 Open access peer-reviewed chapter
Open access peer-reviewed chapter
Abstract
Agriculture is one of the factors contributing to the global depletion of natural resources. A total of 228 bird species were recorded in the savanna of Serengeti National Park and 146 bird species in agricultural areas. The richness of bird species endemic to the Serengeti ecosystem, regional and Africa-wide geographical ranges were all higher inside Serengeti National Park compared to agricultural land. However, a greater proportion of endemic bird species dropped out in agriculture compared to the proportions of species with wider geographic ranges. With respect to the trophic level in agriculture, there was a greater proportional decrease of species (compared to savanna) at the predator trophic level (vertebrate feeders) than at the insectivore and herbivore trophic levels. Therefore, higher feeding levels were less able to tolerate changes in habitat structure due to agriculture than species at lower feeding levels. These results suggest that Park managers should encourage through education and participation of local communities on the importance of improving their agricultural habitats by promoting the regeneration of patches of native vegetation so that more vulnerable species, such as endemics, and raptors can exist there.
Keywords
- conservation
- climate change
- birds’ distribution
- disturbance
- environmental drivers
- human modified habitats
1. Introduction
Agriculture is one of the factors that contribute to the global depletion of natural resources [1, 2, 3, 4]. Land clearing and encroachment are the main contributors to the decline of wildlife and local species extinction worldwide [5, 6, 7, 8]. The increase in human population has resulted in the rise in demand for land for settlement and food to meet the body requirements of people. Over the last four decades, areas adjacent to Serengeti National Park have experienced huge population growth [9, 10]. The period between 1957 and 1977 recorded the highest rate of population increase, some 10% per annum. Of this, only 3.4% was due to natural increase while the rest was due to immigration [9, 10, 11]. The current population in the seven wards to the west of the park is over two million with an annual growth rate exceeding the national average of 2.9% [11, 12].
Human activities such as overgrazing, deforestation, bushfires, mining, urbanization, and cultivation as well as climate change have aggravated the ability of bird species to disperse into new habitats [13, 14]. These two factors, therefore, have resulted in degradation or modification of habitat structure with consequent decline of avian food resources.
Tanzania has reserved about 40% of its land as protected areas [15]; these are distributed throughout the country and are largely located adjacent to rural communities. The majority of these rural communities depend on crops and livestock for their livelihoods; thus, a considerable number of people have settled around buffer zones surrounding protected areas where they believe that the land is fertile and there is sufficient grazing land [9, 16, 17]. Kideghesho et al. [11] showed that there was a 52% increase in livestock from 175,680 to 266,624 from 1990 to 2011, respectively within areas adjacent to the western Serengeti National Park. At the same time, the expansion of arable land and settlement in villages has led to the shrinkage of grazing land for livestock [9, 10]. This density is now well above the livestock carrying capacity, which was already exceeded a decade ago [11, 18]. Livestock numbers have been reported to increase at the peripheral of protected areas likely to impact severely on the native vegetation and expected to influence wildlife daily activities in the park.
1.1 Interaction of environmental change with agriculture areas
In tropical regions, the main climate variable is rainfall rather than the temperature which is the dominant variable at higher latitudes. Rain in the Serengeti ecosystem typically falls in a bimodal pattern, with the long rains during March–May and the short rains during November–December [19]. However, the rains can fuse into one long period, particularly in the north, or the short rains can fail entirely, especially in the southeast [19]. Most importantly the Serengeti ecosystem has a strong gradient in the total annual rainfall from 500 mm in the southeast to 1100 mm in the north and west [19]. The wettest part of the ecosystem is in the agricultural region of northwest Serengeti National Park around Mugumu and Tarime [20, 21]. If hypothetically the Serengeti National Park becomes drier than the savanna bird community within, it would move up the rainfall gradient into the agricultural region. Although most bird species avoid agricultural areas, nearly 30% of all birds regularly or occasionally use such habitats [22, 23], often providing important ecosystem services, such as pest control, pollination, seed dispersal, and nutrient deposition [22, 24].
The main environmental driver in Serengeti National Park is rainfall, with temperature being of lesser importance, although it may still have a subsidiary influence [25, 26]. Animal communities in Serengeti National Park respond to changes in rainfall; if conditions become drier communities will move toward wetter regions. To understand how climate change might affect the Serengeti National Park through changes in rainfall the bird community was taken as a case study. Within the Serengeti National Park, birds are constrained in how far they can move along the rainfall gradient in response to climate change because there are barriers from the human settlement at the edge of the park boundary in the north and west. If conditions become drier, birds will move toward wetter areas which now are severely modified by agriculture. Trees are largely absent, native grasses are replaced by monocultures of domestic crops, and shrubs become abundant along hedgerows. As a result bird species have responded differently to this habitat modification in the village lands [1, 23, 27, 28].
Climate change has already affected bird distributions in the northern temperate regions such as Europe and North America [29, 30, 31, 32, 33]. Far less is known about how climate can affect the distribution of birds in the tropics. It has been postulated that savanna woodlands make up most of Africa’s tropical tree cover, but savannas are widely thought to be declining rapidly under both direct human impacts and indirect human-induced climate change [34, 35, 36]. With climate change, species’ geographic distributions are expected to change. Bird species may become locally extinct in existing protected areas that are no longer environmentally suitable but cannot move into more suitable areas because of habitat modification by humans [36, 37, 38]. Therefore, the conservation of vulnerable bird species may require new areas that are currently both unprotected and climatically unsuitable but could become suitable later [39, 40, 41, 42, 43].
1.2 The importance of disturbance in ecological communities
The number of relatively undisturbed ecosystems in the world is decreasing rapidly and living organisms are confronted with man-made challenges, including climate change and habitat destruction. By the year 2050, 15–37% of existing animal and plant species on earth are predicted to become extinct [37, 44, 45] and half of all species on earth may experience extinction by 2100 [46]. It is important to predict how current man-made challenges will affect key species in ecosystems, so that, if possible, steps can be taken to ameliorate these threats.
There is much scientific concern that the loss of living organisms will reduce the capacity of our ecosystems to provide important services for human populations. Examples of such services are primary production, nutrient cycling, mineral cycling, and pollution control [47, 48, 49, 50]. In addition, the consequences of species extinction on ecosystems may not be detected until it is too late to reverse the loss [51]. Some ecosystems respond to disturbances by losing a few species at first but many more later on; so a need for conservation authorities management to reverse species loss would benefit from early detection [51, 52]. Responses of biota including changes in physiology, productivity, and growth have been caused by human activities and climate change acting upon their habitat [53, 54], and changes in species distribution and abundance due to migration and range shifts have also been caused by habitat loss [55].
