 Open access peer-reviewed chapter
Open access peer-reviewed chapter
Abstract
Plant parasitic nematodes are microscopic organisms that inhabit soil and plant tissues. Among such organisms, those of the genera Meloidogyne, Heterodera, Globodera, and Pratylenchus spp., are the most harmful, as they affect more than 2500 different species of plants, generating worldwide economic losses of over US$100 billion per year. These nematodes constitute a notable threat to the country’s progress and food security. Almost half of the global market for nematicides, which corresponds to US$ 1 billion per year, is used to control these nematodes. Non-fumigant nematicides are the most widely used in their control; however, many of them, such as carbamates and organophosphorus, are banned by environmental protection agencies because of their undesirable effects on non-target organisms. In the last 10 years, a new series of nematicides have emerged with different mechanisms of action than the old non-fumigant nematicides. Tioxazafen and fluazaindolizine are some of the latest new-generation nematicides that have come on the market. The rational design of new nematicides through in silico approaches combined with studies of the genetics and biochemistry of these microorganisms will help to better understand their management and control, aiming to reduce the environmental impact caused by the irrational use of nematicides.
Keywords
- plant parasitic nematodes
- root-knot nematodes
- nematicide
- tioxazafen
- fluazaindolizine
1. Introduction
According to the United Nations (UN), the global population will reach 8.5 billion by 2030 [1]. As a result, the World Bank projects that food consumption will rise by 60–80% [2]. Although the Food and Agriculture Organization of the United Nations (FAO) has as one of its goals to end world poverty, the fact is that food security has been declining rapidly since 2015 [3]. Agricultural pests are clearly one of the many elements leading to a country’s or region’s food security being undermined [4]. Pests and plant diseases, according to the FAO, account for 40% of global food production losses, amounting to more than US$250 billion each year [5]. As a result, dealing with plant pests and diseases is critical to attaining higher agricultural sustainability and security, particularly in the post-pandemic age.
Although microorganisms play a vital role in agriculture, many of them act as phytopathogens, reducing agricultural yield seriously [6, 7]. Plant parasitic nematodes (PPNs) are microorganisms that have a deleterious impact on the harvested of numerous food crops such as soybean (
Because of their complex interaction with host plants, extensive host range, and the degree of damage caused by infection, root-knot nematodes (RKNs), cyst nematodes (CNs), and root-lesion nematodes (RLNs) were at the top of the list of the most economically important species [11]. Thus,
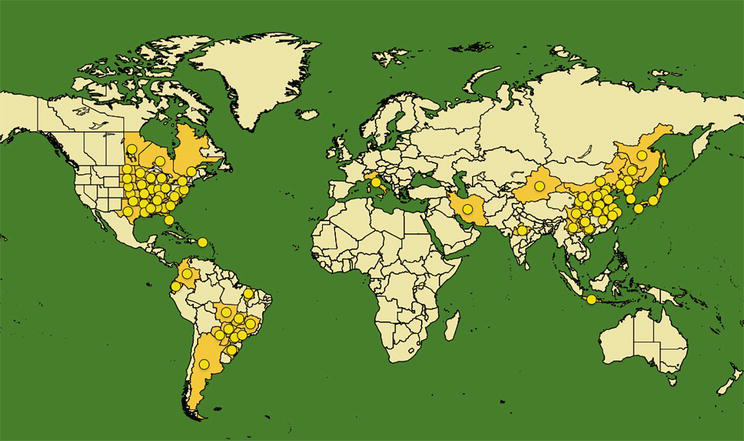
Figure 1.
Geographical distribution map Heterodera glycines. The yellow areas and circles correspond to regions of the world where the nematode is currently present. Source: EPPO global database (2023).
Some of the approaches used to manage and control PPNs include resistant cultivars, crop rotation, biological management, and chemical compounds [8]. Chemical methods are the most efficient and therefore most widely used in the management of PPNs; however, many of the chemicals used are highly hazardous to human and environmental health, including carbamates (CMs), organophosphorus (OPs), organic halides, and pyridinylmethylbenzamides [15].
The arsenal of nematicides accessible for the management of PPNs has been substantially restricted by current environmental protection laws. This has increased demand for novel nematicides that are more selective, less toxic, and compatible with sustainable agricultural guidelines. As a result, various nematicides have been introduced to the market over the last decades. This is the case of tioxazafen, Fluopyram, and fluazaindolizine, which are considered new-generation nematicides as they are less toxic to mammals and the environment, suggesting that they have mechanisms of action distinct from those observed for CMs and OPs types.
2. PPNs: overview
PPNs are microscopic and ubiquitous organisms that live in soil and plant tissues. Despite their inability to travel great distances, anthropogenic actions can lead them to be transmitted via contaminated plant material, soil, and agricultural machinery [11]. In addition to preventing plant roots from absorbing nutrients, PPNs may make plants more vulnerable to secondary phytopathogens such as fungi, bacteria, and viruses [16].
Many agriculturally important PPNs are members of the Tylenchida order, which includes endoparasites from the Heteroderidae and Pratylenchidae families. The Heteroderidae family includes the most important agricultural genera: RKNs (
PPNs, unlike free-living nematodes, have a hollow and protractile stylet that allows them to pass through plant cell walls, but not the plasma membrane, enabling them to feed on cell contents via the feeding tube that is solely connected with the stylet [19]. PPNs release a cocktail of polymeric or non-polymeric effector proteins that degrade and modify cytoplasmic components, causing the host cell’s physiology and morphology to change [20]. The force for the nematode to feed on the cellular contents is provided by a muscular pump in the basal nodes (metacorpus). In this process, a complex plant-parasite interaction occurs, involving a sequence of recognition and response events that vary depending on the kind of parasite and host plant [21]. Although many aspects of the pathogenesis process are unknown, it is evident that acquiring this information is necessary for the creation of new and more effective techniques of treatment and control.
3. Biology and parasitism of the main PPNs of agricultural importance
3.1 Root-knot nematodes (Meloidogyne spp.)
RKN are obligate biotrophic endoparasites that require a host plant to complete their life cycle [20]. There are more than 90 species, but only a few have worldwide agricultural value. The most damaging species include:
The RKNs life cycle starts when an adult female deposits their eggs (~500) in gelatinous-textured sacs (egg masses) generated by six anal glands (Figure 2). This gelatinous sac is made up of glycoproteins, which, in addition to protecting the egg, serve as a regulator of embryonic development by detecting changes in the temperature and humidity of the environment. With the advance in feeding, the female becomes more voluminous and the posterior section of her body is extended in such a way that allows the egg mass to be exposed outside the root, thus forming the nodes or galls [23]. The galls make it easier for microorganisms in the rhizosphere to come in contact with the eggs; however, their gelatinous structure has been shown to have antimicrobial properties that protect them from attack by microorganisms [24].

Figure 2.
Schematic representation of the root-knot nematode life cycle. 1: Egg in development, 2: J2 larvae entering root, 3 and 4: Swollen nematodes feeding in root, 5: Free male on the ground, 6: Mature female breaking through root surface with egg sac.
After embryogenesis, J1 undergoes the first molt (1° ecdysis), which will give birth to the infective form known as the second instar juvenile (J2). When temperature and humidity conditions are favorable, hatching of the eggs occurs, releasing the J2 nematodes. Unlike other PPNs, which require stimuli from plant roots for hatching,
J2 enters and migrates via the vascular cylinder of a vulnerable plant after penetrating very close to the root tip where there is a lack of completely developed endodermis (elongation zone) [30]. During this action, the intermediate lamella softens mechanically and enzymatically [31]. After traveling a short distance into the plant, J2 becomes sessile due to the atrophy of its somatic muscle (except for the head). The head is buried in the periphery of the vascular tissue cells (zone of cell differentiation), where it feeds (through a feeding tube) on the protoxylem and protophloem contents before the cells differentiate into specialized nurse cells known as giant cells (GCs) [32]. After the establishment of a permanent feeding site, the nematode becomes sedentary, growing significantly in size until it reaches maturity.
Although the mechanisms by which GCs are formed are not fully understood in detail, it is known that, during feeding, parasites secrete effector proteins that modify cell division events through reorganization of the cytoskeleton and signaling pathways [33]. This cocktail of effector molecules, which facilitate the establishment of the feeding site, is mainly produced by the dorsal gland. Peptides, virulence factors, β-1,4 endogluconases, polygalacturonases, xylanases, chorismate mutase, among others, are some of the molecules that compose the biochemical arsenal of the RKNs secretome [34].
The cell wall between the daughter nuclei is created in telophase during normal plant cell division; however, in the creation of GCs, the cell splits without cytokinesis and hence without cell wall development. This causes multinuclear GCs to form, as well as the proliferation of other organelles such as endoplasmic reticulum, mitochondria, and ribosomes [35]. These metabolic and physiological changes give GCs the characteristics of metabolically active cells, with the inner growth of the wall forming an extensive labyrinth network that increases the surface area of exchange with the associated membrane of xylem vessels [36]. Although less visible, phloem sieve components develop around large cells to ensure phloem continuity [37].
Compared with uninfected plants, tissue from RKNs-infected plants shows a significant increase in the phytohormone auxin [38]. Auxins, like ethylene, have a role in the formation of cell wall ingrowths by promoting the production of cell wall modifying proteins as well as proteins implicated in acidic growth (proton pumps) [39, 40]. These and others evidence imply that the parasite’s modulation of this phytohormone is a key factor in the creation of GCs. In addition to auxins, cytokinins, which are involved in cell division, are elevated in infected tissue, suggesting that they may actively contribute to the parasite-feeding site formation [41].
