 (1):1538-1551. DOI: 10.1104/pp.105.061663 - 27.
Nonogaki H, Bassel GW, Bewley JD. Germination-still a mystery. Plant Science. 2010; 179 :574-581. DOI: 10.1016/j.plantsci.2010.02.010 - 28.
Pereira do Espirito Santo A, Caixeta Oliveira H, Fernandes Fraceto L, Santaella C. Nanotechnology potential in seed priming for sustainable agriculture. Nanomaterials. 2021; 11 :267. DOI: 10.3390/nano11020267 - 29.
Foschi ML, Juan M, Pascual B, Pascual-Seva N. Influence of seed-covering layers on caper seed germination. Plants. 2023; 12 (3):439. DOI: 10.3390/plants12030439 - 30.
Debeaujon I, Leon-Kloosterziel KM, Koornneef M. Influence of the testa on seed dormancy, germination, and longevity in Arabidopsis. Plant Physiology. 2000; 122 (2):403-414 10.1104/pp.122.2.403 - 31.
Weber H, Borisjuk L, Wobus U. Controlling seed development and seed size in Vicia faba : A role for seed coat-associated invertases and carbohydrate state. The Plant Journal. 1996;10 :823-834 - 32.
Raviv B, Aghajanyan L, Granot G, Makover V, Frenkel O, Gutterman Y, et al. The dead seed coat functions as a long-term storage for active hydrolytic enzymes. PLoS One. 2017; 12 (7):e0181102. DOI: 10.1371/journal.pone.0181102 - 33.
Oidov A, Sukhee B, Agvaantseren T. Some results of the experıment on scarification of seeds of forage legume (the sample of alfalfa seeds). World Science. 2023; 2 (80):1-8. DOI: 10.31435/rsglobal_ws/30062023/7981 - 34.
Ermis S, Demir I. Determining the Effect of some Treatments in Relation to Elemination of Hardseededness and Its Maintenance during Storage in Okra, 5. Çanakkale, Turkey: Sebze Tarımı Sempozyumu; 2004. pp. 101-105 (in Turkish) - 35.
Avci S, Kaya MD. Seed and germination characteristics of wild Onobrychis taxa in Turkey. Turkish Journal of Agriculture and Forestry. 2013; 37 (5):555-560. DOI: 10.3906/tar-1211-29 - 36.
Cuadra CDL, Vidal AK, Lagomarsino F, Peñaloza P, Mansur LM, Huenchuleo C. Effect of temperature and scarification on seed germination of Conanthera spp. (Tecophilaeaceae). Chilean Journal of Agricultural Research. 2019;79 (2):323-329. DOI: 10.4067/S0718-58392019000200323 - 37.
Liyanage GS, Offord CA, Sommerville KD. Techniques for breaking seed dormancy of rainforest species from genus Acronychia. Seed Science and Technology. 2020; 48 (2):159-165. DOI: 10.15258/sst.2020.48.2.03 - 38.
Górnik K, Sas-Paszt L, Seliga Ł, Pluta S, Derkowska E, Głuszek S, et al. The effect of different stratification and scarification treatments on breaking the dormancy of Saskatoon berry seeds. Agronomy. 2023; 13 (2):520. DOI: 10.3390/agronomy13020520 - 39.
Keskin B, Temel S, Gurel G, Ozden E. Effects of some temperature and dormancybreaking applications on germination rates of camelthorn ( Alhagi pseudalhagi (Bieb.) Desv.) seeds. Research in Agricultural Sciences. 2022;54 (1):22-30. DOI: 10.5152/AUAF.2023.220307 - 40.
Demir I, Ermis S. Effect of harvest maturity and drying method on okra seed quality. Seed Technology. 2005; 27 :81-88. Available from:https://www.jstor.org/stable/23433218 - 41.
Okereke CN, Ikegbunam CN, Nwaogaranya UP, Ogbu AC, Francis OA. Iroka CF. Comparative evaluation of the effects of pre-sowing treatments on the germination and growth parameters of Abelmoschus esculenta Linn. Asian Journal of Research Crop Science, 2023;8 (3):108-118. DOI: 10.9734/ajrcs/2023/v8i3172 - 42.
