Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
Translational control of viral gene expression is a fundamental process essential for the vitality of all viruses. In special cases, signals encoded in the mRNA reprogram the ribosome to read the message in a different way, a process termed “translational recoding”. The 2A region of the foot-and-mouth disease virus (FMDV) encodes a short sequence, only 18 amino acids, that mediates self-processing by a novel translational effect “ribosome skipping” rather than proteolysis. Briefly, 2A interacts with the ribosome exit tunnel to inhibit peptide bond formation at the C terminus of the 2A sequence. Translation terminates at this point, but then resumes elongation, creating a second independent protein product. Thus, discrete proteins can be produced from a single transcript. The 2A sequence is particularly useful in vector strategies (AAV and retroviral vectors) where the capacity to incorporate foreign DNA is limited. Use of 2A and “2A-like” peptides to link the sequences encoding several proteins in the same open reading frame has led to their increasing use as important tools in biotechnology and biomedicine. This technology has been crucial for the visual tracking of expressed proteins, human gene therapies targeting cancer, production of induced human pluripotent stem cells for regenerative medicine, creation of transgenic animals and plants and the improvement of CRISPR-Cas9 and TALEN genome editing methods.
Centre for Biomolecular Sciences, School of Biology, University of St Andrews, St Andrews, UK
Martin D. Ryan
Centre for Biomolecular Sciences, School of Biology, University of St Andrews, St Andrews, UK
*Address all correspondence to: gal@st-andrews.ac.uk
1. Introduction
As obligate intracellular parasites with relatively limited coding capacity, viral strategies to sequester the host translation machinery target the initiation, elongation, and termination steps of protein synthesis. Although most translational control strategies operate at the rate-limiting initiation step, different regulatory mechanisms target elongation and termination. Commonly these involve translational control or “translational recoding” defined as instances in which “the rules for decoding are temporarily altered through the action of specific signals built into the mRNA sequences” [1, 2]. The two most common types of recoding elements program ribosomes to either shift the reading frame of an mRNA, typically by one base in either the 5′(−1) or the 3′(+1) direction (programmed ribosomal frameshifting) or reassign the “sense” of stop codons (UAA, UAG or UGA) leading to stop codon “Read-Through”. Less common are recoding elements that cause ribosomes to bypass defined segments of mRNAs (ribosomal “hopping”) or those that skip the synthesis of a specific peptide bond (ribosomal “skipping”). Ribosomal skipping, mediated by a “self-cleaving” 2A oligopeptide that manipulates the ribosome to “skip” the synthesis of the glycyl-prolyl peptide bond at its own carboxyl terminus leading to release of the nascent protein and translation of the downstream sequence. In this chapter we outline the development of this efficient methodology for the co-expression of multiple proteins from a single gene using a small 2A peptide originally identified in viruses.
Among the five distinct types of picornavirus 2A proteins, a group of oligopeptide sequences collectively known as Aphthovirus-like 2A mediate a translational recoding event in cis known as “ribosome skipping”, “StopGo” or “Stop Carry-on” translation [3, 4, 5, 6]. Here a termination event occurs at a sense codon, followed by release of the nascent polypeptide and ribosomal translocation to the next in-frame codon. The function of the 2A sequence was first characterised from the positive-stranded RNA picornavirus Foot-and-Mouth Disease Virus (FMDV). The 2A oligopeptide is only 18 amino acids (aa) long (-LLNFDLLKLAGDVESNPG-), delineated by 3Cpro post-translational cleavage at its N-terminus and co-translational “cleavage” at its C-terminus [7, 8, 9]. The core sequence at the C-terminus of 2A is strongly conserved and contains the canonical motif D12(V/I)E(S/T)NPG2A↓P192B (where P19 refers to the completely conserved first residue of the downstream FMDV protein 2B). The less conserved part of the 2A sequence, located upstream of the motif, appears to be essential for 2A function [9, 10]. During this period the number of genome sequences was limited, but it was apparent that a similar motif was also present at the C-terminus of the longer cardiovirus 2A protein [11]. Analysis of recombinant FMDV polyproteins indicated FMDV 2A, together with the N-terminal proline of protein 2B, appeared sufficient for a co-translational “cleavage” between the upstream (capsid proteins) and downstream (RNA replication protein) domains of the polyprotein (-NPG↓P-) [7, 12]. Following cleavage, the 2A “tag” remains as a C-terminal extension of the upstream protein. The single proline at the N-terminus of the downstream protein confers a long half-life (>20 h) which should help increase protein stability [13]. Early studies showed the FMDV 2A region was not simply a substrate for a virus proteinase (Lpro, 3Cpro), nor a substrate for a host-cell proteinase—“self-cleavage” was a novel translational effect [3, 7, 12]. Due to its mode of action within the ribosome the acronym CHYSEL (cis-acting hydrolase element) was proposed as an alternative name for 2A and 2A-like sequences to avoid confusion with protease mediated cleavage [14].