The plant defense system is also affected by the effector proteins controlling the phytohormone pathway: salicylic acid (SA) and jasmonic acid (JA). In general, the SA pathway protects against biotrophic and hemibiotrophic pathogens such as endoparasitic sedentary [42]. The JA pathway, on the other hand, is involved in plant defenses against necrotrophic pathogens and leaf-chewing insects [43].
The capacity of biotrophic and hemibiotrophic pathogens to modify plant defenses is important to host tissue colonization success. The first line of activated protection against RKNs is the pathogen-associated molecular pattern (PAMPs) triggered immunity (PTI) [44]. In response to the attack (PTI response), the plant triggers the synthesis of reactive oxygen species (ROS), as well as the production of protein kinases (MAPKs) and the activation of the JA and SA pathways [45]. SA regulates the expression of several genes involved in the synthesis of proteins mediating the pathogenic response, while the JA pathway regulates the expression of genes encoding for proteins involved in the synthesis of thionin, defensin, and phytoalexin, among others [45, 46]. The level of response and plant-parasite interaction seems to be influenced by the type of plant, as well as by the type of parasite and its infective state [20].
After the establishment of the feeding site, J2 undergoes significant morphological changes, including two successive molts (J3 and J4). Unlike J2, these last two stages do not feed, as they lack a functional stylet [47]. Under appropriate environmental conditions and with sufficient food availability, J4 undergoes a final ecdysis, the female acquires the classical pear shape (500 times the volume of J2) and the male is left free in the environment. Before reaching full maturity, the cells of the genital primordium of the female divide to give rise to the rectal glands, which will later secrete the gelatinous material with which she will protect her eggs. On the other hand, the male characteristics (testis and vas deferens) differentiate from the posterior end, where they connect with the rectum [25].
It has been recorded that, within the genus Meloidogyne, there are three different forms of reproduction. On the one side, there is the fertilization of oocytes by the male (amphimixis), while, on the other hand, there is parthenogenesis, which can be meiotic or mitotic. In the first case, there is a fusion of the pronucleus of an ovule, generated by meiosis, with a polar body (automixis); while, in the second case, the oocyte divides by mitosis, where one of the nuclei deteriorates and the other remains to give rise to the embryo [48, 49].
As in CNs, sex chromosomes are absent, so sex is strongly influenced, at least in parthenogenetic species, by environmental conditions as well as by host plant conditions. Lack of food, overcrowding, extreme temperatures, and soil dryness are some of the conditions that favor the development of males that reproduce by meiotic parthenogenesis [50]. These males rarely fertilize females, and when they do, mitotic parthenogenesis takes place without any fusion with the sperm nucleus.
3.2 Cyst nematodes (Heterodera and Globodera spp.)
After RKNs, the CNs constitute the second group of sedentary endoparasites with the greatest impact on world agriculture. Among the most important species are
Like RKNs, the life cycle comprises an egg stage, four juveniles (J1,2,3,4), and adults (male and female). However, there are important differences that will be discussed below.
Unlike Meloidogyne, in CNs, the eggs may be retained in the female’s body, or, depending on the species, deposited in a gelatinous sac. After their deaths, the female body transforms into the cyst, which protects and holds the eggs inside. Depending on its maturation stages, it can acquire different shades ranging from black-brown to yellow. These color changes are known to be due to the activity of the enzyme polyphenol oxidases, which, in addition to catalyzing the hydroxylation of phenolic compounds, also catalyze the polymerization of o-quinones on the cyst cuticle [51].
In the cyst, the embryo can remain for many years in a state of metabolic suspension (dormancy stage) when environmental conditions are unfavorable. This dormant state is essential for the survival and pathogenesis of the CNs. In general, normal embryo development requires the action of internal signaling molecules, which are regulated by biotic and abiotic conditions [52]. Not all species have this dormancy stage in their life cycle; however, for those that do, it can be of two types depending on the dormancy strategy used: quiescence and diapause. Quiescence can be an obligate or facultative response to poor environmental conditions, which is reversed when external conditions are ideal. On the other hand, diapause, which can also be obligate or facultative, is a state in which development of the embryo (J1) is completely arrested until the metabolic requirements are not satisfied, so that good environmental conditions are not sufficient to reverse the process [53].
Although the egg may hatch spontaneously, there are, however, factors that stimulate hatching. These may be environmental (temperature, soil moisture, oxygen availability, etc.) or derived from root exudates [54]. The effects of such factors on egg hatching vary between species of CNs [55]. The hatching of
Although the exit of J2 from the cyst constitutes the last stage of the hatching process, it is known that for this to occur, changes in the permeability of the membranes that cover the egg must take place. These changes are essential since they trigger the activation of second messengers that promote the following stages of development, such as the activation of metabolic pathways and J1 ecdysis [59]. Three membranes are known to surround the egg. The inner one is formed mainly by lipids and is semi-permeable to water and small ions; the intermediate, constituted by chitin microfilaments that give resistance and flexibility to the egg; and, finally, an outer one, is formed by lipoproteins (vitelline layer), which are believed to be essential for egg fertilization [60].
Unlike RKNs, where J2 activation can occur before changes in eggshell structure, in CNs, changes in eggshell membrane permeability are required for J2 activation. Within the egg, the movement of the juveniles is reduced by the low turgor pressure of the perivitelline fluid. It is mainly composed of the disaccharide trehalose (α-D-glucopyranosyl 1–1, α-D-glucopyranoside), which not only serves as an energy source for the nematode but also exerts a high osmotic pressure that reduces the water content of the perivitelline fluid [61]. Reduction of the trehalose content, and thus hydration (increased turgor pressure), is a prerequisite for egg hatching [60]. Although this is true for many species, there are others (i.e.,
Free in the soil, J2 is vulnerable to environmental conditions, so it must find a suitable root host plant before its lipid reserves are depleted. Under optimal conditions, localization time for
Once J2 reaches the root of a susceptible plant, it enters through the elongation areas of the root tip, as described for
In general, the process of syncytium formation is a highly complex process, which, as in RKNs, involves changes in gene expression in both the plant and the nematode. After 7 hours of ISC preparation, the sub ventral glands atrophy while the dorsal gland becomes more active [69]. The stylet is then removed from the ISC and reintroduced. It is at this point that the effector proteins contained in the dorsal gland are released into the cytoplasm of the ISCs [68]. As mentioned above, the secreted proteins have the function of establishing syncytium, which is facilitated by the dissolution of the cell wall separating adjacent cells from the ISC [70]. In
Syncytial cells, like GC in RKNs, undergo morphological changes. The nucleus enlarges by endoreduplication, and the central vacuole breaks down and gives rise to a large number of small vacuoles; on the other hand, the cytoplasm expands to accommodate a large number of ribosomes, mitochondria, endoplasmic reticulum, etc. [68]. In addition to these morphological changes, cells also experience metabolic changes that involve the overexpression of genes that are part of primary metabolism. These changes are influenced by effector proteins that modulate the transport and function of phytohormones. Thus, for example, chorismate mutase, an enzyme isolated from several PPNs and phytopathogenic fungi, is implicated in the initial pathogenesis of infection since it is known to alter the shikimic acid pathway, which is essential for plant-nematode interaction, since through it, precursors for auxins, salicylic acid, and a wide variety of phenolic compounds are generated [72, 73].
After the establishment of the feeding site, the nematode develops to the adult stage. The determination of males and females is influenced by environmental conditions and the supply of nutrients from the host plant. Females, unlike males, have a higher demand for nutrients, so their syncytia are larger than those formed by males. As adults, males, unlike females, are mobile and leave the root to find and fertilize a female. On the other hand, the female enlarges considerably to maintain the eggs (~600 inside the cyst and ~ 200 outside it) and thus restart the parasite’s life cycle, which has been estimated at 4 weeks for
3.3 Root-lesion nematodes (Pratylenchus spp.)
The RLNs are obligate biotrophic migratory endoparasites nematodes, without a permanent feeding site. There are more than 90 species registered with a cosmopolitan distribution. After RKNs and CNs, it is the third most important genus in agriculture. The species with the greatest agricultural economic impact are
As migratory endoparasites, they move intra- and inter-cellularly through the root tissues, generating superficial cracking of the root with internal rotting of the tubers that predispose to secondary infections by fungi and bacteria. Unlike the infections generated by RKNs and CNs, where the galls and cysts are observed with the naked eye on infested plants, in RLNs it is not easy to identify signs or symptoms that warn about their presence [74].
Its life cycle is relatively simple. This can last between 3 and 6 weeks, depending on the environmental conditions and the host. The eggs are deposited by the female either in the soil, the roots, or the tubers. Like RKNs and CNs, J2 hatches from the egg, which later develops through J3, J4, and the adult stages: male and female. All are vermiform (worm-like), infective, and motile. This ability to remain mobile during all phases of their cycle allows them to freely enter and exit the host plant [75]. Males commonly reproduce by parthenogenesis, and in some species, they are frequent, while in others they are not [76].
Like other PPNs, they are attracted to root elongation zones by different chemotactic molecules released by the host plant. They have a short, robust, hollow-mouthed stylet, which they use to break the plant cell wall by repeatedly pushing it. J2 and J3 tend to feed on the root hairs; however, they can also do from the epidermal, cortical, or stellar cells [77]. Prior to ingestion of the plant cell’s cytoplasmic contents, the dorsal glands are activated to secrete effector proteins that accumulate in the ampulla behind the buccal stylet. Feeding time can be short or last a few hours. In the latter case the cell may die, however, during the brief feeding, the stylet is removed, and the puncture site is sealed with no contents leaking out. Also, their intracellular displacement is another cause of cell death even when feeding occurs for short periods of time [78, 79].
Initial invasion is followed by the penetration of other nematodes, attracted by compounds released from damaged cells. Migration together with the feeding points generates brown lesions (necrotic areas,), which vary according to the type of host plant and parasite species. This roam feeding behavior leads to loss of plant growth and the appearance of leaf chlorosis and swelling and atrophy of the roots [80].