Paiva PDDO, Silva DPCD, Silva BRD, Sousa IPD, Paiva R, Reis MVD. How scarification, GA3 and graphene oxide influence the in vitro establishment and development of Strelitzia. Plants. 2023; 12 (11):2142. DOI: 10.3390/plants12112142 - 43.
Labbaf N, Rohollahi I, Naji AM. Muscari seed germination enhancement by using sulfuric acid, and stratification priming. Ornamental Horticulture. 2023; 29 :171-180. DOI: 10.1590/2447-536x.v29i2.2548 - 44.
Pergolotti V, Marcellini M, Contreras E, Mezzetti B, Gambardella M, Capocasa F, et al. Standardization of an in vitro seed germination protocol compared to acid scarification and cold stratification methods for different raspberry genotypes. Horticulturae. 2023; 9 (2):153. DOI: 10.3390/horticulturae9020153 - 45.
Razanamandranto S, Tigabu M, Neya S, Odén PC. Effects of gut treatment on recovery and germinability of bovine and ovine ingested seeds of four woody species from the Sudanian savanna in West Africa. Flora-Morphology, Distribution, Functional Ecology of Plants. 2004; 199 :389-397. DOI: 10.1078/0367-2530-00167 - 46.
Jaganathan GK, Yule K, Liu B. On the evolutionary and ecological value of breaking physical dormancy by endozoochory. Perspectives in Plant Ecology, Evolution and Systematics. 2016; 22 :11-22. DOI: 10.1016/j.ppees.2016.07.001 - 47.
Abbasi M, Heidari M, Rahimi M. Improving germination of guava ( Psidium guajava ) seeds by acid scarification. Journal of Horticultural Science. 2014;27 (4):394-399. DOI: 10.22067/JHORTS4.V0I0.30581 - 48.
Gandy YP, Persans MW, Summy KR. An acid-bath technique to break seed dormancy in common sunflower, Helianthus L.annuus (Asteraceae). Subtropical Agriculture and Environments. 2015;66 :23-26 - 49.
Ibrahim HS, Aref Hawramee OK. Impact of acid scarification and cold mist stratification on enhancing seed germination and seedling early growth of Albizia lebbeck (L.) Benth. Mesopotamia Journal of Agriculture. 2019;47 (2):1-13. DOI: 10.33899/magrj.2019.163175 - 50.
Deltalab B, Moghadam NN, Raad MK, Kaviani B. The effect of cold and acid scarification on seed germination of three green space tree species. Journal of Ornamental Plants. 2023; 13 (2):85-97. Available from:https://dorl.net/dor/28210093.2023.13.2.2.5 - 51.
Promwee A, Islam SS, Khomphet T. Effect of dissolved oxygen and chemical scarification on Andrographis paniculata seed germination in macrobubble conditions. International Journal of Agronomy. 2023;2023 :1-13. DOI: 10.1155/2023/3459377 - 52.
Yeh CH, Chen KY, Lee YI. Asymbiotic germination of Vanilla planifolia in relation to the timing of seed collection and seed pretreatments. Botanical Studies. 2021;62 (6):1-12. DOI: DOI.10.1186/s40529-021-00311-y - 53.
Ponert J, Figura T, Vosolsobě S, Lipavská H, Vohník M, Jersáková J. Asymbiotic germination of mature seeds and protocorm development of Pseudorchis albida (Orchidaceae) are inhibited by nitrates even at extremely low concentrations. Botany. 2013;91 :662-670. DOI: 10.1139/cjb-2013-0082 - 54.
Ponert J, Šoch J, Vosolsobě S, Čiháková K, Lipavská H. Integrative study supports the role of trehalose in carbon transfer from fungi to mycotrophic orchid. Front Plant Science. 2021; 12 :793876. DOI: 10.3389/fpls.2021.793876 - 55.
Kindiger B. A method to enhance germination of eastern gamagrass. Maydica. 1994; 39 :53-53 - 56.
Hassan SM, Ghareib HR. Bioactivity of Ulva lactuca L. acetone extract on germination and growth of lettuce and tomato plants. African Journal of Biotechnology. 2009;8 (16):3832-3838 - 57.