Tolerance limits are usually defined in terms of environmental parameters such as temperature, rainfall, altitude, or habitats but they can be complex involving a mixture of all these plus tolerance of predation [56]. Tolerance limits are situations in which living organisms cannot distribute, survive, grow, or reproduce under limited environmental factors [56]. Distribution of organisms can be endemic (with narrow distribution), regional, or Africa-wide ranging depending on the environmental factors of a particular area.
There has been much study of the pattern of wide-ranging or narrow-ranging species in terms of wide tolerance or narrow tolerance of these factors, in order to understand what causes areas of high diversity in the world [57, 58, 59]. This study uses the variation in distribution and hence tolerance limits to predict which species are most susceptible to man-made disturbances in the form of agricultural modification of habitat.
Agriculture around protected areas is one of the major threats to wildlife conservation in Tanzania [1, 9, 60, 61, 62]. Cultivation and livestock husbandry around national parks and game reserves affect natural resource conservation in Tanzania [63, 64]. The confinement of high densities of livestock on village lands causes overgrazing, soil erosion, and siltation of water bodies ultimately causing ecological imbalance, which interferes with ecological resilience [9, 10, 22, 65, 66]. Due to inadequate and poor quality pasture on these village lands, the livestock owners in some villages of western Serengeti National Park have been pressing the government to legalize access to critical grazing and watering points in Grumeti, Ikorongo, and Kijereshi Game Reserves [11, 66]. These villagers, however, are continuing to use the areas illegally in order to survive. Little information is available to describe how birds with different tolerance limits or feeding levels are affected by these habitat disturbances from agricultural modification.
The main objective of the study was to examine the effects of agriculture on the trophic cascade in savanna bird communities. The study had two specific objectives: First, to compare how species of birds with narrow ecological tolerance limits respond to anthropogenic disturbance of habitat (agriculture) in comparison to those with wide tolerance limits (Africa-wide species). Second, to compare how savanna bird species at different trophic levels respond to disturbance in human-modified habitats such as agriculture.
We hypothesized that endemic bird species with narrow ecological tolerance limits lose a greater proportion of species through agricultural modification of habitats than do bird species with wide ecological tolerance limits. Also, bird species in higher feeding guilds lose a greater proportion of species through agricultural modification than do species at lower feeding guilds.
2. Materials and methods
2.1 Study area
The study was conducted in Serengeti National Park (14,763 km2) located between 33°50′–35°20′ E and 1°28′–3°17′ S (Figure 1). The park occupies an extensive upland area varying in elevation from 1162 m at Speke’s Gulf to 1860 m above sea level in the northeast [67]. Serengeti National Park is characterized by grassland, savanna, closed canopy riverine forest, rocky outcrops, rivers, and alkaline lakes (Figure 1).
Figure 1.
Map of Serengeti National Park showing vegetation types. Black dots show bird sampling plots (Map provided by the Tanzania Wildlife Research Institute-GIS, 2023).
The climate shows a relatively constant monthly mean and maximum temperature of 27–28°C taken daily in the morning (9:00 AM) and afternoon (3:00 PM) at Seronera [19]. The minimum temperature varies from 16°C in the hot months of October–March to 13°C during May–August. Rain typically falls in a bimodal pattern, with the long rains during March–May and the short rains in November–December [19]. However, the rains can fuse into one long period, particularly in the north, or the short rains can fail entirely, especially in the southeast of the park [67].
2.2 Bird survey in agricultural and savanna habitats
Data in Serengeti National Park and in agricultural areas immediately outside the Park were obtained by similar methods to allow comparisons of the bird communities in the two study areas. The bird community was sampled along road transects in native savanna and through villages. Both transect and point count methods were used to record bird species, their numbers, and their habitats. Transects in both savanna and in agriculture were established using roads. A total of five set points of 100 m × 100 m along each transect in savanna and agriculture was established, marked by using the Global Positioning System (GPS) Garmin 12 XL made in Taiwan. Points along transects were spaced 2 km apart.
At the five set points established along each transect birds were recorded by sight or by hearing the call for 10 minutes. Transects were driven in the morning from 0615 to 1100 hours. The vehicle traveled at 3 km per hour and observers on each side of the vehicle recorded birds within 50 m on either side of the vehicle. For the agricultural area, only data from those parts of transects that fell within cultivation or habitations were used in this analysis. Cultivation comprised small fields of crops, hedgerows, and small areas used for grazing, usually 2–3 ha in size. Cultivation extended from 0.2 to 1 km on either side of the transects.
Three transects were established in agricultural areas. One was from Fort Ikoma through Mugumu town to Tabora B gate (80 km), a second ran from Mugumu Town to the Park boundary at Nyamburi village (40 km), and the third was from Fort Ikoma park headquarters to Isenye village (60 km). Transects were repeated every year between June 1997 and December 2011, the same observers who worked on the long-term project of the Serengeti Biodiversity Program conducted the surveys. Data from all transects were pooled across years to construct the sample for agricultural land.
Six transects within the Serengeti National Park provided the sample for the native savanna. These were: (i) Along the Mbalageti River (40 km), (ii) Along the southern edge of the woodlands where they meet the plains near Seronera (20 km), (iii) In the central woodlands near Banagi hill (20 km), (iv) In the northeast around Togoro and Lobo (20 km), (v) In the northwest around Tabora (40 km), and (vi) In the far west around Ndabaka (40 km). These were a subset of a much larger sample of permanent road transects surveyed by the Serengeti Biodiversity Program. These transects were counted at intervals of six months from 1997 to December 2011. Data were summed across years for agricultural areas.
3. Data analysis
In comparing species richness between agricultural areas and the savanna of the Serengeti National Park the data were divided into two ways. First, species were examined according to differences in their geographical range within Africa as published in Sinclair and Ryan [68] and the seven volumes of the handbook “The Birds of Africa” [69, 70]. Three groups of geographical ranges were considered: (i) local endemic species found only in the Serengeti ecosystem, (ii) regional species confined to East Africa, and (iii) Africa-wide species (see Appendix 6).
Secondly, bird species were divided according to their feeding guilds, Simberloff and Dayan [71] defined a feeding guild as a group of animals that feeds on similar food types. A feeding guild can be a subset of a trophic level. Birds that feed on other birds, rodents, reptiles, amphibians, and fish were classified as vertebrate feeders (e.g. raptors, owls, and herons). Insectivorous birds were defined (for convenience) in this study as those that feed on arthropods, which includes insects, and also non-insect arthropods such as spiders and centipedes. Birds that feed on herbs, seeds, and fruits were all grouped in a feeding guild of herbivores (see Appendix 6).