Although not all aspects of the biology of parasitism in RLNs are known, it is known that many of the effector proteins are different from those produced by RKNs and CNs. It is to be expected that, due to their migratory behavior and their intermittent feeding, the arsenal of effector proteins to manipulate the response of the host plant is less extensive than that of the sedentary endoparasites. Thus, for example, no orthologous sequences have been found for effector proteins essential for the establishment of infection in sedentary endoparasites such as 19C07 (auxin influx in syncytium], chorismate mutase, 10A06 (antioxidant genes in syncytium), CLE peptide, auxin, peptide hormones (CEPs), cytokinin, among others [74].
Transcriptomic studies in
Like other nematode species,
4. Management: a focus on new nematicides
4.1 Overview
The management and control of PPNs are of crucial importance in agriculture since many of them can endanger the food security of a country due to the loss of productive yield of the crops. After the USA and China, Brazil is the third country with the highest grain production in the world. According to the FAO [88], Brazil leads world soybean production, with 134 million tons produced in 2021, 10% more than that produced by the USA: ~120 million tons. It is estimated that 20% of soybean production in Brazil is affected by
Although crop rotation, genetic improvement, and biological control are some of the alternatives currently available for the management and control of PPNs (Figure 3), chemical control is the most widely used method worldwide [90, 91]. In developing countries, where access to agricultural technology is limited, and where the cost-benefit factor is of paramount importance for the economy, the use of agrochemicals is still the method of choice, due to their ease of implementation and availability [92, 93].

Figure 3.
Flow chart summarizing the different approaches that currently exist for the management and control of PPNs.
Almost half of the global nematicides market, worth US$ 1.3 billion, is used to control species of the genera
OPs and CMs owe their high toxicity to their low selectivity in their mechanism of action, since they are reversible inhibitors of the cholinesterase enzyme (AChE). This is because many are used as broad-spectrum pesticides (insecticides, acaricides, nematicides, and rodenticides) and as chemical weapons [99]. Aldicard, carbofuran, fenamiphos, and oxamyl are some of the OPs and CMs in which cholinesterase inhibition of PPNs has been demonstrated. Since the concentration of nematicide in agricultural soils is not high enough to kill nematodes, many of these compounds exert a nematostatic function with temporary paralysis that interferes with host feeding and egg hatching [100].
Although resistance has been reported under controlled laboratory conditions in PPNs, there are no proven examples in the scientific literature documenting dramatic changes in tolerance to nematicides that lead to suspicions about the development of resistance under field application conditions [100]. On the other hand, field resistance to agrochemicals has been widely demonstrated in different species of agricultural pests; however, it is believed that in PPNs the risk of developing resistance under field conditions is theoretically unlikely [101]. The arguments supporting this hypothesis are based on the application methods of nematicides and the ecological behaviors of PPNs. These characteristics reduce the effects of selective pressure to develop resistance, which can be overcome by the indiscriminate use of nematicides with different mechanisms of action [102].
Despite the fact that the global market for nematicides is much smaller than that of other agrochemicals such as insecticides (US$16.4 billion in 2019), fungicides (US$13.4 billion in 2019), and herbicides (US$32.6 billion in 2019), the fact remains that the market continues to grow despite strong regulatory rules limiting their use. Thus, for example, in 2011 dividends were approximately US$1 billion; however, by 2022 it was US$1.78 billion, an increase of approximately 43% [95, 100]. Post-modern control of PPNs requires cost-effective and selective targeted control strategies that meet the environmental safety demands of both growers and consumers. Thus, in the last 10 years, new nematicides have emerged (Figure 4) with low persistence in the environment and with selective mechanisms of action different from those of OPs and CMs. Of these new nematicides, only fluopyram and cyclobutrifluram have a currently known mechanism of action on PPNs [103, 104].

Figure 4.
Chemical structures of new nematicides.
4.2 Fluopyram and cyclobutrifluram
Marketed as an active ingredient in Verango®, Velum® and Indemnify®, fluopyram (N-[2-[3-chloro-5-(trifluoromethyl)pyridin-2-yl]ethyl]-2-(trifluoromethyl)benzamide) is a new generation nematicide-fungicide developed by Bayer Crop Science in 2012. Like fluopyram, cyclobutrifluram is a trifluoromethylpyridine developed and pantented by Syngenta in 2013 and that is marketed under the names Vaniva®, Tymirium®, and Victrato® [105]. Both interfere with the electron transport chain of the mitochondrial system (complex II) through inhibition of the enzyme succinate dehydrogenase (SDH). This mechanism has been reported for both PPNs and plant pathogenic fungi [106].
In eukaryotic organisms, SDH catalyzes the oxidation of succinate to fumarate as part of a mechanism to transfer electrons through the quinone pool. On the other hand, in nematodes and anaerobic helminths, complex II is associated with another enzyme that performs the reverse reaction (oxidation of reduced quinones) known as fumarate reductase (FRD). This is an adaptation to low oxygen pressures that prevent completely aerobic respiration. Such conditions are typical in intestinal nematodes; however, in free-living nematodes such as
SDH consists of four subunits (A-D) that differ in structure and function. Thus, for example, the SDHA sub-unit is the largest and is where the active site of the enzyme is located. On the other hand, the SDHB subunit contains iron-sulfur groups that mediate electron transfer to ubiquinone, which is located at the interface between the SDHB, SDHC, and SDHD subunits. It is there that many SDHIs bind to interfere with the catalytic activity of complex II [107]. Sequence alignments between sub-units of
Compared to fluensulfone, fluopyram works fast-acting. At a concentration of 2.0 mg/L, 100% of the J2 of
Unlike fluensulfone, abmectin, and aldicard, fluopyram has a poor ovicidal effect. At a concentration of 2.5 mg/L, they have a slight effect on the inhibition of the hatching of
4.3 Fluensulfone
Fluensulfone (5-chloro-2-(3,4,4-trifluorobut-3-enylsulfonyl)-1,3-thiazole) is the active ingredient of Nemitz®, a nematicide belonging to the group of fluroalkenyl sulfones, used for the control of PPNs in a wide variety of agricultural crops [119]. First registered in the USA in 2014 [120], it was developed by ADAMA and Control Solutions for Quali-Pro brand, in the past Makhteshim Agan Industries Ltd. It exists in three commercial formulations, one for turfgrass (Nimitz® Pro G) and two for horticulture: granules (Nimitz® 2% GR) and emulsion (Nimitz® and Nimitz® 480 EC).
Unlike OPs and CMs, fluensulfone is less toxic to humans and non-target organisms. It is moderately toxic after oral exposure, and has low acute toxicity following dermal application or inhalation in rabbits and rats [121]. The lethal dose via oral administration (LD50) in rats is 671 mg/kg body weight, which is five hundred times safer than the old nematicides such as aldicard (1 mg/kg of body weight) [122]. Despite the above, it has been reported to be toxic to aquatic organisms. Thus, for example, the EC50 for
Although there are few studies that demonstrate the impact of fluensulfone on free-living nematodes, it is known to be less toxic to
The immobilization in the form of a straight rod generated by fluensulfone when applied to J2 differs from that produced by OPs, suggesting a different mechanism of action than AChE inhibitors [128]. However, the exact mechanism of action on PPNs remains unknown. Some hypotheses suggest that fluensulfone possibly inhibits the enzyme acyl-CoA dehydrogenase, which is key in mobilizing lipids to obtain energy [129]; however, others suggest that its mechanism of action may be related to that of fluopyram [100, 103]. In insects, the difluoroalkenyl derivatives were found to inhibit β-oxidation of fatty acids in the mitochondria [130].
Compared to OPs and fluopyran, fluensulfone produces a slower but irreversible paralysis.
Like other nematicides, the susceptibility of PPNs to fluensulfone depends on the genus and species, even registering variations between the same species. In migrating nematodes such as
Currently, fluensulfone is approved for the control of different PPNs species (Meloidogyne, Pratylenchus, Hoplolaimus, Globodera, etc.) in a wide variety of crops such as tomato, potato, pepper, cabbage, squash, and strawberry, among others [100, 133]. Depending on the number of nematodes (population density per gram of soil), the type of crop rotation, and the presence of resistant plants, the average application rate is between 1.9 and 3.3 kg/ha [100, 129]. There are different forms of application; however, the most recommended are drip injection or broadcast by mechanical incorporation with a single application 7 days before planting [129]. It has been estimated that its half-life in soil is 36 days, with a dissipation rate (DT50) of between 20 and 50 days, which can vary depending on the type of soil and environmental conditions [134, 135].
4.4 Tioxazafen: seed nematicide
Registered in 2017 as a seed nematicide (NemaStrike™ ST), tioxazafen is a phenyloxadiazole (formally 3-phenyl-5-thiophen-2-yl-1,2,4-oxadiazole) developed by Monsanto Company, now Bayer CropScience, for the management and control of nematodes in cotton, corn, and soybean crops [136]. Unlike other nematicides, the development of tioxazafen involved the use of
Although its mechanism of action is not known with certainty, it is hypothesized that its nematicidal activity is due to ribosomal disruption. Polymorphism studies on
It is known through
Incorporated as an active ingredient in fungicide-insecticide formulations (Acceleron®), tioxazafen has low water solubility (0.0125 mg/mL) and reduced mobility in soil with an octanol/water partition coefficient (Log P) of 3.26. These characteristics make tioxazafen a moderately lipophilic compound that, according to estimates, has an ambient half-life of more than 100 days under aerobic conditions [138]. According to
In studies in growth chambers and in the field, it has been possible to evaluate the larvicidal and ovicidal power of tioxazafen, as well as its ability to inhibit reproduction and colonization of the host plant. The results suggest that tioxazafen exerts a direct nematicidal action on PPNs at doses below those required for OPs and CMs (50 mg/L) [136, 138]. The EC50 for M. incognita and
Trials in growth chambers and greenhouses show that tioxazafen is an excellent controller of several key nematodes when applied to seeds of soybean, corn, and cotton. In field trials with corn and soybean cultivars conducted by the USA EUP (Experimental Use Permit) in 40 locations that had nematode populations three times greater than those needed to cause economically significant damage, it was observed that tioxazafen increased the productive yield more efficiently than those observed for commercial use nematicides [137].
