Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
Human immunodeficiency virus type 1 (HIV-1) is tropic for humans and replicates in virtually none of the other animal species. While various animal models to mimic the conflict between HIV-1 and human hosts have been proposed, nonhuman primates (NHPs) are thought to be most suitable from a purely scientific point of view for the HIV-1/AIDS model studies. Because NHPs are resistant to HIV-1, remodeling the HIV-1 genome is required to validate the productive infection of NHPs. Two types have been reported as retrofitted viruses, that is, SHIVs and HIV-1 derivatives. SHIVs are SIVs (simian immunodeficiency viruses) that carry a small portion of the HIV-1 genome, whereas HIV-1 derivatives are HIV-1 with a minimal sequence/genome modification. SHIVs have been successfully used for studies specifically targeting HIV-1 Pol-RT (reverse transcriptase) and Env proteins. HIV-1 derivatives can induce AIDS in NHPs under certain conditions. More importantly, HIV-1 derivatives contribute to elucidating the HIV-1 adaptation and virus-host interaction through analyzing the process of acquiring replication capacity and pathogenicity in restrictive hosts distinct from natural hosts. In this chapter, we summarize NHP model studies on HIV-1/AIDS using SIV, SHIV, or HIV-1 derivatives and discuss the significance of HIV-1 derivatives toward understanding the HIV-1 biology.
Department of Microbiology, Graduate School of Medicine, Tokushima University, Tokushima, Japan
Naoya Doi
Department of Microbiology, Graduate School of Medicine, Tokushima University, Tokushima, Japan
Bao Quoc Le
Department of Microbiology, Graduate School of Medicine, Tokushima University, Tokushima, Japan
Tomoyuki Kondo
Department of Microbiology, Graduate School of Medicine, Tokushima University, Tokushima, Japan
Akio Adachi
Department of Microbiology, Graduate School of Medicine, Tokushima University, Tokushima, Japan
Masako Nomaguchi*
Department of Microbiology, Graduate School of Medicine, Tokushima University, Tokushima, Japan
*Address all correspondence to: nomaguchi@tokushima-u.ac.jp
1. Introduction
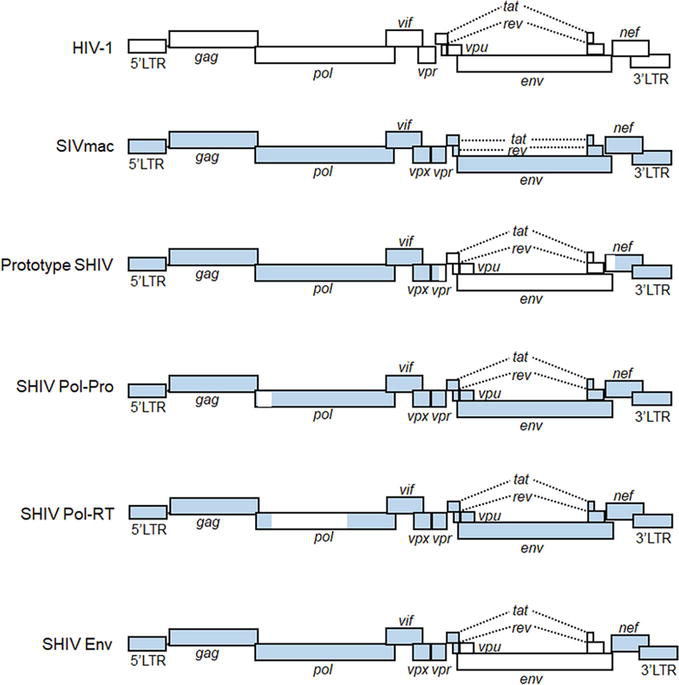
HIV-1 belongs to a subfamily Lentivirinae of the family Retroviridae [1]. As the name indicates, lentiviruses can cause persistent infections in hosts for a long time (almost silent sometimes) with slowly developing symptoms [2, 3, 4]. During the persistent infection state, they are transmissible from infected individuals to the other susceptible host individuals and finally spread to a large population of susceptible hosts. Another important feature of lentiviruses is their unique replication style itself as retroviruses. They reverse transcribe their genomes by their own reverse transcriptase (RT) and integrate them into the host cell DNA in nuclei. Thus, lentiviruses establish a very intimate and lengthy relationship with their hosts. Gene expression (transcription, alternative splicing, nuclear export, and translation) from the integrated genome elaborately proceeds by utilizing cis-acting elements on the viral genome [2, 3, 4]. Within the HIV-1 genome, there are nine genes (two of which are split genes) that can finally produce as many as fifteen proteins (Figure 1). Following the production of viral proteins, two identical genome copies are incorporated into assembled progeny virions, which is unique to the family Retroviridae among viral families. Each retrovirus replication phase, from early (from virus-receptor/coreceptor binding to viral genome integration into host cell DNA) to late (from gene expression of proviral genome to virion production) phases, is a strictly regulated process [2].
Figure 1.
Schematic genome representation of HIV-1, SIVmac, and various SHIVs. White and blue areas indicate sequences from HIV-1 and SIVmac, respectively. Pol-Pro, pol-protease; Pol-RT, pol-reverse transcriptase; LTR, long terminal repeat.
One of the important biological features of HIV-1 is to specifically adapt itself to replication in humans, exhibiting a narrow host range virtually only to humans. Thus, no appropriate experimental animals that are susceptible to authentic HIV-1 infection and pathogenesis are currently available. Unadulterated HIV-1 clones can establish spreading infection in humanized mice (mice reconstituted with human hematopoietic cells), although they are not pathogenic to humanized mice. HIV-1-infected humanized mice are useful for basic and clinical studies such as those on determination of the in vivo activity/function of viral accessory proteins, on analysis of the virus adaptation process, and on evaluation of the efficacy of various antivirals [5, 6, 7, 8, 9]. However, it is still critically required for the humanized mouse system to improve the immune responses in the animals, the acquired immunity in particular [5, 6, 7, 8, 9]. On one hand, simian immunodeficiency viruses (SIVs)- or chimeric viruses of SIV and HIV-1 (SHIVs)-infected macaques have been used for basic/clinical studies (including those regarding the development of vaccines and antivirals) as a model system to mimic viral replication and pathogenesis in HIV-1-infected humans (Figure 1). Macaque-tropic HIV-1 derivatives, which have minimal modifications in the HIV-1 genome and have the ability to infect macaques, also have been developed [10, 11, 12, 13, 14, 15, 16, 17, 18]. Collectively, three distinct classes of viruses, that is, SIVs, SHIVs, and macaque-tropic HIV-1 derivatives, have their own characteristics of replicative and pathogenic properties in macaque cells and individuals. In this chapter, we outline the knowledge on viral replication, adaptation, and pathogenicity obtained from macaque models using SIVs, SHIVs, and macaque-tropic HIV-1 derivatives. We also describe the advantage of macaque-tropic HIV-1 derivatives/macaque infection systems to understand the HIV-1 biology.
2. The emergence of HIV-1 and its narrow host range
HIV-1 exhibits a narrow host range and cannot replicate in macaque cells as described above. The excellent study using SHIV clones first demonstrated that HIV-1 replication in macaque cells is blocked at the post-entry step [19]. Results obtained in the study indicated that Gag-Pol and Vif derived from SIVmac are most likely to be necessary for viral replication in macaque cells [19]. Several years later, intrinsic restriction factors that potently inhibit HIV-1 replication in macaque cells (APOBEC3 protein family, TRIM5 protein family, and tetherin) were identified one after another [20, 21, 22, 23]. APOBEC3 proteins have cytidine deaminase activity, and its major function is to introduce lethal mutations into the HIV-1 genome. TRIM5 proteins target incoming viral cores and perturb the reverse transcription step of HIV-1 replication. Tetherin suppresses virus budding by tethering progeny virions at the plasma membrane as its name suggests. HIV-1 Vif, Gag-CA, and Vpu can antagonize APOBEC3 proteins, TRIM5 proteins, and tetherin in human cells, respectively, but not at all in macaque cells. The actions of these intrinsic restriction factors explain really well why HIV-1 cannot replicate in macaque cells.