Bird data were tested for normality using Kolmogorov-Smirnov (KS) test [72]. The data were not normally distributed even after transformation. Therefore a comparison of birds between agricultural land and the natural savanna of the Serengeti National Park was compared using the non-parametric test [72]. Bird species richness was estimated using the method of rarefaction given in the program of ecological methodology version 7.1 [73]. Because the sample within the protected area was much larger than that for agricultural areas the ‘rarefaction’ method was used to accommodate the difference in sample sizes. Rarefaction is a statistical method for estimating the species richness in a given sample of individuals. It allows two samples of different sizes, representing different sampling efforts, to be compared [74]. For example, within a given category of data, such as vertebrate feeders in agriculture, the program estimates the probable number of species in different sample sizes say 50, 100, 500 individuals etc. up to the total sample size. The same procedure was used for the sample from savanna with which the data for agriculture are being compared. For all tests
Shannon-Wiener diversity (
4. Results
4.1 Agricultural effects on the resilience of the bird community
A total of 228 bird species were recorded in the savanna of Serengeti National Park and 146 bird species in agricultural areas. Estimates for species in the three range categories are presented in Figure 2. In agriculture, a sample of 3233 individuals was obtained which comprised of 88 species. In the savanna, a total of 150 species were recorded but for the same sample size as that for agriculture 97 species were estimated using Rarefaction. The rarefaction curve for Africa-wide birds showed higher species richness inside Serengeti National Park than in agricultural land. Initially, with the small sample size Africa-wide species in agriculture were greater in number than for the same sample size in the Serengeti National Park. However, as sample size increased the number of species in agriculture leveled out while those in Serengeti National Park continued to increase. Although there were more species in Serengeti National Park the difference was not great; some 9.3% of species were not observed in agriculture. The broad habitat niches of these species allowed them to tolerate the habitat modifications from agriculture.
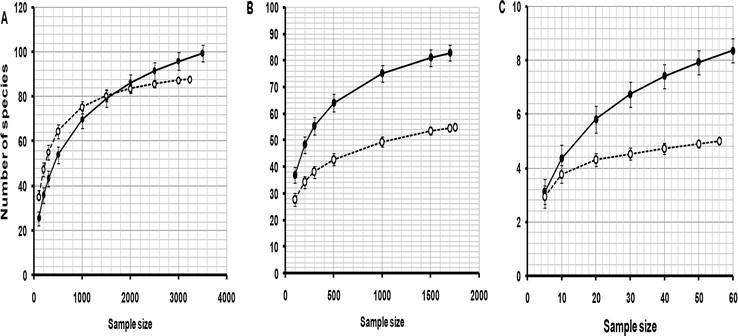
Figure 2.
Rarefaction curves for the number of bird species relative to the number of individuals counted along transects in agriculture (open circles) and the adjacent native savanna of Serengeti National park (closed circles) with respect to their geographical range in Africa. A. Africa-wide species. B. Regional species with an East African distribution. C. Local endemic species confined to the Serengeti ecosystem or areas close by. Vertical lines are standard error.
For regional bird species in East Africa, a total of 55 species were sampled from the agricultural areas. The total number of bird species recorded in the Savanna was 109 but for the same sample as that for agriculture, the sample size in Savanna was reduced to 83 species. For local endemic bird species, 5 species were recorded in agricultural areas and 8 in Savanna for the same sample size but the total number in Savanna was 17. From rarefaction curves in Figure 2, the number of savanna bird species continued to increase as the sample size of individuals increased whereas the number of endemics in agriculture reached an asymptote. This result indicates that if a larger sample size had been obtained in agriculture there would be even larger differences in the number of endemic species. The rarefaction curves show that the more restricted the geographical range of species the greater the loss of species when habitats were modified by agriculture. Thus, the proportion of species not observed in agriculturally modified habitats when compared to savanna was 9% for Africa-wide species, 34% for regional species, and 44% for local endemic species.
4.2 Effects of agriculture on the trophic cascade and food web stability
A comparison of bird diversity using the Shannon-Wiener index shows a difference between Serengeti National Park and agricultural areas. Herbivores (
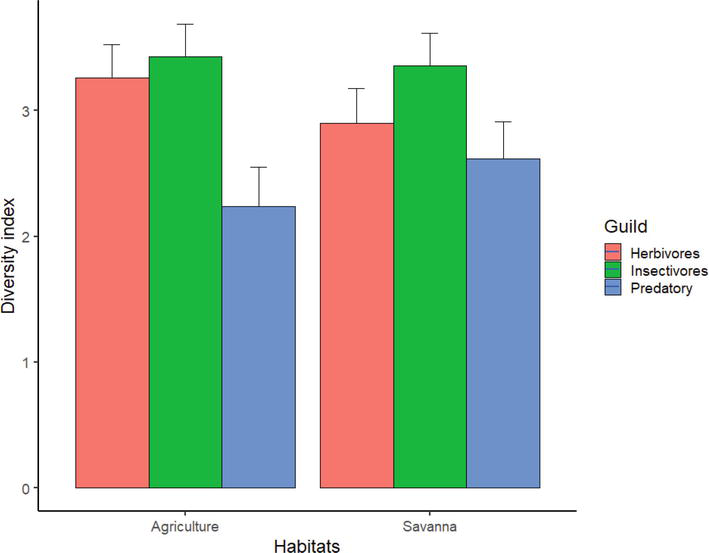
Figure 3.
Diversity indices for bird species observed and categorized according to their feeding guilds along transects in agricultural land and the adjacent native Savanna of Serengeti National Park. Vertical lines represent standard errors.
Rarefaction estimates for species in the three feeding guilds are presented in Figure 4. For herbivores, a total of 51 species were recorded from a sample of 2350. For the same sample size in Savanna, there were 60 species estimated but the total was 79 species in all from the total sample. Insectivores are the dominant guild in the avifauna and so more species were recorded for this group than for herbivores or vertebrate feeders. In agriculture, the total sample of 1983 produced 77 insectivores observed species. For the same sample size in the savanna, rarefaction estimated 90 species although the total number of species was 145 for the complete sample. There were 13 species of vertebrate feeders from a sample of 81 in agriculture. The equivalent number of species in the savanna was 21 from the same sample size with a total of 47 species recorded from the complete savanna sample. In summary, agriculture had 15% fewer herbivore species, 14% fewer insectivore species, and 38% fewer vertebrate feeders than those in savannah (Figure 4).
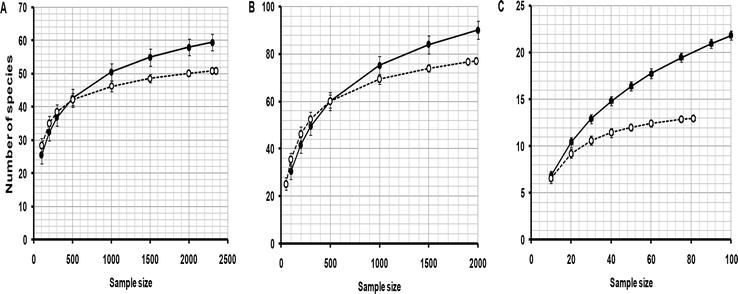
Figure 4.
