Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
Plant parasitic nematodes are major pests of the agricultural industry in developing countries. This group is associated with different parts viz., flower, leaves, buds, roots, trunk etc., of approximately all crucial crops of agronomy due to their omnipresent nature. They are categorized as endo, ecto and semi-endoparasite based on the feeding habits. PPNs possess remarkable characteristics especially, parasitic adaptations which help in combating adverse conditions. Furthermore, they interact with other microorganisms (pathogens) forming complex diseases in crops. For effective management of the PPNs through biological control, it is essential to understand their parasitic mechanism, adaptation of J2 stages, feeding mechanism, host-nematode relationship and identification of associated microbiota. This review focuses on the basic biology of PPN, feeding habits, parasitic potential, molecular insights for understanding host-nematode relationship and their management by exploiting the inherent abilities of biocontrol agents.
Nematology Laboratory, Department of Zoology, Chaudhary Charan Singh University, Meerut, India
Ashok Kumar Chaubey
Nematology Laboratory, Department of Zoology, Chaudhary Charan Singh University, Meerut, India
*Address all correspondence to: himanisadhnasharma@gmail.com
1. Introduction
Plant parasitic nematodes found in association with plants pose a major threat to the agriculture sector and are responsible for approximately $80–$118 billion dollars loss globally [1]. They are obligate parasites of plants which derive their nutrition from them by using their specialized structure viz., stylet. The stylet of PPNs required for feeding and also helps in the establishment of the nematode infection in the host as they used to pierce the plant cell wall for invasion. Furthermore, the stylet also showed connections to the glands present in the pharynx which helps in the production of the molecule’s requisite during the infection, invasion and other fundamental processes [2, 3]. These molecules released from the PPN affect the host immune system which ultimately enhances the parasitic association. These nematodes possess the potential to damage any part of the plant, however; roots found to be most susceptible for the PPN colonization especially endoparasites [4, 5]. The parasitic adaptations, feeding behavior and various lifestyle modes of PPN help in the survival in adverse conditions.
1.1 Morphology
Nematodes are the fascinating animals on earth after insects, with many free living and parasitic forms. It’s a noteworthy characteristic in case of nematodes that all the conceivable habitats viz., marine, soil and aquatic ecosystems are occupied by this amazing group. Diversity can also be seen in their shape and size as they occur in variable shapes such as melon shape (Meloidogyne), reniform (Rotylenchulusreniformis), or swollen shape (Tylenchulussemipenetrans) etc. and their size ranges from microscopic (Paratylenchus 0.18–0.6 mm) to several meters long (Paralongidorus maximus ∼11.0 mm). The general body structure of nematodes is elongated, cylindrical and tapered at both ends as suggested by their name ‘nema’ which means thread like [6, 7]. The basic body design consists of an inner alimentary canal which is surrounded by an outer tube of body wall which is commonly referred as ‘tube within a tube’ structure. The alimentary canal basically comprises gonads and digestive tract. Body wall consists of a variety of muscles viz., ventral and dorsal longitudinal muscles which regulate nematode locomotion as well as these muscles are activated by the dorsal and ventral nerves. Unlined, pressurized, fluid filled cavity which is present between the tubes is known as ‘pseudocoelom’ formed from the blastula surrounding the gut cavity. Its primary function is to provide the turgor pressure for the entire body and subsequently aids the transfer of oxygen, nutrients and metabolic products. Exoskeleton of nematodes, also known as cuticle, is made up of collagens, glycoproteins, insoluble proteins and lipids.
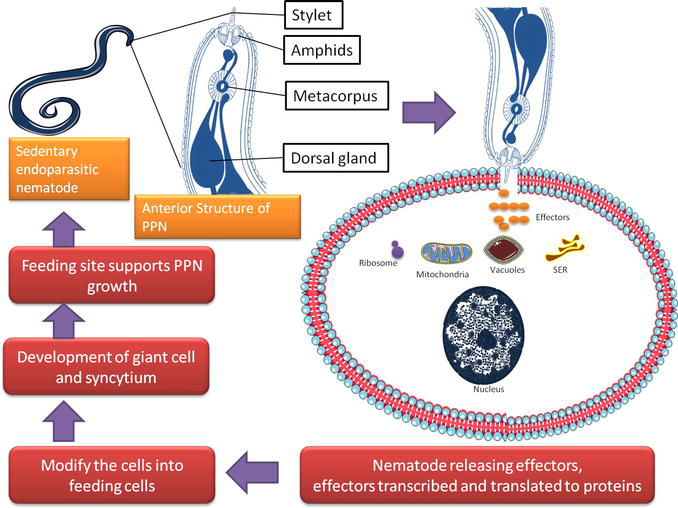
Plant parasitic nematodes possess an accessory structure known as stylet which is utilized for association with the plants and deriving the nutrients. The shape, size and position of this stylet are of taxonomic importance for the identification of this nemic fauna. The stylet of PPN showed connections with the intestine because of attachment to the pharynx anteriorly (Figure 1). Order Tylenchida and Dorylaimida are found to be the pathogen of plants, invertebrates and fungi [8] and as parasites of agricultural crops and forest trees they have great economic importance. All possible habitats or ecological niches have been occupied by Tylenchids. The infection of PPN can be found in all the possible habitats; however, diversity occurred in the root parasites [5]. Over 4000 species of plant parasitic nematodes have been identified but it is very interesting to note that only few genera possess the potential of economic loss viz., Meloidogyne, Helicotylenchus, Hoplolaimus, Heterodera, Globodera, Criconemoides, Hemicycliophora etc. [1]. The genus Meloidogyne comprises 105 species, out of which 10 species found to cause potent damage to the agricultural sector [9].
Figure 1.
The anterior end of plant parasitic nematode. The protrusible stylet helps in piercing the plant cell wall. The dorsal gland and other organs produced effectors which modify the surrounding cells for the development of feeding site.
1.2 Life cycle of plant parasitic nematodes
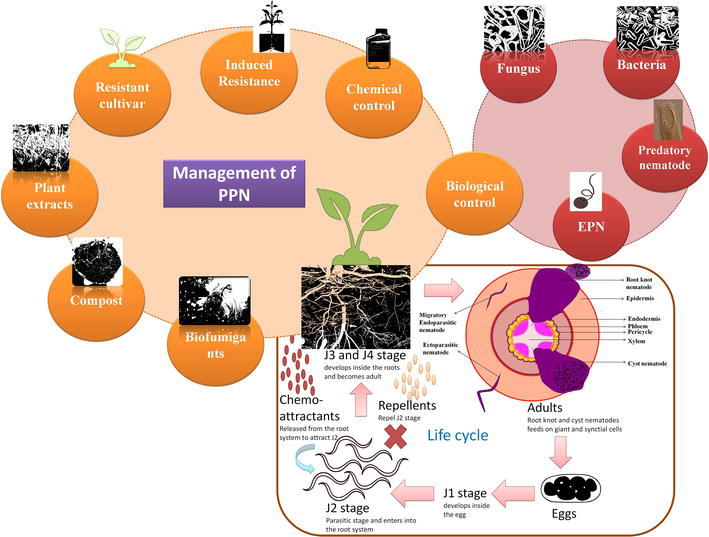
The life cycle of both root knot and cyst nematodes consists of 25–30 days on average. The second juvenile stage (J2) of the PPN finds the host roots for feeding. This stage showed attraction towards the roots and their penetration inside the host cells for satiating the nutrition needs developed by the synctial cells. The synctial comprises the host cells approximately 200 which lost their cell wall and contributed their protoplasts to the large feeding site [10]. After entering into the host cells, the J2 stage molts to J3, J3 molts to J4 and J4 molts to adults. The J2 stage once started feeding becomes larger in size and finally transforms to the males. Among all the larval stages, only the J2 stage showed mobility and others were immobile. Males possess a vermiform body and find their way out from the plants. The exterior environment consists of various chemical cues/pheromones released from the female’s body. These cues help males in locating the females for fertilization. Female body comprises eggs produced during fertilization. These eggs were protected in a cyst and all the cysts have the potential to release J2 stage. In case of cyst nematodes, J2 reached to the vascular cells and feeding site developed consists of syncytium cell, however, in root knot nematodes, J2 stage migrated to cortical cells where feeding site developed from the continuous mitosis without following cytokinesis [11]. The process leads to the development of a giant cell which becomes the feeding site for the J2 [12]. The J2 molts thrice and finally becomes an adult. Fertilization is not observed in the root knot, however, males formed during this entire process. Parthenogenesis observed in the root knot nematodes. Eggs are not present inside the female body, instead found outside the body in a protected matrix released from the female body itself. Feeding site either synctial cell or giant cells required repeat stimulus from J2 stage and both nematodes development and life cycle found dependent on these sites (Figure 2). This repeated thrust and maintenance of the feeding sites affects the roots which in turn affected the nutrient and water supply to the various parts of the plants, ultimately, affecting the yields.
Figure 2.
Different feeding strategies and life cycle of plant parasitic nematode. The role of chemoattrants and repellents on the parasitic J2 stage during the host localization. Management of PPN using different strategies and biological control.
1.3 Feeding behavior strategies of PPN
PPNs are dependent on the plants for their survival and they are found in close proximity to the plant parts in order to complete their life cycle [6]. They are broadly categorized into above ground feeders and below ground feeders based on their feeding strategies adopted. Further, below ground feeders are classified into ectoparasites, semi-endoparasites and endoparasites.
