Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
Cellular and cell-free therapies have provided novel therapeutic opportunities for treating various incurable diseases. This is because the mesenchymal stromal/stem cells (MSCs) produce a plethora of bioactive molecules able to target different biological pathways through extracellular vesicle-mediated paracrine mechanisms simultaneously. However, to share transcriptomic signatures with their origin tissue, it is expected that each MSC population has a unique molecular profiler. In this sense, to analyze the transcriptome of therapeutic cells, it is crucial to identify the molecular profiler of these cells to predict the potential clinical benefits promoted by these cells. Herein, we discuss the application of high-throughput RNA-sequencing (RNA-Seq) in the Pharma Intelligence Era, discussing and exemplifying how the combination of molecular biology with Analytics can revolutionize the Research, Development, & Innovation (RD&I) of advanced medicinal products.
Genetics Laboratory, Butantan Institute, São Paulo, SP, Brazil
Postgraduate Program in Structural and Functional Biology, Paulista School of Medicine (EPM), Federal University of São Paulo (UNIFESP), São Paulo, Brazil
BioDecision Analytics Limited, São Paulo, Brazil
João Rafael Dias Pinto
BioDecision Analytics Limited, São Paulo, Brazil
Getulio Vargas Foundation, São Paulo, Brazil
Irina Kerkis
Genetics Laboratory, Butantan Institute, São Paulo, SP, Brazil
Postgraduate Program in Structural and Functional Biology, Paulista School of Medicine (EPM), Federal University of São Paulo (UNIFESP), São Paulo, Brazil
*Address all correspondence to: rodrigo.pinheiro.araldi@gmail.com
1. Introduction
Since their discovery, mesenchymal stromal/stem cells (MSCs) have been proposed as the best candidate for treating non-curable diseases [1, 2, 3, 4, 5, 6, 7].
Stem cells can be obtained from both embryonic and adult tissues. Embryonic stem cells (ESCs) are derived from the inner cell mass of the human blastocyst at the pre-implantation stage. However, hESC are pluripotent cells and can differentiate into all cell types, which compose adult organisms. Therefore, when transplanted in an animal model, these cells form teratomas—benign tumors that contain derivate of all three germ layers, the most typical ones being bone, cartilage, neuronal rosettes, and epithelium of the airways and gut [8]. By contrast, human adult stem cells (hASCs) comprise a population of undifferentiated precursor cells that naturally reside in all tissues and organs. However, hASCs show limited differentiating capability (multipotent cells), and they do not confer any risk of forming teratomas. This is one of the reasons why hASCs are the most studied type of stem cell for therapeutic purposes.
The bone marrow hASCs are classified as stromal cells or mesenchymal stem cells (MSCs). Stromal cells are progenitors of the hematopoiesis-supporting stroma and are components of skeletal tissue such as bone, cartilage, and adipocytes. MSCs can be isolated from numerous adult and perinatal tissues, such as bone marrow (BM), umbilical cord (UC) vein, Wharton’s jelly, adipose and placental tissues, menstrual blood (MB), liver, spleen, and dental pulp (DP) [2, 3, 9, 10]. Moreover, MSCs can be easily expanded in vitro and in accordance with Good Manufacturing Process (GMP) conditions, which is crucial to guarantee a large-scale production of therapeutic cells for therapeutic purposes.
For a long time, it has been suggested that the therapeutic properties of stem cells were based on the capacity of these cells to replace dead cells in injured tissues [3, 11]. However, studies showed that transplanted MSCs remain in the injury site for a few days and, subsequently, are not found in the tissue [11, 12, 13]. These results were later confirmed by numerous independent studies [10, 14, 15], which reported that less than 1% of transplanted MSCs survive for more than 1 week after systemic administration [16, 17]. Historically, these studies broke the paradigm that the therapeutic action of these cells is based on their capability to replace dead cells. Despite the therapeutic properties and the facilities to isolate and produce these cells, the mechanism of action (MoA) of the MSCs remains not completely understood.
Supporting this breaking paradigm, Takahashi et al. [18] showed that injection of MSC conditioned culture medium (CCM) increases the capillary density, reduces the damage size, and improves the cardiac function in murine models for heart attack. This result, combined with previous evidence that less than 1% of MSCs survive for more than 1 week after systemic administration, provided strong evidence that the therapeutic properties of MSCs are mediated by the “secretome” of these cells, which is composed of a plethora of bioactive molecules [2, 19, 20].
To date, several bioactive molecules have already been identified in the MSCs’ secretome, such as chemokines cytokines, interleukins, growth factors, lipid steroids, nucleotides, nucleic acids, ions, and metabolites [2, 21, 22]. These molecules can be found in free form (when directly released into the extracellular space) or within extracellular vesicles (EVs), which are recognized as a key component of paracrine secretion [6, 22, 23].
The rediscovery of EVs and the discovery that these vesicles mediate cell-to-cell communication led to the conceptualization of the paracrine stimulation theory [11, 24]. This theory postulates that MSCs release a variety of bioactive molecules into the intercellular space, playing a key role in cross-talk communication between the cells and the surrounding tissues [1, 2, 3, 10, 25]. Thus, the MSCs’ secretome is the key mediator of the regenerative action of these cells [6, 11, 21, 26]. For this reason, in 2017, Prof. Dr. Arnold Caplan, who isolated for the first time the MSC population for therapeutic purposes, proposed to rename these cells to signaling cells [27].
The paracrine stimulation theory not only brought novel horizons to elucidate the MoA of stem cells but also allowed us to explore the biotechnological potential of these EVs in a new therapeutic modality known as cell-free therapy [2, 6, 22, 28].
2. Cell-free therapy: novel horizons for the treatment of non-curable diseases
The discovery that the MSC conditioned culture medium (CCM) elicits therapeutic properties similar to the MSCs [18] aroused the interest in exploring the content of this CCM as an alternative to cell therapy. However, CCM contains components such as bovine fetal serum, antibiotics, and pH indicators that limit the direct application of this medium for clinical purposes. For this reason, the CCM is discarded as a subproduct of the MSC manufacturing process. Thus, biopharmaceutical companies are discarding a valuable source of feedstock rich in bioactive molecules that could be explored for clinical purposes in cell-free therapy [6, 28].