Nasreen S, Khan MA, Zia M, Ishaque M, Uddin S, Arshad M, et al. Response of sunflower to various pre-germination techniques for breaking seed dormancy. Pakistan Journal of Botany. 2015; 47 (2):413-416 - 58.
Agostini RTD, Abrantes FL, Machado-Neto NB, Custódio CC. Ethanol and hormones in physiological conditioning on germination and seed dormancy of Urochloa humidicola cv. Llanero. Journal of Seed Science. 2022; 44 :e202244022. DOI: 10.1590/2317-1545v44261411 - 59.
Aliloo AA, Darabinejad S. Evaluation of different techniques for breaking seed dormancy of Heliotropium europaeum L. (Boraginaceae). Journal of Biological and Environmental Science. 2013;7 (20):87-91 - 60.
Rodrigues-Junior AG, Mello ACM, Baskin CC, Baskin JM, Oliveira DM, Garcia QS. A function for the pleurogram in physically dormant seeds. Annals of Botany. 2019; 23 (5):867-876. DOI: 10.1093/aob/mcy222 - 61.
Baskin CC, Baskin JM. Seeds: Ecology, Biogeography, and, Evolution of Dormancy and Germination. Lexington, Kentucky, US: Academic Press, Elsevier; 1998. 666 p. DOI: 10.1016/B978-0-12-080260-9.X5000-3 - 62.
Mohammadi G, Khah EM, Honarmand SJ, Shirkhani A, Shabani G. Effects of seed hardness breaking techniques on okra ( Abelmoschus esculentus L.) germination. International Journal of Agriculture and crop sciences. 2012;4 (6):264-273 - 63.
Schelin M, Tigabu M, Eriksson I, Sawadogo L, Oden PC. Effects of scarification, gibberellic acid and dry heat treatments on the germination of Balanites aegyptiaca seeds from the Sudanian savanna in Burkina Faso. Seed Science and Technology. 2003;31 (3):605-617. DOI: 10.15258/sst.2003.31.3.10 - 64.
Tung LD, Serrano EP. Effects of warm water in breaking dormancy of rice seed. Omonrice. 2011; 18 (1):129-136 - 65.
Demir I. Development of seed quality during seed development in okra. In: Proceedings of the International Symposium on Agrotechnics and Storage of Vegetable and Ornamental Seeds. Bari, Italy; Vol. 362. Jun 1994. pp. 125-132 - 66.
Demir I. The effects of heat treatment on hardseededness of serially harvested okra seed lots at optimum and low temperatures. Scientia Horticulturae. 2001; 89 (1):1-7. DOI: 10.1016/S0304-4238(00)00216-8 - 67.
Giasson C, Baretta C, Sobral LS, Baldissera R. Dormancy breaking, germination, and production of Mimosa bimucronata (DC.) Kuntze seedlings. Cerne. 2019;25 :68-75. DOI: 10.1590/01047760201925012612 - 68.
Chaodumrikul S, Kaewsorn P, Chulaka P, Chanprasert W. Breaking seed dormancy in smooth loofah ( Luffa cylindrica (L.) M. Roem.) using scarification and dry heat treatment. Agriculture and Natural Resources. 2016;50 (2):85-88. DOI: 10.1016/j.anres.2015.09.003 - 69.
Taghizadeh M, Sajadi FS. Effects of dormancy breaking methods on germination of Cercis siliquastrum andSpartium junceum and seedling growth. Ornamental Horticulture. 2023;29 :28-36. DOI: 10.1590/2447-536X.v29i1.2528 - 70.
Baskin JM, Baskin CC. A classification system for seed dormancy. Seed Science Research. 2004; 14 (1):1-16. DOI: /10.1079/ssr2003150 - 71.
Ellis RH, Hong TD, Roberts EH. Handbook of Seed Technology for Genebanks. Vol. 1. Principles and Methodologies. Rome: IBPGR; 1985. 210 p - 72.
Jayasuriya K, Baskin JM, Baskin C. Cycling of sensitivity to physical dormancy-break in seeds of Ipomoea lacunosa (Convolvulaceae) and ecological significance. Annals of Botany. 2008;101 (3):341-352. DOI: 10.1093/aob/mcm285 - 73.