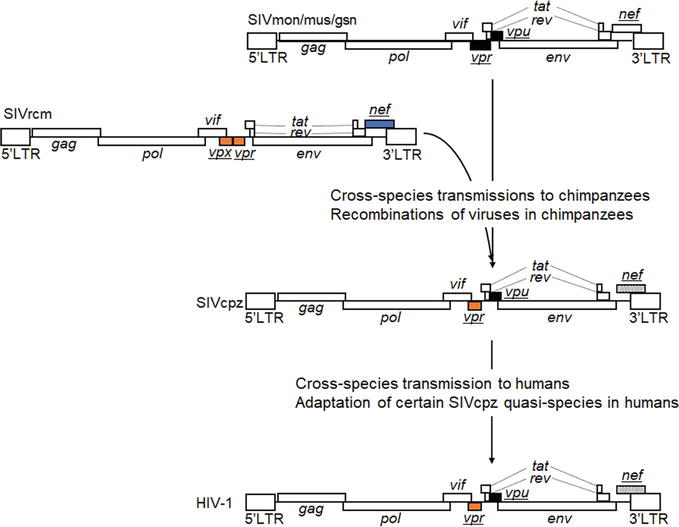
While HIV-1 replication is completely blocked in macaques, HIV-1 has been shown to have emerged through the repeated cross-species transmissions of various SIVs from their natural hosts to the new hosts and the viral recombination/adaptation in transmitted hosts (Figure 2) [20, 24, 25]. In brief, SIVmon/mus/gsn, which parasitize Mona monkey, Mustached monkey, and Greater spot-nosed monkey as natural hosts, respectively, and SIVrcm from Red-capped mangabey co-infected chimpanzees by cross-species transmissions. SIVcpz arose by their recombination in chimpanzees and resulted in the elimination of vpx from SIVrcm and also in the acquisition of vpu and removal of env-nef overlapping from SIVmon/mus/gsn. Then, certain SIVcpz quasi-species that were produced during continuous replication in chimpanzees cross-species transmitted to humans [20, 24, 25]. To investigate experimentally the emergence process of HIV-1, for example, what functional/structural changes occurred in SIVs during cross-species transmission and adaptation in chimpanzees and humans, may help uncover virus-host interactions that are key determinants for the HIV-1 emergence, that is, the basis for species-specific tropism of HIV-1.
Figure 2.
The emergence of HIV-1 from ancestral SIVs. HIV-1 emergence has been suggested to have resulted from cross-species transmissions of SIVs and their recombinations in transmitted hosts [20, 24, 25]. For phylogenic relationships between HIV-1 and related SIVs, refer to references [26, 27]. Genome organizations of various SIVs and HIV-1 are shown. Accessory genes other than vif are underlined and colored to see which gene have derived from each SIV. Nef genes of SIVcpz and HIV-1 are highlighted because of the loss of env-nef overlapping from other SIVs.
Taking the strict human tropism of HIV-1 into consideration, researchers have made every effort to generate appropriate model study systems for the HIV-1/AIDS research by selecting viruses, selecting host animals, genetically altering viruses, and/or genetically altering host animals [10, 11, 12, 13, 14, 15, 16, 17, 18]. A wide variety of animal species from mice to primates and of lentivirus species have been proposed for the surrogate models and extensively tested for their usability and for their scientific value. Among these, NHP models using primate immunodeficiency viruses (SIVmac and SHIV) (Figure 1) are most suitable for the HIV-1/AIDS research from various points of view, such as pathogenic outcome and robust immune responses [13, 17, 28]. Due to the close similarity in genetic background and physiological status, researchers have plenty and successful experience in the NHP system to carry out various basic research on viruses and/or to do translational research to develop drugs, vaccines, and other interventions [10, 11, 12, 13, 14, 15, 16, 17, 18]. Of the three macaque species (rhesus, cynomolgus, and pigtailed macaques) frequently used for experimental virus infections, the pigtailed and rhesus macaques are known to be most susceptible and resistant to the SIV infection, respectively. Pigtailed macaques genetically lack the restriction of TRIM5alpha against viruses via the Gag-CA. On one hand, the cynomolgus macaque rarely develops AIDS upon infection with SIV. Indeed, rhesus macaques responded to infections with SIVmac and SHIV similarly as observed for the case of HIV-1 and humans, and some of them finally came down with AIDS. Only the rhesus macaque and SIV/SHIV system consistently give this result [13, 16].
To construct a novel class of infectious molecular clones carrying the HIV-1 genes that replicate in macaque cells, two different approaches have been taken. One is the SHIV (Figure 1), that is, a virus clone constructed in the background of SIVmac genome, and another is the HIV-1 derivative, that is, a variant HIV-1 clone with a minimal sequence modification in the context of HIV-1 genome [29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44]. While the former virus clones commonly have been designated SHIV [31], the latter clones have been variously called as HIV-1 derivative [34], stHIV-1 [33], HIV-1mt [39], HSIV [40], or HIV-1rmt [43]. Research on the SHIV preceded study on the HIV-1 derivative. As for SHIVs, it is reasonable to construct clones carrying the HIV-1 genes/domain sequences that are not determinants for its species tropism. Even if the chimeric constructs mainly consist of SIVmac sequences/genes, the activity and/or function of the inserted HIV-1 genes in the replication cycle can be readily assessed individually or combinedly in vivo in a demonstrative way. In addition, the mutations/adaptations of the inserted HIV-1 genes in various in vivo environments can be experimentally monitored. Thus, SHIV-RT (reverse transcriptase) [32], SHIV-Pro [36], and SHIV-Env [38, 41] of a direct progeny of the prototype SHIV [29] in a sense were generated (Figure 1) and used for experiments. It was highly expected that these SHIVs are very useful to study the basic virus property in vivo, the effect of antiviral drugs in vivo, the emergence of drug-resistance variants, and/or the host immunological responses associated with the concerned viral proteins. In fact, a large number of scientific reports on SHIV-RTs have been published (for example, see references [45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59]). More impressively, SHIV-Env constructs have played a critical role in the HIV-1/AIDS research activity as fully revealed by selected, only a small portion of scientific articles published so far [60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81]. From a medical point of view, SHIV constructs need to have RT and Env sequences derived from HIV-1 for in vivo RT and Env studies, respectively. Since there are intrinsic and critical differences in biological and molecular properties of RT and Env proteins between HIV-1 and SIVmac, authentic SIVmac and its equivalent viruses cannot be used or are of a significantly limited value as a model virus of HIV-1 for the medical research on some specific subjects [10, 12]. Importantly, SIVmac RT significantly differs in the susceptibility to anti-RT drugs from HIV-1 RT [32]. And also, SIVmac Env is so virologically and immunologically distinct from HIV-1 Env [76], as is natural for the well-recognized variable nature of primate lentiviral Env proteins. The distinct outcomes of infection have been observed in SIVs/SHIVs-infected NHPs used for HIV research, for example, development of AIDS-like diseases with rapid progression (in a few months) or with several months/years of chronic phases. The outcomes can be determined by the complex interaction between viruses and hosts depending on materials and methods used for infection, that is, virus species/strains, NHP species, infection routes, numbers of virus challenges, virus doses, and host restriction factors such as MHC and intrinsic restriction factors (for SIVs/SHIVs pathogenesis, see well-organized figures and tables in reviews [13, 17, 28, 82, 83]). In total, various SIV/SHIV clones have successfully paved the way to the HIV-1/macaque model infection system. There are excellent reviews on experimental infection studies using NHPs (macaques) and SIV/SHIV clones and also on their applications such as understanding the HIV-1 latency and establishing the HIV-1 cure strategies [28, 82].
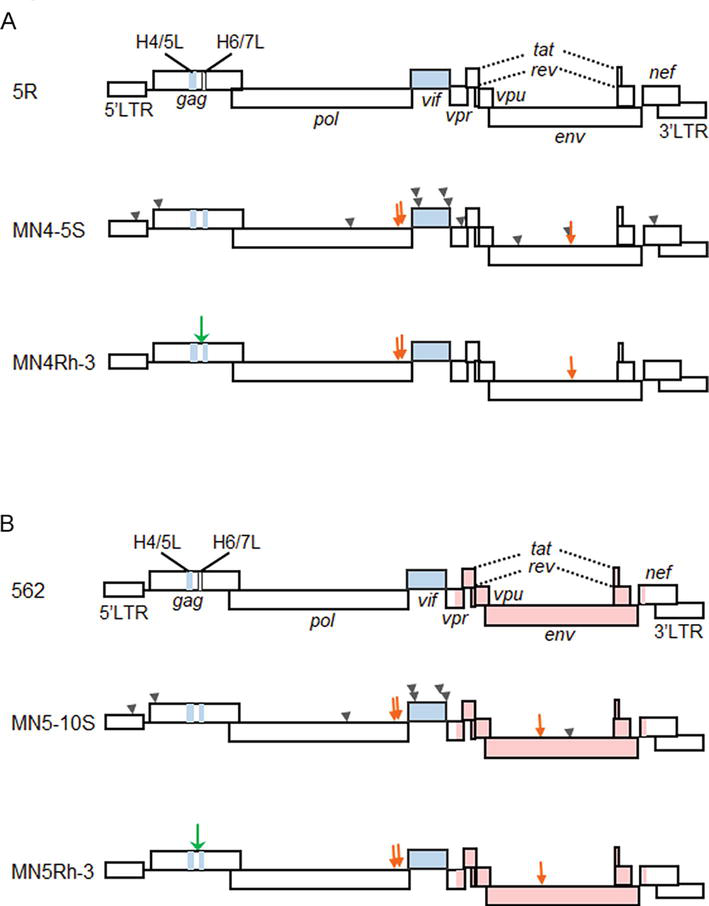
It has been well-recognized and generalized that the restriction of viral replication occurs at the entry step on the cell surface and/or at some intracellular stage(s) [84]. As clarified by our early studies, the HIV-1 species tropism is determined at the early post-entry phase, within the viral DNA synthesis event [19, 29]. The viral determinants for the tropism were predicted to be Gag-CA and Vif as described above. To generate a novel class of HIV-1 that can replicate in macaque cells, it was necessary to genetically modify the two viral genomic regions. We first checked whether a small portion of gag-CA region of HIV-1 genome encoding CypA-binding region was replaceable with the corresponding site of SIVmac genome [85, 86]. The proviral clones containing the chimeric Gag-CA of various lengths were found to be viable in human cells but not in macaque cells [85, 86]. We next constructed a new clone designated NL-ScaVR and also generated its adapted version in macaque cells designated NL-DT5R (5R in Figure 3). The two constructs are CXCR4-tropic as a parental HIV-1 NL4–3 clone [89, 90], and carry a small portion of gag-CA region and the entire vif gene derived from SIVmac. The HIV-1 derivative clone NL-DT5R was able to establish infections in mononuclear cells (PBMCs) and individuals of the pigtailed macaque [34, 35]. We also observed that NL-DT5R does not replicate virtually in rhesus macaque PBMCs. In parallel with our work, another group constructed a similar HIV-1 clone designated stHIV-1 containing most of the gag-CA region and the entire vif region from SIVmac239 [33] and showed that it replicates quite well in rhesus macaque cells but does not cause any diseases in pigtailed macaques [33, 37].