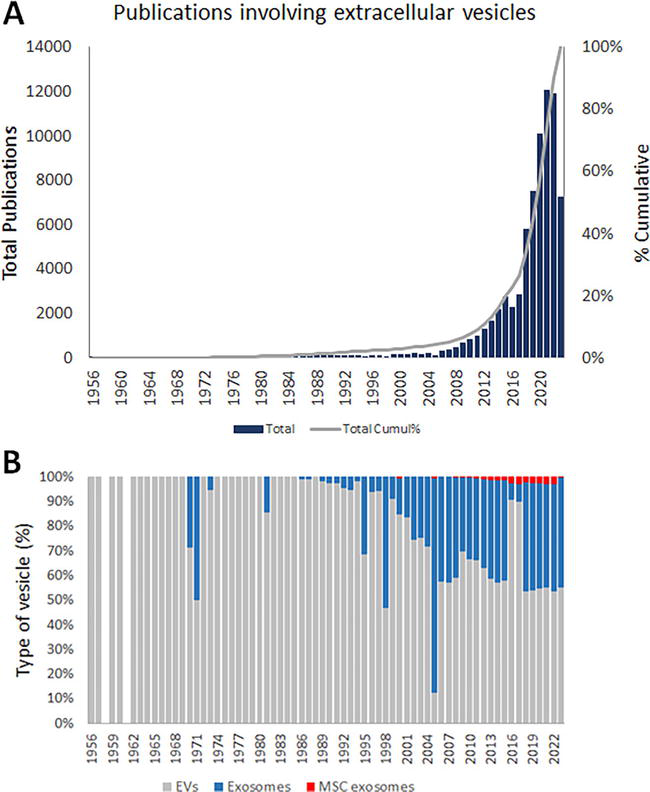
Although the MSC-derived bioactive molecules could be found in a free form diluted in the CCM, generally, in this form these bioactive molecules are easily degraded or oxidized. By contrast, when released within EVs, these bioactive molecules are preserved, increasing the half-life of these molecules [6, 28]. Therefore, it is not surprising that cell-free therapy is based on the therapeutic use of these EVs, justifying the increased interest in these vesicles, as illustrated by the number of publications involving EVs shown in Figure 1.
Figure 1.
Number of publications involving extracellular vesicles from 1956 to October 10, 2023 (A). Numbers show an increased interest in these vesicles, which has increased since 2008. Results also show an increased interest in exosomes from 2004 (B), a specific class of EV. However, the number of publications using exosomes derived from MSCs has been reduced since 2010 (B).
Although the interest in these vesicles had emerged two decades ago (Figure 1), the EVs were discovered in the second half of the 1940s, when Chargaff, studying blood coagulation, observed small “membrane debris” sedimented at high-speed centrifugation of plasma supernatant [29, 30]. Twenty years later, after Chargaff’s report, Peter Wolf confirmed the existence of corpuscles, “platelet dust,” through electron microscopy [31]. However, the biological nature of these corpuscles remained unknown until 1974, when Nunez et al. [32] identified structures with a size under 1000 nm, called multivesicular bodies (MVBs), opening up the path in the identification of a subtype of EVs that later was called exosomes or small EVs (30–150 nm) [33].
The EVs comprise a heterogenous group of small cell-released particles [34], which are classified according to their size and biogenesis in exosomes (30–200 nm), microvesicles (150–1500 nm), and apoptotic bodies (1000–5000 nm) [6, 35, 36]. Among these EVs, exosomes are the most studied and useful type of EV for therapeutic purposes [6, 36]. For this reason, we will focus on this class of EV.
Exosomes are nanosized vesicles, surrounded by a phospholipid membrane, containing cholesterol, sphingomyelin, ceramide, lipid rafts, and evolutionarily conserved markers such as tetraspanins (CD9, CD63, and CD81), heat shock proteins (HSP60, HSP70, and HSP90), major histocompatibility component (MHC) classes I and II, Alix, TSG101, lactadherin, and lysosome-associated membrane glycoprotein 2 [6].
Exosomes constitutively originate from late endosomes, which are formed by inward budding of the limited multivesicular body (MVB) [6, 37, 38].
MVBs and late endosomes are a subset of specialized endosomal compartments rich in intraluminal vesicles (ILVs) [39]. During the ILV formation process, certain proteins are incorporated into the invaginating membrane, while the cytosolic components are engulfed and enclosed within the ILV [38, 40, 41].
Along the biogenesis, the endosomal sorting complex required for transport (ESCRT), which consists of ESCRT-0, -I, -II, -III, combined with the vacuolar protein sorting 4-vesicle trafficking 1 (VPS4-VTA1), and some accessory proteins such as the ALG-2-interacting protein X (ALIX) homodimer [42] promotes the sorting of specific biomolecules to the exosomes [43].
Upon maturation, the MVBs can be transported to the plasma membrane via the cytoskeletal and microtubule network and undergo exocytosis post-fusion with the cell surface, whereby the ILVs get secreted as exosomes [40, 44]. Alternatively, MVBs can also follow a degradation pathway either by direct fusion with lysosomes or by fusion with autophagosomes followed by lysosomes [39]. Although both the secretory and degradative MVB pathways coexist, the mechanism that influences these pathways remains unclear [39].
Once released into the extracellular space, exosomes interact with the extracellular matrix and recipient cells, serving as natural vehicles for the nanodelivery of nucleic acids, proteins, metabolites, and lipids [6, 36, 39].
Considering that all cell types naturally produce and secrete exosomes, all cell types can be considered a source of exosomes [6, 28]. However, for therapeutic purposes, the MSCs emerge as the most important source of therapeutic exosomes. This is because, as previously discussed, MSCs can be easily isolated and produced on a large scale. In addition, these cells naturally produce a plethora of bioactive molecules that can elicit therapeutic responses. Thus, considering that during the biogenesis, the ESCRT complex promotes the sorting of specific molecules that may be driven to the lumen of these nanosized vesicles, it is expected that the molecular cargo of exosomes changes according to their origin. Therefore, no prototype MSCs can serve as a universal source of exosomes to treat all types of diseases. For this reason, knowing the molecular profiler (transcriptomic signature of MSCs) can help us to identify appropriate exosomes for the treatment of a set of diseases. In this regard, RNA-Seq emerges as a useful tool to identify the molecular profiler of these cells and their respective exosomes and, with this, to predict the clinical application of these cells and/or exosomes.