1.3.1 Above ground feeders
1.3.1.1 Stem and bulb nematodes
Ditylenchus dipsac found responsible for causing disease in the plants as it largely affects the stem and bulb of the host hence referred to as stem and bulb nematodes. Hyacinth’s brown ring disease caused by this nematode in the plants where wilting, collapsing and yellowish color in the leaves was observed. In the garlic plants, the weight of the bulb decreases and they turn brownish and shriveled. These nematodes affect the storage of these plant bulbs ultimately decreasing the yield [7].
1.3.1.2 Seed gall nematodes
This nematode targets the seeds of the host plant affecting the yield. The disease which affects the seeds of wheat is caused by the Anguinatritici called ear-cockleseed or seed gall, hence commonly referred as seed gall nematode. They are ectoparasites but affect other areas of the plants as soon as it invades the seeds and inflorescence. The diseased plant showed symptoms such as wrinkles, swollen and bulged leaves, whitening, yellowish color of the stems with bending etc. [7].
1.3.1.3 Foliar nematodes
The nematodes belonging to genus Aphelenchoides spp. specifically target the leaves of the plants. The symptoms include the yellowish leaves followed by drying and wrinkling, the younger leaves can be spotted with whitish or yellowish patches. Further, it affects the kernels of the plants which become smaller and distortion in their shape is also observed during infection. The grains of the infected plants showed discoloration and became weak due to which cracks were observed [7].
1.3.2 Below ground feeders
1.3.2.1 Ectoparasites
These nematodes found on the surface of the host and derive their nutrition from the root’s epidermis. They are further classified into sedentary ectoparasites and migratory ectoparasites. The sedentary nematodes showed specificity in the host and maintain a permanent attachment with the host, for example, Belonolaimus sp., Xiphenema sp., Trichodorus sp. etc. The J2 stage found to be the infective stage in these and only free-living stage. The migratory ectoparasites derive their nutrition from the roots itself but they do not maintain a permanent attachment with the host. Instead, if the roots get disturbed, they will show detachment from the roots and be found in the soil as free nematodes, for example, Criconemella sp., Paratylenchus sp., Hemicycliophora sp. etc. [7].
1.3.2.2 Semi endoparasites
These nematodes were attached with the host anteriorly, however, their posterior body was found free in the soil. They are further classified into sedentary and migratory semi-endoparasites. Insedentary semi-endoparasites, the anterior body showed permanent attachment with the host for example, Tylenchorchynchs, Hoplolaimus etc. TheJ3/J4–adult are the infective stages of sedentary nematodes. The migratory semi-endoparasites does not form any permanent attachment with the host andJ2/J4 are the infective stages, for example, Rotylenchulus sp., Tylenchulus sp. etc. [7].
1.3.2.3 Endoparasites
The endoparasites further categorized into migratory and sedentary. The migratory endoparasites enter into the host through roots and migrate to other areas as they feed on the cells, for exmple, Pratylenchus sp, Radopholus sp. etc. On the contrary, the sedentary endoparsites also entered through root systems but lost their mobility as they feed on special cells viz., giant cell or syncytial cells, for example, Meloidogyne sp., Heterodera sp, Naccobus sp. The infective stage of sedentary endoparsites is the J2 stage for spreading the infection [7].
2.1 Parasitic genes and effectors molecule of plant parasitic nematodes
2.1.1 Root-knot nematodes
PPN release effectors molecules to weaken the host immune system. One of the important and harmful root-knot nematodes viz., Meloidogyne releases a number of effectors which helps in their survival by combating the defense system. MiPFN3 gene in Meloidogyne incognita found to be coding for profilin (involved in disassembly of actin) which has the potential to bind the motor actin filament. The effectors molecule binds with the actin filament of the host cell disrupting its normal functioning facilitating the PPN survival [13]. The other effectors are secreted, encoded by gene MilSE5, which disrupts and interrupts important pathways of metabolism [14]. One of the gene viz., Misp12 secreted effectors which directly interfere with the defenses system of the host [15]. The Inflorescence Deficient in Abscission (IDA) like peptide is also encoded by the M. incognita geneMilDL1 [16]. The immune system of the host was compromised when effectors, encoded by gene Mh265, released in M. hapla [17]. In M. javanica, one of the gene MjTTL5 responsible for the production of transthyretin-like protein which helps in suppressing the defense system of the host cell [18]. In M. graminicola, effectors and the irrespective genes were identified helping in colonization and combating the defense of the host. The host system was suppressed by the effectors encoded by the genes MgGPP and Mg16820 [19, 20]. The innate immunity is also compromised by the effectors encoded by gene Mg01965 [21]. The apoptosis process in the plant cells were suppressed by the effectors (gene MeTCTP) released by the M. enterolobii [22].
2.1.2 Cyst nematodes
Cyst nematodes also released these effectors molecules which enhancing their survival inside the plant’s cells and tissues. The gene Hs30D08 in Heterodera schachtii codes for the effectors interfering with spliceosome machinery which leads to the changes in those genes responsible for the development of feeding site [23]. Other gene mainly, HsPDI coding for the enzyme disulfide isomerase found to alter the redox potential of plant cell/tissue [24]. Another gene reported from the H. schachtii viz., HsTyr encoded tyrosinase enzyme which plays important role in maintenance of homeostasis [25]. The gene HsCLEB found coding for the B-type CLE peptide which plays key role in cell signaling in plants. These peptides had been regulating the vascular cells playing important role in the development of feeding area for the PPN [26]. There are some DNA binding proteins coded by the gene HsGLAND4 which inhibit the transcription of lipid transfer protein as they bind to the promoters observed in Arabidopsis [27]. The other gene viz., Hs32E03 coding those molecules which are responsible for the chromatin modifications in order to induce alteration in the ribosomal RNAs of host [28]. There are some effectors which induce alteration in the plant cell wall encoded by the gene HaEXPB2 in H. avenae [29]. The immune system of the host found to be compromised especially hypersensitive responses in the presence of effectors coded by HgGLAND18 in H. glycines [30]. In Globodera pallida, two genes viz., GpSPRY-414-2 and GpGS have been identified for releasing SPRYSEC effector and glutathione synthetase enzyme, respectively [31, 32].
2.1.3 Lesion nematodes
Lesion nematodes, one of migratory parasitic nematodes, releases effector molecules helping in either metabolism or suppression of the host defense system. The gene Ppen12895_c0_seq1 (FAR) encodes for the molecules that play a role in the metabolism of fatty acids in Pratylenchus penetrans [33]. Another gene reported from the P. penetrans also facilitates survival inside the host [33]. In P. zeae, molecules encoded by the gene Vap-1 are found responsible for suppressing the immune system of the host [34]. On the contrary, another gene viz., Sec-2 from the same species played a significant role in combating the defense system of the host [34].
2.1.4 Burrowing nematodes
The burrowing nematode especially, Radopholus similis had known to harbor some genes which have facilitated the entry, survival of the nematode and infection to the host. The gene Rs-scp-1 coding the effectors helping the nematode development, facilitating entry into the host and also contribute to the pathogenesis [35]. Another gene viz., Rs-csp responsible for facilitating the juvenile stages entry and also enhances their pathogenicity [36]. Furthermore, the Rs-cb-1 gene is known to code for the molecule playing an essential role during the reproduction and ease invasion of the nematode [37]. Likewise, molecules encoded by the Rs-crt gene facilitate the reproduction and increase pathogenic potential of the nematode [38]. Rs-far-1 gene is found to be a coding effector which contributes during the entry, developmental stages, disrupting the defense system of the host [39].
2.1.5 Potato rot nematodes
In Ditylenchus destructor, two genes DD03093 (VAP-1) and DDC03397 (VAP-2) had been reported for coding the molecules which play an essential role in suppressing the host defense system [40]. The other gene viz., DD03835 (Sec-2) products facilitates the nematodes in overcoming the defense system of the host [40].
2.1.6 Pine wood/wilt nematodes
From the genome of Bursaphelenchus xylophilus, six genes have been reported playing a crucial role in parasitism. The first gene, BxSapB1, provides those molecules which enhance the pathogenicity of the nematode and also contribute to the apoptosis of the host cell [41]. One of the genes encodes the 1–3-endoglucanaseenzyme and other encodes expansin-like protein found responsible for degrading the plant cell wall at the time of invasion [42]. To overcome the defense barriers of the host, detoxifying enzymes also released viz., peroxiredoxin, glutathione-S-transferase and cytochrome P450 [43].
2.2 Adaptation of J2 stage during host invasion and colonization
2.2.1 Migratory endoparasites
The juvenile stages of PPNs such as Ditylenchus, Anguina and Pratylenchus require hosts for their survival purposes as they feed on them. Feeding is important for molting which ensures their survival. During extremities of temperature, pH, rain etc. the J2 stage enters into a dormant stage and possesses the ability of surviving without host for a certain period. The quiescent stage in Radopholus similis found to be 6 months [44]. The juveniles of Anguina sp withstand the temperature extremities until it finds the host [45]. Some of the PPN juveniles adopt the anhydrobiosis mode which is commonly observed in the entomopathogenic nematodes. The anhydrobiosis process is a gradual release of the moisture from the nematode body to a level where a dried powder form is observed. This process enhances the survival chances of the juveniles for a longer duration without a host. This condition is observed in Ditylenchus sp. The other way of enhancing survival is the arrest of the juvenile stage. This was observed in Ditylenchus dipsaci in which the last juvenile stage arrested and showed variations from the normal juveniles. The arrested stage was found to be larger in size with a higher amount of lipid reserves which somehow showed similarity with the Caenorhabditis elegans larvae [46]. To understand the similarity between the Ditylenchus and Caenorhabditis, researchers focused on the molecular studies to resolve this mystery. There are some overlapping regions found in this duo which raises the possibility of convergent evolution [47].