In Brazil, the nematicidal power of tioxazafen on
4.5 Fluazaindolizine: selective nematicide
Corteva Agriscience™, an agricultural division of the DowDuPont company (formerly DuPont Crop Protection), discovered and developed Salibro™, based on the new active substance fluazaindolizine (Reklemel™ Active). It is the first member of the new chemical class of N-phenylsulfonylimidazopyridine-2-carboxamide nematicides [148]. Fluazaindolizine, which was first registered with the EPA in July 2021, is a pesticide used to control PPNs on crops such as carrots, squash, tomatoes, eggplant, potatoes, and taro, as well as some fruits such as oranges, peaches, almonds, and grapes [149]. Their discovery began with high-throughput screening of an internal compound library against RKNs. After evaluating different 2,5-disubstituted arylsulfonamides, fluazaindolizine was selected due to its ecotoxicological and nematicidal profile [148]. It is suitable for a broad range of application techniques, including drip irrigation, bed spraying, and soil incorporation, among others [150].
Under
Greenhouse and field trials on tomato, strawberry, carrot, etc. plants show that fluazaindolizine is an effective nematicide for the treatment of
Although the selective effect of fluazaindolizine on non-target organisms is known, there is still toxicological information on the safety of this new nematicide on human and environmental health. The LD50 in rats via oral administration is 1187 mg/kg [100]. The main target organs in mammals are the urinary tract, hematopoietic tissue, and liver or gallbladder [161]. It is carcinogenic in high concentrations and has little impact on the reproductive system of rats and rabbits. Despite the possible toxicological effects of fluazaindolizine for mammals, it is known to be an easily photodegradable compound. The pH does not affect its degradation in soil; however, humic acid, iron (Fe-III), and nitrate (NO3−) ions negatively affect the photodegradation of fluazaindolizine. It has been proposed that the possible mechanism of photodegradation of fluazaindolizine involves imidazole-ring, dechlorination, hydroxyl substitution, ring-opening, cleavage, oxidation, and decarboxylation [162, 163].
5. Conclusion
Because of their cosmopolitan nature and high reproductive success, PPNs are a problem for medium- and large-scale agriculture. Although the leading countries in high-precision agriculture have the technical and economic resources to deal with this problem, this is not the case in developing countries, where traditional pest control methods are insufficient to adequately manage PPNs. Understanding the molecular basis of the plant-nematode interaction is essential for proper control, as this will allow the development of more rational and specific methods for PPN control. Omics and
Acknowledgments
The author Acknowledgments Chemaxon Ltd. for the license granted for the use of the MarvinSketch software (v.22.18) with which the chemical structures of new nematicides were designed.
References
- 1.
UNDESA. Department of Economic and Social Affairs. World Population Prospects 2022: Summary of Results Ten Key Messages [Internet]. 2022. Available from: https://www.un.org/development/desa/pd/sites/www.un.org.development.desa.pd/files/undesa_pd_2022_wpp_key-messages.pdf [Accessed: June 01, 2023] - 2.
Andree BPJ, Chamorro A, Kraay A, Spencer P, Wang D. Predicting Food Crises. World Bank, Washington, DC; 2020. Available from: http://hdl.handle.net/10986/34510 [Accessed: 2023-06-01] - 3.
FAO. Thinking about the Future of Food Safety – A Foresight Report [Internet]. Rome, Italy. 2022. Available from: 10.4060/cb8667en [Accessed: 2021-06-01] - 4.
Ireland KB, van Klinken R, Cook DC, Logan D, Jamieson L, Tyson JL, et al. Plant pest impact metric system (PPIMS): Framework and guidelines for a common set of metrics to classify and prioritise plant pests. Crop Protection. 2020; 128 :105003. DOI: 10.1016/j.cropro.2019.105003 - 5.
FAO. Climate Change Fans Spread of Pests and Threatens Plants and Crops, New FAO Study [Internet]. 2021. Available from: https://www.fao.org/news/story/en/item/1402920/icode/ [Accessed: June 01, 2021] - 6.
Lindsey APJ, Murugan S, Renitta RE. Microbial disease management in agriculture: Current status and future prospects. Biocatalysis Agricultural Biotechnology. 2020; 23 :1-12. DOI: 10.1016/j.bcab.2019.101468 - 7.
Patil BL. Plant viral diseases: Economic implications. In: Bamford DH, Zuckerman M, editors. Encyclopedia of Virology. 4nd ed. USA: Academic Press; 2020. pp. 81-97. DOI: 10.1016/B978-0-12-809633-8.21307-1 - 8.
Pan L, Li X-Z, Sun D-A, Jin H, Guo H-R, Qin B. Design and synthesis of novel coumarin analogs and their nematicidal activity against five phytonematodes. Chinese Chemical Letters. 2016; 27 :375-376. DOI: 10.1016/j.cclet.2016.01.029 - 9.
Singh S, Singh B, Singh AP. Nematodes: A threat to sustainability of agriculture. Procedia Environmental Sciences. 2015; 29 :215-216. DOI: 10.1016/j.proenv.2015.07.270 - 10.
Karuri H. Root and soil health management approaches for control of plant-parasitic nematodes in sub-Saharan Africa. Crop Protection. 2022; 152 :1-9. DOI: 10.1016/j.cropro.2021.105841 - 11.
Abd-Elgawad MM, Askary TH. Impact of phytonematodes on agriculture economy. In: Askary TH, Martinelli PR, editors. Biocontrol Agents of Phytonematodes. 1st ed. Washington: CABI; 2015. pp. 3-49. DOI: 10.1079/9781780643755.0003 - 12.
Colussi J, Schnitkey G. New Soybean Record: Historical growing of production in Brazil [Internet]. 2021. Available from: https://farmdocdaily.illinois.edu/wpcontent/uploads/2021/03/fdd300321.pdf [Accessed: June 01, 2021] - 13.
Moura F, da Silva EH, Silva Antolin LA, Zanon AJ, Soares Andrade A, Antunes de Souza H, et al. Impact assessment of soybean yield and water productivity in Brazil due to climate change. European Journal of Agronomy. 2021; 129 :1-11. DOI: 10.1016/j.eja.2021.126329 - 14.
Tylka GL, Marett CC. Known distribution of the soybean cyst nematode, Heterodera glycines , in the United States and Canada in 2020. Plant Health Progress. 2021;22 :72-74. DOI: 10.1094/PHP-10-20-0094-BR - 15.
Ganguly RK, Mukherjee A, Chakraborty SK, Verma JP. Impact of agrochemical application in sustainable agriculture. In: Verma JP, Macdonald CA, Gupta VK, Podile AR, editors. New and Future Developments in Microbial Biotechnology and Bioengineering. 1st ed. Uttar Pradesh: Elsevier B.V; 2021. pp. 15-24. DOI: 10.1016/B978-0-444-64325-4.00002-X - 16.
Khanna K, Kohli SK, Ohri P, Bhardwaj R. Plants-nematodes-microbes crosstalk within soil: A trade-off among friends or foes. Microbiological Research. 2021; 248 :1-23. DOI: 10.1016/j.micres.2021.126755 - 17.
Ahuja A, Somvanshi VS. Diagnosis of plant-parasitic nematodes using loop-mediated isothermal amplification (LAMP): A review. Crop Protection. 2020; 147 :1-10. DOI: 10.1016/j.cropro.2020.105459 - 18.
Ali MA, Azeem F, Li H, Bohlmann H. Smart parasitic nematodes use multifaceted strategies to parasitize plants. Frontiers in Plant Science. 2017; 8 :1699. DOI: 10.3389/fpls.2017.01699 - 19.
Hammond-Kosack KE, Jones JDG. Responses to plant pathogens. In: Buchanan B, Gruissem W, Jones RL, editors. Biochemistry and Molecular Biology of Plants. 2nd ed. Chichester: John Wiley & Sons; 2015. pp. 984-1049 - 20.
Ibrahim HMM, Ahmad EM, Martínez-Medina A, Aly MAM. Effective approaches to study the plant-root knot nematode interaction. Plant Physiology and Biochemistry. 2019; 141 :332-342. DOI: 10.1016/j.plaphy.2019.06.009 - 21.
Mateille T, Tavoillot J, Goillon C, Pares L, Lefèvre A, Védie H, et al. Competitive interactions in plant-parasitic nematode communities affecting organic vegetable cropping systems. Crop Protection. 2020; 135 (105206):105206. DOI: 10.1016/j.cropro.2020.105206 - 22.
Jones JT, Haegeman A, Danchin EGJ, Gaur HS, Helder J, Jones MGK, et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Molecular Plant Pathology. 2013; 14 (9):946-961. DOI: 10.1111/mpp.12057 - 23.
Verdejo-Lucas S, Talavera M. Root-knot nematodes on zucchini ( Cucurbita pepo subsp.pepo ): Pathogenicity and management. Crop Protection. 2019;126 :104943. DOI: 10.1016/j.cropro.2019.104943 - 24.
Orion D, Kritzman G, Meyer SLF, Erbe EF, Chitwood DJ. A role of the gelatinous matrix in the resistance of root-knot nematode ( Meloidogyne spp.) eggs to microorganisms. Journal of Nematology. 2001;33 (4):203-207. DOI: PMC2620502 - 25.