3. RNA-Seq analysis applied to identification of molecular profiler of ATMPs
The advent of high-throughput techniques presents a multitude of opportunities to the biopharmaceutical industry [45, 46]. Among these techniques, transcriptome, which includes RNA sequencing (RNA-Seq) and single-cell RNA sequencing (scRNA-Seq), stands out due to their capability to assess qualitative and quantitatively the gene expression levels, serving as important tools to characterize MSCs and MSC-derived exosomes with therapeutic interest. While RNA-Seq can be applied to characterize a cell or exosome population, scRNA-Seq is useful to analyze the heterogenicity of cell type in the MSC population. This chapter will focus on RNA-Seq since this technique is most useful to characterize MSC or MSC-derived exosome populations and predict their therapeutic properties.
Due to the complexity of the RNA-Seq data, several recommendations have been proposed to guarantee the quality of the results obtained with this technique [46, 47, 48, 49]. In general, these recommendations are focused on experimental questions, such as the number of samples, sequencing depth, quality control, read alignment, and quantification of gene levels [47, 48, 49]. However, there is no single analysis pipeline to be used in all cases. In this sense, the multiple bioinformatic tools, especially those applied to assess the differential expression, make the RNA-Seq data analysis a laborious task, requiring basic knowledge of genetics, bioinformatics, and statistics [46].
Statistical analyses comprise a mandatory step to identify differentially expressed genes (DEGs), which can be used to characterize the molecular profile and predict the therapeutic potential of MSCs or MSC-derived exosomes. Although the statistical techniques required for these purposes are included in the packages used to analyze the RNA-Seq, the choice for inappropriate controls and/or the lack of the most powerful analytics techniques, particularly those based on artificial intelligence (AI), can negatively affect the power of the transcriptome to predict the therapeutic potential of cell and cell-free products.
However, the combined use of traditional statistical methods with Analytics tools to analyze Pharma Big Data such as those generated by the RNA-Seq has revolutionized both pharmaceutical and biopharmaceutical industries. The combination of these techniques applied to analyze (bio)pharmaceutical data led to the advent of Pharma Intelligence, which has revolutionized the research, development, & innovation (RD&I) in life sciences [46].
4. Pharma Intelligence applied to the identification of transcriptomic signature of different types of MSCs
Pharma Intelligence (PI) is a generic term employed to refer to the combined use of mathematics, statistics, Data Analytics (or Analytics), Business Intelligence (BI), bioinformatics, and artificial intelligence (AI) to analyze and extract valuable information from big data produced in the multi-Omics Era [46].
Pharma Intelligence emerged as a need of the pharmaceutical sector that, over the last two decades, evidenced numerous disadvantages of the traditional empirical drug development model [46]. The traditional empirical drug development model is based on meeting regulatory requirements to guarantee the investigational product (IP) gets market access instead of providing the most useful details about the IP, such as the eventual side effects and even the mechanism of action (MoA), which could predict the drug repositioning, accelerating the RD&I in the pharmaceutical area [46]. For this reason, historical evidence of the traditional empirical drug development model not only increases the time to obtain the registry of new (bio)pharmaceutical products for the treatment of NCDs but also causes economic losses to the entire pharmaceutical supply chain. In this regard, technologies that could predict the eventual risk of the IP, and MoA are crucial to mitigate the failure rate of the IP during the clinical trials.
On the one hand, the advent of “Omics” technologies (genomics, transcriptome, proteome, and metabolome) revolutionized the RD&I in the (bio)pharmaceutical sector; on the other hand, these multi-Omics tools have contributed to the generation of high amount of data (Big data) that are not efficiently used by (bio)pharmaceutical industry to generate valuable insights [46, 47, 50, 51]. Moreover, due to the complex nature of the pharmaceutical data, which combine structured (tabular data) and non-structured data (image bank, sound, and video recording from clinical trial), the conventional statistical techniques are not sufficient to efficiently explore these datasets [46]. This occurs because standard statistical tests assume that a single hypothesis is tested. However, when multiple hypotheses are tested simultaneously, there is a higher probability of a type I error (false-positive)—an incorrect rejection of a true null hypothesis [46, 52]. However, PI has allowed us to overcome the challenges imposed by the Pharmaceutical Big Data Era, helping to select variables of interest, revealing unexpected correlations among these variables, and significantly reducing the time and cost of drug discovery [46, 52, 53].
In addition, the PI possibilities the reanalyze of datasets available on public data banks, such as the Sequence Read Archive (SRA) of the National Institute of Health (NIH, https://www.ncbi.nlm.nih.gov/sra), facilitating the RD&I of ATMPs.
5. Pharma intelligence applied to prediction of therapeutic potential of MSCs and MSC-derived exosomes
To share a transcriptomic signature with their origin tissue, each MSC population is expected to possess different molecular profiles. Thus, it is not surprising that there is no universal MSC that can confer therapeutic benefits for all diseases. For this reason, identifying the molecular profile (or transcriptomic signature) of each MSC population is crucial to predicting the therapeutic potential, as well as the mechanism of action (MoA) of these cells.
In this sense, the SRA can serve as a useful source of RNA-Seq datasets of MSCs and MSC-derived exosomes to predict the appropriate use of each MSC or MSC-derived exosome for target disease.
Generally, standard analysis of RNA-Seq, using conventional packages (DSeq2, NOISeq, edgeR, etc.) to assess the genes differentially expressed, provides noise results. This is because these packages are based on statistical tests that provide p-values. Thus, all gene that shows a p-value <0.05 is recognized as differentially expressed. However, slight biological expression differences result in a significant p-value (p-value <0). Although commonly employed, analysis using this analytical approach can result in the identification of numerous genes/transcripts with low biological relevance. Alternatively, some bioinformaticians have calculated the log2 fold-chance (log2FC) and adopted a cutoff value that could justify a phenotypic difference (i.e., a biological response), assuming that this cutoff represents a statistically significant difference. However, using appropriate analytical strategies based on PI, it is possible to combine the log2FC to statistical tests to select genes of interest accurately for downstream prediction analyses.