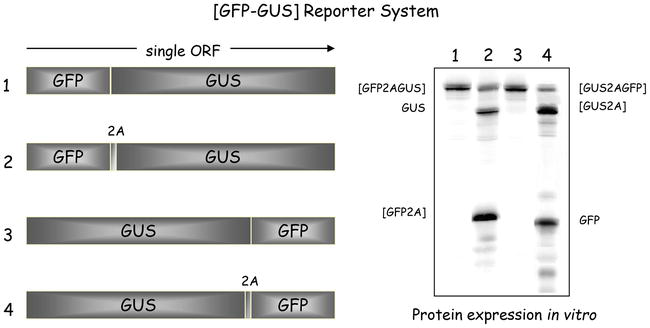
To monitor gene expression, artificial “self-processing” polyproteins were constructed with these 18 codons, plus the first aa (proline) of 2B (referred to as “2A”), separating green fluorescent protein (GFP—stop codon deleted) and β-glucuronidase (GUS) reporter-encoding sequences in a single open reading frame (ORF) encoding [GFP2AGUS] (Figure 1, pSTA1) [3, 15, 16]. In the absence of all other FMDV proteins, such artificial polyproteins comprised well-characterised, metabolically stable “reporter” proteins and used to program in vitro translation systems. These produced a simple translation profile that could be readily interpreted—and the accumulation of the processing products quantified. A control construct encoding [GFPGUS] produced only the fusion protein [GFPGUS], whereas the (GFP2AGUS) was shown to be highly active in mediating cleavage, producing the uncleaved polyprotein product (GFP2AGUS) (~5% of radiolabel incorporated) and the cleavage products (GFP2A) and [GUS], together accounting for ~95% incorporation of radiolabel. This approach gave the surprising result that the GFP reporter protein upstream of 2A accumulated to a higher level than that downstream of 2A (GUS) (Figure 1). Reversing the gene order (GUS2AGFP) produced the same type of imbalance. Having eliminated “trivial” causes for this imbalance (e.g. protein/RNA degradation, premature termination of transcription/translation) [3], these data were at variance with a proteolytic mechanism which predicts a unitary stoichiometry of the products. Inhibition of protein synthesis by the antibiotic puromycin produced a distinct product with a size corresponding to the upstream protein, indicating a pause on the mRNA at the site where the nascent chain is released [3]. The ribosome pause seen by ribosome toeprinting occurred at the end of the 2A coding sequence (-NPG↓P-), with glycine and proline in the P- and A- sites, respectively [17]. This imbalance was due to different levels of biosynthesis of each portion of the ORF and represents a novel type of recoding [5, 18].
Figure 1.
Analysis of 2A-mediated “cleavage”. Artificial reporter polyproteins (boxed areas) used to programme in vitro translation systems are shown together with translation profiles obtained from rabbit reticulocyte lysates (right). The control pGFPGUS and pGUSGFP constructs produce only a single translation product—the [GFPGUS] and [GUSGFP] fusion proteins respectively. The translation profile from the pGFP2AGUS construct shows three major products: uncleaved [GFP2AGUS] and the cleavage products [GFP2A] and [GUS]. The profile from pGUS2AGFP also shows three major products: uncleaved [GUS2AGFP] and the cleavage products [GUS2A] and [GFP].