Karssen G, Wesemael W, Moens M. Root-knot nematodes. In: Perry RN, Moens M, editors. Plant Nematology. 2nd ed. London UK: CAB International; 2013. pp. 73-83 - 26.
Perry RN, Wright DJ, Chitwood DJ. Reproduction, physiology and biochemistry. In: Perry RN, Moens M, editors. Plant Nematology. 2nd ed. London UK: CAB International; 2013. pp. 235-242 - 27.
Pline M, Dusenbery DB. Responses of plant-parasitic nematode Meloidogyne incognita to carbon dioxide determined by video camera-computer tracking. Journal of Chemical Ecology. 1987; 13 (4):873-888. DOI: 10.1007/BF01020167 - 28.
Rasmann S, Hiltpold I, Ali J. The role of root-produced volatile secondary metabolites in mediating soil interactions. In: Montanaro G, Dichio B, editors. Advances in Selected Plant Physiology Aspects. 1st ed. Rijeka, Croatia: IntechOpen; 2012. pp. 270-281. DOI: 10.5772/34304 - 29.
Ali JG, Alborn HT, Stelinski LL. Constitutive and induced subterranean plant volatiles attract both entomopathogenic and plant parasitic nematodes. Journal of Ecology. 2010; 99 (1):26-35. DOI: 10.1111/j.1365-2745.2010.01758.x - 30.
Čepulyte R, Danquah WB, Bruening G, Williamson VM. Potent attractant for root-knot nematodes in exudates from seedling root tips of two host species. Scientific Reports. 2018; 8 (1):1-10. DOI: 10.1038/s41598-018-291 - 31.
Dutta TK, Ganguly AK, Gaur HS. Global status of rice root-knot nematode, Meloidogyne graminicola . African Journal of Microbiology Research. 2012;6 (32):6016-6021. DOI: 10.5897/AJMR12.707 - 32.
Tapia-Vázquez I, Montoya-Martínez AC, De los Santos-Villalobos S, Ek-Ramos MJ, Montesinos-Matías R, Martínez-Anaya C. Root-knot nematodes ( Meloidogyne spp.) a threat to agriculture in Mexico: Biology, current control strategies, and perspectives. World Journal of Microbiology and Biotechnology. 2022;38 (2):1-18. DOI: 10.1007/s11274-021-03211-2 - 33.
Eves-van den Akker S. Plant–nematode interactions. Current Opinion in Plant Biology. 2021; 62 :102035. DOI: 10.1016/j.pbi.2021.102035 - 34.
Khan M, Khan AU. Plant parasitic nematodes effectors and their crosstalk with defense response of host plants: A battle underground. Rhizosphere. 2021; 17 :100288. DOI: 10.1016/j.rhisph.2020.100288 - 35.
Truong NM, Nguyen CN, Abad P, Quentin M, Favery B. Function of root-knot nematode effectors and their targets in plant parasitism. In: Escobar C, Fenoll C, editors. Advances in Botanical Research. Vol. 73. Toledo: Elsevier Ltd; 2015. pp. 294-319 - 36.
Escobar C, Barcala M, Cabrera J, Fenoll C. Overview of root-knot nematodes and giant cells. In: Escobar C, Fenoll C, editors. Advances in Botanical Research. Vol. 73. Toledo: Elsevier Ltd; 2015. pp. 5-24 - 37.
Jones MGK, Goto DB. Root-knot nematodes and Giant cells. In: Jones J, Gheysen G, Fenoll C, editors. Genomic and Molecular Genetics of Plant-Nematode Interactions. 1st ed. Dordrecht, Netherlands: Springer Science+Business Media; 2011. pp. 83-99 - 38.
Gheysen G, Mitchum MG. Phytoparasitic nematode control of plant hormone pathways. Plant Physiology. 2019; 179 (4):1212-1226. DOI: 10.1104/pp.18.01067 - 39.
Kyndt T, Goverse A, Haegeman A, Warmerdam S, Wanjau C, Jahani M, et al. Redirection of auxin flow in Arabidopsis thaliana roots after infection by root-knot nematodes. Journal of Experimental Botany. 2016;67 (15):4559-4570. DOI: 10.1093/jxb/erw230 - 40.
Yuan J, Bateman P, Gutierrez-Marcos J. Genetic and epigenetic control of transfer cell development in plants. Journal of Genetics and Genomics. 2016; 43 (9):533-539. DOI: 10.1016/j.jgg.2016.08.002 - 41.
Dowd CD, Chronis D, Radakovic ZS, Siddique S, Schmülling T, Werner T, et al. Divergent expression of cytokinin biosynthesis, signaling and catabolism genes underlying differences in feeding sites induced by cyst and root-knot nematodes. The Plant Journal. 2017; 92 (2):211-228. DOI: 10.1111/tpj.13647 - 42.
Leonetti P, Zonno MC, Molinari S, Altomare C. Induction of SA-signaling pathway and ethylene biosynthesis in Trichoderma harzianum -treated tomato plants after infection of the root-knot nematodeMeloidogyne incognita . Plant Cell Reports. 2017;36 (4):621-631. DOI: 10.1007/s00299-017-2109-0 - 43.
Nahar K, Kyndt T, De Vleesschauwer D, Höfte M, Gheysen G. The Jasmonate pathway is a key player in systemically induced defense against root knot nematodes in rice. Plant Physiology. 2011; 157 (1):305-316. DOI: 10.1104/pp.111.177576 - 44.
Jaouannet M, Magliano M, Arguel MJ, Gourgues M, Evangelisti E, Abad P, et al. The root-knot nematode calreticulin mi-CRT is a key effector in plant defense suppression. Molecular Plant-Microbe Interactions. 2013; 26 (1):97-105. DOI: 10.1094/MPMI-05-12-0130-R - 45.
Przybylska A, Obrępalska-Stęplowska A. Plant defense responses in monocotyledonous and dicotyledonous host plants during root-knot nematode infection. Plant and Soil. 2020; 451 :239-260. DOI: 10.1007/s11104-020-04533-0 - 46.
Molinari S, Fanelli E, Leonetti P. Expression of tomato salicylic acid (SA)-responsive pathogenesis-related genes in Mi-1-mediated and SA-induced resistance to root-knot nematodes. Molecular Plant Pathology. 2013; 15 (3):255-264. DOI: 10.1111/mpp.12085 - 47.
Manzanilla-Lopez RH, Evans K, Bridge J. Plant diseases caused by nematode. In: Chen Z, Chen S, Dickson DW, editors. Nematology - Advances and Perspectives. Vol. 2nd. Wallingford, UK: CABI Publishing; 2004. pp. 636-716 - 48.
Castagnone-Sereno P, Danchin EGJ, Perfus-Barbeoch L, Abad P. Diversity and evolution of root-knot nematodes, genus Meloidogyne : New insights from the genomic era. Annual Review of Phytopathology. 2013;51 (1):203-220. DOI: 10.1146/annurev-phyto-082712-102300 - 49.
Liu QL, Thomas VP, Williamson VM. Meiotic parthenogenesis in a root-knot nematode results in rapid genomic homozygosity. Genetics. 2007; 176 (3):1483-1490. DOI: 10.1534/genetics.107.071134 - 50.
Chitwood DJ, Perry RN. Reproduction, physiology and biochemistry. In: Perry NR, Moens M, Starr JL, editors. Roo-Knot Nematodes. 1st ed. London, UK: CABI Publishing; 2009. pp. 182-194 - 51.
Baldwin JG, Handoo ZA. General morphology of cyst nematodes. In: Perry RN, Moens M, Jones JT, editors. Cyst Nematodes. 1st ed. London, UK: CABI Publishing; 2018. pp. 337-359 - 52.
Smiley RW, Dababat AA, Iqbal S, Jones MGK, Maafi ZT, Peng D, et al. Cereal cyst nematodes: A complex and destructive group of heterodera species. Plant Disease. 2017; 101 (10):1692-1720. DOI: 10.1094/PDIS-03-17-0355-FE - 53.
Palomares-Rius JE, Hedley P, Cock PJA, Morris JA, Jones JT, Blok VC. Gene expression changes in diapause or quiescent potato cyst nematode, Globodera pallida , eggs after hydration or exposure to tomato root diffusate. Peer J. 2016;4 :e1654. DOI: 10.7717/peerj.1654 - 54.
Shimizu K, Kushida A, Akiyama R, Lee HJ, Okamura Y, Masuda Y, et al. Hatching stimulation activity of steroidal glycoalkaloids toward the potato cyst nematode, Globodera rostochiensis . Plant Biotechnology. 2020;37 (3):319-325. DOI: 10.5511/plantbiotechnology.20.0516a - 55.
Ngala B, Mariette N, Ianszen M, Dewaegeneire P, Denis MC, Porte C, et al. Hatching induction of cyst nematodes in bare soils drenched with root exudates under controlled conditions. Frontiers in Plant Science. 2021; 11 :602825. DOI: 10.3389/fpls.2020.602825 - 56.
Shimizu K, Akiyama R, Okamura Y, Ogawa C, Masuda Y, Sakata I, et al. Solanoeclepin B, a hatching factor for potato cyst nematode. Science Advances. 2023; 9 (11):1-11. DOI: 10.1126/sciadv.adf4166 - 57.
Nonaka S, Katsuyama T, Kondo T, Sasaki Y, Asami T, Yajima S, et al. 1,10-phenanthroline and its derivatives are novel hatching stimulants for soybean cyst nematodes. Bioorganic and Medicinal Chemical Letters. 2016; 1 :5240-5243. DOI: 10.1016/j.bmcl.2016.09.052 - 58.
Perry RN. Plant signal in nematode hatching and attraction. In: Fenoll C, Grundler FMW, Ohl SA, editors. Cellular and Molecular Aspects of Plant-Nematode Interactions. Vol. 10. Dordrecht, Netherlands: Kluwer Academic Publishers; 1997. pp. 38-46 - 59.