This strategy was currently used to assess the transcriptional profile of human immature dental pulp stem cells (hIDPSCs) produced on a large scale [54]. In order to increase the predictor power of this method, the study compared the hIDPSCs transcriptional profile with the transcriptome signature of other MSCs obtained from adipose tissue (AD-MSC, BM-MSC (HD-MSC, hepatocyte-derived-MSC), menstrual blood (MB-MSC), UC-MSC and vertebral tissue (vMSC) [54]. For this, the study employed a dataset comprised of more than 130 samples from the SRA [54]. Results obtained by the PI showed that the hIDPSCs have a unique transcriptome when compared with other analyzed MSCs [54]. The study also identified that the hIDPSCs express 375 unique genes (Table 1), which are not expressed by other MSCs [54]. These unique genes were subjected to the biological functional enrichment analysis. Interestingly, the results of these analyses suggest that the hIDPSCs have a higher capacity to mediate cell communication when compared to other MSCs and that the genes exclusively expressed by the hIDPSCs regulate metabolic processes and neurogenesis [54]. To confirm experimentally these results, primary motor neurons from transgenic mice for amyotrophic lateral sclerosis (ALS) were treated with three different concentrations of exosomes isolated from the conditioned culture medium of these cells for up to 14 days. Results showed that the exosomes derived from the hIDPSCs were able to promote both neurite and axon guidance length in a concentration- and time-dependent manner [54]. These data provide evidence that the PI applied to the analysis of RNA-Seq of therapeutic cells has an accurate predictor power.
Genes commonly e exclusively expressed in each MSC population.
In relation to the total genes identified/MSC population.
Using this same analytical approach (based on PI), herein we reanalyzed these data, showing that the MSCs share the expression of 5912 genes in common, which represents between 61.62% (MB-MSC) to 85.53% (BM-MSC) of the genes encoded by these cells (Table 1).
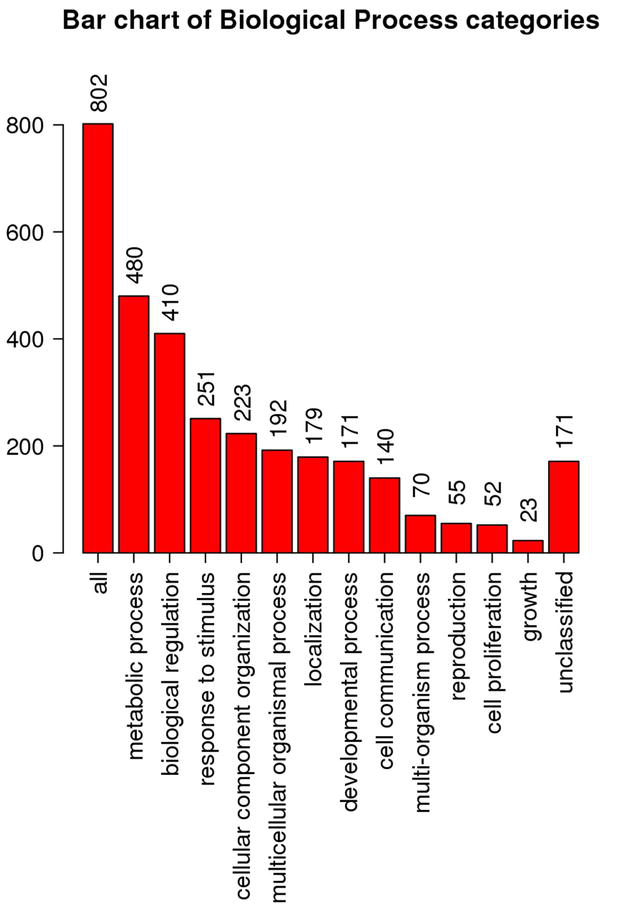
Aiming to predict the biological pathways in which these 5912 genes commonly expressed by all MSCs, it was employed the same functional enrichment technique used to predict the hIDPSCs therapeutic properties. Results showed that 98.34% (5814/5912) genes commonly expressed by all MSCs encode proteins, which are involved in (i) metabolic process (4246) and (ii) biological regulation (3877), (iii) response to stimulus (2811), (iv) cellular component organization (2557), and (v) cell communication (1968) (Figure 2). The high number of cell communication-related genes identified as commonly expressed by the MSCs reinforces the paracrine stimulation theory, suggesting that the therapeutic effects of MSCs can be mediated by the bioactive molecules naturally produced and secreted by these cells within EVs.
Figure 2.
Biological functional enrichment in terms of biological process of the 5814 genes commonly expressed by the MSC populations analyzed. Results were obtained using the WEB gestalt tool, available from: https://www.webgestalt.org/.
Biological function enrichment also showed that the genes commonly expressed by the MSCs are involved in (i) transcriptional and (ii) translational regulation, being associated with protein folding and polyubiquitination (Table 2). Interestingly, the products of these genes are also involved in (iii) mitochondrial regulation, (iv) cell survival, (v) angiogenesis, and (vi) vesicle-mediated transport (Table 2). These data show that MSCs have a complex mechanism of action since the proteins encoded by these cells can act synergically and simultaneously in multiple biological pathways, which are found deregulated in most diseases.