Rarefaction curves for the number of bird species relative to the number of individuals counted along transects in agriculture (open circles) and the adjacent native savanna of Serengeti National Park (closed circles) with respect to their feeding guild. A. Herbivore species. B. Insectivore species. C. Birds that feed on vertebrates. Vertical lines represent standard errors.
Differences in the abundances of birds in agriculture (
5. Discussion
This chapter examines the effects of disturbance on the distribution of bird communities in the savanna, and in particular the effects of habitat modification through agriculture. In the introduction, it was predicted that if wide-ranging species had broad habitat tolerance then such species should be able to tolerate agricultural modifications better than bird species with small ranges and narrow habitat requirements. Secondly, it was predicted that since the effects of disturbance are magnified up the trophic food chain [77] agriculture should have greater effects through the loss of species on higher trophic levels. These two hypotheses are discussed in detail below.
5.1 Agricultural effects on the resilience of the bird communities
These results indicate that birds in agricultural land may be affected by human activities through the planting of crops, the change in the herbaceous and shrub communities, and the removal of trees, all of which birds use for foraging, nesting, and perching. In general, the evidence supports the hypothesis that major changes to habitats such as agricultural modification causes bird communities to change, with species with broad niches and wide tolerance limits being more able to withstand the change than those with narrow ecological niches. Using the range of a species in Africa as the indicator of wide or narrow tolerance limits it was predicted that a greater proportion of local endemics would be lost in agricultural habitat than would species with broader habitat preferences, the Africa-wide species. The results are consistent with this hypothesis.
Local bird species with narrow geographical ranges have a lower tolerance to habitat change [78] and therefore are less able to survive in agricultural land compared to Africa-wide species with wide geographical ranges and ecological tolerance. The greater decrease of local endemics bird species when habitats changed from natural Savanna to agriculture was probably due to a combination of changes in habitat structure in agricultural land, such as loss of trees, monocultures of crops, and perhaps more exotic plants which do not provide suitable habitat for their survival. Habitat loss, overhunting, and animal disease could be factors contributing to the greater loss of local endemic birds in agricultural land. These factors promote protected area isolation by restricting birds’ movement and dispersal.
These findings are consistent with other studies. Waltert et al. [79] reported a decline of endemic birds; they observed species composition gradually changed as the habitat transformed from natural forest to agricultural land in Sulawesi, Indonesia. Bolwig et al. [80] recorded biodiversity change in the forest under agricultural intensification in Uganda’s farmed landscapes and reported a decrease in bird abundance and species richness with land use intensification. Neuschulz et al. [81] showed that patches of natural forest in South Africa were essential for the conservation of local endemic bird species while generalist species were able to expand into and tolerate modified and degraded forest habitat.
Agricultural activities outside protected areas have resulted in changes in vegetation from diverse natural plant assemblages to cultivated land with large areas of single domestic crops and low diversity of indigenous plant species; crops also result in an increase in exotic plant species that are not suitable for the survival of local endemic bird species [82, 83]. Many studies of birds that compared protected areas with agricultural land have shown a dramatic decrease in bird species in the agricultural area [1, 20, 84, 85, 86, 87]. Similarly, Moreau [88] reported that intense habitat alteration by human activities has contributed to the disappearance of many species of birds. Land-use changes have also led to land degradation, which in turn negatively affects the species richness of birds [89, 90, 91, 92, 93, 94, 95].
5.2 Agricultural effects on the trophic cascade and food web stability
It was predicted that disturbance was intensified up the food chain so that more species would be lost at higher trophic levels due to agriculture. There was a drop in vertebrate feeders when compared to the lower trophic levels. The sharp decrease in the number of vertebrate feeders in agricultural land may have resulted from an increase in human activities through the removal of trees that birds use for nesting and perching. Another possibility is that the food items present in agricultural land may differ in type, abundance, and availability. Because disturbance becomes magnified up the trophic level, vertebrate feeder experiences greater effects than species at lower trophic levels and the results of this study have indicated this effect. This change in the trophic structure could result in more marked fluctuations in the biota if there is a top-down control in the food chain of the Serengeti ecosystem. Top-down control is defined as the limitation of population numbers at one level by species at higher trophic levels [96, 97]. There is considerable evidence that top-down control commonly occurs in all terrestrial ecosystems, as synthesized in Terborgh and Estes [98] and Estes et al. [99]. Top-down control in the Serengeti ecosystem is continually occurring. Nkwabi et al. [100] observed that insectivorous birds appear to limit insect numbers in grasslands and predicted that any removal of birds that feed on insects along agricultural land could result in an increased insect population which are pests for crops, and could become a problem for agricultural food production.
Another evidence reported by Byrom et al. [101] who studied the importance of black-winged kites formerly was black-shouldered kites (
These results suggested that any decrease in insectivore and herbivore bird species diversity in Serengeti National Park will result not from a decline in the number of species but from the very large numbers of some individual populations, particularly those of granivorous such as queleas, bishops, widowbirds, and whydahs. These numbers have reported to be resulted in lower diversity estimates, although the overall richness was higher in Savanna.
The lower species diversity and richness of vertebrate feeders in agricultural land were probably due to increased human activities including deforestation, overgrazing, and increase in pesticide use, poisoning of predators, overhunting, and direct harassment. This finding agrees with the hypothesis that top-feeding guild bird species are less able to tolerate change in habitat compared to species at lower feeding guilds. Generally, disturbance becomes magnified up the feeding guild so that birds that feed on vertebrates experience disturbance more than do herbivores [102, 103]. It has been shown that agricultural activities are among the factors that influence raptor species richness and abundance, raptor numbers within 30 km of protected area boundaries were 40% lower than those in the interior protected area [104]. Agriculture also has an effect on populations inside protected areas close to boundaries [103, 105, 106].
The decrease in species richness of herbivorous and insectivorous birds in agricultural land was probably due to a reduced number and diversity of native tree, shrub and herb species, and the diversity and abundance of invertebrates which are a source of food for most bird species, as has been shown around Serengeti National Park [1]. In addition, a few granivorous such as queleas and widowbirds have increased in abundance by taking advantage of the grain crops. Hulme and Cresswell [107] observed that agricultural habitat with monocultures of cereal crops like maize, sorghum, and millet was more attractive to some bird species than the woodland and shrubs of native Savanna around Jos in Nigeria. In contrast, Dhindsa and Saini [108] reported an increase in the richness of some granivorous bird species in the agricultural habitats in India.