2.2.2 Sedentary endoparasites
The migratory stage in these PPNs is the second juvenile stage (J2) which is responsible for spreading the infection in the fields. The hatching of J2 stage from the egg found regulated from various chemical cues released from the specific host. The pre-J2 stage of sedentary nematodes can be quiescent in the environment for a longer period of time until it hatches out from the egg [48]. The signals from the host help in the hatching of the J2 from the egg and they showed similarity with the dauer larvae of free-living nematodes [49]. The J2 stage of cyst nematodes exhibit variations in the cuticle on the basis of lipophilicity as they perceive cues from the host [50]. These J2 stages are either pre- parasitic or parasitic, found to derive their energy from the deposited fat [51, 52]. The larvae of Heterodera glycines showed resistance to various harmful toxic substances ensuring its survival [53]. The pre-J2 stage can be dormant for a longer duration; however, the parasitic stage needs to find the suitable host for its survival, development within a short period of time. The parasitic J2 stage hatching should have occurred in the presence of the host. The cues released from the host helps the J2 stages in finding the host. The J2 stages also showed responses towards the chemical signals released from the host parts such as roots and these signals travel larger distances. The other signals possess a shorter range and these signals sensed by the J2 nearer to the host [54]. The J2 stage of Meloidogyne hatched out in response to extreme moisture or heat and does not require signals from the plants. This could be the reason behind their broad host range helping them in maintaining their population during unsuitable conditions. Though cyst nematodes J2 stages hatched out in response to environmental conditions, host signals also played an important role. The potato cyst nematode parasitic stage requires specific signals from the host plant for the hatching [55].
2.3 Importance of chemotaxis in host recognition
Previous studies revealed that plant parasitic nematodes especially root knot nematodes (RKN) showed attraction to certain chemicals released from the roots exudates which helps them in finding their specific host [56, 57]. The chemotaxis behavior played an important role in the hatching, survival and development of the plant parasitic nematodes. Though chemicals/compounds released from the host helps in the attraction of J2 stage but some of them work as repellents, some act as stimulants for hatching of the J2 stage and some act as inhibitors [58, 59]. The chemoattractants of RKN found to be released from other parts of the plants also [60]. Likewise, cyst nematodes (CN) also showed chemotactic behavior in response to the compounds secreted from the different parts of the plants [59].
2.3.1 Factors influencing hatching
The hatching in Heterodera glycines found stimulated by the compounds Glycinoeclepin A (Pentanor triterpenoid) secreted from the Kidney bean (Phaseolus vulgaris) root extracts [54, 61, 62]. 1,10-phenanthroline and tannic acid are synthetic chemicals also known for stimulating hatching in the H. glycines [63, 64]. Further, Root exudates of ryegrass (Lolium multiflorum) also stimulate the hatching in H. glycines [65]. Studies also showed that H. glycines cyst wall and egg homogenates and rinsates act as stimulants for the J2 stage hatching from the egg [66]. In Globodera rostochiensis, hatching found to be stimulated by the tetranortriterpenoid compound (Solanoeclepin A) and carbohydrate (glucose and fructose) secreted from the Solanum tuberosum and Solanum lycopersicum root diffusates [67, 68, 69]. The compound Asparagusic acid (Dithiolane) secreted from the Asparagus officinalis root extract is known to play a role in the inhibition of H. glycines and G. rostochiensis J2 stage hatching from the egg [70]. The compound allyl isothiocyanate (Isothiocyanate) is a synthetic chemical and formed from the degradation of allyl glucosinolates from Brassica plants found to inhibit the H. glycines and H. schachtii J2 hatching [71]. The root diffusates from Anthriseussilvestris, Arehangeliealitoralis, Pimpinella major, Pimpinella saxifraga, Heracleum sibirieum acts as hatching inhibitors for the CN mainly G. rostochiensis [72]. The synthetic chemical (E)-Chalcone (flavonoid) is also known for inhibiting the G. rostochiensis and G. pallida J2 hatching [73, 74]. The studies revealed that G. rostochiensis J2 hatching can be inhibited from the fermented Myrothecium verrucaria and root diffusates from tomato plants with the ABC-G33 and ABC-C6 transporter gene knocked down [75, 76]. The compound Bursehemin and Matairesinol extracted from the leaf of Bupleurum salicifolium inhibit the egg hatching in G. pallida [70]. The compound allyl isothiocyanate extracted from the Oil of black mustard affected the egg hatching in G. pallida [77]. The compounds secreted from the Bacillus cereus and Achromobacter xylosoxidans also inhibited the hatching of H. filipjevi [78].
2.3.2 Chemo attractants
Meloidogyne incognita found attracted to alcohol (Mannitol), organic acids (Vanillic acid, lauric acid), amino acids (Argenine, lysine), phenolics (Tannic acid), phytohormones (6-Dimethylallylaminopurine, salicylic acid, gibberellic acid, Indole-3-acetic acid), organic amines (Cadaverine, 1,3-diaminopropane, putrescine), Calcium chloride [79, 80]. The root diffusates and extracts from marigold, pepper, or soybean seedlings acts as chemoattractants and H. glycines J2 stages showed attraction to these [80]. The secondary metabolites of ethylene pathway in Arabidopsis thaliana also attracted parasitic stages of H. schachtii [81]. The parasitic J2 stage of G. rostochiensis is attracted to α-solanine (glycoalkaloids) and linalool [82, 83]. The G. pallida J2 stages showed attraction to synthetic compounds viz., indole acetic acid, ethephon, salicylic acid, mannitol, methyl jasmonate, α-solanine and linalool [82, 83].
2.3.3 Repellents
There are some compounds which act as repellents in order to prevent the host from PPN infection. The bulb extracts of Narcissus tazetta and metabolites produced during the ethylene pathway in Soybean and Arabidopsis thaliana repelled the population of H. glycines [84, 85]. The secondary metabolites released during Strigolactone signaling in Arabidopsis thaliana are found to repel the population of H. schachtii [86]. Further, diffusates from tomato plants root with the ABC-G33 and ABC-C6 transporter gene knocked down in the laboratory conditions revealed the repellent properties against the G. rostochiensis and G. pallida [76].
This is considered as one of the important genera in the filamentous fungi for controlling the PPN population. They worked against both root knot and cyst nematodes. They are parasitic to the developmental stages of cyst nematodes. They penetrate the developmental stages by exploiting the enzymes chitinase and protease which break down the extracellular layer. Consequently, the egg number decreases due to fungus parasitism and low level of parasitic stage hatching occurred [87]. In South Africa, Romulus products are prepared from the T. harzianum wettable powder formulation by Dagutat Biolab for controlling the population. In India, there are two commercial products viz., ECOSOM® and commander fungicide prepared from the T. harzianum wettable powder formulation by Agri Life and H.T.C Impex Private Limited, respectively. In Columbia, two commercial products Trichobiol and Trifesol based on the T. harzianum wettable powder formulation were prepared by the Control Biologico Integrado; Mora Jaramillo Arturo Orlando—Biocontrol and Biocultivos Agricultura Sostenible, respectively, as fungal nematicide [88]. The potential of the Trichoderma genus against PPN was presented in Table 1.
Different species of Trichoderma genus against PPN and their effects.
3.1.1.2 Mycorrhizal fungi
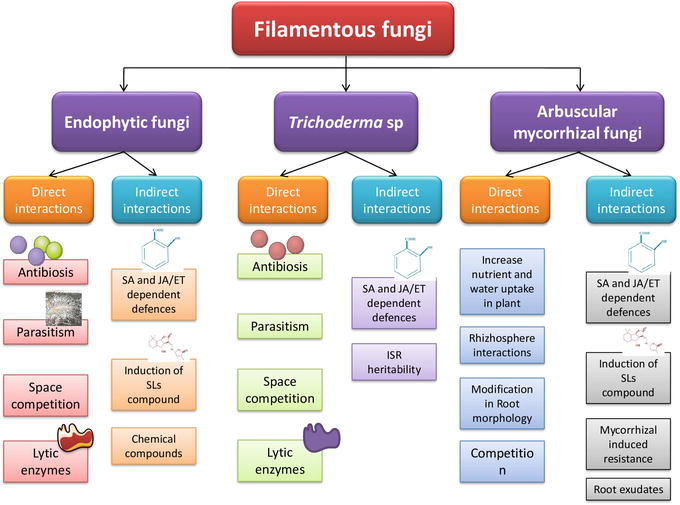
Previous findings on this genus did not report direct protection to the plants against PPN; however, it more efficiently worked on the plants, either by inducing morphological alterations in the roots, providing surplus nutrition or altering the environment for various interactions (Figure 3) [97, 98]. Recently, it’s been reported that they affect the PPN population and can be used in controlling the PPN infection. The Rhizophagus intraradices and Funneliformis mosseae found effective in controlling the population of RKN associated with Solanum lycopersicum [99]. Likewise, R. intraradices, R. mosseae, and R. etunicatum controlled the population of Meloidogyne sp. found in the association with peaches [100]. On the contrary, the increased population of some mycorrhizal fungi enhances the PPN population in the maize field [101]. There are some factors which affect the population of both PPN and fungi and their interaction with each other. The ecological and edaphic factors played a major role in the effects of mycorrhizal fungi against phytoparasitica as the concentration of N and P decreases the biocontrol potential of fungi and increases the population of PPN in the fields [102]. There are contradictions regarding the biocontrol potential of these fungi; however, several studies revealed the biocontrol of these against PPN in different fields [103, 104].