Enrichment result
FDR
p-value
Gene set size
Overlap
Geneontology (GO)
Regulation of catabolic process (GO:0009894)
0.000
0.000
875
510
Positive regulation of catabolic process (GO:0031331)
0.000
0.000
348
204
Vesicle-mediated transport (GO:0016192)
0.000
0.000
1942
899
Translational initiation (GO:0006413)
0.000
0.000
192
154
RNA binding (GO:0003723)
0.000
0.000
1603
1056
RNA processing (GO:0006397)
0.000
0.000
487
337
Protein targeting (GO:0006605)
0.000
0.000
412
264
Protein folding (GO:0006457)
0.000
0.000
210
131
Protein polyubiquitination (GO:0000209)
0.000
0.000
255
155
Ubiquitin ligase complex (GO:0000151)
0.000
0.000
273
166
Mitochondrial protein complex (GO:0098798)
0.000
0.000
266
167
Pathway (KEGG, Pnather, Reactome, Wikipathay)
Protein processing in endoplasmic reticulum (has04141)
0.000
0.000
165
125
Ubiquitin-mediated proteolysis (hsa04120)
0.000
0.000
136
100
Ubiquitin proteasome pathway (P00060)
0.000
0.000
44
37
p38 MAPK pathway (P05918)
0.000
0.000
34
24
RAS pathway (P04393)
0.000
0.000
70
48
PI3 kinase pathway (P00048)
0.000
0.000
47
30
PDGF signaling pathway (P00047)
0.000
0.000
125
72
Integrin signaling pathway (P000340
0.000
0.000
166
95
P53 pathway (P00059)
0.000
0.000
79
44
Transcription regulation of TP53 (R-HAS-3700989)
0.000
0.000
365
221
Vesicle-mediated transport (R-HAS-5653656)
0.000
0.000
667
397
Cellular response to stress (R-HAS-2262752)
0.000
0.000
426
242
PTEN regulation (R-HAS-6807070)
0.000
0.000
140
104
mRNA processing (WP411)
0.000
0.000
126
108
Electron transport chain (OXOPHOS system in mitochondria) (WP111)
0.000
0.000
103
71
VEGFA-VEGFR2 signaling pathway (WP3888)
0.000
0.000
431
280
EGF/EGFR signaling pathway (WP437)
0.000
0.000
162
109
Table 2.
Biological functional enrichment of the genes commonly by all analyzed MSC population.
FDR – false discovery ratio; GO – Geneontology; has – KEGG; P – Panther; R-HAS – Reactome; WP – Wikipathway.
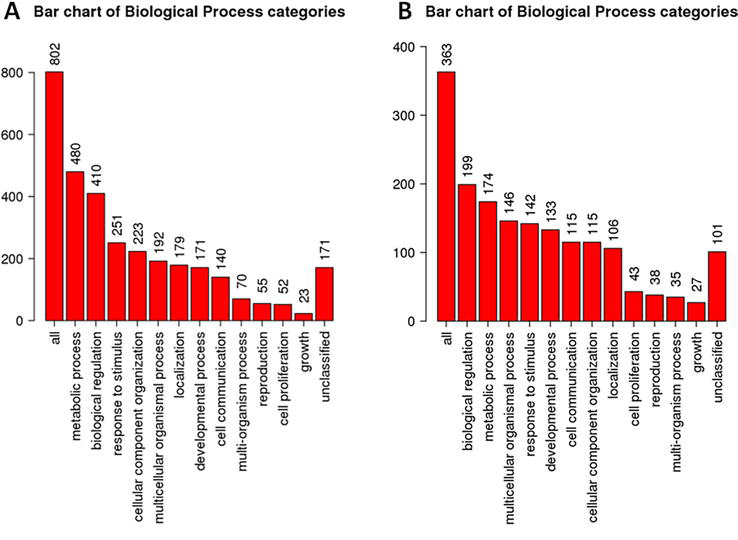
Using the same approach, we also analyzed the set of genes identified as exclusively expressed by the MB-MSCs, since this MSC population showed the highest number of unique genes, followed by the hIDPSCs (Table 1). Results showed that 96.39% of the genes exclusively expressed by the MB-MSCs (802/832) encode proteins, which are related to (i) metabolic process (480), (ii) biological regulation (410), (iii) response to stimulus 251), (iv) component organization (223), and other biological process, including (v) cell communication (Figure 3).
Figure 3.
Biological functional enrichment in terms of biological process of the genes exclusively expressed by the MB-MSCs (A) and hIDPSCs (B) results were obtained using the WEB gestalt tool, available from: https://www.webgestalt.org/.
Biological function enrichment analyses of the genes exclusively expressed by the MB-MSCs showed that most of the products encoded by these genes bind to the DNA, acting as transcriptional factors, regulating cellular responses to DNA damages, or are involved in DNA replication, as part of the replication fork (Table 3).
Enrichment result
FDR
p-value
Overlap
Geneontology (GO)
RNA modification (GO:0009541)
0.000
0.000
41
ncRNA processing (GO:0034470)
0.000
0.000
38
DNA repair (GO:0006281)
0.000
0.000
43
Cellular response to DNA damage stimulus (GO:0006974)
Interestingly, the reanalyzing of the 375 genes exclusively expressed by the hIDPSCs showed that, although the unique genes found in hIDPSCs are involved in the same biological process regulated by the unique genes found in MB-MSCs (Figure 3), they govern distinct biological pathways (Table 4), conferring different therapeutic properties for these two populations of MSC. This is because the unique genes expressed by hIDPSCs are involved in neurogenesis and synapsis (Table 4), suggesting that these cells can be explored for the treatment of neurological disorders, such as neurodegenerative diseases.
Enrichment results
FDR
p-value
Overlap
GO
Neurogenesis (GO:0022008)
0,000
0,000
51
Neuron spine (GO:0044309)
0,007
0,000
10
Dendritic spine (GO:0043197)
0,0000
0,007
10
Neuron to neuron synapsis (GO:0098984)
0,019
0.000
13
Table 4.
Biological functional enrichment of the genes exclusively expressed by hIDPSC population.
FDR – false discovery ratio; GO – Geneontology.
Confirming the predictor power of the Pharma Intelligence applied to cell therapy, preclinical studies involving the intravenous administration of the hIDPSCs in rat models for Huntington’s (HD, neurodegenerative disorder characterized by a progressive loss of medium spiny neurons (MSNs) in the striatum, causing motor, cognitive and neuropsychiatric disorders) provided evidence that these cells can cross the brain-blood barrier and engraft within the brain areas involved in the HD pathophysiology. Besides this, the study showed that the hIDPSCs administration increases the expression of MSN biomarkers, suggesting a neuroregenerative action [55]. In another independent study, using a rat model for Parkinson’s disease, it was also observed that the intravenous administration of these cells ameliorated the motor function of these animals only 3 days after the cell transplantation [56]. Similar results were also demonstrated in phase I clinical trial for HD, which provided evidence that the transplantation of hIDPSCs can improve the motor function of patients with HD [57].