Li X, Baskin JM, Baskin CC. Anatomy of two mechanisms of breaking physical dormancy by experimental treatments in seeds of two north American Rhus species (Anacardiaceae). American Journal of Botany. 1999; 86 :1505-1511. DOI: 10.2307/2656788 - 74.
Rodriguez Araujo ME, Pérez D, Aronson J, Cross A. Filling gaps on seed germination and species selection in drylands of Argentina: Work in progress and reflections on intelligent tinkering. Multequina. 2021; 30 :165-180 - 75.
Nandi OI. Ovule and seed anatomy of Cistaceae and related Malvanae. Plant Systematics and Evolution. 1998; 209 :239-264. Available from:https://www.jstor.org/stable/23643179 - 76.
Brits GJ, Manning JC. Seed structure and physiology in relation to recruitment ecology in Leucospermum (Proteaceae) in fynbos. Australian Journal of Botany. 2019; 67 (4):290-308. DOI: 10.1071/BT18199 - 77.
Van Staden J, Jäger AK, Light ME, Burger BV, Brown NAC, Thomas TH. Isolation of the major germination cue from plant-derived smoke. South African Journal of Botany. 2004; 70 (4):654-659. DOI: 10.1016/S0254-6299(15)30206-4 - 78.
Kulkarni MG, Light ME, Van Staden J. Plant-derived smoke: Old technology with possibilities for economic applications in agriculture and horticulture. South African Journal of Botany. 2011; 77 (4):972-979. DOI: 10.1016/j.sajb.2011.08.006 - 79.
Khatoon A, Rehman SU, Aslam MM, Jamil M, Komatsu S. Plant-derived smoke affects biochemical mechanism on plant growth and seed germination. International Journal of Molecular Sciences. 2020; 21 (20):7760. DOI: 10.3390/ijms21207760 - 80.
Gokdas Z, Yildirim E, Gupta S, Demir I. Karrikinolide stimulated seed germination of artificially aged marrow, cabbage and pepper seeds through repair of cell structure and enzyme activity. South African Journal of Botany. 2022; 151 :208-213. DOI: 10.1016/j.sajb.2022.09.049 - 81.
Hradilová I, Duchoslav M, Brus J, Pechanec V, Hýbl M, Kopecký P, et al. Variation in wild pea ( Pisum sativum subsp.elatius ) seed dormancy and its relationship to the environment and seed coat traits. PeerJ. 2019;7 (e6263):2-32. DOI: 10.7717/peerj.6263 - 82.
Tavşanoğlu Ç, Ergan G, Çatav ŞS, Zare G, Küçükakyüz K, Özüdoğru B. Multiple fire-related cues stimulate germination in Chaenorhinum rubrifolium (Plantaginaceae), a rare annual in the Mediterranean Basin. Seed Science Research. 2017;27 (1):26-38. DOI: 10.1017/S0960258516000283 - 83.
Downes KS, Light ME, Pošta M, Van Staden J. Fire-related cues and the germination of eight Conostylis (Haemodoraceae) taxa, when freshly collected, after burial and after laboratory storage. Seed Science Research. 2015; 25 (3):286-298. DOI: 10.1017/S0960258515000227 - 84.
Liyanage GS, Ooi MK. Do dormancy-breaking temperature thresholds change as seeds age in the soil seed bank? Seed Science Research. 2017; 27 (1):1-11. DOI: 10.1017/S0960258516000271 - 85.
Harrington GT. Agricultural value of impermeable seeds. Journal of Agricultural Research. 1916; 6 :761-796 - 86.
Roh MS, Bentz JA, Wang P, Li E, Koshioka M. Maturity and temperature stratification affect the germination of Styrax japonicus seeds. The Journal of Horticultural Science and Biotechnology. 2004; 79 :645-651. DOI: 10.1080/14620316.2004.11511820 - 87.
Ozden E, Light ME, Demir I. Alternating temperatures increase germination and emergence in relation to endogenous hormones and enzyme activities in aubergine seeds. South African Journal of Botany. 2021; 139 :130-139. DOI: 10.1016/j.sajb.2021.02.015 - 88.