Figure 3.
Direct and indirect interactions of filamentous fungi with root system responsible for inducing induced resistance in plants against the PPN [87]. SA = salicylic acid; JA = jasmonic acid; SLs = strigolactones; ET = ethylene; ISR = induced systemic resistance.
3.1.1.3 Endophytic fungi
The endophytic fungi are also considered for controlling the PPN infection in the field. The studies revealed that these fungi possess the potential of attacking, killing, immobilizing, repelling, interfering with the developmental cells and arouse confusion during host seeking in order to control the PPN population [105]. Acremonium implicatum is found regulating the population M. incognita present in the rhizhosphere of Solanum lycopersicum whereas, F. oxysporum regulates the Pratylenchus goodeyi population in the fields [106]. Both of these fungi worked against the PPN and reduced their population by parasitizing and killing the developmental stages. Furthermore, they produced secondary metabolites against the PPN population which restricts their growth. The Chaetomium globosum releases these compounds which affects the growth of Meloidogyne species in the field [107]. Daldinia cf. concentric produced volatile compounds in the field against the Meloidogyne species which in turn restrict the growth [108]. The compound of alternariol 9-methyl ether released by the fungi regulates the population of B. xylophilus [109]. Though they are promising biocontrol agents in controlling the PPN, some of the entomopathogenic fungus (EPF) found to favor the PPN population. Pochonia sp. found to be a potent biocontrol agent in controlling the RKN and CN population. The species of this genus exploited in the formation of commercial products for controlling the PPN infection in the fields. The commercial product KlamiC® (Rothamsted Research and Centro Nacional de Sanidad Agropecuaria) based on granulate formulation from UK & Cuba, PcMR-1 strain (Clamitec, Myco solutions, Lda) based on liquid formulation from Portugal, Xianchongbike (Laboratory for Conservation and Utilization of Bio-resources, Yunnan University) based on liquid formulation from China and IPP-21 product from Italy. Paecilomyces lilacinus has been used in commercial products for controlling the PPN population in various countries. The commercial products viz., BIOACT®WP based on water-dispersible powder developed by Bayer Crop Science in USA, PL Gold based on wettable powder formulation by BASF Worldwide, Becker Underwood & PL 251based on water-dispersible granulate by Biological Control Products in South Africa, Stanes Bio Nematon based on liquid or powder Imported from T. Stanes and Company Limited (India) and Gaara Company (Egypt) and BIOCON based on wettable powder developed by Asiatic Technologies Incorporation, Philippines. In India, different companies developed commercial products based on the wettable powder and liquid formulation viz., Shakti Paecil (Shakti Biotech), PAECILO® (Agri Life) and Nematofree (International Panaacea Limited) from India developed for the PPN control. The other formulations from India are Paecilon (Enpro Bio Sciences Private Limited) based on liquid formulation and Gmaxbioguard (GreenmaxAgro Tech) based on talc-based carrier formulation [88].
3.1.1.4 Aspergillus
This genus fungus showed parasitism to the PPN developmental stages and provided protection to the plants [110]. The Aspergillus niger control the population of M. incognita associated with mung bean, okra and eggplant [111]. Further, M. javanica associated with pigeonpea and tomato population was negatively affected in the presence of A. niger [112]. The M. arenaria population associated with tomatoes was reduced in the presence of A. niger [113]. The commercial products of A. niger have been developed in India using different formulations. The soluble (liquid) concentrate and suspension concentrate formulation has been used for developing Kalisena products by Cadila Pharmaceutical Limited. The other commercial products viz., Beej Bandhu, Pusa Mrida and Kalasipahi have been developed from the wettable powder and capsule formulation [88].
3.1.2 Bacteria
There are some bacterial species which are found efficacious against the phytoparasitica nematode. The Pasteuria penetrans attaches itself to the PPN and their germination leads to the filling of the PPN body with endospores. The Pseudomonas fluorescens regulate the population through antibiotics mediating suppression of PPN. The Bacillus firmus suppress the population through many ways such as providing protection to the roots, releasing phytohormone, destruction of chemoattractants etc. The B. thuringiensis mediates the protection through their proteins which acts against the nematodes. The B. subtilis mediates protection through the antibiotics mainly surfactin and iturin. The commercial products of these biocontrol agents were prepared by various companies in different countries. The commercial product based on P. penetrans viz., Econem by Nematech Company from Japan and Pasteuria Bioscience from USA. The commercial product based on Bacillus subtilis is Stanes Sting (Imported from T. Stanes and Company Limited, India, by Gaara company, Egypt), Quartzo, Nemix C and Presense (FMC Química do Brasil Ltda., Brazil) and Pathway Consortia® (Pathway Holdings, USA) [88].
3.1.3 Entomopathogenic nematodes
Entomopathogenic nematodes (EPNs) gaining attention from the past few years due to their inherent ability of controlling the pest population. However, the characteristics of EPN as controlling agent of pest population also provide safer options from switching harmful insecticides and pesticides [114]. The omnipresent nature of EPN makes them suitable for including in the Integrated Pest Management (IPM) programs. The EPN belongs to two major families that is, Heterorhabditidae and Steinernematidae which includes 19 species from the Heterorhabditis, 1 Neosteinernema and 126 Steinernema species [115, 116]. The parasitic nature of these nematodes observed in the insect’s host, however; several present studies suggested that they can also exploit in regulating plant parasitic nematodes population [117]. The detailed list of the PPN controlled by the EPN is provided in Table 2.
The predatory nematodes feed on the plant parasitic nematode and can be exploited in controlling the PPN infection [130]. Most of the predators belong to Mononchida, Dorylaimida, Diplogasterida and Aphelenchida and each of the order possess specialized feeding apparatus [131]. Clarkuspapillatus feeds on Aphelenchoides sp., Hemicriconemoides sp., Heteroderaschachtii, M. hapla, T. semipenetrans, Subanguinaradicicola [132]. Aporcelaimllus nivalis feeds on Anguinatritici, Aphelenchoides sp., Basiria sp., Helicotylenchus indicus, H. mangiferae, Heteroderamothi, H. oryzae, Hoplolaimus indicus, Longidorus sp., M. incognita, P. citri, Scutellonema sp., T. mashhoodi, T. semipenetrans, Trichodorus sp., X. americanum, X. insigne in laboratory conditions [132]. Aquatidesthornei found efficacious against A. tritici, H. indicus, H. mothi, H. oryzae, Longidorus sp., M. incognita, P. citri, T. mashhoodi, Paratrichodorus sp., X. americanum [132]. Studies on the Labronemavulvapapillatus suggested that it feeds on A. tritici, G. rostochiensis, M. naasi, X. index in laboratory conditions [132]. The studies on Mesodorylaimus sp., Aporcelaimellus sp., Neoactinolaimus sp., against R. reniformis, R. similis and M. incognita suggested their potential as a biocontrol agent [132]. Despite these experiments, limited studies conducted on the predatory potential of these groups if we compare the publication records from 2000 to 2019 [132]. Their biocontrol potential in greenhouse and in vivo conditions need to be explored in order to use them against the PPN.
3.2 Nematicides
3.2.1 Bionematicides
3.2.1.1 Antagonistic cultivated plants
There are some plants known for the production of compounds which restricts the growth of PPN [133]. These antagonistic/anthelmintic compounds are released in the soil where they regulate the PPN infection and protect the plants. There are so many species that release these anthelmintic compounds but some are found to play an important role in limiting the PPN infection [133]. The genus Tagetes releases compounds such as bithienyl and alpha-terthienyl derivatives helping in suppressing the PPN infection. There is another mechanism opted by this genus for controlling the PPN population such as favoring the animal species/plants which are natural predators of nematodes or inhibiting the population, which does not serve for the nutrition requirements to the PPN etc. [134]. The species of this genus found efficacious in controlling the root knot and lesion nematode in tomato plants [133]. Azadirahta indica found to be efficient in regulating the nematode population. Further, cruciferous plants have been known for releasing the harmful nematicidal compound. The compound isothiocynates released from these plants when tested against the nematodes found efficacious in population regulation. The usage of this compound as biofumigant against the phytoparasitica nematode was also tested and proved efficient. These compounds are not limited to these plants but other plants also release these volatile molecules acting against the PPN population. The hydrolysed cyanogenic glycoside (cyanide) released from the herbaceous plants found acting against the phytoparasitica nematode [134]. Crotalaria species are also exploited for regulating the infection in the fields. The galls produced during root knot nematode infection are found to be decreased in the presence of Crotalaria longirostrata by utilizing the harmful compounds released from the microbes [134]. Zea may affect the population of lesion nematodes when tested with oats and radish. The nematode population was affected due to the lowering reproductive rate [134]. Likewise, the plant health increases when Zea mays cropped with velvet or jack beans and lesion nematode population decreases in response to this [134].