6. Perspectives of the use of pharma intelligence in cell and cell-free therapy
Although already commonly used by pharmaceutical companies to accelerate the RD&I in drug discovery, the complex nature of MSCs and/or MSC-derived exosomes still has limited the application of Pharma Intelligence as part of the characterization and, as a tool to predict the therapeutic potential of cells and exosomes. However, fortunately, today, we already have specialists and facilities providing innovative solutions that combine mathematical, statistics, analytics, and bioinformatic tools to extract valuable information from multi-Omics, preclinical, and even clinical datasets. Although the number of specialists and companies dedicated to employing Pharma Intelligence in RD&I of cell and cell-free therapy remains reduced when compared to the number of companies offering services dedicated to drug discovery, PI is no longer a promise for the future of these therapies. PI is the New Era of advanced medical therapies.
Moreover, the capability of the algorithms to predict results that can drive the correct use of each MSC population or even each MSC-derived exosome has improved, particularly with the combined use of AI. Section 6 exemplifies how the appropriate use of PI can drive the correct use of MSC populations for clinical purposes.
However, the recent emergence of ChatGPT (which is a type of AI) and the idealization of the use of AI brought by science fiction (sci-fi) could lead to a false interpretation of the power of these techniques, creating an over-expectation about the PI.
In this sense, PI may be understood as a powerful predictor tool to characterize molecular cells and exosome candidates for clinical purposes that can bring many advantages to the R&D of cell and cell-free therapies. Thus, PI serves as a tool to facilitate decision-making, allowing us to identify the best application or predict the MoA of a specific MSC population or MSC-derived exosome.
On the one hand, the PI brings novel opportunities, including job offers. On the other hand, it requires qualified professionals who have expertise in life science (particularly with expertise in cell and cell-free therapy) and analytics. This is because the predictive value of any result obtained using PI depends on many factors, such as (i) the data quality, (ii) data organization and availability, advanced knowledge in (iii) programming, (iv) statistics, (v) bioinformatics, (vi) molecular and cellular biology, and in (vii) the pathophysiology of the target disease. Despite this complexity, which requires a multidisciplinary effort to extract valuable information from data, PI can be considered a revolutionary tool for the biopharmaceutical sector, accelerating the time to obtain the registry of an investigational product and reducing costs with non-necessary experiments needed to bring the proof of concept (POC) required for Regulatory Agencies.
References
1.Baek G, Choi H, Kim Y, Lee HC, Choi C. Mesenchymal stem cell-derived extracellular vesicles as therapeutics and as a drug delivery platform. Stem Cells Translational Medicine. 2019;8:880-886
2.L. PK, Kandoi S, Misra R, Vijayalakshmi S, Rajagopal K, Verma RS. The mesenchymal stem cell secretome: A new paradigm towards cell-free therapeutic mode in regenerative medicine. Cytokine Growth Factor Review [Internet]. 2019;46(April):1-9. Available from: doi:10.1016/j.cytogfr.2019.04.002
3.Cheng L, Zhang K, Wu S, Cui M, Xu T. Focus on mesenchymal stem cell-derived exosomes: Opportunities and challenges in cell-free therapy. Stem Cells International. 2017;2017:1-10
4.Evangelista A, Soares MBP, Villarreal CF. Cell-free therapy: A neuroregenerative approach to sensory neuropathy? Neural Regeneration Research. 2019;14(8):1383-1384
5.Bianco P, Robey O, Simmons P. Mesenchymal stem cells: Revisiting history, concepts and assays. Cell Stem Cell. 2008;2(4):313-319
6.Araldi RP, D’Amelio F, Vigerelli H, de Melo TC, Kerkis I. Stem cell-derived exosomes as therapeutic approach for neurodegenerative disorders: From biology to biotechnology. Cells [Internet]. 2020;9(12):2663. Available from: https://www.mdpi.com/2073-4409/9/12/2663
7.Kerkis I, Araldi R, Wenceslau C, Mendes T. Advances in cellular and cell-free therapy medicinal products for Huntington’s disease treatment. In: From Physiopathology to Treatment of Huntington’s Disease [Internet]. London, UK: IntechOpen; 2022. pp. 1-27. DOI: 10.1016/j.colsurfa.2011.12.014
8.Calabrese EJ. Hormesis and embryonic stem cells. Chemico-Biological Interactions [Internet]. 2022;352:109783. Available from: https://linkinghub.elsevier.com/retrieve/pii/S000927972100421X
9.Lee JY, Hong S-H. Hematopoietic stem cells and their roles in tissue regeneration. International Journal of Stem Cells [Internet]. 2020;13(1):1-12. Available from: http://www.ijstemcell.com/journal/view.html?doi=10.15283/ijsc19127
10.Fujita Y, Kadota T, Araya J, Ochiya T, Kuwano K. Clinical application of mesenchymal stem cell-derived extracellular vesicle-based therapeutics for inflammatory lung diseases. Journal of Clinical Medicine. 2018;7(10):355
11.Gomzikova MO, Rizvanov AA. Current trends in regenerative medicine: From cell to cell-free therapy. Bionanoscience [Internet]. 2017;7(1):240-245. DOI: 10.1007/s12668-016-0348-0
12.Duffield JS, Park KM, Hsiao L-L, Kelley VR, Scadden DT, Ichimura T, et al. Restoration of tubular epithelial cells during repair of the postischemic kidney occurs independently of bone marrow-derived stem cells. Journal of Clinical Investigation [Internet]. 2005;115(7):1743-1755. Available from: http://www.jci.org/articles/view/22593