Figure 3.
Schematic representation of various HIV-1mt genomes. CXCR4-tropic and CCR5-tropic clones are shown in panels (A) and (B), respectively. Green and orange arrows indicate the structure guided and spontaneously occurred mutations with growth-enhancing effects, respectively [87, 88]. Arrow heads are the neutral/negative mutations. Pink areas show the region derived from CCR5-tropic virus clone [89]. H4/5L, Gag-CA loop between helices 4 and 5; H6/7L, Gag-CA loop between helices 6 and 7.
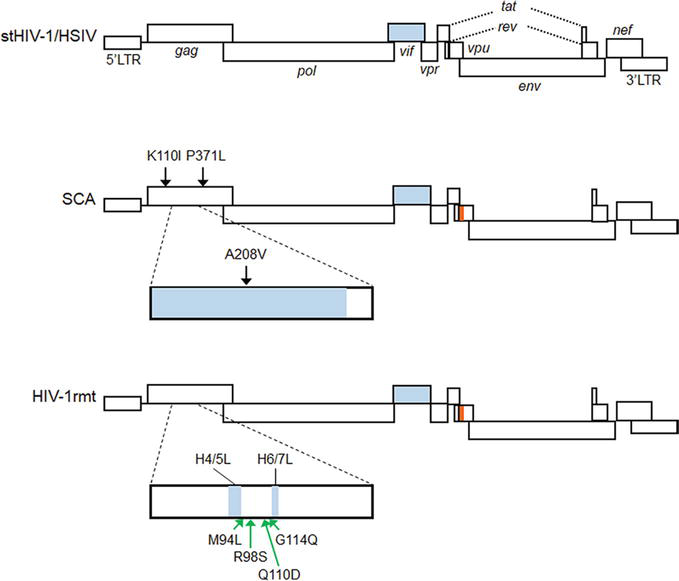
In order to improve the replication ability in macaque cells of NL-DT5R, and also to construct CCR5-tropic clones, we altered virus genomes in a stepwise manner by the sequence/structure-guided mutagenesis and by virus adaptation in cells (Figure 3). NL-DT5R was found to be rather sensitive, if not completely, to cellular restriction factors TRIM5 and APOBEC3 proteins [91]. First, we replaced the loop domain of HIV-1 Gag-CA between helices 6 and 7 with the corresponding site of SIVmac (MN4-5S in Figure 3A) [39, 92] to increase resistance to TRIM5, and then randomly modify the viral genome by the adaptation in macaque cells and structure-guided mutagenesis (MN4-Rh-3 in Figure 3A) [42, 87]. CCR5-tropic MN5Rh-3 that carries the env gene from the HIV-1 SF162 clone was similarly constructed (Figure 3B). At each modification step, the viral replication abilities of both in vitro and in vivo were experimentally confirmed to be enhanced in parallel. However, the growth potential, even for MN4Rh-3 and MN5Rh-3, was still insufficiently augmented to grow well in rhesus macaque cells. Considering these clones have the intact vif gene from SIVmac sufficiently active against the rhesus APOBEC3 restriction factor, Gag-CA was thought to be the main target for further modification to counteract the rhesus TRIM5 restriction factor. Another cellular restriction factor tetherin may affect the HIV-1 growth in macaque cells. To overcome the severe restriction imposed by rhesus cells, we finally modified the gag-CA region of the MN4Rh-3 version and replaced its 5′ portion of the vpu gene with the corresponding part of SIVgsn (SIV isolated from the greater spot-nosed monkey) [43]. The resultant proviral clone was designated LSDQ (HIV-1rmt in Figure 4). LSDQ grew comparably well in rhesus PBMCs with a standard SIVmac clone (MA239) pathogenic for rhesus macaques [43].
Figure 4.
Schematic representation of HIV-1rmt genomes. For comparison, the genomes of stHIV-1 (pig-tailed macaque version) [37, 93, 94] and HSIV [40, 95] are shown at the top. White, blue, and orange areas indicate sequences from HIV-1, SIVmac, and SIVgsn, respectively. Neutral and growth-enhancing mutations relative to parental clones are shown by black and green arrows, respectively. See references [43, 96] for detail.
Finally, the growth potential of CXCR4-tropic and CCR5-tropic HIV-1rmt clones was examined in rhesus macaques [44]. Both clones grew comparably well in rhesus PBMCs. When inoculated into rhesus macaques, the two virus clones certainly initiated productive infection (peak viremia ~105 copies/mL and ~ 104 copies/mL for CXCR4- and CCR5-tropic viruses, respectively). However, virus production gradually became undetectable for all the animals tested (5–6- and 3–4-weeks post-infection for CXCR4- and CCR5-tropic viruses, respectively), indicating that the virus replication level at the initial phase was insufficient for viral persistence [44]. The peak level of virus production was significantly low relative to that by SIVmac. On one hand, Hatziioannou et al. have reported that they successfully generated a pathogenic molecular clone (stHIV-A19) upon infection to CD8+ cell-depleted pigtailed macaques [93, 94]. The molecular clone stHIV-A19 was obtained from a pathogenic viral swarm by repeated animal-to-animal (CD8+ cell-depleted pigtailed macaque) virus passage [94]. The virus level in the animal is maintained to be high only with the depletion treatment, being consistent with the appearance of AIDS. Clearly, this is the first study to describe the HIV-1 derivative molecular clone that can cause AIDS in animals. In vivo replication ability of various macaque-tropic HIV-1 derivative clones has been well summarized in the review [18].