Finally, the foraging efficiency of insectivorous and herbivorous birds can be influenced by predation risk due to changes in vegetation structure and abundance. It has been suggested that increased predation rates may result in declines of some bird species in agricultural land [109, 110].
6. Conclusion and recommendation
The main objective of the study was to examine the effects of agriculture on two aspects of the Savanna bird community. First, how did agriculture affect species that differed in range within Africa? The study aimed at finding out whether birds that are endemic, regional, and Africa-wide ranges differed in their sensitivity to habitat change due to agriculture. Second, how did agriculture affect species at different trophic levels? The study tested the prediction that birds at top feeding levels (vertebrate feeders) were less able to tolerate change in habitat compared to those species at lower feeding guilds (insectivores and herbivores). The results showed a greater proportional loss of endemic species compared to wider ranging species when the habitat was changed to agriculture. In general, local endemic bird species in the Serengeti ecosystem showed less tolerance to habitat change in agricultural land.
Secondly, top feeding guilds lost proportionately more species than lower feeding guilds when habitat changed to agriculture. Thus, vertebrate feeders were less able to tolerate changes in habitat structure in agricultural land compared to the lower trophic levels of insectivores and herbivores. This result is important because there is a known top-down control by predators on lower feeding guilds in the food chain. Vertebrate feeders appear to limit rodent numbers [101] and insectivorous birds control insects [111] in Savanna compared to agriculture. Therefore, the disappearance of vertebrate feeders and insectivores could result in higher levels of rodents and some insect pests in agricultural land. The effect of food chain collapse becomes a pest control problem facing farmers.
Findings from this study imply that if climate change causes the system to move up the rainfall gradient, birds will possibly also move into agricultural areas and so lose the fragile species. These movements of birds in East Africa toward wetter areas (because of environmental change) have now been documented by Beale et al. [112]; thus, these losses of birds have taken place since then. If such losses of birds are typical of agricultural landscapes in East Africa, then many of the African savannah birds will become confined to protected areas in the next few decades [112] and some of these species could be threatened with extinction because they are only found in these protected areas.
Acknowledgments
This work was funded by a Canadian Natural Sciences and Engineering Research Council grant to Prof. A.R.E. Sinclair and Dr. Simon A.R. Mduma. We thank the Serengeti National Park, Department of Zoology and Wildlife Conservation of University of Dar es Salaam, Serengeti Wildlife Research Centre, Tanzania Wildlife Research Institute, Tanzania National Parks and Tanzania Commission for Science and Technology for their help and permission to conduct this research. We are grateful to Stephen Makacha, John Mchetto, Joseph Masoy, and Dr Bukombe John for their assistance with fieldwork.
References
- 1.
Sinclair ARE, Mduma SAR, Arcese P. Protected areas as biodiversity benchmarks for human impact: agriculture and the Serengeti avifauna. Proceedings of the Royal Society of London. Series B: Biological Sciences. 2002; 269 (1508):2401-24 - 2.
Wunderlich SM, Martinez NM. Conserving natural resources through food loss reduction: production and consumption stages of the food supply chain. International Soil and Water Conservation Research. 2018; 6 (4):331-339 - 3.
Singh RL, Singh PK. Global environmental problems. Principles and Applications of Environmental Biotechnology for a Sustainable Future. 2017:13-41 - 4.
Wang L, Vo XV, Shahbaz M, Ak A. Globalization and carbon emissions: is there any role of agriculture value-added, financial development, and natural resource rent in the aftermath of COP21? Journal of Environmental Management. 2020; 268 :110712 - 5.
Moorcroft PR, Levin SA. Biodiversity patterns in managed and natural landscapes. In: Levin SA, Carpenter SR, Godfray HCJ, Kinzig AP, Loreau M, Losos JB, Walker B, Wilcove DS, editors. The Princeton Guide to Ecology. Princeton, New Jersey.: Princeton University Press; 2009. pp. 445-457 - 6.
Reside AE, Beher J, Cosgrove AJ, Evans MC, Seabrook L, Silcock JL, et al. Ecological consequences of land clearing and policy reform in Queensland. Pacific Conservation Biology. 2017; 23 (3):219-230 - 7.
Saridnirun G, Sukumal N, Grainger MJ, Savini T. Living with human encroachment: status and distribution of Green Peafowl in northern stronghold of Thailand. Global Ecology and Conservation. 2021; 28 :e01674 - 8.
Bukombe JK, Nkwabi AK, Mangewa LJ, Sweke EA, Kavana PY, Liseki SD, et al. Alien invasive species in Tanzania. In: Pullaiah T, Ielmini M, editors. Invasive Alien Species: Observations and Issues from Around the World. Wiley-Blackwell: USA; 2021. pp. 263-290 - 9.
Estes AB, Kuemmerle T, Kushnir H, Radeloff VC, Shugart HH. Land-cover change and human population trends in the greater Serengeti ecosystem from 1984-2003. Biological Conservation. 2012; 147 (1):255-263 - 10.
Estes AB, Kuemmerle T, Kushnir H, Radeloff VC, Shugart HH. Agricultural expansion and human population trends in the greater Serengeti ecosystem from 1984-2003. In: Sinclair ARE, Metzger KL, Fryxell JM, Mduma SAR, editors. Serengeti IV: Sustaining Biodiversity in a Coupled Human-Natural System. Chicago: University of Chicago Press; 2015. pp. 513-531 - 11.
Kideghesho JR, Nyahongo JW, Hassan SN, Tarimo TC, Mbije NE. Factors and ecological impacts of wildlife habitat destruction in the Serengeti ecosystem in northern Tanzania. African Journal of Environmental Assessment and Management. 2006; 11 :17-32 - 12.
Sinclair A, Hopcraft JGC, Olff H, Mduma SA, Galvin KA, Sharam GJ. Historical and future changes to the Serengeti ecosystem. In: Sinclair ARE, Packer C, Mduma SA, Fryxell JM, editors. Serengeti III: Human Impacts on Ecosystem Dynamics. Chicago: Chicago Press; 2008. pp. 7-46 - 13.
Walther BA, Van Niekerk A, Rahbek C. Long-term population declines of Palearctic passerine migrant birds: a signal from the Sahel. In: British Ornithologists’ Union’s (BOU) Annual Conference. 2011. http://www.bou.org.uk/bouproc-net/migratory-birds/walther-etal.pdf. [Accessed: 12 December 2013] - 14.
Sinclair AR, Nkwabi A, Mduma SA, Magige F. Responses of the Serengeti avifauna to long-term change in the environment. Ostrich. 2014; 85 (1):1-11 - 15.
Mkumbukwa AR. The evolution of wildlife conservation policies in Tanzania during the colonial and post-independence periods. Development Southern Africa. 2008; 25 (5):589-600 - 16.