Perry RN. Hatching. In: Lee DL, editor. The Biology of Nematodes. 1st ed. London, UK: Taylor & Francis Group; 2002. pp. 294-335 - 60.
Bohlmann H. Introductory chapter on the basic biology of cyst nematodes. In: Escobar C, Fenoll C, editors. Advances in Botanical Research. Vol. 73. Toledo: Elsevier Ltd; 2015. pp. 2-21. DOI: 10.1016/bs.abr.2014.12.001 - 61.
Masler EP, Perry RN. Hatch, survival and sensory perception. In: Perry RN, Moens M, Jones JT, editors. Cyst nematodes. 1st ed. London, UK: CABI International; 2018. pp. 44-73 - 62.
Perry RN, Clarke A, Hennessy J. The influence of osmotic pressure on the hatching of Heterodera schachtii. Revue Nematol. 1980; 3 (1):3-9 - 63.
Li T, Tang BZ, Li B, Mobbs D. The hatching of cereal cyst nematode ( Heterodera avenae ) in response to different inorganic ions. Scientia Agricola. 2022;79 (1):1-7. DOI: 10.1590/1678-992X - 64.
Turner SJ, Subbotin SA. Cyst nematodes. In: Perry RN, Moens M, editors. Plant nematology. 2nd ed. London, UK: CABI International; 2013. pp. 109-141 - 65.
Price JA, Coyne D, Blok VC, Jones JT. Potato cyst nematodes Globodera rostochiensis andG. pallida . Molecular Plant Pathology. 2021;22 (5):495-507. DOI: 10.1111/mpp.13047 - 66.
Curtis RHC. Plant-nematode interactions: Environmental signals detected by the nematode’s chemosensory organs control changes in the surface cuticle and behaviour. Parasite. 2008; 15 (3):310-316. DOI: 10.1051/parasite/2008153310 - 67.
Mende NV, Nobre MJ, Perry RN. Host finding, invasion and feeding. In: Sharma SB, editor. The Cyst Nematodes. 1st ed. Dordrecht, Netherlands: Springer Science+Business Media; 1998. pp. 217-238 - 68.
Sobczak M, Golinowski W. Cyst nematodes and syncytia. In: Jones J, Gheysen G, Fenoll C, editors. Genomic and Molecular Genetics of Plant-Nematode Interactions. 1st ed. Dordrecht, Netherlands: Springer Science+Business Media; 2011. pp. 61-80 - 69.
Wyss U. Observations on the feeding behaviour of Heterodera schachtii throughout development, including events during moulting. Fundamental and Applied Nematology. 1992;15 :75-89 - 70.
Subbotin SA, Mundo-Ocampo M, Baldwin JG. Biology and evolution. In: Subbotin SA, Mundo-Ocampo M, Baldwin JG, editors. Systematics of Cyst Nematodes (Nematoda: Heteroderinae), Part a. 1st ed. Leiden, Netherlands: BRILL; 2010. pp. 43-70 - 71.
Sobczak M, Golinowski W. Structure of cyst nematode feeding sites. In: Berg RH, Taylor CG, editors. Cell Biology of Plant Nematodes Parasitism. Vol. 15. Heidelberg, Germany: Springer-Verlag; 2009. pp. 153-187 - 72.
Mitchum MG, Hussey RS, Baum TJ, Wang X, Elling AA, Wubben M, et al. Nematode effector proteins: An emerging paradigm of parasitism. New Phytologist. 2013; 199 (4):879-894. DOI: 10.1111/nph.12323 - 73.
Karczmarek A, Overmars H, Helder J, Goverse A. Feeding cell development by cyst and root-knot nematodes involves a similar early, local and transient activation of a specific auxin-inducible promoter element. Molecular Plant Pathology. 2004; 5 (4):343-346. DOI: 10.1111/j.1364-3703.2004.00230.x - 74.
Jones MGK, Fosu-Nyarko J. Molecular biology of root lesion nematodes ( Pratylenchus spp.) and their interaction with host plants. Annals of Applied Biology. 2014;164 (2):163-181. DOI: 10.1111/aab.12105 - 75.
Bucki P, Qing X, Castillo P, Gamliel A, Dobrinin S, Alon T, et al. The genus Pratylenchus (Nematoda: Pratylenchidae) in Israel: From taxonomy to control practices. Plants. 2020;9 (11):1475. DOI: 10.3390/plants9111475 - 76.
Mokrini F, Viaene N, Waeyenberge L, Dababat AA, Moens M. Root-lesion nematodes in cereal fields: Importance, distribution, identification, and management strategies. Journal of Plant Diseases and Protection. 2018; 126 (1):1-11. DOI: 10.1007/s41348-018-0195-z - 77.
Orlando V, Grove IG, Edwards SG, Prior T, Roberts D, Neilson R, et al. Root-lesion nematodes of potato: Current status of diagnostics, pathogenicity and management. Plant Pathology. 2020; 69 (3):405-417. DOI: 10.1111/ppa.13144 - 78.
Zunke U. Ectoparasitic feeding behaviour of the root lesion nematode, Pratylenchus penetrans , on root hairs of different host plants. Revue Nématologie. 1990;13 (3):331-337. DOI: 10670/1.u6a71m - 79.
Ducan LW, Moens M. Migratory endoparasitic nematodes. In: Perry RN, Moens M, editors. Plant nematology. 2nd ed. London UK: CAB International; 2013. pp. 145-178 - 80.
Fosu-Nyarko J, Jones MP. Advances in understanding the molecular mechanisms of root lesion nematode host interactions. Annual Review of Phytopathology. 2016; 54 (1):253-278. DOI: 10.1146/annurev-phyto-080615-100257 - 81.
Vieira PC, Shao J, Vijayapalani P, Maier T, Pellegrin C, Eves-Van S, et al. A new esophageal gland transcriptome reveals signatures of large scale de novo effector birth in the root lesion nematode Pratylenchus penetrans . BMC Genomics. 2020;21 (1):1-16. DOI: 10.1186/s12864-020-07146-0 - 82.
Devi AN, Ponnuswami V, Sundararaju P, Soorianathasundaram K, Sathiamoorthy S, Uma S, et al. Mechanism of resistance in banana cultivars against root lesion nematode, Pratylenchus coffeae . Indian Journal of Nematology. 2007;37 :138-144 - 83.
Perry RN. Desiccation survival of parasitic nematodes. Parasitology. 1999; 119 :S19-S30. DOI: 10.1017/S0031182000084626 - 84.
Thompson JP, Rostad HE, Whish JPM. Survival of root-lesion nematodes ( Pratylenchus thornei ) after wheat growth in a vertisol is influenced by rate of progressive soil desiccation. Annals of Applied Biology. 2016;170 (1):78-88. DOI: 10.1111/aab.12316 - 85.
Ribeiro LM, Campos HD, Neves DL, Dias-Arieira CR. Survival of Pratylenchus brachyurus under dry soil conditions. Heliyon. 2020;6 (9):1-6. DOI: 10.1016/j.heliyon.2020.e05075 - 86.
Townshend JL. Anhydrobiosis in Pratylenchus penetrans . Journal of Nematology. 1984;16 (3):282-289. DOI: PMC2618384/?tool=EBI - 87.
Storey RMJ, Glazer I, Orion D. Lipid utilisation by starved and anhydrobiotic individuals of Pratylenchus thornei . Nematologica. 1982;28 (4):373-378. DOI: 10.1163/187529282X00178 - 88.
FAO. FAOSTAT [Internet]. www.fao.org . 2022. Available from:https://www.fao.org/faostat/en/#data/QCL - 89.
Favoreto L, Meyer MC, Dias-Arieira CR, Machado AC, Santiago DC, Ribeiro N. Diagnose e manejo de fitonematoides na cultura da soja. Informe Agropecuario. 2019; 306 :18-29 - 90.
Pires D, Vicente CS, Menéndez E, Faria JMS, Rusinque L, Camacho MJ, et al. The fight against plant-parasitic nematodes: Current status of bacterial and fungal biocontrol agents. Pathogens. 2022; 11 (10):1-22. DOI: 10.3390/pathogens11101178 - 91.
Rajak P, Roy S, Ganguly A, Mandi M, Dutta A, Das K, et al. Agricultural pesticides – Friends or foes to biosphere? Journal of Hazardous Materials Advances. 2023; 10 :100264. DOI: 10.1016/j.hazadv.2023.100264 - 92.
Zasada IA, Halbrendt JM, Kokalis-Burelle N, LaMondia J, McKenry MV, Noling JW. Managing nematodes without methyl bromide. Annual Review of Phytopathology. 2010; 48 (1):311-328. DOI: 10.1146/annurev-phyto-073009-114425 - 93.
Sharma A, Kumar V, Shahzad B, Tanveer M, Sidhu GPS, Handa N, et al. Worldwide pesticide usage and its impacts on ecosystem. SN Applied Sciences. 2019; 1 (11):11446. DOI: g/10.1007/s42452-019-1485-1 - 94.
Wram CL, Zasada IA. Short-term effects of sublethal doses of nematicides on Meloidogyne incognita . Phytopathology. 2019;109 (9):1605-1613. DOI: 10.1094/PHYTO-11-18-0420-R - 95.
Spherical Insights LLP. Global Nematicides Market Size to Worth USD 4.28 Billion By 2032 | CAGR of 9.1% [Internet]. Available from: https://www.sphericalinsights.com/request-sample/1768 . [Accessed: May 21, 2023] - 96.
Taylor RWD. Methyl bromide—Is there any future for this noteworthy fumigant? Journal of Stored Products Research. 1994; 30 (4):253-260. DOI: 10.1016/S0022-474X(94)90317-4 - 97.