5. Adaptation of macaque-tropic HIV-1 derivatives to macaque cells/individuals
Macaque-tropic HIV-1 derivatives are useful not only for the establishment of HIV-1-macaque infection models but also serve as model systems to investigate the inherent HIV-1 property, the high mutation/adaptation abilities. It is because one can analyze how HIV-1 adapts to replication-restrictive environments imposed by macaque cells. Studies on the adaptation process, for example, what mutations emerged in the genome of macaque-tropic HIV-1 derivatives in macaques as a new host and how the mutations altered the activity/function of viral proteins, would provide pivotal insights into determinants related to HIV-1 replication and pathogenesis. As described earlier, evading the potent restriction from intrinsic factors such as APOBEC3 and TRIM5 proteins in macaque cells was essential for the generation of macaque-tropic HIV-1 clones. The restriction from APOBEC3 proteins can be overcome by the replacement of the entire vif gene with the corresponding gene from SIVmac239. Meanwhile, various efforts had been made to construct HIV-1 capsids resistant to rhesus macaque TRIM5 proteins (Table 1) since the introduction of the entire gag-CA gene from SIVmac239 into the HIV-1 genome is lethal [98]. Of such constructs, SCA was generated by the introduction of SIVmac239 genomic region coding for a part of Gag-CA (1 to 204 amino acid residues), not a full-length, into the HIV-1 genome and then acquired some adaptive mutations that had emerged through adaptation in cells (Table 1) [33]. Research was also performed to construct HIV-1 Gag-CA with minimal genetic modifications that confer resistance against rhesus macaque TRIM5 proteins. Bieniasz’s group made rhesus TRIM5-resistant HIV-1 clones with various Gag-CAs by the assisted evolution method that combined PCR mutagenesis and adaptation in rhesus TRIM5alpha-expressing cells [97]. Of the HIV-1 clones they made, a clone designated LNEIE (with mutations M10L, I91N, A92E, M96I, and G116E in Gag-CA) showed the best growth in PBMCs from rhesus macaques (Table 1). Aside from this, we sought to alter HIV-1 Gag-CA through the combination of amino acid sequence comparison and in silico comparative structural analysis between HIV-1 (NL4–3) and SIVmac239 Gag-CAs. Of the clones we generated, LSDQ (with mutations M94L, R98S, and G114Q in Gag-CA) was the best as tested for rhesus TRIM5alpha resistance and growth in rhesus macaque cells (Table 1) [43, 87, 96]. Interestingly, it has been shown that the acquisition of R97S (corresponding to R98S in HIV-1 NL4–3) in Gag-CA of TRIM5alpha-susceptible SIV clone contributes to evasion from rhesus TRIM5alpha restriction [99]. The residue R98 in Gag-CA would be one of major determinants for rhesus TRIM5alpha susceptibility. Rhesus TRIM5alpha resistance was enhanced for both LNEIE and LSDQ clones carrying distinct sets of substituted amino residues. This may be compatible with a model that dimeric TRIM5alpha proteins recognize and interact with retroviral capsid lattice rather than bind to viral capsid via targeting particular residues within Gag-CA [100, 101, 102, 103]. Investigation using macaque-tropic HIV-1 derivatives revealed that there are several ways to genetically modify HIV-1 Gag-CA to avoid restriction from rhesus TRIM5 proteins.
Gag-CA containing a loop between helices 4 and 5/a loop between helices 6 and 7 loop from SIVmac239, and amino acid substitutions (M94L, R98S, and G114Q)
Structure-guided Gag-CA mutation (Q110D) based on an adaptive mutation (Gag-CA G114E)
Adaptation experiments of our prototype macaque-tropic HIV-1 derivatives NL-DT5R and NL-DT562 in macaque cells led to the identification of a novel genomic region that can determine the Vif expression level. Initially, we found replication-enhancing adaptive mutations within integrase (Pol-IN) C-terminal domain (CTD) that frequently and reproducibly emerged during independent adaptation experiments (Table 2) [88, 104, 105]. Extensive virological and sequence analyses of these adaptive mutations showed: 1) viral growth potential can be altered by naturally occurring synonymous single-nucleotide mutations (nsSNMs) within the region surrounding identified adaptive mutations, 2) these identified nsSNMs result in variations in Vif expression levels, and thus, 3) variations in viral replication by the nsSNMs occur dependently on both Vif and APOBEC3G expression levels. Moreover, we found that changes in Vif expression levels were due to the effect of these nsSNMs on the splicing efficiency at the splicing acceptor 1 (SA1) within pol-IN CTD, which is the important site for vif mRNA production. It has been reported that there are splicing regulatory elements that affect vif production, that is, ESEVif, ESEM1/M2, G4 motif, ESS2b, ESE2b, and GI2–1 [109, 110, 111, 112, 113]. We newly identified several nsSNMs outside of these splicing regulatory elements that did vary viral replication potential via alteration in Vif expression levels and named the region where such nsSNMs were found as SA1D2prox [104, 105, 114]. More importantly, the results in these experiments indicate that HIV-1 can adapt itself to given environments only by acquiring nucleotide mutations, synonymous silent mutations.
In vitro and in vivo adaptation of macaque-tropic HIV-1 derivative clones.
Other adaptive mutations within Env-gp120 (L124F, N132K, G150R, I307V) have been identified (references [88, 107]).
Other adaptive mutations within Env-gp120 (L124F, F174L, P181T, G308E, G310R) have been identified Refs. [88, 107].
As expected, we found numerous growth-enhancing mutations in the Env proteins of the viral clones that were obtained from adaptation experiments in macaque cells using NL-DT5R/NL-DT562 (Table 2). Our in silico structural modeling revealed that the adaptive mutations in Env are located within the regions involved in the interaction with receptor CD4 and/or coreceptor CCR5 [88, 106, 107]. While most of the adaptive mutations identified in Env increased viral growth potentials in both macaque and human cells, viral replication of NL-DT562 carrying Env-gp120 G310R was enhanced only in macaque cells and was abolished in human cells by a drastic reduction in the interaction with human CD4 and CCR5 [108]. The analyses of adaptive mutations that emerged in Env led to detailed understanding of the interplay of Env with cellular receptor/coreceptors and of changes in the structure and function of Env by the mutations.
The stHIV-1 clone, which is currently the only AIDS-inducible clone, also has been shown to adapt to CD8-depleted macaque individuals (Table 2) [93, 94]. After the animal-to-animal adaptation using stHIV-1 clones carrying four distinct env genes and pigtailed macaques, the deletion of four amino acid residues in the Env-gp120 V3 region was found. This deletion within Env was shown to be associated with a change in the coreceptor usage from CCR5 to CXCR4 (called coreceptor switch) that is seen in HIV-1-infected patients [93]. In addition, stHIV-1 acquired two amino acid mutations in Vpu (one insertion and one substitution) during the animal-to-animal adaptation. The HIV-1 Vpu carrying these mutations gained antagonizing activity against the tetherin of pigtailed macaques [93]. A pathogenic molecular clone (stHIV-A19) harbored mutations in the Nef and Env regions during adaptation in CD8-depleted pigtailed macaques, whereas adaptive mutations found in the Gag-CA have been shown to enhance the resistance against a host restriction factor Mx2 [94]. Studies using pathogenic HIV-1 derivatives disclosed what changes are required for efficient viral replication and pathogenicity in macaques as a new host and what are their determinants in both virus and host.
The infection system consisting of HIV-1 derivative viruses and macaque hosts may sophisticatedly reflect, as an experimental model, the interaction between HIV-1 and its host humans. In an effort to establish HIV-1-infected macaque models, a noteworthy result was obtained that stHIV-1/stHIV-1-A19 could induce AIDS in CD8-depleted pigtailed macaques after some animal-to-animal adaptations. However, this is the only case that showed the pathogenicity of the HIV-1 derivative virus in macaques as fully described above. Furthermore, the viruses were unable to cause the disease in naïve animals. And, it is not clarified yet whether certain viral gene(s) and/or viral genomic region(s) are linked to viral pathogenicity. Totally, based on the experimental results obtained so far, we may predict that the pathogenic viruses such as some SIVs and SHIVs grew quite well at the early infection phase in individuals so as to obtain variations sufficient to persist in host individuals in the presence of strong host antiviral immunity. If the viruses could persist and maintain the critical set-point level in hosts, they might finally cause AIDS and AIDS-related complex. Further experimental studies are required to confirm this perspective.
It should be mentioned here as a virologically critically important matter that through the generation of macaque-tropic HIV-1 derivatives and a series of their infection experiments in macaque cells and individuals, we and others could learn how HIV-1 mutate and adapt itself to restrictive environments. Numerous synonymous and non-synonymous mutational changes in the HIV-1 genome that significantly affect viral replication and pathogenicity have been successfully identified. The identified alterations are closely linked to our understanding as to how HIV-1 replicates in host cells through modulating functional domains/regions/activity of its genome and proteins such as viral RNA splicing and viral proteins’ binding to cellular receptors and to some other replication-relevant cellular factors. Findings obtained using macaque-tropic HIV-1 derivatives have already greatly contributed to and would also play a significant role in understanding the HIV-1 biology.
We thank Yayoi Shono (Tokushima University) for experimental assistance. We also thank Kazuko Yoshida (Tokushima University) and Kyoko Inui (Tokushima University) for editorial and administrative assistance.
AA and MN conceived the idea and wrote a draft/the final manuscript. TaK, ND, BQL, and ToK reviewed it and discussed its content. All authors approved submission.
This work was supported in part by JSPS KAKENHI Grant Numbers 21K07042 to MN, 22K07102 to TaK, 21K08491 to ND, and 20K18484 to ToK, and by grants from the Takeda Science Foundation and The Uehara Memorial Foundation to TaK.
Our recent studies described in this chapter have been done in collaboration with the following researchers: Masaru Yokoyama, Osamu Kotani, Hironori Sato, Kei Miyakawa, and Akihide Ryo (National Institute of Infectious Diseases, Japan). We are indebted to these scientists for their critical contribution to our work. We also thank our all staffs in our department and the other institutions who have supported our work.