3.2.1.2 Plant-related materials and compounds
There are some compounds isolated from the plants possessing anthelmintic properties that can be used for controlling the PPN infection. These compounds can be prepared from any part of the plant or sometimes whole plant for effective control. The prepared compound can be broadly categorized into acids, oils, alcohol etc. Though, most of these compounds have been prepared from the isolated metabolites of plants but this is not a necessary condition that these will be prepared only from plants. These can be prepared from other microorganisms as they also produce some similar compounds during metabolism [135]. The acetic acid production in the plants occurred during the metabolic pathways but this compound is also produced by the bacterium viz., Lactobacillus brevis [136]. This compound possesses the ability to damage the J2stage of root knot nematodes leading to decreased population. Besides this, compounds produced from decomposition of other materials in the soil such as amino acids, butyric acids etc. also cause damage to the phytoparsitica nematode [134]. The compounds released from the T. viride and Secale cereale affected the PPN infection [134]. Various compounds extracted from the neem which found efficacious against the PPN. Furthermore, the presence of these compounds in the rhizosphere of plants provides protection to the host as the absorption of these compounds in the roots develops resistance against these nematodes. Extensive research has been carried out in this field in order to find the new and effective compounds for management of PPN [134]. The essential oils extracted from the plants were tested against the PPN and differences were found [134]. The essential oils namely carvone, terpinen-4-ol etc. found efficacious against the RKN species in laboratory conditions. The other PPN populations of R. reniformis, Criconemella and Hoplolaimus found resistant against these oils and less mortality observed. The essential oils of Dysphania ambrosioides found efficacious in managing these parasitic nematodes [134]. Compounds extracted from the Paenoiarockii and Camellia oleifera were found efficient in controlling the developmental stages of the RKN [134].
3.3 Host resistance
ISR is an induced systemic resistance in the hosts against the parasites and pathogens causing disease. In this, the host defense system strengthens against the agents’ causing diseases through certain compounds [137]. The resistance in the host developed from a particular infection after suffering from the infection earlier and termed as induced resistance [138]. The induced resistance is further classified into two broad categories mainly SAR and ISR [139]. The SAR, that is, Systemic acquired resistance controls the disease-causing agent and decreases the harmful effects of the when compared with other plants. This SAR in the host plants found to be in coordination of the production of salicylic acid during the diseases [140]. The production of salicylic acid mediates the induction of PR-1 gene causing the degradation of pathogen walls. The ISR resistance is found to be regulated by the Jasmonic acid and ethylene pathways. The ISR does not depend upon the salicylic acid and does not show association with the PR genes expression [141]. There are some chemicals identified which provide resistance to the host plants against these parasites [142]. The successful management of lesion and burrowing nematodes found to be based on these chemicals which decrease the PPN infection and enhance the plant growth by providing resistance [143].
The PPN infection in the agricultural sectors is one of the major problems in the world. The exploitation of biological control agents especially, fungi, bacteria and EPN could be useful in management of PPN as suggested from research. Further, the use of these agents with other microorganisms or chemical/bionematicide can enhance their efficacy against the PPN. Till now, the use of chemical nematicide against the PPN has some drawbacks which draw the attention towards using the biopesticide. To use EPN agents in the fields it’s necessary to develop a successful formulation and extensive studies on their virulence activity against the PPN in in vivo condition are required. On the contrary, fungal and bacterial nematicides based commercial products can be easily assessed in the management of PPN.
1.Bernard GC, Egnin M, Bonsi C. The impact of plant-parasitic nematodes on agriculture and methods of control. Nematology-Concepts, Diagnosis and Control. 2017;1:121-151
2.Jones JT, Haegeman A, Danchin EG, Gaur HS, Helder J, Jones MG, et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Molecular Plant Pathology. 2013;14(9):946-961
3.Mejias J, Truong NM, Abad P, Favery B, Quentin M. Plant proteins and processes targeted by parasitic nematode effectors. Frontiers in Plant Science. 2019;10:970
4.Zinovieva SV. Co-adaptation mechanisms in plant-nematode systems. Parazitologiia. 2014;48(2):110-130
5.Vieira P, Gleason C. Plant-parasitic nematode effectors—Insights into their diversity and new tools for their identification. Current Opinion in Plant Biology. 2019;50:37-43
6.Hartman GL, Rupe JC, Sikora EJ, Domier LL, Davis JA, Steffey KL. Compendium of Soybean Diseases and Pests. St. Paul, MN: American Phytopathological Society; 2015
7.El-Saadony MT, Abuljadayel DA, Shafi ME, Albaqami NM, Desoky ES, El-Tahan AM, et al. Control of foliar phytoparasitic nematodes through sustainable natural materials: Current progress and challenges. Saudi Journal of Biological Sciences. 2021;28(12):7314-7326
8.Sasser JN, Freckman DW. World perspective on nematology: The role of society. In: Veech JA, Dickson DW, editors. Vistas on Nematology: A Commemoration of the 25th Anniversary of the Society of Nematologists. Hyattsville, Maryland: Society of Nematologists Inc.; 1987. pp. 7-14
9.Ghaderi R, Karssen G. An updated checklist of Meloidogyne Göldi, 1887 species, with a diagnostic compendium for second-stage juveniles and males. Journal of Crop Protection. 2020;9(2):183-193
10.Jones MG. Host cell responses to endoparasitic nematode attack: Structure and function of giant cells and syncytia. Annals of Applied Biology. 1981;97(3):353-372
11.Von Mende N. Invasion and migration behaviour of sedentary nematodes. In: Fenol C, Grundler FMW, Ohl SA, editors. Cellular and Molecular Aspects of Plant-Nematode Interactions. The Netherlands: Kluwer Academic Publishers; 1997. pp. 51-64
12.Jagdale S, Rao U, Giri AP. Effectors of root-knot nematodes: An arsenal for successful parasitism. Frontiers in Plant Science. 2021;12:800030
13.Leelarasamee N, Zhang L, Gleason C. The root-knot nematode effector MiPFN3 disrupts plant actin filaments and promotes parasitism. PLoS Pathogens. 2018;14(3):e1006947
14.Shi Q , Mao Z, Zhang X, Zhang X, Wang Y, Ling J, et al. A Meloidogyne incognita effector MiISE5 suppresses programmed cell death to promote parasitism in host plant. Scientific Reports. 2018;8(1):7256
15.Xie J, Li S, Mo C, Wang G, Xiao X, Xiao Y. A novel Meloidogyne incognita effector Misp12 suppresses plant defense response at latter stages of nematode parasitism. Frontiers in Plant Science. 2016;7:964
16.Wubben MJ, Gavilano L, Baum TJ, Davis EL. Sequence and spatiotemporal expression analysis of CLE-motif containing genes from the reniform nematode (Rotylenchulusreniformis Linford & Oliveira). Journal of Nematology. 2015;47(2):159
17.Gleason C, Polzin F, Habash SS, Zhang L, Utermark J, Grundler FM, et al. Identification of two Meloidogyne hapla genes and an investigation of their roles in the plant-nematode interaction. Molecular Plant-Microbe Interactions. 2017;30(2):101-112
18.Lin B, Zhuo K, Chen S, Hu L, Sun L, Wang X, et al. A novel nematode effector suppresses plant immunity by activating host reactive oxygen species-scavenging system. New Phytologist. 2016;209(3):1159-1173
19.Chen J, Lin B, Huang Q , Hu L, Zhuo K, Liao J. A novel Meloidogyne graminicola effector, MgGPP, is secreted into host cells and undergoes glycosylation in concert with proteolysis to suppress plant defenses and promote parasitism. PLoS Pathogens. 2017;13(4):e1006301
20.Naalden D, Haegeman A, de Almeida-Engler J, Birhane Eshetu F, Bauters L, Gheysen G. The Meloidogyne graminicola effector Mg16820 is secreted in the apoplast and cytoplasm to suppress plant host defense responses. Molecular Plant Pathology. 2018;19(11):2416-2430