13.Biancone L, Bruno S, Deregibus MC, Tetta C, Camussi G. Therapeutic potential of mesenchymal stem cell-derived microvesicles. Nephrology Dialysis Transplantation [Internet]. 2012;27(8):3037-3042. Available from: https://academic.oup.com/ndt/article-lookup/doi/10.1093/ndt/gfs168
14.Gao J, Dennis JE, Muzic RF, Lundberg M, Caplan AI. The dynamic in vivo distribution of bone marrow-derived mesenchymal stem cells after infusion. Cells Tissues Organs [Internet]. 2001;169(1):12-20. Available from: https://www.karger.com/Article/FullText/47856
15.Lee RH, Pulin AA, Seo MJ, Kota DJ, Ylostalo J, Larson BL, et al. Intravenous hMSCs improve myocardial infarction in mice because cells embolized in lung are activated to secrete the anti-inflammatory protein TSG-6. Cell Stem Cell [Internet]. 2009 Jul;5(1):54-63. Available from: https://linkinghub.elsevier.com/retrieve/pii/S1934590909002124
16.Eggenhofer E, Benseler V, Kroemer A, Popp FC, Geissler EK, Schlitt HJ, et al. Mesenchymal stem cells are short-lived and do not migrate beyond the lungs after intravenous infusion. Frontiers in Immunology [Internet]. 2012;3:3. Available from: http://journal.frontiersin.org/article/10.3389/fimmu.2012.00297/abstract
17.Song YS, Lee HJ, Doo SH, Lee SJ, Lim I, Chang K-T, et al. Mesenchymal stem cells overexpressing hepatocyte growth factor (HGF) inhibit collagen deposit and improve bladder function in rat model of bladder outlet obstruction. Cell Transplantion [Internet]. 2012 Aug;21(8):1641-1650. Available from: http://journals.sagepub.com/doi/full/10.3727/096368912X637488
18.Takahashi M, Li T-S, Suzuki R, Kobayashi T, Ito H, Ikeda Y, et al. Cytokines produced by bone marrow cells can contribute to functional improvement of the infarcted heart by protecting cardiomyocytes from ischemic injury. American Journal of Physiology-Heart and Circulatory Physiology [Internet]. 2006;291(2):H886-H893. Available from: https://www.physiology.org/doi/10.1152/ajpheart.00142.2006
19.Gurunathan S, Kang M-H, Jeyaraj M, Qasim M, Kim J-H. Review of the isolation, characterization, biological function, and multifarious therapeutic approaches of exosomes. Cell. 2019;8(4):307
20.Haraszti RA, Miller R, Stoppato M, Sere YY, Coles A, Didiot MC, et al. Exosomes produced from 3D cultures of MSCs by tangential flow filtration show higher yield and improved activity. Molecular Therapy [Internet]. 2018;26(12):2838-2847. DOI: 10.1016/j.ymthe.2018.09.015
21.Haider KH, Aramini B. Mircrining the injured heart with stem cell-derived exosomes: An emerging strategy of cell-free therapy. Stem Cell Research & Therapy. 2020;11(1):1-12
22.Teixeira MR, Alievi AL, da Costa VR, Dias Pinto JR, Araldi RP. Therapeutic potential of extracellular vesicles in Parkinson’s disease. Exploration of Neuroscience [Internet]. 2023;29:106-122. Available from: https://www.explorationpub.com/Journals/en/Article/100616
23.Han C, Sun X, Liu L, Jiang H, Shen Y, Xu X, et al. Exosomes and their therapeutic potentials of stem cells. Stem Cells International. 2016;2016:1-11
24.Baraniak PR, McDevitt TC. Stem cell paracrine actions and tissue regeneration. Regenerative medicine [Internet]. 2010 Jan;5(1):121-143. Available from: https://www.futuremedicine.com/doi/10.2217/rme.09.74
25.Vizoso FJ, Eiro N, Cid S, Schneider J, Perez-Fernandez R. Mesenchymal stem cell secretome: Toward cell-free therapeutic strategies in regenerative medicine. International Journal of Molecular Sciences. 2017;18(9):1852
26.Hu P, Yang Q , Wang Q , Shi C, Wang D, Armato U, et al. Mesenchymal stromal cells-exosomes: A promising cell-free therapeutic tool for wound healing and cutaneous regeneration. Burn Trauma. 2019;7:1-10
27.Caplan AI. Mesenchymal stem cells: Time to change the name! Stem Cells Translational Medicine [Internet]. 2017;6(6):1445-1451. Available from: https://academic.oup.com/stcltm/article/6/6/1445/6448531
28.da Costa VR, Araldi RP, Vigerelli H, D’Ámelio F, Mendes TB, Gonzaga V, et al. Exosomes in the tumor microenvironment: From biology to clinical applications. Cells [Internet]. 2021;10(10):2617. Available from: https://www.mdpi.com/2073-4409/10/10/2617
29.Chargaff E. Cell structure and the problem of blood coagulation. Journal of Biological Chemistry [Internet]. 1945;160(1):351-359. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0021925818431316
30.Chargaff E, West R. The biological significant of the thromboplastic protein of blood. The Journal of Biological Chemistry. 1946;166(1):189-197
31.Wolf P. The nature and significance of platelet products in human plasma. British Journal of Haematology [Internet]. 1967;13(3):269-288. Available from: https://onlinelibrary.wiley.com/doi/10.1111/j.1365-2141.1967.tb08741.x
32.Nunez EA, Wallis J, Gershon MD. Secretory processes in follicular cells of the bat thyroid. III. The occurrence of extracellular vesicles and colloid droplets during arousal from hibernation. American Journal of Anatomy [Internet]. 1974;141(2):179-201. Available from: https://onlinelibrary.wiley.com/doi/10.1002/aja.1001410203
33.Palazzolo S, Canzonieri V, Rizzolio F. The history of small extracellular vesicles and their implication in cancer drug resistance. Frontiers in Oncology [Internet]. 2022;24:12. Available from: https://www.frontiersin.org/articles/10.3389/fonc.2022.948843/full
34.Théry C, Witwer KW, Aikawa E, Alcaraz MJ, Anderson JD, Andriantsitohaina R, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV 2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. Journal of Extracellular Vesicles. 2018;7(1):1535750