Many original articles reporting the scientifically new and important findings could not be cited due to the tremendous numbers of publications and the space limitations. We express our sincere regret over these omissions based on rather subjective considerations.
apolipoprotein B mRNA editing enzyme, catalytic polypeptide-like 3
stHIV-1
simian-tropic HIV-1
HIV-1mt
macaque-tropic HIV-1
HSIV
pigtailed macaque-tropic HIV-1
HIV-1rmt
rhesus macaque-tropic HIV-1
References
1.Goff SP. Retroviridae. In: Knipe DM, Howley PM, editors. Fields virology. 6th ed. Philadelphia PA: Lippincott Williams & Wilkins; 2013. pp. 1424-1473
2.Freed EO, Martin MA. Human immunodeficiency viruses: Replication. In: Knipe DM, Howley PM, editors. Fields virology. 6th ed. Philadelphia PA: Lippincott Williams & Wilkins; 2013. pp. 1502-1560
3.Kuritzkes DR, Koup RA. HIV-1: Pathogenesis, clinical manifestations, and treatment. In: Knipe DM, Howley PM, editors. Fields Virology. 6th ed. Philadelphia PA: Lippincott Williams & Wilkins; 2013. pp. 1561-1583
4.Evans DT, Elder JH, Desrosiers RC. Nonhuman Lentiviruses. In: Knipe DM, Howley PM, editors. Fields Virology. 6th ed. Philadelphia PA: Lippincott Williams & Wilkins; 2013. pp. 1584-1612
5.Victor GJ. Humanized mice for HIV and AIDS research. Current Opinion in Virology. 2016;19:56-64. DOI: 10.1016/j.coviro.2016.06.010
6.Marsden MD, Zack JA. Humanized mouse models for human immunodeficiency virus infection. Annual Reviews of Virology. 2017;4:393-412. DOI: 10.1146/annurev-virology-101416-041703
7.Gillgrass A, Wessels JM, Yang JX, Kaushic C. Advances in humanized mouse models to improve understanding of HIV-1 pathogenesis and immune responses. Frontiers in Immunology. 2021;11:617516. DOI: 10.3389/fimmu.2020.617516
8.Terahara K, Iwabuchi R, Tsunetsugu-Yokota Y. Perspectives on non-BLT humanized mouse models for studying HIV pathogenesis and therapy. Viruses. 2021;13:776. DOI: 10.3390/v13050776
9.Baroncini L, Bredl S, Nicole KP, Speck RF. The humanized mouse model: What added value does it offer for HIV research? Pathogens. 2023;12:608. DOI: 10.3390/pathogens12040608
10.Ambrose Z, KewalRamani VN, Bieniasz PD, Hatziioannou T. HIV/AIDS: In search of an animal model. Trends in Biotechnology. 2007;25:333-337. DOI: 10.1016/j.tibtech.2007.05.004
11.Nomaguchi M, Doi N, Kamada K, Adachi A. Species barrier of HIV-1 and its jumping by virus engineering. Reviews in Medical Virology. 2008;18:261-275. DOI: 10.1002/rmv.576
12.Nomaguchi M, Doi N, Fujiwara S, Adachi A. Macaque-tropic HIV-1 derivatives: A novel experimental approach to understand viral replication and evolution in vivo. In: Chang TL, editor. HIV-Host Interactions. London, UK, Rijeka, Croatia: IntechOpen; 2011. pp. 325-348. DOI: 10.5772/23624
14.Misra A, Thippeshappa R, Kimata JT. Macaques as model hosts for studies of HIV-1 infection. Frontiers in Microbiology. 2013;4:176. DOI: 10.3389/fmicb.2013.00176
15.Saito A, Akari H. Macaque-tropic human immunodeficiency virus type 1: Breaking out of the host restriction factors. Frontiers in Microbiology. 2013;4:187. DOI: 10.3389/fmicb.2013.00187
16.Garcia-Tellez T, Huot N, Ploquin MJ, Rascle P, Jacquelin B, Müller-Trutwin M. Non-human primates in HIV research: Achievements, limits and alternatives. Infection, Genetics and Evolution. 2016;46:324-332. DOI: 10.1016/j.meegid.2016.07.012
17.Nishimura Y, Martin MA. Of mice, macaques, and men: Broadly neutralizing antibody immunotherapy for HIV-1. Cell Host & Microbe. 2017;22:207-216. DOI: 10.1016/j.chom.2017.07.010
18.Thippeshappa R, Kimata JT, Kaushal D. Toward a macaque model of HIV-1 infection: Roadblocks, progress, and future strategies. Frontiers in Microbiology. 2020;11:882. DOI: 10.3389/fmicb.2020.00882
19.Shibata R, Sakai H, Kawamura M, Tokunaga K, Adachi A. Early replication block of human immunodeficiency virus type 1 in monkey cells. Journal of General Virology. 1995;76:2723-2730. DOI: 10.1099/0022-1317-76-11-2723
20.Kirchhoff F. Immune evasion and counteraction of restriction factors by HIV-1 and other primate lentiviruses. Cell Host & Microbe. 2010;8:55-67. DOI: 10.1016/j.chom.2010.06.004
21.Blanco-Melo D, Venkatesh S, Bieniasz PD. Intrinsic cellular defenses against human immunodeficiency viruses. Immunity. 2012;37:399-411. DOI: 10.1016/j.immuni.2012.08.013
22.Harris RS, Hultquist JF, Evans DT. The restriction factors of human immunodeficiency virus. Journal of Biological Chemistry. 2012;287:40875-40883. DOI: 10.1074/jbc.R112.416925
23.Malim MH, Bieniasz PD. HIV restriction factors and mechanisms of evasion. Cold Spring Harbor Perspectives in Medicine. 2012;2:a006940. DOI: 10.1101/cshperspect.a006940
24.Kirchhoff F. Is the high virulence of HIV-1 an unfortunate coincidence of primate lentiviral evolution? Nature Reviews Microbiology. 2009;7:467-476. DOI: 10.1038/nrmicro2111
25.Sharp PM, Hahn BH. Origins of HIV and the AIDS pandemic. Cold Spring Harbor Perspectives in Medicine. 2011;1:a006841. DOI: 10.1101/cshperspect.a006841
26.Sakai Y, Miyake A, Doi N, Sasada H, Miyazaki Y, Adachi A, et al. Expression of profiles of Vpx/Vpr proteins are co-related with the primate lentiviral lineage. Frontiers in Microbiology. 2016;7:1211. DOI: 10.3389/fmicb.2016.01211
27.Sakai Y, Doi N, Miyazaki Y, Adachi A, Nomaguchi M. Phylogenetic insights into the functional relationship between primate lentiviral reverse transcriptase and accessory proteins Vpx/Vpr. Frontiers in Microbiology. 2016;7:1655. DOI: 10.3389/fmicb.2016.01655
28.Kleinman AJ, Pandrea I, Apetrei C. So pathogenic or so what?-a brief overview of SIV pathogenesis with an emphasis on cure research. Viruses. 2022;14:135. DOI: 10.3390/v14010135
29.Shibata R, Kawamura M, Sakai H, Hayami M, Ishimoto A, Adachi A. Generation of a chimeric human and simian immunodeficiency virus infectious to monkey peripheral blood mononuclear cells. Journal of Virology. 1991;65:3514-3520. DOI: 10.1128/JVI.65.7.3514-3520.1991
30.Sakuragi S, Shibata R, Mukai R, Komatsu T, Fukasawa M, Sakai H, et al. Infection of macaque monkeys with a chimeric human and simian immunodeficiency virus. Journal of General Virology. 1992;73:2983-2987. DOI: 10.1099/0022-1317-73-11-2983
31.Li J, Lord CI, Haseltine W, Letvin NL, Sodroski J. Infection of cynomolgus monkeys with a chimeric HIV-1/SIVmac virus that expresses the HIV-1 envelope glycoproteins. Journal of Acquired Immune Deficiency Syndromes. 1988;1992(5):639-646
32.Uberla K, Stahl-Hennig C, Böttiger D, Mätz-Rensing K, Kaup FJ, Li J, et al. Animal model for the therapy of acquired immunodeficiency syndrome with reverse transcriptase inhibitors. Proceedings of the National Academy of Sciences of the United States of America. 1995;92:8210-8214. DOI: 10.1073/pnas.92.18.8210
33.Hatziioannou T, Princiotta M, Piatak M Jr, Yuan F, Zhang F, Lifson JD, et al. Generation of simian-tropic HIV-1 by restriction factor evasion. Science. 2006;314:95. DOI: 10.1126/science.1130994
34.Kamada K, Igarashi T, Martin MA, Khamsri B, Hatcho K, Yamashita T, et al. Generation of HIV-1 derivatives that productively infect macaque monkey lymphoid cells. Proceedings of the National Academy of Sciences of the United States of America. 2006;103:16959-16964. DOI: 10.1073/pnas.0608289103