Owing to the degeneracy of the genetic code, a protein sequence can be encoded by many different synonymous mRNA coding sequences in protein biogenesis. Furthermore, many organisms, including viruses, tend to have biases towards certain synonymous codons (the “codon bias”) in their genes. As described by Gao et al. [19] a marked codon bias within the FMDV genome is evident—the amino acids E, S, N, P, G, P tend to use GAG, TCC, AAC, CCT, GGG and CCC respectively. However, in the context of a synthetic reporter polyprotein, synonymous codon usage of this conserved motif does not affect apparent cleavage efficiencies whereas introducing non-synonymous changes impairs cleavage activity. Frameshifting the 2A oligopeptide with respect to the reporter proteins completely abolishes cleavage activity [3, 10, 15, 20]. Further, synonymous codon usage patterns for G2A at the 2A/2B cleavage site (GGG, GGC, GGA, GGT) have no effect on the cleavage efficiency [19]. This supports the view that it is the amino acid residue rather than the nucleotide sequence which is critical for the activity of 2A. In contrast to these studies, Kjær and Belsham found that in FMDV genomes codon bias operates in encoding the NPGP motif, raising the possibility that the RNA sequence could also contribute to the recoding event [21]. It’s noteworthy that conservative mutation of all potential nucleophilic amino acids within the highly conserved D12(V/I)E(S/T)NPG2A↓P192B motif are active—further proof that 2A-mediated cleavage is not a proteolytic mechanism [15]. The take home message from mutagenesis experiments is that 2A is largely intolerant to sequence alteration over most of its length—2A peptides are fine tuned to function as a whole [5, 9, 10, 15, 22]. Our own extensive mutagenesis indicates that several variations of the conserved motif still display some level of activity [10, 15]. However, certain 2A sequences have not evolved to produce a simple binary outcome. “Sub-optimal” 2A sequences usually result in either “standard” translation (generating a fusion protein), or, a molar excess of products upstream of 2A—due to dissociation of ribosomes after peptide release, rather than recommencing elongation. It is not difficult to imagine clear advantages associated with incomplete StopGo activity (e.g. dual protein targeting, described in Roulston et al. [23]), but also the possibility of “regulating” the ratio of up- to down-stream products (e.g. “sensor” of translational stress, described in [24]).
Various studies have demonstrated that synonymous codon usage bias plays an important role in the translation of certain mRNAs [25, 26]. While codon optimisation is routinely used to increase translational efficiency, with regard to 2A peptides, wild type sequences may be evolutionary optimised to enhance the StopGo mechanism. The length of the FMDV 2A used is also important for cleavage in vitro/in vivo—N-terminally truncated forms of 2A in our reporter system showed that the minimal length required for activity was 12aa (12 N-terminal and 1 C-terminal to the G↓P site) [9, 15]. In the case of shorter 2As, cleavage efficiency has been improved by insertion of various spacer sequences immediately upstream of the 2A sequence such as a glycine-serine linker (e.g. -GSG- or -SGSG-, [27, 28, 29]; a 3XFLAG epitope tag -DYKDHDG-DYKDHDI-DYKDDDDK-, [30]; or a V5 epitope tag -GKPUPNPLLGLDST- [31]. These “flexible” linkers create a space between the N-terminal protein and the 2A peptide, favouring a conformation which facilitates efficient cleavage [32]. By contrast, an optimised Kozak sequence from the silkworm (Bombyx mori) placed immediately downstream of the 2A peptide, improved the expression of the sequence downstream of 2A [33]. Longer versions with extra sequences (>5aa) derived from the FMDV capsid protein (“1D”) immediately upstream of 2A produce higher levels of cleavage [12, 15, 16]. After “tweaking” of the FMDV 2A sequence, 2A (+11aa 1D) proved to be the most favourable in terms of both length and cleavage efficiency [34, 35]. GSG linkers and longer versions of 2A sequences to improve “skipping” efficiencies are not fail-safe solutions and should be assessed empirically for every protein in context. Previously described position effects have also shown that expression of genes flanking 2A is highly dependent on their arrangement [33, 36, 37, 38]. For example, the yield of β-carotene by engineered yeasts could be altered by exchanging the order of codon-optimised carotenogenic genes GGPPS, CARB and CARRP [39].