21.Zhuo K, Naalden D, Nowak S, Xuan Huy N, Bauters L, Gheysen G. A Meloidogyne graminicola C-type lectin, Mg01965, is secreted into the host apoplast to suppress plant defence and promote parasitism. Molecular Plant Pathology. 2019;20(3):346-355
22.Zhuo K, Chen J, Lin B, Wang J, Sun F, Hu L, et al. A novel Meloidogyne enterolobii effector MeTCTP promotes parasitism by suppressing programmed cell death in host plants. Molecular Plant Pathology. 2017;18(1):45-54
23.Verma A, Lee C, Morriss S, Odu F, Kenning C, Rizzo N, et al. The novel cyst nematode effector protein 30D08 targets host nuclear functions to alter gene expression in feeding sites. New Phytologist. 2018;219(2):697-713
24.Habash SS, Sobczak M, Siddique S, Voigt B, Elashry A, Grundler FM. Identification and characterization of a putative protein disulfide isomerase (HsPDI) as an alleged effector of Heteroderaschachtii. Scientific Reports. 2017;7(1):13536
26.Vieira P, Eves-Van Den Akker S, Verma R, Wantoch S, Eisenback JD, Kamo K. The Pratylenchus penetrans transcriptome as a source for the development of alternative control strategies: Mining for putative genes involved in parasitism and evaluation of in planta RNAi. PLoS One. 2015;10(12):e0144674
27.Barnes SN, Wram CL, Mitchum MG, Baum TJ. The plant-parasitic cyst nematode effector GLAND4 is a DNA-binding protein. Molecular Plant Pathology. 2018;19(10):2263-2276
28.Vijayapalani P, Hewezi T, Pontvianne F, Baum TJ. An effector from the cyst nematode Heteroderaschachtii derepresses host rRNA genes by altering histone acetylation. The Plant Cell. 2018;30(11):2795-2812
29.Liu J, Peng H, Cui J, Huang W, Kong L, Clarke JL, et al. Molecular characterization of a novel effector expansin-like protein from Heteroderaavenae that induces cell death in Nicotiana benthamiana. Scientific Reports. 2016;6(1):35677
30.Noon JB, Qi M, Sill DN, Muppirala U, Eves-van den Akker S, Maier TR, et al. A plasmodium-like virulence effector of the soybean cyst nematode suppresses plant innate immunity. New Phytologist. 2016;212(2):444-460
31.Mei Y, Wright KM, Haegeman A, Bauters L, Diaz-Granados A, Goverse A, et al. The Globodera pallida SPRYSEC effector Gp SPRY-414-2 that suppresses plant defenses targets a regulatory component of the dynamic microtubule network. Frontiers in Plant Science. 2018;9:1019
32.Lilley CJ, Maqbool A, Wu D, Yusup HB, Jones LM, Birch PR, et al. Effector gene birth in plant parasitic nematodes: Neofunctionalization of a housekeeping glutathione synthetase gene. PLoS Genetics. 2018;14(4):e1007310
33.Vieira P, Maier TR, Eves-van den Akker S, Howe DK, Zasada I, Baum TJ, et al. Identification of candidate effector genes of Pratylenchus penetrans. Molecular Plant Pathology. 2018;19(8):1887-1907
34.Fosu-Nyarko J, Tan JA, Gill R, Agrez VG, Rao U, Jones MG. D e novo analysis of the transcriptome of P ratylenchuszeae to identify transcripts for proteins required for structural integrity, sensation, locomotion and parasitism. Molecular Plant Pathology. 2016;17(4):532-552
35.Huang X, Xu CL, Chen WZ, Chen C, Xie H. Cloning and characterization of the first serine carboxypeptidase from a plant parasitic nematode, Radopholussimilis. Scientific Reports. 2017;7(1):4815
36.Wang K, Li Y, Huang X, Wang DW, Xu CL, Xie H. The cathepsin S cysteine proteinase of the burrowing nematode Radopholussimilis is essential for the reproduction and invasion. Cell & Bioscience. 2016;6:1-5
37.Li Y, Wang K, Xie H, Wang DW, Xu CL, Huang X, et al. Cathepsin B cysteine proteinase is essential for the development and pathogenesis of the plant parasitic nematode Radopholussimilis. International Journal of Biological Sciences. 2015;11(9):1073
38.Li Y, Wang K, Xie H, Wang YT, Wang DW, Xu CL, et al. A nematode calreticulin, Rs-CRT, is a key effector in reproduction and pathogenicity of Radopholussimilis. PLoS One. 2015;10(6):e0129351
39.Zhang C, Xie H, Cheng X, Wang DW, Li Y, Xu CL, et al. Molecular identification and functional characterization of the fatty acid-and retinoid-binding protein gene Rs-far-1 in the burrowing nematode Radopholussimilis (Tylenchida: Pratylenchidae). PLoS One. 2015;10(3):e0118414
40.Peng H, Gao BL, Kong LA, Yu Q , Huang WK, He XF, et al. Exploring the host parasitism of the migratory plant-parasitic nematode Ditylenchus destuctor by expressed sequence tags analysis. PLoS One. 2013;8(7):e69579
41.Hu LJ, Wu XQ , Li HY, Zhao Q , Wang YC, Ye JR. An effector, BxSapB1, induces cell death and contributes to virulence in the pine wood nematode Bursaphelenchusxylophilus. Molecular Plant-Microbe Interactions. 2019;32(4):452-463
42.Shinya R, Morisaka H, Kikuchi T, Takeuchi Y, Ueda M, Futai K. Secretome analysis of the pine wood nematode Bursaphelenchusxylophilus reveals the tangled roots of parasitism and its potential for molecular mimicry. PLoS One. 2013;8(6):e67377
43.Espada M, Silva AC, Eves van den Akker S, Cock PJ, Mota M, Jones JT. Identification and characterization of parasitism genes from the pinewood nematode Bursaphelenchusxylophilus reveals a multilayered detoxification strategy. Molecular Plant Pathology. 2016;17(2):286-295
44.Tarjan AC. Longevity of Radopholussimilis (cobb) in host-free soil. Nematologica. 1961;6:170-175
45.Bird AF, Buttrose MS. Ultrastructural changes in the nematode Anguinatritici associated with an hydrobiosis. Journal of Ultrastructure Research. 1974;48(2):177-189
46.Perry RN, Moens M. Survival of parasitic nematodes outside the host. In: Molecular and Physiological Basis of Nematode Survival. Wallingford UK: CAB International; 2011. pp. 1-27
47.Van Megen H, Van Den Elsen S, Holterman M, Karssen G, Mooyman P, Bongers T, et al. A phylogenetic tree of nematodes based on about 1200 full-length small subunit ribosomal DNA sequences. Nematology. 2009;11:927-950
48.Hunt DJ, Luc M, Manzanilla-López RH. Plant parasitic nematodes in subtropical and tropical agriculture. In: Luc M, Sikora RA, Bridge J, editors. Identification, Morphology and Biology of Plant Parasitic Nematodes. 2nd ed. Wallingford: CABI publishing; 2005. pp. 11-52
49.Yang D, Chen C, Liu Q , Jian H. Comparative analysis of pre- and postparasitic transcriptomes and mining pioneer effectors of heteroderaavenae. Cell & Bioscience. 2017;7:1-18
50.Proudfoot L, Kusel JR, Smith HV, Harnett W, Worms MJ, Kennedy MW. Rapid changes in the surface of parasitic nematodes during transition from pre- to post-parasitic forms. Parasitology. 1993;107:107-117
51.McCarter JP, Mitreva MD, Martin J, Dante M, Wylie T, Rao U, et al. Analysis and functional classification of transcripts from the nematode meloidogyne incognita. Genome Biology. 2003;4:1-19
52.Popeijus H, Blok VC, Cardle L, Bakker E, Phillips MS, Helder J, et al. Analysis of genes expressed in second stage juveniles of the potato cyst nematodes globoderarostochiensis and G. pallida using the expressed sequence tag approach. Nematology. 2000;2:567-574
53.Schroeder NE, Mac Guidwin AE. Behavioural quiescence reduces the penetration and toxicity of exogenous compounds in second-stage juveniles of heteroderaglycines. Nematology. 2010;12:277-287
54.Sikder MM, Vestergård M. Impacts of root metabolites on soil nematodes. Frontiers in Plant Science. 2020;10:1-18
55.Perry RN, Clarke AJ. Hatching mechanisms of nematodes. Parasitology. 1981;83:435-449
56.Reynolds AM, Dutta TK, Curtis RH, Powers SJ, Gaur HS, Kerry BR. Chemotaxis can take plant-parasitic nematodes to the source of a chemo-attractant via the shortest possible routes. Journal of the Royal Society Interface. 2011;8(57):568-577
57.Liu W, Jones AL, Gosse HN, Lawrence KS, Park SW. Validation of the chemotaxis of plant parasitic nematodes toward host root exudates. Journal of Nematology. 2019;51(1):1-10
58.Oota M, Tsai AY, Aoki D, Matsushita Y, Toyoda S, Fukushima K, et al. Identification of naturally occurring polyamines as root-knot nematode attractants. Molecular Plant. 2020;13(4):658-665
59.Ochola J, Coyne D, Cortada L, Haukeland S, Ng'ang'a M, Hassanali A, et al. Cyst nematode bio-communication with plants: Implications for novel management approaches. Pest Management Science. 2021;77(3):1150-1159
60.Tsai AY, Iwamoto Y, Tsumuraya Y, Oota M, Konishi T, Ito S, et al. Root-knot nematode chemotaxis is positively regulated by l-galactose sidechains of mucilage carbohydrate rhamnogalacturonan-I. Science Advances. 2021;7(27):eabh 4182
61.Masler E, Perry R. In: Perry RN, Moens M, Jones JT, editors. Hatch, Survival and Sensory Perception Cyst Nematodes. Wallingford, UK: C.A.B.I; 2018. pp. 44-73