Sharma A, Shukla A, Attri K, Kumar M, Kumar P, Suttee A, et al. Global trends in pesticides: A looming threat and viable alternatives. Ecotoxicology and Environmental Safety. 2020; 201 :110812. DOI: 10.1016/j.ecoenv.2020.110812 - 98.
Costa LG, Giordano G, Guizzetti M, Vitalone A. Neurotoxicity of pesticides: A brief review. Frontiers in Bioscience. 2008; 13 (13):1240. DOI: 10.2741/2758 - 99.
Gupta RC, Sachana M, Mukherjee IM, Doss RB, Malik JK, Milatovic D. Organophosphates and carbamates. In: Gupta RC, editor. Veterinary Toxicology. 3nd ed. London: Academic Press; 2018. pp. 495-507. DOI: 10.1016/B978-0-12-811410-0.00037-4 - 100.
Oka Y. From old-generation to next-generation nematicides. Agronomy. 2020; 10 (9):1387. DOI: 10.3390/agronomy10091387 - 101.
Sparks TC, Crossthwaite AJ, Nauen R, Banba S, Cordova D, Earley F, et al. Insecticides, biologics and nematicides: Updates to IRAC’s mode of action classification - a tool for resistance management. Pesticide Biochemistry and Physiology. 2020; 167 :104587. DOI: 10.1016/j.pestbp.2020.104587 - 102.
Grange R, Mandiriza G, Zyl C. Nematodes, nematicides and resistance management. In: Insecticide Resistence Action Committee. 2021; South Africa. p. 1-23. Available from: https://irac-online.org/documents/nematicides-and-resistance-management/ [Accessed:2023-05-21] - 103.
Faske TR, Hurd K. Sensitivity of Meloidogyne incognita and Rotylenchulus reniformis to Fluopyram. Journal of Nematology. 2015; 47 (4):316-321 - 104.
Desaeger J, Wram C, Zasada I. New reduced-risk agricultural nematicides - rationale and review. Journal of Nematology. 2020; 52 (1):1-16. DOI: 10.21307/jofnem-2020-091 - 105.
Xue Z, Zhong S, Shen J, Sun Y, Gao X, Wang X, et al. Multiple mutations in SDHB and SDHC2 subunits confer resistance to the succinate dehydrogenase inhibitor cyclobutrifluram in Fusarium fujikuroi . Journal of Agricultural and Food Chemistry. 2023;71 (8):3694-3704. DOI: 10.1021/acs.jafc.2c08023 - 106.
Liang P, Shen S, Xu Q , Zhang L, Jin S, Lu H, et al. Design, synthesis biological activity, and docking of novel fluopyram derivatives containing guanidine group. Bioorganic and Medical Chemistry. 2021; 29 :115846-115846. DOI: 10.1016/j.bmc.2020.115846 - 107.
Harada S, Inaoka DK, Ohmori J, Kita K. Diversity of parasite complex II. Biochimica et Biophysica Acta (BBA) – Bioenergetics. 2013; 1827 (5):658-667. DOI: 10.1016/j.bbabio.2013.01.005 - 108.
Kuramochi T, Hirawake H, Kojima S, Takamiya S, Furushima R, Aoki T, et al. Sequence comparison between the flavoprotein subunit of the fumarate reductase (complex II) of the anaerobic parasitic nematode, Ascaris suum and the succinate dehydrogenase of the aerobic, free-living nematode,Caenorhabditis elegans . Molecular and Biochemical Parasitology. 1994;68 (2):177-187. DOI: 10.1016/0166-6851(94)90163-5 - 109.
Karavaeva V, Sousa FL. Modular structure of complex II: An evolutionary perspective. Biochimica et Biophysica Acta (BBA) - Bioenergetics. 2022; 1864 :1-11. DOI: 10.1016/j.bbabio.2022.148916 - 110.
Lümmen P, Fürsch H. Fluopyram a novel nematicide for the control of root-knot nematodes. In: Jeschke P, Witschel M, Krämer W, Schirmer U, editors. Modern Crop Protection Compounds. 3nd ed. Weinheim: Wiley-VCH Verlag GmbH & Co. KGaA; 2019. pp. 1630-1640 - 111.
Burns AR, Luciani GM, Musso G, Bagg R, Yeo M, Zhang Y, et al. Caenorhabditis elegans is a useful model for anthelmintic discovery. Nature Communications. 2015;6 (1):1-11. DOI: 10.1038/ncomms8485 - 112.
Oka Y, Saroya Y. Effect of fluensulfone and fluopyram on the mobility and infection of second-stage juveniles of Meloidogyne incognita andM. javanica . Pest Management Science. 2019;75 (8):2095-2106. DOI: 10.1002/ps.5399 - 113.
Schleker ASS, Rist M, Matera C, Damijonaitis A, Collienne U, Matsuoka K, et al. Mode of action of fluopyram in plant-parasitic nematodes. Scientific Reports. 2022; 12 (1):1-14. DOI: 10.1038/s41598-022-15782-7 - 114.
Storelli A, Keiser A, Eder R, Jenni S, Kiewnick S. Evaluation of fluopyram for the control of Ditylenchus dipsaci in sugar beet. Journal of Nematology. 2020;52 (1):1-10. DOI: 10.21307/jofnem-2020-071 - 115.
Feist E, Kearn J, Gaihre Y, O’Connor V, Holden-Dye L. The distinct profiles of the inhibitory effects of fluensulfone, abamectin, aldicarb and fluopyram on Globodera pallida hatching. Pesticide Biochemistry and Physiology. 2020;165 :104541. DOI: 10.1016/j.pestbp.2020.02.007 - 116.
Hawk T, Faske TR. Effect of seed-applied fluopyram on Meloidogyne incognita infection and maturity in cotton and soybean. Journal of Nematology. 2021;52 :1-7. DOI: 10.21307/jofnem-2020-083 - 117.
Grabau ZJ, Liu C, Schumacher LA, Small IM, Wright DL. In-furrow fluopyram nematicide efficacy for Rotylenchulus reniformis management in cotton production. Crop Protection. 2021;140 :105423. DOI: 10.1016/j.cropro.2020.105423 - 118.
Waisen P, Wang KH, Uyeda J, Myers RY. Effects of fluopyram and azadirachtin integration with sunn hemp on nematode communities in zucchini, tomato and sweet potato in Hawaii. Journal of Nematology. 2021; 53 (1):1-15. DOI: 10.21307/jofnem-2021-030 - 119.
Jeanmart S, Edmunds A, Lamberth C, Pouliot M. Synthetic approaches to the 2010-2014 new agrochemicals. Bioorganic and Medical Chemistry. 2016; 24 (3):317-341. DOI: 10.1016/j.bmc.2015.12.014 - 120.
USEPA. Pesticide Product Label, FLUENSULFONE 480EC [Internet]. 2014. Available from: https://www3.epa.gov/pesticides/chem_search/ppls/066222-00243-20140911.pdf [Accessed: June 01, 2023] - 121.
Dewhurst I, Tasheva M. Fluensulfone. In: WHO. Pesticide Residues in Food – 2013: Toxicological Evaluations. 1st ed. Geneva: World Health Organization; 2015. pp. 271-309 - 122.
Proença P, Teixeira H, De Mendonça MC, Castanheira F, Marques EP, Corte-Real F, et al. Aldicarb poisoning: One case report. Forensic Science International. 2004; 146 :S79-S81. DOI: 10.1016/j.forsciint.2004.09.028 - 123.
APVMA. Public Release Summery on the Evaluation of the New Active Fluensulfone in the Product Nimitz 480 EC Nematicide; Australian Pesticides and Veterinary Medicines Authority: Kingstone, Australia, 2015; p. 47. Available from: http://apvma.gov.au/node/15596 [Accessed: 2023-06-01] - 124.
Giannakou IO, Panopoulou S. The use of fluensulfone for the control of root-knot nematodes in greenhouse cultivated crops: Efficacy and phytotoxicity effects. Cogent Food & Agriculture. 2019; 5 (1):1643819. DOI: 10.1080/23311932.2019.1643819 - 125.
Kearn J, Ludlow E, Dillon J, O’Connor V, Holden-Dye L. Fluensulfone is a nematicide with a mode of action distinct from anticholinesterases and macrocyclic lactones. Pesticide Biochemistry and Physiology. 2014; 109 :44-57. DOI: 10.1016/j.pestbp.2014.01.004 - 126.
Kawanobe T, Fujita H. Evaluation of Nematicidal activity of Fluensulfone against non-target free-living nematodes under field conditions. Agronomy. 2019; 9 (12):853. DOI: 10.3390/agronomy9120853 - 127.
Waldo BD, Grabau ZJ, Mengistu TM, Crow WT. Nematicide effects on non-target nematodes in bermudagrass. Journal of Nematology. 2019; 51 :e2019-e2009. DOI: 10.21307/jofnem-2019-009 - 128.
Oka Y, Shuker S, Tkachi N. Nematicidal efficacy of MCW-2, a new nematicide of the fluoroalkenyl group, against the root-knot nematode Meloidogyne javanica . Pest Management Science. 2009;65 (10):1082-1089. DOI: 10.1002/ps.1796 - 129.
Maienfisch P, Loiseleur O, Slaats B. Recent nematicides. In: Jeschke P, Witschel M, Krämer W, Schirmer U, editors. Modern Crop Protection Compounds. 3nd ed. Weinheim: Wiley-VCH Verlag GmbH & Co. KGaA; 2019. pp. 1630-1640 - 130.
Pitterna T, Böger M, Maienfisch P. Gem-difluorovinyl derivatives as insecticides and acaricides. Chimia. 2004; 58 (3):108-116. DOI: 10.2533/000942904777678163 - 131.