35.Igarashi T, Iyengar R, Byrum RA, Buckler-White A, Dewar RL, Buckler CE, et al. Human immunodeficiency virus type 1 derivative with 7% simian immunodeficiency virus genetic content is able to establish infections in pig-tailed macaques. Journal of Virology. 2007;81:11549-11552. DOI: 10.1128/JVI.00960-07
36.Ishimatsu M, Suzuki H, Akiyama H, Miura T, Hayami M, Ido E. Construction of a novel SHIV having an HIV-1-derived protease gene and its infection to rhesus macaques: A useful tool for in vivo efficacy tests of protease inhibitors. Microbes and Infection. 2007;9:475-482. DOI: 10.1016/j.micinf.2007.01.005
37.Hatziioannou T, Ambrose Z, Chung NP, Piatak M Jr, Yuan F, Trubey CM, et al. A macaque model of HIV-1 infection. Proceedings of the National Academy of Sciences of the United States of America. 2009;106:4425-4429. DOI: 10.1073/pnas.0812587106
38.Nishimura Y, Shingai M, Willey R, Sadjadpour R, Lee WR, Brown CR, et al. Generation of the pathogenic R5-tropic simian/human immunodeficiency virus SHIVAD8 by serial passaging in rhesus macaques. Journal of Virology. 2010;84:4769-4781. DOI: 10.1128/JVI.02279-09
39.Saito A, Nomaguchi M, Iijima S, Kuroishi A, Yoshida T, Lee YJ, et al. Improved capacity of a monkey-tropic HIV-1 derivative to replicate in cynomolgus monkeys with minimal modifications. Microbes and Infection. 2011;13:58-64. DOI: 10.1016/j.micinf.2010.10.001
40.Thippeshappa R, Polacino P, Yu Kimata MT, Siwak EB, Anderson D, Wang W, et al. Vif substitution enables persistent infection of pig-tailed macaques by human immunodeficiency virus type 1. Journal of Virology. 2011;85:3767-3779. DOI: 10.1128/JVI.02438-10
41.Shingai M, Donau OK, Schmidt SD, Gautam R, Plishka RJ, Buckler-White A, et al. Most rhesus macaques infected with the CCR5-tropic SHIV(AD8) generate cross-reactive antibodies that neutralize multiple HIV-1 strains. Proceedings of the National Academy of Sciences of the United States of America. 2012;109:19769-19774. DOI: 10.1073/pnas.1217443109
42.Saito A, Nomaguchi M, Kono K, Iwatani Y, Yokoyama M, Yasutomi Y, et al. TRIM5 genotypes in cynomolgus monkeys primarily influence inter-individual diversity in susceptibility to monkey-tropic human immunodeficiency virus type 1. Journal of General Virology. 2013;94:1318-1324. DOI: 10.1099/vir.0.050252-0
43.Nomaguchi M, Yokoyama M, Kono K, Nakayama EE, Shioda T, Doi N, et al. Generation of rhesus macaque-tropic HIV-1 clones that are resistant to major anti-HIV-1 restriction factors. Journal of Virology. 2013;87:11447-11461. DOI: 10.1128/JVI.01549-13
44.Doi N, Miura T, Mori H, Sakawaki H, Koma T, Adachi A, et al. CXCR4- and CCR5-tropic HIV-1 clones are both tractable to grow in rhesus macaques. Frontiers in Microbiology. 2018;9:2510. DOI: 10.3389/fmicb.2018.02510
45.Rosenwirth B, ten Haaft P, Bogers WM, Nieuwenhuis IG, Niphuis H, Kuhn EM, et al. Antiretroviral therapy during primary immunodeficiency virus infection can induce persistent suppression of virus load and protection from heterologous challenge in rhesus macaques. Journal of Virology. 2000;74:1704-1711. DOI: 10.1128/jvi.74.4.1704-1711.2000
46.Hofman MJ, Higgins J, Matthews TB, Pedersen NC, Tan C, Schinazi RF, et al. Efavirenz therapy in rhesus macaques infected with a chimera of simian immunodeficiency virus containing reverse transcriptase from human immunodeficiency virus type 1. Antimicrobial Agents and Chemotherapy. 2004;48:3483-3490. DOI: 10.1128/AAC.48.9.3483-3490.2004
47.Ambrose Z, Boltz V, Palmer S, Coffin JM, Hughes SH, Kewalramani VN. In vitro characterization of a simian immunodeficiency virus-human immunodeficiency virus (HIV) chimera expressing HIV type 1 reverse transcriptase to study antiviral resistance in pigtail macaques. Journal of Virology. 2004;78:13553-13561. DOI: 10.1128/JVI.78.24.13553-13561.2004
48.North TW, Van Rompay KK, Higgins J, Matthews TB, Wadford DA, Pedersen NC, et al. Suppression of virus load by highly active antiretroviral therapy in rhesus macaques infected with a recombinant simian immunodeficiency virus containing reverse transcriptase from human immunodeficiency virus type 1. Journal of Virology. 2005;79:7349-7354. DOI: 10.1128/JVI.79.12.7349-7354.2005
49.Van Rompay KK, Johnson JA, Blackwood EJ, Singh RP, Lipscomb J, Matthews TB, et al. Sequential emergence and clinical implications of viral mutants with K70E and K65R mutation in reverse transcriptase during prolonged tenofovir monotherapy in rhesus macaques with chronic RT-SHIV infection. Retrovirology. 2007;4:25. DOI: 10.1186/1742-4690-4-25
50.Ambrose Z, Palmer S, Boltz VF, Kearney M, Larsen K, Polacino P, et al. Suppression of viremia and evolution of human immunodeficiency virus type 1 drug resistance in a macaque model for antiretroviral therapy. Journal of Virology. 2007;81:12145-12155. DOI: 10.1128/JVI.01301-07
51.Shao W, Kearney M, Maldarelli F, Mellors JW, Stephens RM, Lifson JD, et al. RT-SHIV subpopulation dynamics in infected macaques during anti-HIV therapy. Retrovirology. 2009;6:101. DOI: 10.1186/1742-4690-6-101
52.North TW, Higgins J, Deere JD, Hayes TL, Villalobos A, Adamson L, et al. Viral sanctuaries during highly active antiretroviral therapy in a nonhuman primate model for AIDS. Journal of Virology. 2010;84:2913-2922. DOI: 10.1128/JVI.02356-09
53.Kearney M, Spindler J, Shao W, Maldarelli F, Palmer S, Hu SL, et al. Genetic diversity of simian immunodeficiency virus encoding HIV-1 reverse transcriptase persists in macaques despite antiretroviral therapy. Journal of Virology. 2011;85:1067-1076. DOI: 10.1128/JVI.01701-10
54.Boltz VF, Ambrose Z, Kearney MF, Shao W, Kewalramani VN, Maldarelli F, et al. Ultrasensitive allele-specific PCR reveals rare preexisting drug-resistant variants and a large replicating virus population in macaques infected with a simian immunodeficiency virus containing human immunodeficiency virus reverse transcriptase. Journal of Virology. 2012;86:12525-12530. DOI: 10.1128/JVI.01963-12
55.Gavegnano C, Detorio M, Montero C, Bosque A, Planelles V, Schinazi RF. Ruxolitinib and tofacitinib are potent and selective inhibitors of HIV-1 replication and virus reactivation in vitro. Antimicrobial Agents and Chemotherapy. 2014;58:1977-1986. DOI: 10.1128/AAC.02496-13
56.North TW, Villalobos A, Hurwitz SJ, Deere JD, Higgins J, Chatterjee P, et al. Enhanced antiretroviral therapy in rhesus macaques improves RT-SHIV viral decay kinetics. Antimicrobial Agents and Chemotherapy. 2014;58:3927-3933. DOI: 10.1128/AAC.02522-14
57.Kearney MF, Anderson EM, Coomer C, Smith L, Shao W, Johnson N, et al. Well-mixed plasma and tissue viral populations in RT-SHIV-infected macaques implies a lack of viral replication in the tissues during antiretroviral therapy. Retrovirology. 2015;12:93. DOI: 10.1186/s12977-015-0212-2
58.Melody K, McBeth S, Kline C, Kashuba AD, Mellors JW, Ambrose Z. Low frequency of drug-resistant variants selected by long-acting rilpivirine in macaques infected with simian immunodeficiency virus containing HIV-1 reverse transcriptase. Antimicrobial Agents and Chemotherapy. 2015;59:7762-7770. DOI: 10.1128/AAC.01937-15
59.Akiyama H, Ramirez NP, Gibson G, Kline C, Watkins S, Ambrose Z, et al. Interferon-inducible CD169/siglec1 attenuates anti-HIV-1 effects of alpha interferon. Journal of Virology. 2017;91:e00972-e00917. DOI: 10.1128/JVI.00972-17