Ogutu JO, Owen-Smith N, Piepho HP, Said MY. Continuing wildlife population declines and range contraction in the Mara region of Kenya during 1977-2009. Journal of Zoology. 2011; 285 (2):99-109 - 17.
Nkwabi AK, Bukombe J, Maliti H, Liseki S, Lesio N, Kija H. An overview of biodiversity of tanzania and conservation efforts. In: Pullaiah T, editor. Global Biodiversity Volume 3, Selected Countries in Africa. Waretown, USA.: Apple Academic Press; 2018. pp. 297-342 - 18.
Kauzeni AS, Kiwasila HL. Serengeti Regional Conservation Strategy: A Socio-Economic Study. Tanzania: Institute of Resource Assessment, University of Dar es Salaam; 1994 - 19.
Norton-Griffiths M, Herlocker D, Pennycuick L. The patterns of rainfall in the Serengeti ecosystem, Tanzania. East African Wildlife Journal. 1975; 13 (3-4):347-374 - 20.
Sinclair ARE, Mduma SAR, Hopcraft JGC, Fryxell JM, Hilborn R, Thirgood S. Long-term ecosystem dynamics in the Serengeti: lessons for conservation. Conservation Biology. 2007; 21 (3):580-590 - 21.
Sinclair ARE, Mduma SAR, Hopcraft JGC, Galvin K, Sharam GJ. Historical and future changes to the Serengeti ecosystem. In: Sinclair ARE, Packer C, Mduma SAR, Fryxell JM, editors. Serengeti III: Human Impacts on Ecosystem Dynamics. Chicago: University of Chicago Press; 2008. pp. 7-46 - 22.
Morell V. Counting creatures of the Serengeti, great and small. Science. 1997; 278 (5346):2058-2060 - 23.
Estes A. Avian Diversity in Serengeti National Park: How do Community Differ Between Interior Savanna and Grassland and Exterior Agricultural Landscape [MSc Thesis]. Madison: University of Wisconsin. 2000 - 24.
Sekercioglu CH. Increasing awareness of avian ecological function. Trends in Ecology and Evolution. 2006; 21 (8):464-471 - 25.
Sinclair ARE, Metzger KL, Fryxell JM, Packer C, Byrom A, Craft ME, et al. Asynchronous food-web pathways could buffer the response of Serengeti predators to El Niño Southern Oscillation. Ecology. 2013; 94 (5):1123-1130 - 26.
Metzger KL, Sinclair ARE, Macfarlane A, Coughenour MB, Ding J. Scales of Change in the Greater Serengeti Ecosystem. In: Sinclair ARE, Metzger KL, Mduma SAR, editors. Serengeti IV: Sustaining Biodiversity in a Coupled Human-Natural System. Chicago: University of Chicago Press; 2015. pp. 33-71 - 27.
Ikin K, Beaty RM, Lindenmayer DB, Knight E, Fischer J, Manning AD. Pocket parks in a compact city: how do birds respond to increasing residential density? Landscape Ecology. 2013; 28 :45-56 - 28.
Newbold T, Scharlemann JP, Butchart SH, Şekercioğlu ÇH, Alkemade R, Booth H, et al. Ecological traits affect the response of tropical forest bird species to land-use intensity. Proceedings of the Royal Society B: Biological Sciences. 2013; 280 (1750):2012-2131 - 29.
Sykes MT, Prentice IC, Cramer W. A bioclimatic model for the potential distributions of north European tree species under present and future climates. Journal of Biogeography. 1996; 23 (2):203-233 - 30.
Pearson RG, Dawson TP. Predicting the impacts of climate change on the distribution of species: are bioclimate envelope models useful? Global Ecology and Biogeography. 2003; 12 (5):361-371 - 31.
Both C, Visser ME. The effect of climate change on the correlation between avian life-history traits. Global Change Biology. 2005; 11 (10):1606-1613 - 32.
Williams P, Hannah L, Andelman S, Midgley G, AraúJo M, Hughes G, et al. Planning for climate change: identifying minimum-dispersal corridors for the Cape Proteaceae. Conservation Biology. 2005; 19 (4):1063-1074 - 33.
Burger C, Belskii E, Eeva T, Laaksonen T, Mägi M, Mänd R, et al. Climate change, breeding date and nestling diet: how temperature differentially affects seasonal changes in pied flycatcher diet depending on habitat variation. Journal of Animal Ecology. 2012; 81 (4):926-936 - 34.
Grainger A. Constraints on modelling the deforestation and degradation of tropical open woodlands. Global Ecology and Biogeography. 1999; 8 (3-4):179-190 - 35.
Homewood K, Brockington D. Biodiversity, conservation and development in Mkomazi Game Reserve, Tanzania. Global Ecology and Biogeography. 1999; 8 (3-4):301-313 - 36.
Thomas CD, Gillingham PK. The performance of protected areas for biodiversity under climate change. Biological Journal of the Linnean Society. 2015; 115 (3):718-730 - 37.
Thomas CD, Cameron A, Green RE, Bakkenes M, Beaumont LJ, Collingham YC, et al. Extinction risk from climate change. Nature. 2004; 427 (6970):145-148 - 38.
Sekercioglu CH, Primack RB, Wormworth J. The effects of climate change on tropical birds. Biological Conservation. 2012; 148 (1):1-18 - 39.
Cowling R, Pressey R, Lombard A, Desmet P, Ellis A. From representation to persistence: requirements for a sustainable system of conservation areas in the species-rich mediterranean-climate desert of southern Africa. Diversity and Distributions. 1999; 5 (1-2):51-71 - 40.
Rutherford M, Powrie L, Schulze R. Climate change in conservation areas of South Africa and its potential impact on floristic composition: a first assessment. Diversity and Distributions. 1999; 5 (6):253-262 - 41.
Araújo MB, Cabeza M, Thuiller W, Hannah L, Williams PH. Would climate change drive species out of reserves? An assessment of existing reserve-selection methods. Global Change Biology. 2004; 10 (9):1618-1626 - 42.
Gill JA. Approaches to measuring the effects of human disturbance on birds. Ibis. 2007; 149 (Suppl. 1):9-14 - 43.
Hole DG, Willis SG, Pain DJ, Fishpool LD, Butchart SH, Collingham YC, et al. Projected impacts of climate change on a continent-wide protected area network. Ecology Letters. 2009; 12 (5):420-431 - 44.
Carey C. The impacts of climate change on the annual cycles of birds. Philosophical Transactions of the Royal Society B: Biological Sciences. 2009; 364 (1534):3321-3330 - 45.
Sillett TS, Holmes RT, Sherry TW. Impacts of a global climate cycle on population dynamics of a migratory songbird. Science. 2000; 288 (5473):2040-2042 - 46.