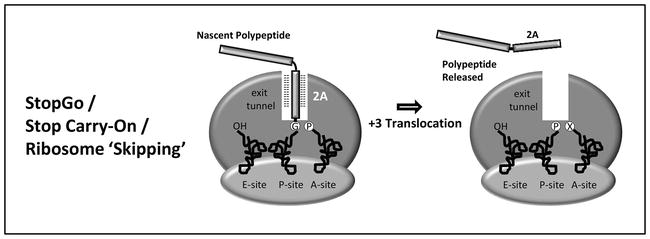
Based on dynamic molecular modelling and results from experiments with in vitro translation systems, the model of 2A-mediated translational recoding [3, 20, 40] proposes the nascent 2A peptide (~25aa) interacts with the exit tunnel of the ribosome such that the C-terminal portion is sterically constrained within a region of the peptidyl transferase centre (PTC) of the ribosome that cannot be accessed by prolyl-tRNAPro. The ribosomal PTC resides in the large ribosomal subunit and catalyses the two principle chemical reactions of protein synthesis: peptide bond formation and peptide release. It should be noted that the unique case of proline the nucleophile (nitrogen) is also sterically constrained since it is part of a ring structure. It has been shown that prolyl-tRNAPro is the poorest nucleophile among all aminoacyl tRNAs—polypeptide elongation is paused at this specific site. Our StopGo model predicts that (i) prolyl-tRNAPro dissociates from the A-site, (ii) the nascent polypeptide is released at this site by termination factors eRF1/3 and (iii) prolyl-tRNAPro (re)enters the ribosome a the A-site and must by translocated to the P-site to allow the next aminoacyl-tRNA to enter for resumption of translation and the synthesis of the downstream product (Figure 2) [17, 41]. Machida and colleagues showed however that processing of encephalomyocarditis virus proteins 2A and 2B in a reconstituted in vitro translation system requires neither eukaryotic initiation or termination (release) factors [42].
Figure 2.
Model of 2A translational “recoding” activity. In ribosome “skipping”, the 2A sequence is thought to interact with the ribosome exit such that the stereochemistry in the peptidyl transferase centre is altered: the peptide bond is not formed. The model proposes that prolyl-tRNA exits the A-site, eRF1/3 enters the A-site (at a sense codon) and terminates translation. After exit of eRF1, prolyl-tRNA then re-enters the A-site and is then (pseudo) translocated such that the A-site is now vacant and elongation can resume.
Our proposed model of this StopGo translational recoding event predicts that three alternative outcomes arise; having synthesised sequences upstream of 2A, ribosomes either (i) release the nascent peptide and then resume translation of the downstream sequences, (ii) release the nascent peptide and translation is terminated at that point, or, (iii) that no translational recoding occurs: the glycyl-prolyl peptide bond is formed and the protein is synthesised in the normal manner. The ratio of these translation products is dependent upon the specific 2A-like sequence in question—a view supported by the observations from many of the laboratories that have used StopGo protein co-expression applications. Initially studied in mammalian cell cultures (HeLa, [14]; HEK293, [43]; CHO, [44]) 2A-mediated ribosome skipping functions in all eukaryotic expression systems tested to date: amoeba [45], fungi [38, 46, 47, 48, 49], algae [50, 51], plants [52, 53] and animals [33, 54, 55, 56] but not prokaryotic systems: the structure of the ribosome and the exit tunnel is highly conserved among eukaryotes but differs between eu-and prokaryotes.
4.2 Targeted expression of recombinant proteins
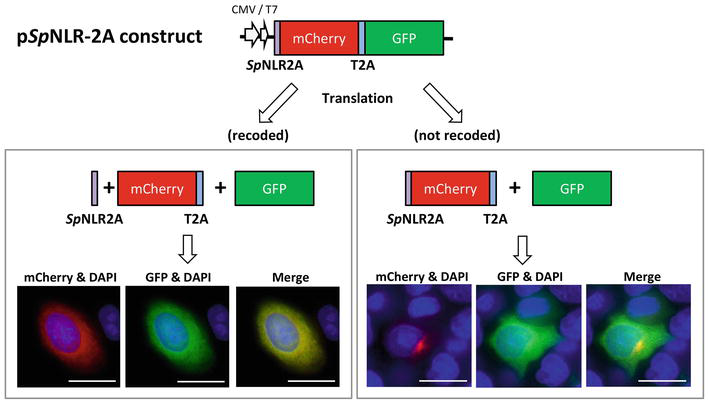
Our model of the 2A mechanism predicted this “cleavage” would occur within the ribosome: that any protein downstream of 2A would emerge from the ribosome with a nascent N-terminus. Therefore, if one included a co-translational signal sequence immediately downstream of 2A, then this would be recognised by the signal recognition particle (SRP) as if it were generated during “normal” translation. In a proof-of-concept study this was the case: polyproteins were constructed comprising internal signal sequences and we were able to show that such proteins could be both co-expressed and independently targeted to sub-cellular sites (or secreted), greatly increasing the utility of the 2A co-expression system [57, 58]. To verify the applicability of this method to developmental studies, Trichas et al. designed a 2A bicistronic reporter that correctly processed a membrane-localised TdTomato gene (Myr-TdTomato) and nuclear-localised enhanced green fluorescent protein (EGFP) gene (H2B-GFP) in cell culture and transgenic mice [59]. Additionally, this approach produced functional expression of Myr-mCherry and H2B-GFP in the branchipod crustacean Daphnia magna [60]. Meanwhile, we demonstrated the (partially) active 2A-like sequences present at the N-terminus of a number of NOD-like receptor proteins (NLRs) within the purple sea urchin (Strongylocentrotus purpuratus) also play a dual role as exocytic pathway signal peptides. If 2A mediates translational recoding, the 2A signal sequence is synthesised as a discrete translation product separate from the downstream product which is localised to the cytoplasm. If 2A does not mediate translational recoding, the 2A signal sequence is fused to the downstream translation product, functions as a signal sequence and targets the fusion protein to the exocytic pathway—a novel form of dual protein targeting (Figure 3) [23].