60.Luciw PA, Pratt-Lowe E, Shaw KE, Levy JA, Cheng-Mayer C. Persistent infection of rhesus macaques with T-cell-line-tropic and macrophage-tropic clones of simian/human immunodeficiency viruses (SHIV). Proceedings of the National Academy of Sciences of the United States of America. 1995;92:7490-7494. DOI: 10.1073/pnas.92.16.7490
61.Letvin NL, Montefiori DC, Yasutomi Y, Perry HC, Davies ME, Lekutis C, et al. Potent, protective anti-HIV immune responses generated by bimodal HIV envelope DNA plus protein vaccination. Proceedings of the National Academy of Sciences of the United States of America. 1997;94:9378-9383. DOI: 10.1073/pnas.94.17.9378
62.Shibata R, Igarashi T, Haigwood N, Buckler-White A, Ogert R, Ross W, et al. Neutralizing antibody directed against the HIV-1 envelope glycoprotein can completely block HIV-1/SIV chimeric virus infections of macaque monkeys. Nature Medicine. 1999;5:204-210. DOI: 10.1038/5568
63.Baba TW, Liska V, Hofmann-Lehmann R, Vlasak J, Xu W, Ayehunie S, et al. Human neutralizing monoclonal antibodies of the IgG1 subtype protect against mucosal simian-human immunodeficiency virus infection. Nature Medicine. 2000;6:200-206. DOI: 10.1038/72309
64.Barouch DH, Santra S, Schmitz JE, Kuroda MJ, Fu TM, Wagner W, et al. Control of viremia and prevention of clinical AIDS in rhesus monkeys by cytokine-augmented DNA vaccination. Science. 2000;290:486-492. DOI: 10.1126/science.290.5491.486
65.Walker LM, Sok D, Nishimura Y, Donau O, Sadjadpour R, Gautam R, et al. Rapid development of glycan-specific, broad, and potent anti-HIV-1 gp120 neutralizing antibodies in an R5 SIV/HIV chimeric virus infected macaque. Proceedings of the National Academy of Sciences of the United States of America. 2011;108:20125-20129. DOI: 10.1073/pnas.1117531108
66.Georgiev IS, Doria-Rose NA, Zhou T, Kwon YD, Staupe RP, Moquin S, et al. Delineating antibody recognition in polyclonal sera from patterns of HIV-1 isolate neutralization. Science. 2013;340:751-756. DOI: 10.1126/science.1233989
67.Shingai M, Nishimura Y, Klein F, Mouquet H, Donau OK, Plishka R, et al. Antibody-mediated immunotherapy of macaques chronically infected with SHIV suppresses viraemia. Nature. 2013;503:277-280. DOI: 10.1038/nature12746
68.Del Prete GQ , Eilers B, Moldt B, Keele BF, Estes JD, Rodriguez A, et al. Selection of unadapted, pathogenic SHIVs encoding newly transmitted HIV-1 envelope proteins. Cell Host & Microbe. 2014;16:412-418. DOI: 10.1016/j.chom.2014.08.003
69.Ko SY, Pegu A, Rudicell RS, Yang ZY, Joyce MG, Chen X, et al. Enhanced neonatal fc receptor function improves protection against primate SHIV infection. Nature. 2014;514:642-645. DOI: 10.1038/nature13612
70.Shingai M, Donau OK, Plishka RJ, Buckler-White A, Mascola JR, Nabel GJ, et al. Passive transfer of modest titers of potent and broadly neutralizing anti-HIV monoclonal antibodies block SHIV infection in macaques. The Journal of experimental medicine. 2014;211:2061-2074. DOI: 10.1084/jem.20132494
71.Klein F, Nogueira L, Nishimura Y, Phad G, West AP Jr, Halper-Stromberg A, et al. Enhanced HIV-1 immunotherapy by commonly arising antibodies that target virus escape variants. The Journal of Experimental Medicine. 2014;211:2361-2372. DOI: 10.1084/jem.20141050
72.Francica JR, Sheng Z, Zhang Z, Nishimura Y, Shingai M, Ramesh A, et al. Analysis of immunoglobulin transcripts and hypermutation following SHIV(AD8) infection and protein-plus-adjuvant immunization. Nature. Communications. 2015;6:6565. DOI: 10.1038/ncomms7565
73.Reeves RK, Li H, Jost S, Blass E, Li H, Schafer JL, et al. Antigen-specific NK cell memory in rhesus macaques. Nature Immunology. 2015;16:927-932. DOI: 10.1038/ni.3227
74.Gautam R, Nishimura Y, Pegu A, Nason MC, Klein F, Gazumyan A, et al. A single injection of anti-HIV-1 antibodies protects against repeated SHIV challenges. Nature. 2016;533:105-109. DOI: 10.1038/nature17677
75.Li H, Wang S, Kong R, Ding W, Lee FH, Parker Z, et al. Envelope residue 375 substitutions in simian-human immunodeficiency viruses enhance CD4 binding and replication in rhesus macaques. Proceedings of the National Academy of Sciences of the United States of America. 2016;113:E3413-E3422. DOI: 10.1073/pnas.1606636113
76.Nishimura Y, Gautam R, Chun TW, Sadjadpour R, Foulds KE, Shingai M, et al. Early antibody therapy can induce long-lasting immunity to SHIV. Nature. 2017;543:559-563. DOI: 10.1038/nature21435
77.Gautam R, Nishimura Y, Gaughan N, Gazumyan A, Schoofs T, Buckler-White A, et al. A single injection of crystallizable fragment domain-modified antibodies elicits durable protection from SHIV infection. Nature Medicine. 2016;24:610-616. DOI: 10.1038/s41591-018-0001-2
78.Madani N, Princiotto AM, Mach L, Ding S, Prevost J, Richard J, et al. A CD4-mimetic compound enhances vaccine efficacy against stringent immunodeficiency virus challenge. Nature Communications. 2018;9:2363. DOI: 10.1038/s41467-018-04758-9
79.Borducchi EN, Liu J, Nkolola JP, Cadena AM, Yu WH, Fischinger S, et al. Antibody and TLR7 agonist delay viral rebound in SHIV-infected monkeys. Nature. 2018;563:360-364. DOI: 10.1038/s41586-018-0600-6
80.Fan J, Liang H, Ji X, Wang S, Xue J, Li D, et al. CTL-mediated immunotherapy can suppress SHIV rebound in ART-free macaques. Nature Communications. 2019;10:2257. DOI: 10.1038/s41467-019-09725-6
81.Pegu A, Borate B, Huang Y, Pauthner MG, Hessell AJ, Julg B, et al. A meta-analysis of passive immunization studies shows that serum-neutralizing antibody titer associates with protection against SHIV challenge. Cell Host & Microbe. 2019;26:336-346.e3. DOI: 10.1016/j.chom.2019.08.014
82.Terrade G, Huot N, Petitdemange C, Lazzerini M, Orta Resendiz A, Jacquelin B, et al. Interests of the non-human primate models for HIV cure research. Vaccines (Basel). 2021;9:958. DOI: 10.3390/vaccines9090958
83.Liang B, Li H, Li L, Omange RW, Hai Y, Luo M. Current advances in HIV vaccine preclinical studies using Macaque models. Vaccine. 2019;37:3388-3399. DOI: 10.1016/j.vaccine.2019.04.094
84.Nomaguchi M, Doi N, Matsumoto Y, Sakai Y, Fujiwara S, Adachi A. Species tropism of HIV-1 modulated by viral accessory proteins. Frontiers in Microbiology. 2012;3:267. DOI: 10.3389/fmicb.2012.00267
85.Fujita M, Yoshida A, Miyaura M, Sakurai A, Akari H, Koyama AH, et al. Cyclophilin A-independent replication of a human immunodeficiency virus type 1 isolate carrying a small portion of the simian immunodeficiency virus SIV(MAC) gag capsid region. Journal of Virology. 2001;75:10527-10531. DOI: 10.1128/JVI.75.21.10527-10531.2001
86.Kamada K, Yoshida A, Khamsri B, Piroozmand A, Yamashita T, Uchiyama T, et al. Construction of gag-chimeric viruses between HIV-1 and SIVmac that are capable of productive multi-cycle infection. Microbes and Infection. 2006;8:1075-1081. DOI: 10.1016/j.micinf.2005.11.006
87.Nomaguchi M, Yokoyama M, Kono K, Nakayama EE, Shioda T, Saito A, et al. Gag-CA Q110D mutation elicits TRIM5-independent enhancement of HIV-1mt replication in macaque cells. Microbes and Infection. 2013;15:56-65. DOI: 10.1016/j.micinf.2012.10.013
88.Nomaguchi M, Doi N, Fujiwara S, Saito A, Akari H, Nakayama EE, et al. Systemic biological analysis of the mutations in two distinct HIV-1mt genomes occurred during replication in macaque cells. Microbes and Infection. 2013;15:319-328. DOI: 10.1016/j.micinf.2013.01.005