Myers N, Knoll AH. The biotic crisis and the future of evolution. Proceedings of the National Academy of Sciences. 2001; 98 (10):5389-5392 - 47.
Walker BH. Biodiversity and ecological redundancy. Conservation Biology. 1992; 6 (1):18-23 - 48.
Walker BH. Conserving biological diversity through ecosystem resilience. Conservation Biology. 1995; 9 (4):747-752 - 49.
Rapport D, Costanza R, McMichael A. Assessing ecosystem health. Trends in Ecology & Evolution. 1998; 13 (10):397-402 - 50.
Chapin FS, Zavaleta ES, Eviner VT, Naylor RL, Vitousek PM, Reynolds HL, et al. Consequences of changing biodiversity. Nature. 2000; 405 (6783):234-242 - 51.
May RM. Thresholds and breakpoints in ecosystems with a multiplicity of stable states. Nature. 1977; 269 (5628):471-477 - 52.
Holling CS. Resilience and stability of ecological systems. Annual Review of Ecology and Systematics. 1973; 4 :1-23 - 53.
Cannell M, Thornley J, Mobbs D, Friend A. UK conifer forests may be growing faster in response to increased N deposition, atmospheric CO2 and temperature. Forestry. 1998; 71 (4):277-296 - 54.
Pavlacky DC, Possingham HP, Lowe AJ, Prentis PJ, Green DJ, Goldizen AW. Anthropogenic landscape change promotes asymmetric dispersal and limits regional patch occupancy in a spatially structured bird population. Journal of Animal Ecology. 2012; 81 (5):940-952 - 55.
Parmesan C. Climate and species' range. Nature. 1996; 382 (6594):765-766 - 56.
Jezkova T. The Ecological Niche and Ecological Niche Modeling. Las Vegas: School of Life Sciences, University of Nevada; 2012 - 57.
Mathews S, Bonser SP. Life histories, ecological tolerance limits, and the evolution of geographic range size in Eucalyptus (Myrtaceae). Australian Journal of Botany. 2005; 53 (6):501-508 - 58.
Gilchrist GW. Specialists and generalists in changing environments. I. Fitness landscapes of thermal sensitivity. American Naturalist. 1995; 146 (2):252-270 - 59.
Gaston KJ. Geographic range limits of species. Proceedings of the Royal Society B: Biological Sciences. 2009; 276 (1661):1391-1393 - 60.
Nkwabi AK, Bukombe JK, Kija HK, Liseki SD, Ndimuligo SA, Kavana PY. Avifauna in relation to habitat disturbance in Wildlife Management Areas of the Ruvuma miombo ecosystem, Southern Tanzania. In: Mikkola H, editor. Birds - Challenges and Opportunities for Business, Conservation and Research. London, UK, London: IntechOpen; 2021. pp. 1-20 - 61.
Nkwabi AK, John B, Kija H, Otsyina RM, Monjare JF, Kajuni AR. Abundance and distribution of small mammals relative to human activities in the wildlife management areas of Ruvuma landscape, southern Tanzania. International Journal of Fauna and Biological Studies. 2018; 5 (2):156-162 - 62.
Thonfeld F, Steinbach S, Muro J, Hentze K, Games I, Naeschen K, et al. The impact of anthropogenic land use change on the protected areas of the Kilombero catchment, Tanzania. ISPRS Journal of Photogrammetry and Remote Sensing. 2020; 168 :41-55 - 63.
Kideghesho JR, Rija A, Mwamende K, Selemani I. Emerging issues and challenges in conservation of biodiversity in the rangelands of Tanzania. Nature Conservation. 2013; 6 :1-29. DOI: 10.3897/natureconservation.6.5407 - 64.
Mwalyosi RB. Land-use Changes and Resource Degradation in South-West Masailand, Tanzania. Environmental Conservation. 1992; 19 (02):145-152 - 65.
Rybicki J, Hanski I. Species–area relationships and extinctions caused by habitat loss and fragmentation. Ecology Letters. 2013; 16 (Suppl.1):27-38 - 66.
Kideghesho JR, Kaltenborn BP, Røskaft E, Tarimo TM. Serengeti shall not die: can the ambition be sustained? The International Journal of Biodiversity Science and Management. 2005; 1 (3):150-166 - 67.
Sinclair ARE, Norton-Griffiths M, editors. Serengeti: Dynamics of an Ecosystem. Chicago: University of Chicago Press; 1979 - 68.
Sinclair I, Ryan P. Birds of Africa: South of the Sahara. Cape Town: Random House, Struik; 2010 - 69.
Fry CH, Keith S, Urban EK, editors. Birds of Africa Volume VI. Picathartes to Oxpeckers. San Diego, California: Academic Press; 2000 - 70.
Fry CH, Keith S. The birds of Africa Volume VII. Sparrows to buntings. London: Christopher Helm; 2004 - 71.
Simberloff D, Dayan T. The guild concept and the structure of ecological communities. Annual Review of Ecology and Systematics. 1991; 22 :115-143 - 72.
Hammer Ø, Harper DAT, Ryan PD. PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica. 2001; 4 (1):1-9 - 73.
Krebs CJ, Kenney AJ. Programs for Ecological Methodology. Menlo Park: Bennjamin/Cummings; 2009 - 74.
Krebs CJ. Ecological Methodology. New York, NY: Harper and Row; 2001 - 75.
Seaby RMH, Henderson PA. Species Diversity and Richness Version 4.1.2. 2007, Pisces Conservation Ltd: Lymington - 76.
Solow AR. A simple test for change in community structure. Journal of Animal Ecology. 1993; 62 (1):191-193 - 77.
Terborgh J, Lopez L, Nunez P, Rao M, Shahabuddin G, Orihuela G, et al. Ecological meltdown in predator free forest fragments. Science. 2001; 294 (5548):1923-1926 - 78.
Bonier F, Martin PR, Wingfield JC. Urban birds have broader environmental tolerance. Biology Letters. 2007; 3 (6):670-673 - 79.
Waltert M, Mardiastuti A, Mühlenberg M. Effects of land use on bird species richness in Sulawesi, Indonesia. Conservation Biology. 2004; 18 (5):1339-1346 - 80.
Bolwig S, Pomeroy D, Tushabe H, Mushabe D. Crops, trees, and birds: biodiversity change under agricultural intensification in Uganda's farmed landscapes. Geografisk Tidsskrift, Danish Journal of Geography. 2006; 106 (2):115-130 - 81.
Neuschulz EL, Botzat A, Farwig N. Effects of forest modification on bird community composition and seed removal in a heterogeneous landscape in South Africa. Oikos. 2011; 120 (9):1371-1379 - 82.