62.Bohlmann H. Introductory chapter on the basic biology of cyst nematodes. Advances in Botanical Research. 2015;73:33-59
63.Nonaka S, Katsuyama T, Kondo T, Sasaki Y, Asami T, Yajima S, et al. 1, 10-Phenanthroline and its derivatives are novel hatching stimulants for soybean cyst nematodes. Bioorganic and Medicinal Chemistry Letters. 2016;26(21):5240-5243
64.Renčo M, Sasanelli N, Papajová I, Maistrello L. Nematicidal effect of chestnut tannin solutions on the potato cyst nematode Globoderarostochiensis (Woll.) Barhens. Helminthologia. 2012;49:108-114
65.Riga E, Topp E, Potter J, Welacky T, Anderson T, Tenuta A. The impact of plant residues on the soybean cyst nematode, Heterodera glycines. Canadian Journal of Plant Pathology. 2001;23(2):168-173
66.Charlson DV, Tylka GL. Heteroderaglycines cyst components and surface disinfestants affect H. Glycines hatching. Journal of Nematology. 2003;35(4):458
67.Tanino K, Takahashi M, Tomata Y, Tokura H, Uehara T, Narabu T. Total synthesis of Solanoeclepin A. Nature Chemistry. 2011;3:484-488
68.Jones P, Byrne J, Devine K. In vitro studies on the relative availability and mobility in soil of natural hatching factors for the potato cyst nematodes, Globodera rostochiensis and G. pallida. Nematology. 2001;3(1):75-83
69.Ganapati Reddy P, Chun B-K, Zhang H-R, Rachakonda S, Ross BS, Sofia MJ. Stereoselective synthesis of PSI-352938: A β-D-2′-deoxy-2′-α-fluoro-2′-β-C-methyl-3′,5′-cyclic phosphate nucleotide prodrug for the treatment of HCV. Journal of Organic Chemistry. 2011;76:3782-3790
70.Zhou L, Wang J, Wang K, Xu J, Zhao J, Shan T, et al. Secondary metabolites with antinematodal activity from higher plants. Studies in Natural Products Chemistry. 2012;37:67-114
71.Yu Q , Tsao R, Chiba M, Potter J. Selective nematicidal activity of allyl isothiocyanate. Journal of Food, Agriculture and Environment. 2005;3:218-221
72.Ghisalberti EL. Secondary metabolites with antinematodal activity. Studies in Natural Products Chemistry. 2002;26:425-506
73.Díaz-Tielas C, Graña E, Reigosa MJ, Sánchez-Moreiras AM. Biological activities and novel applications of chalcones. Planta Daninha. 2016;34:607-616
74.Silva FJ, Campos VP, Oliveira DF, Gomes VA, Barros AF, Din ZU, et al. Chalcone analogues: Synthesis, activity against Meloidogyne incognita, and in silico interaction with cytochrome P 450. Journal of Phytopathology. 2019;167(4):197-208
75.Perry R, Warrior P, Kerry B, Twomey U. Effects of the biological nematicide, DiTera, on hatching of Globoderarostochiensis and G. Pallida. Journal of Nematology. 2000;2:355-362
76.Cox D, Dyer S, Weir R, Cheseto X, Sturrock M, Coyne D, et al. ABC transporters alter plant-microbe-parasite interactions in the rhizosphere. Science Reports. 2019;9:526582
77.Wood C, Kenyon DM, Cooper JM. Allyl isothiocyanate shows promise as a naturally produced suppressant of the potato cyst nematode, Globodera pallida, in biofumigation systems. Nematology. 2017;19(4):389-402
78.Zhang J, Li Y, Yuan H, Sun B, Li H. Biological control of the cereal cyst nematode (Heteroderafilipjevi) by Achromobacterxylosoxidans isolate 09X01 and Bacillus cereus isolate 09B18. Biological Control. 2016;92:1-6
79.Fleming TR, Maule AG, Fleming CC. Chemosensory responses of plant parasitic nematodes to selected phytochemicals reveal long-term habituation traits. Journal of Nematology. 2017;49(4):462
80.Wang C, Masler EP, Rogers ST. Responses of Heteroderaglycines and Meloidogyne incognita infective juveniles to root tissues, root exudates, and root extracts from three plant species. Plant Disease. 2018;102(9):1733-1740
81.Kammerhofer N, Radakovic Z, Regis JM, Dobrev P, Vankova R, Grundler FM, et al. Role of stress-related hormones in plant defence during early infection of the cyst nematode Heteroderaschachtii in Arabidopsis. New Phytologist. 2015;207(3):778-789
82.Būda V, Čepulytė-Rakauskienė R. The effects of α-solanine and zinc sulphate on the behaviour of potato cyst nematodes Globoderarostochiensis and G. Pallida. Nematology. 2015;17(9):1105-1111
83.Būda V, Čepulyt Ė-Rakauskien Ė R. The effect of linalool on second-stage juveniles of the potato cyst nematodes Globoderarostochiensis and G. Pallida. Journal of Nematology. 2011;43(3-4):149
84.Hu Y, You J, Li C, Williamson VM, Wang C. Ethylene response pathway modulates attractiveness of plant roots to soybean cyst nematode heteroderaglycines. Scientific Reports. 2017;7(1):41282
85.Hu Y, You J, Li C, Pan F, Wang C. Assessing the effects of water extract of Narcissus tazetta bulb on hatching, mortality, chemotaxis and reproduction of the soybean cyst nematode, Heterodera glycines. Nematology. 2019;22(1):53-62
86.Escudero Martinez CM, Guarneri N, Overmars H, van Schaik C, Bouwmeester H, Ruyter-Spira C, et al. Distinct roles for strigolactones in cyst nematode parasitism of Arabidopsis roots. European Journal of Plant Pathology. 2019;154:129-140
87.Poveda J, Abril-Urias P, Escobar C. Biological control of plant-parasitic nematodes by filamentous fungi inducers of resistance: Trichoderma, mycorrhizal and endophytic fungi. Frontiers in Microbiology. 2020;11:992
88.Abd-Elgawad MM, Askary TH. Fungal and bacterial nematicides in integrated nematode management strategies. Egyptian Journal of Biological Pest Control. 2018;28(1):1-24
89.Zhang S, Gan Y, Xu B, Xue Y. The parasitic and lethal effects of Trichoderma longibrachiatum against Heterodera avenae. Biological Control. 2014;72:1-8
90.Zhang S, Gan Y, Ji W, Xu B, Hou B, Liu J. Mechanisms and characterization of Trichoderma longibrachiatum T6 in suppressing nematodes (Heterodera avenae) in wheat. Frontiers in Plant Science. 2017;8:1491
91.Kumar P, Chand R. Bio-efficacy of Trichoderma harzianum against root knot nematode Meloidogyne incognita on Brinjal. Annals of Plant Protection Sciences. 2015;23(2):361-364
92.Brahma U, Borah A. Management of Meloidogyne incognita on pea with bioagents and organic amendment. Indian Journal of Nematology. 2016;46(1):58-61
93.Devi TS, Mahanta B, Borah A. Comparative efficacy of Glomus fasciculatum, Trichoderma harzianum, carbofuran and carbendazim in management of Meloidogyne incognita and Rhizoctonia solani disease complex on brinjal. Indian Journal of Nematology. 2016;46(2):161-164
94.Deori R, Borah A. Efficacy of Glomus fasciculatum, Trichoderma harzianum for the management of Meloidogyne incognita and Rhizoctonia solani disease complex in green gram. Indian Journal of Nematology. 2016;46(1):61-64
95.Contina JB, Dandurand LM, Knudsen GR. Use of GFP-tagged Trichoderma harzianum as a tool to study the biological control of the potato cyst nematode Globodera pallida. Applied Soil Ecology. 2017;1(115):31-37
96.Khan MR, Ahmad I, Ahamad F. Effect of pure culture and culture filtrates of Trichoderma species on root-knot nematode, Meloidogyne incognita infesting tomato. Indian Phytopathology. 2018;71:265-274
97.Schouteden N, De Waele D, Panis B, Vos CM. Arbuscular mycorrhizal fungi for the biocontrol of plant-parasitic nematodes: A review of the mechanisms involved. Frontiers in Microbiology. 2015;6:1280
98.Wani KA, Manzoor J, Shuab R, Lone R. Arbuscular mycorrhizal fungi as biocontrol agents for parasitic nematodes in plants. In: Mycorrhiza-nutrient uptake, biocontrol, ecorestoration. Cham: Springer; 2017. pp. 195-210
99.Marro N, Caccia M, Doucet ME, Cabello M, Becerra A, Lax P. Mycorrhizas reduce tomato root penetration by false root-knot nematode Nacobbus aberrans. Applied Soil Ecology. 2018;124:262-265
100.Calvet C, Pinochet J, Hernández-Dorrego A, Estaún V, Camprubí A. Field microplot performance of the peach-almond hybrid GF-677 after inoculation with Arbuscular mycorrhizal fungi in a replant soil infested with root-knot nematodes. Mycorrhiza. 2001;10:295-300
101.Brito OD, Hernandes I, Ferreira JC, Cardoso MR, Alberton O, Dias-Arieira CR. Association between Arbuscular mycorrhizal fungi and Pratylenchus brachyurus in maize crop. Chilean Journal of Agricultural Research. 2018;78(4):521-527
102.Ferreira BS, Santana MV, Macedo RS, Silva JO, Carneiro MA, Rocha MR. Co-occurrence patterns between plant-parasitic nematodes and arbuscular mycorrhizal fungi are driven by environmental factors. Agriculture, Ecosystems and Environment. 2018;265:54-61
103.Emery SM, Reid ML, Bell-Dereske L, Gross KL. Soil mycorrhizal and nematode diversity vary in response to bioenergy crop identity and fertilization. GCB Bioenergy. 2017;9(11):1644-1656
104.Alvarado-Herrejón M, Larsen J, Gavito ME, Jaramillo-López PF, Vestberg M, Martínez-Trujillo M, et al. Relation between arbuscular mycorrhizal fungi, root-lesion nematodes and soil characteristics in maize agroecosystems. Applied Soil Ecology. 2019;135:1-8