89.Kawamura M, Ishizaki T, Ishimoto A, Shioda T, Kitamura T, Adachi A. Growth ability of human immunodeficiency virus type 1 auxiliary gene mutants in primary blood macrophage cultures. Journal of General Virology. 1994;75:2427-2431. DOI: 10.1099/0022-1317-75-9-2427
90.Adachi A, Gendelman HE, Koenig S, Folks T, Willey R, Rabson A, et al. Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone. Journal of Virology. 1986;59:284-291. DOI: 10.1128/JVI.59.2.284-291.1986
91.Kamada K, Yamashita T, Hatcho K, Adachi A, Nomaguchi M. Evasion from CypA- and APOBEC-mediated restrictions is insufficient for HIV-1 to efficiently grow in simian cells. Microbes and Infection. 2009;11:164-171. DOI: 10.1016/j.micinf.2008.11.002
92.Kuroishi A, Saito A, Shingai Y, Shioda T, Nomaguchi M, Adachi A, et al. Modification of a loop sequence between alpha-helices 6 and 7 of virus capsid (CA) protein in a human immunodeficiency virus type 1 (HIV-1) derivative that has simian immunodeficiency virus (SIVmac239) vif and CA alpha-helices 4 and 5 loop improves replication in cynomolgus monkey cells. Retrovirology. 2009;6:70. DOI: 10.1186/1742-4690-6-70
93.Hatziioannou T, Del Prete GQ , Keele BF, Estes JD, McNatt MW, Bitzegeio J, et al. HIV-1-induced AIDS in monkeys. Science. 2014;344:1401-1405. DOI: 10.1126/science.1250761
94.Schmidt F, Keele BF, Del Prete GQ , Voronin D, Fennessey CM, Soll S, et al. Derivation of simian tropic HIV-1 infectious clone reveals virus adaptation to a new host. Proceedings of the National Academy of Sciences of the United States of America. 2019;116:10504-10509. DOI: 10.1073/pnas.1818059116
95.Thippeshappa R, Ruan H, Wang W, Zhou P, Kimata JT. A variant macaque-tropic human immunodeficiency virus type 1 is resistant to alpha interferon-induced restriction in pig-tailed macaque CD4+ T cells. Journal of Virology. 2013;87:6678-6692. DOI: 10.1128/JVI.00338-13
96.Nomaguchi M, Nakayama EE, Yokoyama M, Doi N, Igarashi T, Shioda T, et al. Distinct combinations of amino acid substitutions in N-terminal domain of Gag-capsid afford HIV-1 resistance to rhesus TRIM5α. Microbes and Infection. 2014;16:936-944. DOI: 10.1016/j.micinf.2014.08.017
97.Soll SJ, Wilson SJ, Kutluay SB, Hatziioannou T, Bieniasz PD. Assisted evolution enables HIV-1 to overcome a high TRIM5α-imposed genetic barrier to rhesus macaque tropism. PLoS Pathogens. 2013;9:e1003667. DOI: 10.1371/journal.ppat.1003667
98.Dorfman T, Göttlinger HG. The human immunodeficiency virus type 1 capsid p2 domain confers sensitivity to the cyclophilin-binding drug SDZ NIM 811. Journal of Virology. 1996;70:5751-5757. DOI: 10.1128/JVI.70.9.5751-5757.1996
99.McCarthy KR, Schmidt AG, Kirmaier A, Wyand AL, Newman RM, Johnson WE. Gain-of-sensitivity mutations in a Trim5-resistant primary isolate of pathogenic SIV identify two independent conserved determinants of Trim5α specificity. PLoS Pathogens. 2013;9:e1003352. DOI: 10.1371/journal.ppat.1003352
100.Ganser-Pornillos BK, Chandrasekaran V, Pornillos O, Sodroski JG, Sundquist WI, Yeager M. Hexagonal assembly of a restricting TRIM5alpha protein. Proceedings of the National Academy of Sciences of the United States of America. 2011;108:534-539. DOI: 10.1073/pnas.1013426108
101.Biris N, Yang Y, Taylor AB, Tomashevski A, Guo M, Hart PJ, et al. Structure of the rhesus monkey TRIM5α PRYSPRY domain, the HIV capsid recognition module. Proceedings of the National Academy of Sciences of the United States of America. 2012;109:13278-13283. DOI: 10.1073/pnas.1203536109
102.Yang H, Ji X, Zhao G, Ning J, Zhao Q , Aiken C, et al. Structural insight into HIV-1 capsid recognition by rhesus TRIM5α. Proceedings of the National Academy of Sciences of the United States of America. 2012;109:18372-18377. DOI: 10.1073/pnas.1210903109
103.Biris N, Tomashevski A, Bhattacharya A, Diaz-Griffero F, Ivanov DN. Rhesus monkey TRIM5α SPRY domain recognizes multiple epitopes that span several capsid monomers on the surface of the HIV-1 mature viral core. Journal of Molecular Biology. 2013;425:5032-5044. DOI: 10.1016/j.jmb.2013.07.025
104.Nomaguchi M, Miyake A, Doi N, Fujiwara S, Miyazaki Y, Tsunetsugu-Yokota Y, et al. Natural single-nucleotide polymorphisms in the 3′ region of the HIV-1 pol gene modulate viral replication ability. Journal of Virology. 2014;88:4145-4160. DOI: 10.1128/JVI.01859-13
105.Nomaguchi M, Doi N, Sakai Y, Ode H, Iwatani Y, Ueno T, et al. Natural single-nucleotide variations in the HIV-1 genomic SA1prox region can alter viral replication ability by regulating Vif expression levels. Journal of Virology. 2016;90:4563-4578. DOI: 10.1128/JVI.02939-15
106.Doi N, Yokoyama M, Koma T, Kotani O, Sato H, Adachi A, et al. Concomitant enhancement of HIV-1 replication potential and neutralization-resistance in concert with three adaptive mutations in Env V1/C2/C4 domains. Frontiers in Microbiology. 2019;10:2. DOI: 10.3389/fmicb.2019.00002
107.Yokoyama M, Nomaguchi M, Doi N, Kanda T, Adachi A, Sato H. In silico analysis of HIV-1 Env-gp120 reveals structural bases for viral adaptation in growth-restrictive cells. Frontiers in Microbiology. 2016;7:110. DOI: 10.3389/fmicb.2016.00110
108.Koma T, Yokoyama M, Kotani O, Doi N, Nakanishi N, Okubo H, et al. Species-specific valid ternary interactions of HIV-1 Env-gp120, CD4, and CCR5 as revealed by an adaptive single-amino acid substitution at the V3 loop tip. Journal of Virology. 2021;95:e0217720. DOI: 10.1128/JVI.02177-20
109.Kammler S, Otte M, Hauber I, Kjems J, Hauber J, Schaal H. The strength of the HIV-1 3′ splice sites affects rev function. Retrovirology. 2006;3:89. DOI: 10.1186/1742-4690-3-89
110.Exline CM, Feng Z, Stoltzfus CM. Negative and positive mRNA splicing elements act competitively to regulate human immunodeficiency virus type 1 vif gene expression. Journal of Virology. 2008;82:3921-3931. DOI: 10.1128/JVI.01558-07
111.Mandal D, Exline CM, Feng Z, Stoltzfus CM. Regulation of vif mRNA splicing by human immunodeficiency virus type 1 requires 5′ splice site D2 and an exonic splicing enhancer to counteract cellular restriction factor APOBEC3G. Journal of Virology. 2009;83:6067-6078. DOI: 10.1128/JVI.02231-08
112.Widera M, Erkelenz S, Hillebrand F, Krikoni A, Widera D, Kaisers W, et al. An intronic G run within HIV-1 intron 2 is critical for splicing regulation of vif mRNA. Journal of Virology. 2013;87:2707-2720. DOI: 10.1128/JVI.02755-12
113.Brillen AL, Walotka L, Hillebrand F, Müller L, Widera M, Theiss S, et al. Analysis of competing HIV-1 splice donor sites uncovers a tight cluster of splicing regulatory elements within exon 2/2b. Journal of Virology. 2017;91:e00389-e00317. DOI: 10.1128/JVI.00389-17
114.Nomaguchi M, Doi N, Yoshida T, Koma T, Adachi S, Ode H, et al. Production of HIV-1 vif mRNA is modulated by natural nucleotide variations and SLSA1 RNA structure in SA1D2prox genomic region. Frontiers in Microbiology. 2017;8:2542. DOI: 10.3389/fmicb.2017.02542
Written By
Takaaki Koma, Naoya Doi, Bao Quoc Le, Tomoyuki Kondo, Akio Adachi and Masako Nomaguchi
Submitted: 05 August 2023Reviewed: 16 August 2023Published: 30 September 2023
 Open access peer-reviewed chapter
Open access peer-reviewed chapter