Oka Y. Sensitivity to fluensulfone of inactivated Meloidogyne spp. second-stage juveniles. Pest Management Science. 2020;76 (7):2379-2387. DOI: 10.1002/ps.5774 - 132.
Oka Y. Nematicidal activity of fluensulfone against some migratory nematodes under laboratory conditions. Pest Management Science. 2014; 70 (12):1850-1858. DOI: 10.1002/ps.3730 - 133.
Grabau ZJ, Noling JW, Navia Gine PA. Fluensulfone and 1,3-dichloroprene for plant-parasitic nematode management in potato production. Journal of Nematology. 2019; 51 :1-12. DOI: 10.21307/jofnem-2019-038 - 134.
Ludlow K. Public Release Summary on the Evaluation of the New Active Fluensulfone in Product NIMITZ 480 EC Nematicide. Australian Pesticides and Veterinary Medicines Authority, Kingston [Internet]. 2015. Available from: https://apvma.gov.au/sites/default/files/publication/15596-prs-fluensulfone.pdf [Accessed: June 01, 2023] - 135.
Norshie PM, Grove IG, Back MA. Persistence of the nematicide fluensulfone in potato ( Solanum tuberosum ssp. tuberosum) beds under field conditions. Nematology. 2017;19 (6):739-747. DOI: 10.1163/15685411-00003085 - 136.
Faske T, Brown KA, Kelly JC. Toxicity of Tioxazafen to Meloidogyne incognita andRotylenchulus reniformis . Journal of Nemtology. 2022;54 (1):1-6. DOI: 10.2478/jofnem-2022-0007 - 137.
Slomczynska U, South MS, Bunkers GJ, Edgecomb D, Wyse-Pester D, Selness S, et al. Tioxazafen: A new broad-spectrum seed treatment nematicide. In: ACS Symposium Series. Washington, DC: American Chemical Society; 2015. pp. 129-147. DOI: 10.1021/bk-2015-1204.ch010 - 138.
South SM, Wilson D, Spal S, Slomczynska U, Bunkers GJ, et al. Development of Tioxazafen as a new broad spectrum Nematicide. In: Jeschke P, Witschel M, Kramer W, Schirmer U, editors. Modern Crop Protection Compounds. 1st ed. Germany: Wiley-VCH; 2019. pp. 1615-1628. DOI: 10.1002/9783527644179 - 139.
Gopalsamy A, Shi M, Golas J, Vogan E, Jacob J, Johnson M, et al. Discovery of benzisoxazoles as potent inhibitors of chaperone heat shock protein 90. Journal Medical Chemistry. 2008; 51 (3):373-375. DOI: 10.1021/jm701385c - 140.
Lamberth C. Oxazole and isoxazole chemistry in crop protection. Journal of Heterocyclic Chemistry. 2018; 55 (9):2035-2045. DOI: 10.1002/jhet.3252 - 141.
Wayne N, Mishra P, Bolon DN. Hsp90 and client protein maturation. In: Calderwood SK, Prince TL, editors. Molecular Chaperones: Methods in Molecular Biology. Vol. 787. New York: Springer Science+Business Media; 2011. pp. 33-44. DOI: 10.1007/978-1-61779-295-3_3 - 142.
Devaney E, O’Neill K, Harnett W, Whitesell L, Kinnaird JH. Hsp90 is essential in the filarial nematode Brugia pahangi . International Journal for Parasitology. 2005;35 (6):627-636. DOI: 10.1016/j.ijpara.2005.01.007 - 143.
Gelmedin V, Delaney A, Jennelle L, Hawdon JM. Expression profile of heat shock response factors during hookworm larval activation and parasitic development. Molecular and Biochemical Parasitology. 2015; 202 (1):1-14. DOI: 10.1016/j.molbiopara.2015.08.003 - 144.
Somogyvári M, Gecse E, Sőti C. DAF-21/Hsp90 is required for C. elegans longevity by ensuring DAF-16/FOXO isoform a function. Scientific Reports. 2018;8 (1):12048. DOI: 10.1038/s41598-018-30592-6 - 145.
Wolterink G, Boobis AR. Tioxazafen. In: Joint Meeting of the FAO Panel of Experts on Pesticide Residues in Food and the Environment and the WHO Core Assessment Group on Pesticide Residues. Pesticide Residues in Food - 2018: Toxicological Evaluations. Berlin, Germany, 18-27 September 2018. Geneva: World Health Organization and Food and Agriculture Organization of the United Nations; 2019. Licence: CC BY-NC-SA 3.0 IGO. Available from: https://inchem.org/documents/jmpr/jmpmono/v2018pr01.pdf [Accessed: 2023-06-01] - 146.
Uzuele EL. Eficácia do tratamento de sementes com o nematicida tioxazafen no controle dos nematoides Heterodera glycines ,Meloidogyne incognita , M. javanica, Pratylenchus brachyurus e P. zeae [thesis]. Piracicaba: Universidade de São Paulo; 2016 - 147.
Walker DM, Graham CR, Miller WR, Koeller KJ. Three step, one-pot process to prepare thiophene-2-carbonyl chloride (TCC), a key raw material in the manufacture of tioxazafen (Nemastrike®). Tetrahedron Letters. 2019; 60 (12):834-838. DOI: 10.1016/j.tetlet.2019.02.015 - 148.
Lahm GP, Wiles JA, Cordova D, Thoden T, Desaeger J, Smith BK. Fluazaindolizine: A new active ingredient for the control of plant-parasitic nematodes. In: Jeschke P, Witschel M, Krämer W, Schirmer U, editors. Modern Crop Protection Compounds. 3nd ed. Weinheim: Wiley-VCH Verlag GmbH & Co. KGaA; 2019. pp. 1630-1640 - 149.
EPA. Memorandum Supporting Revised Proposed Decision to Approve Registration for the New Active Ingredient Fluazaindolizine [Internet]. 2023. Available from: https://www.regulations.gov/search?filter=EPA-HQ-OPP-2020-0065 [Accessed: June 04, 2023] - 150.
Groover WL, Lawrence KS. Evaluation of a new chemical nematicide, fluazaindolizine (Reklemel™ active), for plant-parasitic nematode management in bermudagrass. Journal of Nematology. 2021; 53 :1-14. DOI: 10.21307/jofnem-2021-043 - 151.
Leonardi R, Jackowski S. Biosynthesis of pantothenic acid and coenzyme a. EcoSal Plus. 2007; 2 (2):1-22. DOI: 10.1128/ecosalplus.3.6.3.4 - 152.
Craig JP, Bekal S, Niblack T, Domier L, Lambert KN. Evidence for horizontally transferred genes involved in the biosynthesis of vitamin B(1), B(5), and B(7) in Heterodera glycines . Journal of Nematology. 2009;41 (4):281-290 - 153.
Siddique S, Radakovic ZS, Hiltl C, Pellegrin C, Baum T, Beasley H, et al. The genome and lifestage-specific transcriptomes of a plant-parasitic nematode and its host reveal susceptibility genes involved in trans-kingdom synthesis of vitamin B5. Nature Communications. 2022; 13 (1):6190. DOI: 10.1038/s41467-022-33769-w - 154.
Thoden TC, Wiles JA. Biological attributes of Salibro™, a novel sulfonamide nematicide. Part 1: Impact on the fitness of Meloidogyne incognita ,M. hapla andAcrobeloides buetschlii . Nematology. 2019;21 (6):625-639 - 155.
Thoden T, Pardavella IV, Tzortzakakis EA. In vitro sensitivity of different populations of Meloidogyne javanica andM. incognita to the nematicides Salibro™ and Vydate®. Nematology. 2019;21 (8):889-893 - 156.
Wu HY, de Oliveira SJ, Becker JS, Becker JO. Fluazaindolizine mitigates plant-parasitic nematode activity at sublethal dosages. Journal of Pest Science. 2020; 94 (2):573-583. DOI: 10.1007/s10340-020-01262-2 - 157.
Qiao K, Liu Q , Zhang S. Evaluation of fluazaindolizine, a new nematicide for management of Meloidogyne incognita in squash in calcareous soils. Crop Protection. 2021; 143 :105469. DOI: 10.1016/j.cropro.2020.105469 - 158.
Watson TT, Desaeger JA. Evaluation of non-fumigant chemical and biological nematicides for strawberry production in Florida. Crop Protection. 2019; 117 :100-107. DOI: 10.1016/j.cropro.2018.11.019 - 159.
Desaeger JA, Watson TT. Evaluation of new chemical and biological nematicides for managing Meloidogyne javanica in tomato production and associated double-crops in Florida. Pest Management Science. 2019;75 (12):3363-3370. DOI: 10.1002/ps.5481 - 160.
Regmi H, Desaeger J. Integrated management of root-knot nematode ( Meloidogyne spp.) in Florida tomatoes combining host resistance and nematicides. Crop Protection. 2020;134 :105170. DOI: 10.1016/j.cropro.2020.105170 - 161.
USEPA. Memorandum Supporting Revised Proposed Decision to Approve Registration for the New Active Ingredient Fluazaindolizine [Internet]. 2023. Available from: https://www.regulations.gov/search?filter=EPA-HQ-OPP-2020-0065 - 162.
Lin H, Pang K, Ma Y, Hu J. Photodegradation of fluazaindolizine in water under simulated sunlight irradiation: Identification of transformation products and elucidation of transformation mechanism. Chemosphere. 2019; 214 :543-552. DOI: 10.1016/j.chemosphere.2018.09.151 - 163.
Pang N, Lin H, Hu J. Photodegradation of fluazaindolizine in aqueous solution with graphitic carbon nitride nanosheets under simulated sunlight illumination. Ecotoxicology and Environmental Safety. 2019; 170 :33-38. DOI: 10.1016/j.ecoenv.2018.11.019