Figure 3.
Cleavage and signal sequence functions of 2A. The wild-type and point mutant version was cloned upstream of sequences encoding cherry FP and GFP linked via Thosea asigna virus (TaV) 2A into a single ORF. Image analysis shows the wild-type 2A produces a high proportion of cherry localised in the cytoplasm (and diffuses into the nucleus). The mutant inactive 2A acts as a signal targeting cherry to the exocytic pathway (bars ~5μm. Nuclei stained with DAPI are shown in blue).
4.3 The unwanted “tags”
The addition of partial 2A peptide sequences to the C-terminus of proteins may interfere with enzymatic activity [61]. For secreted proteins, inclusion of a furin recognition sequence (e.g. ↓-RRR-, ↓-RKRR-, ↓-RRKR-) between the first gene and 2A results in efficient removal of 2A residues—furin is a ubiquitous serine protease localised on the trans-Golgi networks of virtually all cell types [62]. Proteins expressed in plants/fungi could have this tag removed by endogenous proteinases acting on similar hybrid linker peptides, LP4-2A in plants (SN↓AADEVAT) [63], TEV-2A in fungi (ENLYFQ↓S) [49]. Although not a full-proof solution, a GSG linker used in conjunction with a furin recognition site (e.g -furin-GSG-2A-, -GSG-furin-GSG-2A-) reportedly enhanced cleavage efficiency via increased exposure of the -RRKR- site [32, 44, 64]. The “unwanted” tag may remain—recognised by monoclonal antibodies, peptide epitopes can characterise, purify, and localise proteins of interest in vitro and in vivo [65, 66]. Commercial antibodies have been raised against the consensus 2A sequence, which thus serves as a useful target for identifying 2A-tagged proteins in biochemical assays—validated research applications include ICC, immunofluorescence, IP, and Western Blot [67, 68]. The presence of a proline amino acid residue (or often several additional amino acids, derived from restriction site sequences introduced during cloning) at the N-terminus of co-expressed proteins may affect protein stability within polycistronic constructs [69, 70, 71]. In this case, proteins that require authentic termini can be introduced at the first gene position.
The presence of 2A-like sequences has been reported in a number of viral genomes within different genera of the Picornaviridae, other positive strand viruses such as the Dicistroviridae and Iflaviridae [16, 72], double-strand RNA viruses belonging to the Totiviridae/Reoviridae families [73], and surprisingly in a tentatively assigned negative-sense single-stranded RNA virus of the Bunyaviridae family [72]. The positive-strand RNA viruses typically possess one 2A/2A-like sequence, but some viruses have two, three or more motifs—the picornavirus duck egg-reducing syndrome virus (DERSV) was shown to have a total of seven 2A proteins, the first six separated by the “DxExNPGP” sequence motif (Table 1) [74]. Translation in vitro with our pSTA1 dual reporter system [GFP-2A-GUS] shows some of these multiple 2ANPGP sequences perform just as well as the commonly used FMDV 2A peptide: IMNV 2A1 and 2A2 [16]; Aalivirus A1/B1 2A1-2A4; Grusopivirus A1/C 2A1-2A3; Limnipivirus A1 2A1, B1 2A2, C1 2A1, D1 2A1-3; Mosavirus B1 2A1 and 2A2 (unpublished data). The advantage of releasing more than one protein from the same ORF suggests that host infection mechanisms in viruses with multiple 2A peptides in their genomes may be more complex [6]. Probing databases with this conserved motif revealed cellular 2A-like sequences within non-LTR retrotransposons (non-LTRs) of Trypanosoma brucei, T. cruzi, T. vivax and T. congolense, and, a range of simple aquatic species: Xenopus tropicalis (African claw-toed frog, vertebrate), Branchiostoma floridae (Amphioxus, Florida lancelet, cephalochordate), Aplysia californica (California sea slug, mollusc), Crassostrea gigas (Pacific oyster, mollusc), Lottia gigantean (Owl limpet, mollusc), Nematostella vectensis (sea anemone, cnidarian) and S. purpuratus (echinoderm) [75, 76, 77]. To allay public fears and bypass the use of animal-virus sequences, non-viral 2A variants (e.g. those found in the California sea slug and purple sea urchin) have been used just as effectively for coordinated multi-protein co-expression [78].