105.Schouten A. Mechanisms involved in nematode control by endophytic fungi. Annual Review of Phytopathology. 2016;54:121-142
106.Mwaura P, Dubois T, Losenge T, Coyne D, Kahangi E. Effect of endophytic Fusarium oxysporum on paralysis and mortality of Pratylenchus goodeyi. African Journal of Biotechnology. 2010;9(8):1130-1134
107.Khan B, Yan W, Wei S, Wang Z, Zhao S, Cao L, et al. Nematicidal metabolites from endophytic fungus Chaetomium globosum YSC5. FEMS Microbiology Letters. 2019;366(14):fnz 169
108.Liarzi O, Bucki P, Braun Miyara S, Ezra D. Bioactive volatiles from an endophytic Daldinia cf. concentrica isolate affect the viability of the plant parasitic nematode Meloidogyne javanica. PLoS One. 2016;11(12):e0168437
109.Lou J, Yu R, Wang X, Mao Z, Fu L, Liu Y, et al. Alternariol 9-methyl ether from the endophytic fungus Alternaria sp. Samif 01 and its bioactivities. Brazilian Journal of Microbiology. 2016;47:96-101
110.Askary TH. Nematophagous fungi as biocontrol agents of phytonematodes. In: Biocontrol Agents of Phytonematodes. Wallingford, UK: CABI; 2015
111.Bhat MY, Wani AH. Bio-activity of fungal culture filtrates against root-knot nematode egg hatch and juvenile motility and their effects on growth of mung bean (Vigna radiata L. Wilczek) infected with the root-knot nematode, Meloidogyne incognita. Archives of Phytopathology and Plant Protection. 2012;45(9):1059-1069
112.Zareen A, Zaki MJ, Khan NJ. Effect of fungal filtrates of aspergillus species on development of root-knot nematodes and growth of tomato (Lycopersicon esculentum mill). Pakistan Journal of Biological Sciences. 2001;4(8):995-999
113.Mokbel AA, Obad IM, Ibrahim IK. The role of antagonistic metabolites in controlling root-knot nematode, Meloidogyne arenaria on tomato. Alexandria Journal of Agricultural Research. 2009;54(1):199-205
114.Khan RR, Ali RA, Ali A, Arshad M, Majeed S, Ahmed S, et al. Compatibility of entomopathogenic nematodes (Nematoda: Rhabditida) and the biocide, spinosad for mitigation of the armyworm, Spodoptera litura (F.)(Lepidoptera: Noctuidae). Egyptian Journal of Biological Pest Control. 2018;28:1-6
115.Sharma H, Rana A, Bhat AH, Chaubey AK. Entomopathogenic nematodes: Their characterization, bio-control properties and new perspectives. In: Nematodes-Recent Advances, Management and New Perspectives. London: Books on Demand; 2021. p. 187
116.Malan AP, Knoetze R, Kapp C, Tiedt L, Steinernemabakwenae n. sp. (Rhabditida: Steinernematidae), a new entomopathogenic nematode from South Africa. Nematology. 2023;25:275-293
117.El Aimani A, Houari A, Laasli SE, Mentag R, Iraqi D, Diria G, et al. Antagonistic potential of Moroccan entomopathogenic nematodes against root-knot nematodes, Meloidogyne javanica on tomato under greenhouse conditions. Scientific Reports. 2022;12(1):2915
118.Grewal PS, Martin WR, Miller RW, Lewis EE. Suppression of plant-parasitic nematode populations in turfgrass by application of entomopathogenic nematodes. Biocontrol Science and Technology. 1997;7:393-399
119.Grewal PS, Lewis EE, Venkatachari S. Allelopathy: A possible mechanism of suppression of plant-parasitic nematodes by entomopathogenic nematodes. Nematology. 1999;1(7):735-743
120.Fallon DJ, Kaya HK, Gaugler R, Sipes BS. Effects of etomopathiogenic nematodes on Meloidogyne javanica on tomatoes and soybeans. Journal of Nematology. 2002;34(3):239
121.Pérez EE, Lewis EE. Use of entomopathogenic nematodes to suppress Meloidogyne incognita on greenhouse tomatoes. Journal of Nematology. 2002;34(2):171
122.Molina JP, Dolinski C, Souza RM, Lewis EE. Effect of entomopathogenic nematodes (Rhabditida: Steinernematidae and Heterorhabditidae) on Meloidogyne mayaguensis Rammah and Hirschmann (Tylenchida: Meloidoginidae) infection in tomato plants. Journal of Nematology. 2007;39(4):338
123.Caccia M, Lax P, Doucet ME. Effect of entomopathogenic nematodes on the plant-parasitic nematode Nacobbus aberrans. Biology and Fertility of Soils. 2013;49:105-109
124.Javed N, Khan SA, Atiq M, Kamran M. Effect of Steinernema glaseri and Heterorhabditis indica on the plant vigour and root knot nematodes in tomato roots at different densities and time of applications. Pakistan Journal of Zoology. 2012;44(4):1165-1170
125.del Valle EE, Lax P, Rondán Dueñas J, Doucet ME. Effects of insect cadavers infected by Heterorhabditis bacteriophora and Steinernema diaprepesi on Meloidogyne incognita parasitism in pepper and summer squash plants. Agricultural Science and Research. 2013;40:109-118
126.Kepenekci I, Hazir S, Lewis EE. Evaluation of entomopathogenic nematodes and the supernatants of the in vitro culture medium of their mutualistic bacteria for the control of the root-knot nematodes Meloidogyne incognita and M. arenaria. Pest Management Science. 2016;72(2):327-334
127.Kepenekci I, Hazir S, Oksal E, Lewis EE. Application methods of Steinernema feltiae, Xenorhabdusb ovienii and Purpureocillium lilacinum to control root-knot nematodes in greenhouse tomato systems. Crop Protection. 2018;108:31-38
128.Khan SA, Javed N, Kamran M, Abbas H, Safdar A, ul Haq I. Management of Meloidogyne incognita race 1 through the use of entomopathogenic nematodes in tomato. Pakistan Journal of Zoology. 2016;48(3):763
129.Osman HA, Ameen HH, Hammam MM, El-Sayed GM, Elkelany US, Abd-Elgawad MM. Antagonistic potential of an Egyptian entomopathogenic nematode, compost and two native endophytic bacteria isolates against the root-knot nematode (Meloidogyne incognita) infecting potato under field conditions. Egyptian Journal of Biological Pest Control. 2022;32(1):137
130.Esser E. Biological control of nematodes by nematodes I. Dorylaims (Nematoda: Dorylaimida). Nematology Circular. 1987;144:4
131.Pervez R, Bilgrami AL, Yoshiga T, Kondo E. Feeding, attraction and aggregation behaviour of Mesodorylaimus bastiani and Aquatides thornei at the feeding site using Hirschmanniella oryzae as prey. International Journal of Nematology. 2000;10(2):207-214
132.Kanwar RS, Patil JA, Yadav S. Prospects of using predatory nematodes in biological control for plant parasitic nematodes—A review. Biological Control. 2021;160:104668
133.Grubišić D, Uroić G, Ivošević A, Grdiša M. Nematode control by the use of antagonistic plants. Agriculturae Conspectus Scientificus. 2018;83(4):269-275
134.Abd-Elgawad MM. Optimizing safe approaches to manage plant-parasitic nematodes. Plants. 2021;10(9):1911
135.Ntalli N, Ratajczak M, Oplos C, Menkissoglu-Spiroudi U, Adamski Z. Acetic acid, 2-undecanone, and (E)-2-decenal ultrastructural malformations on Meloidogyne incognita. Journal of Nematology. 2016;48(4):248
136.Seo HJ, Park AR, Kim S, Yeon J, Yu NH, Ha S, et al. Biological control of root-knot nematodes by organic acid-producing Lactobacillus brevis wikim 0069 isolated from kimchi. The Plant Pathology Journal. 2019;35(6):662
137.Conrath U. Systemic acquired resistance. Plant Signaling & Behavior. 2006;1(4):179-184
138.Hammerschmidt R. Introduction: Definitions and some history. In: Induced Resistance for Plant Defense. United States: Wiley; 2014. pp. 1-10
139.El-Deeb TS, Bakkar SM, Eltoony L, Zakhary MM, Kamel AA, Nafee AM, et al. The adipokine chemerin and fetuin-a serum levels in type 2 diabetes mellitus: Relation to obesity and inflammatory markers. The Egyptian Journal of Immunology. 2018;25(1):191-202
140.Ryals JA, Neuenschwander UH, Willits MG, Molina A, Steiner HY, Hunt MD. Systemic acquired resistance. The Plant Cell. 1996;8(10):1809
141.Vallad GE, Goodman RM. Systemic acquired resistance and induced systemic resistance in conventional agriculture. Crop Science. 2004;44(6):1920-1934
142.Malamy J, Sanchez-Casas P, Hennig J, Guo A, Klessig F. Dissection of the salicylic acid signaling pathway in tobacco. American Phytopathological Society. 1996;9:474-482
143.Chałańska A, Łabanowski G, Maciorowski R. Control efficacy of selected natural products against chrysanthemum foliar nematode–Aphelenchoides ritzemabosi (Schwartz, 1911) Steiner & Buhrer, 1932 Ocena skutecznościdziałanianiektórychsubstancjiipreparatówpochodzenianaturalnego w zwalczaniuwęgorkachryzantemowca–Aphelenchoidesritzemabosi (Schwartz, 1911) Steiner i Buhrer, 1932. Progress in Plant Protection. 2013;53(3):563-567
Written By
Himani Sharma and Ashok Kumar Chaubey
Submitted: 08 September 2023Reviewed: 10 September 2023Published: 08 November 2023
 Open access peer-reviewed chapter
Open access peer-reviewed chapter