35.Sheta M, Taha EA, Lu Y, Eguchi T. Extracellular vesicles: New classification and tumor immunosuppression. Biology (Basel) [Internet]. 2023;12(1):110. Available from: https://www.mdpi.com/2079-7737/12/1/110
36.Da Costa VR, Araldi RP, Vigerelli H, D’Ámelio F, Mendes TB, Gonzaga V, et al. Exosomes in the tumor microenvironment: From biology to clinical applications. Cell. 2021;10(10):1-27
37.Sarko DK, McKinney CE. Exosomes: Origins and therapeutic potential for neurodegenerative disease. Frontiers in Neuroscience. 2017;11:1-7
38.Zhang Y, Liu Y, Liu H, Tang WH. Exosomes: Biogenesis, biologic function and clinical potential. Cell & Bioscience [Internet]. 2019;9(1):19. Available from: https://cellandbioscience.biomedcentral.com/articles/10.1186/s13578-019-0282-2
39.Gurung S, Perocheau D, Touramanidou L, Baruteau J. The exosome journey: From biogenesis to uptake and intracellular signalling. Cellular Communication Signalling [Internet]. 2021;19(1):1-19. Available from: https://biosignaling.biomedcentral.com/articles/10.1186/s12964-021-00730-1
40.Han Q-F, Li W-J, Hu K-S, Gao J, Zhai W-L, Yang J-H, et al. Exosome biogenesis: Machinery, regulation, and therapeutic implications in cancer. Molecular Cancer [Internet]. 2022;21(1):207. Available from: https://molecular-cancer.biomedcentral.com/articles/10.1186/s12943-022-01671-0
41.Bebelman MP, Smit MJ, Pegtel DM, Baglio SR. Biogenesis and function of extracellular vesicles in cancer. Pharmacology & Therapeutics [Internet]. 2018;188:1-11. DOI: 10.1016/j.pharmthera.2018.02.013
42.Ju Y, Bai H, Ren L, Zhang L. The role of exosome and the ESCRT pathway on enveloped virus infection. International Journal of Molecular Sciences. 2021;22(16):9060
43.Wang M, Yu F, Ding H, Wang Y, Li P, Wang K. Emerging function and clinical values of exosomal microRNAs in cancer. Molecular Therapy—Nucleic Acids [Internet]. 2019;16:791-804. Available from: https://linkinghub.elsevier.com/retrieve/pii/S2162253119301209
44.Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478):eaau6977
45.Vamathevan J, Clark D, Czodrowski P, Dunham I, Ferran E, Lee G, et al. Applications of machine learning in drug discovery and development. Nature Reviews Drug Discovery [Internet]. 2019;18(6):463-477. Available from: http://www.nature.com/articles/s41573-019-0024-5
46.Faustinoni-Neto B, Dias-Pinto JR, Araldi RP. Pharma data analytics (pharma intelligence): Opportunities and challenges in the multi-omics era of drug discovery and development. Pharmacologia. 2023;14(1):29-39
47.Conesa A, Madrigal P, Tarazona S, Gomez-Cabrero D, Cervera A, McPherson A, et al. A survey of best practices for RNA-seq data analysis. Genome Biology. 2016;17(1):1-19
48.Tarazona S, Furió-Tarí P, Turrà D, Di PA, Nueda MJ, Ferrer A, et al. Data quality aware analysis of differential expression in RNA-seq with NOISeq R/bioc package. Nucleic Acids Research [Internet]. 2015;43(21):e140. Available from: https://academic.oup.com/nar/article-lookup/doi/10.1093/nar/gkv711
49.Tarazona S, García F, Ferrer A, Dopazo J, Conesa A. NOIseq: A RNA-seq differential expression method robust for sequencing depth biases. EMBnet Journal [Internet]. 2012;17(B):18. Available from: http://journal.embnet.org/index.php/embnetjournal/article/view/265
50.Chung M, Bruno VM, Rasko DA, Cuomo CA, Muñoz JF, Livny J, et al. Best practices on the differential expression analysis of multi-species RNA-seq. Genome Biology [Internet]. 2021;22(1):121. Available from: https://genomebiology.biomedcentral.com/articles/10.1186/s13059-021-02337-8
51.Luecken MD, Theis FJ. Current best practices in single-cell RNA-seq analysis: A tutorial. Molecular Systems Biology [Internet]. 2019;15(6):e8746. Available from: https://onlinelibrary.wiley.com/doi/10.15252/msb.20188746
52.Hernandez I, Zhang Y. Using predictive analytics and big data to optimize pharmaceutical outcomes. American Journal of Health-System Pharmacy [Internet]. 2017;74(18):1494-1500. Available from: https://academic.oup.com/ajhp/article/74/18/1494/5102866
53.Parikh S, Patel R, Khunt D, Chavda VP, Vora L. Data analytics and data visualization for the pharmaceutical industry. In: Bioinformatics Tools for Pharmaceutical Drug Product Development [Internet]. Beverly, MA, USA: Wiley; 2023. pp. 55-76. DOI: 10.1002/9781119865728.ch4
54.Araldi RP, Viana M, Colozza-gama GA. Unique transcriptional signatures observed in stem cells from the dental pulp of deciduous teeth produced on a large scale. Pharmacologia. 2023;14(1):72-95
55.Wenceslau CV, de Souza DM, Mambelli-Lisboa NC, Ynoue LH, Araldi RP, da Silva JM, et al. Restoration of BDNF, DARPP32, and D2R expression following intravenous infusion of human immature dental pulp stem cells in Huntington’s disease 3-NP rat model. Cells [Internet]. 2022;11(10):1664. Available from: https://www.mdpi.com/2073-4409/11/10/1664
56.Araldi RP, Ramos AT, Alievi AL, Policíquio B, Teixei MR, Mendes TB, et al. NestaCell® promotes motor, cognitive and neuropsychiatric functions amelioration and dopaminergic neurons restoration in a preclinical model of Parkinson’s disease. Cytotherapy [Internet]. 2022;24(10):S3. Available from: https://linkinghub.elsevier.com/retrieve/pii/S1465324922008490
57.Macedo J, Pagani E, Wenceslau C, Ferrara L, Kerkis I. A phase I clinical trial on intravenous administration of immature human dental pulp stem cells (NestaCell) to Huntington’s disease patients. Cytotherapy [Internet]. 2021;23(4):1. Available from: https://linkinghub.elsevier.com/retrieve/pii/S1465324921000657
Written By
Rodrigo Pinheiro Araldi, João Rafael Dias Pinto and Irina Kerkis
Submitted: 25 September 2023Reviewed: 10 October 2023Published: 06 March 2024
 Open access peer-reviewed chapter
Open access peer-reviewed chapter