Van Dorp D, Opdam P. Effects of patch size, isolation and regional abundance on forest bird communities. Landscape Ecology. 1987; 1 (1):59-73 - 83.
Lepczyk CA, Flather CH, Radeloff VC, Pidgeon AM, Hammer RB, Liu J. Human impacts on regional avian diversity and abundance. Conservation Biology. 2008; 22 (2):405-416 - 84.
Soini E. Bird diversity and land use on the slopes of Mt Kilimanjaro and the adjacent plains, Tanzania. African Zoology. 2006; 41 (2):193-209 - 85.
Newmark WD. Isolation of African protected areas. Frontiers in Ecology and the Environment. 2008; 6 (6):321-328 - 86.
Devictor V, Godet L, Julliard R, Couvet D, Jiguet F. Can common species benefit from protected areas? Biological Conservation. 2007; 139 (1):29-36 - 87.
Sheehan DK, Sanderson FJ. Seeing the bigger picture: how anthropogenic landscape modification in Africa affects declining migratory birds and the need for trans-continental research and conservation. Ibis. 2012; 154 (4):659-662 - 88.
Moreau RE. Mt Kilimanjaro and Mt Kenya: some comparisons with special reference to mammals and birds with a note on Mt Meru (with bibliography of Kilimanjaro by C. Gillman). Tanganyika Notes and Records. 1944; 14 :28-59 - 89.
Misana SB, Majule AE, Lyaruu HV, Change LU. Linkages between changes in land use, biodiversity and land degradation on the slopes of Mount Kilimanjaro, Tanzania. LUCID Project, International Livestock Research Institute. 2003 - 90.
Atkinson PW, Buckingham D, Morris AJ. What factors determine where invertebrate-feeding birds forage in dry agricultural grasslands? Ibis. 2004; 146 (Suppl. 2):99-107 - 91.
Long P, Székely T, Kershaw M, O'Connell M. Ecological factors and human threats both drive wildfowl population declines. Animal Conservation. 2007; 10 (2):183-191 - 92.
Woodroffe R, Ginsberg JR. Edge effects and the extinction of populations inside protected areas. Science. 1998; 280 :2126-2128 - 93.
Barnes AD, Allen K, Kreft H, Corre MD, Jochum M, Veldkamp E, et al. Direct and cascading impacts of tropical land-use change on multi-trophic biodiversity. Nature Ecology & Evolution. 2017; 1 (10):1511-1519 - 94.
Rurangwa ML, Aguirre-Gutiérrez J, Matthews TJ, Niyigaba P, Wayman JP, Tobias JA, et al. Effects of land-use change on avian taxonomic, functional and phylogenetic diversity in a tropical montane rainforest. Diversity and Distributions. 2021; 27 (9):1732-1746 - 95.
Müller S, Gossner MM, Penone C, Jung K, Renner SC, Farina A, et al. Land-use intensity and landscape structure drive the acoustic composition of grasslands. Agriculture, Ecosystems & Environment. 2022; 328 :107845 - 96.
Sinclair ARE. Population regulation in animals, ecological concepts. Symposium of the British Ecological Society. 1989; 29 :197-241 - 97.
Sinclair ARE, Metzger K, Brashares JS, Nkwabi A, Sharam G, Fryxell JM, et al. Trophic cascades in African savanna: Serengeti as a case study. In: Terborgh J, Estes J, editors. Trophic Cascades: Predators, Prey, and the Changing Dynamics of Nature. Washington, DC.: Island Press; 2010. pp. 255-274 - 98.
Terborgh J, Estes JA, editors. Trophic Cascades: Predators, Prey, and the Changing Dynamics of Nature. 2010 - 99.
Estes JA, Terborgh J, Brashares JS, Power ME, Berger J, Bond WJ, et al. Trophic downgrading of planet earth. Science. 2011; 333 (6040):301-306 - 100.
Nkwabi AK, Sinclair ARE, Metzger KL, Mduma SAR. The effect of natural disturbances on the avian community of the Serengeti woodlands. In: Sinclair ARE, Metzger K, Mduma SAR, Fryxell J, editors. Serengeti IV: Sustaining Biodiversity in a Coupled Human-Natural System. Chicago: University of Chicago; 2015. pp. 395-418 - 101.
Byrom AE, Ruscoe W, Nkwabi AJK, Metzger K, Forrester GJ, Durant S, et al. Small mammal diversity and population dynamics in the greater Serengeti ecosystem. In: Sinclair ARE, Metzger K, Mduma SAR, Fryxell J, editors. Serengeti IV: Sustaining Biodiversity in a Coupled Human-Natural System. Chicago: University of Chicago; 2015. pp. 323-357 - 102.
Tuule E, Tuule A, Lõhmus A. Fifty-year dynamics in a temperate raptor assemblage. Estonian Journal of Ecology. 2011; 60 :132-142 - 103.
Anadón JD, Sánchez-Zapata JA, Carrete M, Donázar JA, Hiraldo F. Large-scale human effects on an arid African raptor community. Animal Conservation. 2010; 13 (5):495-504 - 104.
Herremans M, Herremans-Tonnoeyr D. Land use and the conservation status of raptors in Botswana. Biological Conservation. 2000; 94 (1):31-41 - 105.
Thiollay JM. The decline of raptors in West Africa: long-term assessment and the role of protected areas. Ibis. 2006; 148 (2):240-254 - 106.
Ogada DL, Keesing F. Decline of raptors over a three-year period in Laikipia, central Kenya. Journal of Raptor Research. 2010; 44 (2):129-135 - 107.
Hulme MF, Cresswell W. Density and behaviour of Whinchats Saxicola rubetra on African farmland suggest that winter habitat conditions do not limit European breeding populations. Ibis. 2012; 154 (4):680-692 - 108.
Dhindsa MS, Saini HK. Agricultural ornithology: an Indian perspective. Journal of Biosciences. 1994; 19 (4):391-402 - 109.
Whittingham MJ, Evans KL. The effects of habitat structure on predation risk of birds in agricultural landscapes. Ibis. 2004; 146 (Suppl. 2):210-220 - 110.
Marsden SJ, Symes CT, Mack AL. The response of a New Guinean avifauna to conversion of forest to small-scale agriculture. Ibis. 2006; 148 (4):629-640 - 111.
Nkwabi A, Sinclair AE, Metzger K, Mduma S. Disturbance, species loss and compensation: wildfire and grazing effects on the avian community and its food supply in the Serengeti Ecosystem, Tanzania. Austral Ecology. 2011; 36 (4):403-412 - 112.
Beale CM, Baker NE, Brewer MJ, Lennon JJ. Protected area networks and savannah bird biodiversity in the face of climate change and land degradation. Ecology Letters. 2013; 16 (8):1061-1068