Fenner M, Thompson K. The Ecology of Seeds. Cambridge, UK: Cambridge University Press; 2005. 250 p. DOI: 10.1017/CBO9780511614101 - 89.
Widell KO, Vogelmann TC. Fiber optic studies of light gradients and spectral regime within Lactuca sativa achenes. Physiologia Plantarum. 1988;72 (4):706-712. DOI: 10.1111/j.1399-3054.1988.tb06369.x - 90.
Brady SM, McCourt P. Hormone cross-talk in seed dormancy. Journal of Plant Growth Regulation. 2003; 22 :25-31. DOI: 10.1007/s00344-003-0018-7 - 91.
Casal JJ, Sánchez RA. Phytochromes and seed germination. Seed Science Research. 1998; 8 (3):317-329. DOI: 10.1017/S0960258500004256 - 92.
Sanchez RA, Mella RA. The Exit from Dormancy and the Induction of Germination: Physiological and Molecular Aspects. Handbook of Seed Physiology Application to Agriculture. 1st ed. New York: Food Product Press and The Haworth Press, Inc; 2004. 479 p - 93.
Lamont BB, Pausas JG. Seed dormancy revisited: Dormancy-release pathways and environmental interactions. Functional Ecology. 2023; 37 (4):1106-1125. DOI: 10.1111/1365-2435.14269 - 94.
Motsa MM, Slabbert MM, Van Averbeke W, Morey L. Effect of light and temperature on seed germination of selected African leafy vegetables. South African Journal of Botany. 2015; 99 :29-35. DOI: 10.1016/j.sajb.2015.03.185 - 95.
Gao S, Chu C. Gibberellin metabolism and signaling: Targets for improving agronomic performance of crops. Plant and Cell Physiology. 2020; 61 (11):1902-1911. DOI: 10.1093/pcp/pcaa104 - 96.
Kaneko M, Itoh H, Ueguchi-Tanaka M, Ashikari M, Matsuoka M. The α-amylase induction in endosperm during rice seed germination is caused by gibberellin synthesized in epithelium. Plant Physiology. 2002; 128 (4):1264-1270. DOI: 10.1104/pp.010785 - 97.
Shaik SS, Carciofi M, Martens HJ, Hebelstrup KH, Blennow A. Starch bioengineering affects cereal grain germination and seedling establishment. Journal of Experimental Botany. 2014; 65 (9):2257-2270. DOI: 10.1093/jxb/eru107 - 98.
Thomas TH. Some reflections on the relationship between endogenous hormones and light-mediated seed dormancy. Plant Growth Regulation. 1992; 11 :239-248 - 99.
Lee SY, Park K, Jang BK, Ji B, Lee H, Baskin CC, et al. Exogenous gibberellin can effectively and rapidly break intermediate physiological dormancy of Amsonia elliptica seeds. Frontiers in Plant Science. 2022;13 :1043897. DOI: 10.3389/fpls.2022.1043897 - 100.
Ozden E, Demir I. GA3 enhanced seed germination of Solanum torvum . Radovi Poljoprivrednog Fakulteta Univerziteta u Sarajevu (Works of the Faculty of Agriculture University of Sarajevo. 2016;61:316-320 - 101.
He XQ , Wang YR, Hu XW, Baskin CC, Baskin JM, Lv YY. Seed dormancy and dormancy-breaking methods in Leymus chinensis (Trin.) Tzvel (Poaceae) Grass and Forage. Science. 2016;71 (4):641-648. DOI: 10.1111/gfs.12220 - 102.
Chen JZ, Huang XL, Xiao XF, Liu JM, Liao XF, Sun QW, et al. Seed dormancy release and germination requirements of Cinnamomum migao , an endangered and rare woody plant in Southwest China. Frontiers in Plant Science. 2022;13 :770940. DOI: 10.3389/fpls.2022.770940 - 103.
Silva ECD, Villa F, Silva DFD, Possenti JC, Silva LSD, Ritter G. Araticum accessions: Effect of gibberellic acid concentrations and soaking times on seed dormancy overcoming. Revista Caatinga. 2021; 34 :614-620. DOI: 10.1590/1983-21252021v34n313rc