Of the many 2A peptides identified to date, four viral 2As have been widely used in biotechnology and biomedicine: FMDV (“F2A”), equine rhinitis A virus (ERAV, “E2A”), porcine teschovirus-1 (PTV-1, “P2A”), and Thosea asigna virus (TaV, “T2A”) (see Table 2) [27, 54, 59, 79]. Comparing the in vitro activity of different 2As inserted between GFP and GUS, T2A20 has the highest cleavage efficiency followed by E2A20, P2A20 and F2A20 [15]. In human cell lines, mice and zebrafish, cleavage and targeting of NLS-EGFP-2A-mCherry-CAAX was most efficient in P2A19 linked constructs followed by T2A18, E2A20 and F2A22 [43]. To test the performance of selected 2A peptides in insect cell cultures (B.mori) the cleavage efficiency of six types of 2A with a GSG linker were analysed: P2A, T2A, E2A, F2A, BmCPV2A and BmIFV2A. For the most used sequences P2A22 exhibited the highest cleavage efficiency in all insect cell lines tested followed by E2A23, F2A25 and T2A21 [32]. Interestingly, P2A19 and T2A18 functions in Drosophila are approximately equivalent both in cultured cells and in vivo [80]—given their poor efficiency at polypeptide separation, E2A and F2A may not be useful in Drosophila [80, 81]. A study to characterise the 2A system for metabolic engineering applications in Saccharomyces cerevisiae showed that the F2A19, T2A18, and P2A19 sequences are functional in S. cerevisiae cells [82]—earlier analysis in S. cerevisiae indicated that ERBV-1 (Equine rhinitis B virus 1) 2A had the highest cleaving efficiency among 22 viral 2A sequences tested [48]. For potential upcoming amoeba-based bioprocesses codon optimised P2A19, T2A18, E2A20 and F2A22 were screened for activity in Dictyostelium discoideum [45]. In this work P2A and T2A performed the best with cleavage rates of 99.6% and 97.4% respectively, followed by E2A (95.5%), and F2A (66.9%). The availability of a range of 2A sequences can be useful—to minimise the risk of homologous recombination, it is wise to use different 2A peptide sequences if more than two genes are being linked.
Table 2.
2A/2A-like sequences most commonly used for protein co-expression.
Current strategies for multi-gene co-expression include: (i) pre-mRNA splicing; (ii) proteolytic cleavage sites; (iii) fusion proteins; (iv) IRESs (internal ribosomal entry sites); (v) self-cleaving 2A peptides and (vi) a new-kid-on-the-block termed HACKing (Highly efficient and Accessible system by CracKing genes into the genome) [83]. Among the six systems listed, the most popular strategies are 2A/2A-like sequences and viral IRES elements. Like the 2A peptide, the IRES was identified first among picornaviruses but in a different group, the Enteroviruses, typified by poliovirus (PV) [84]. Since then, IRES elements have been identified in other positive-sense RNA viruses such as encephalomyocarditis virus (EMCV) [85], DNA viruses [86, 87] and a growing number of cellular mRNAs involved in growth control, cell cycle progression and apoptosis [88]. In the majority of eukaryotic cells the mRNA 5′ 7meG “cap” is a signal for eukaryotic initiation factors (eIFs) to recruit ribosomes and initiate translation [89]. IRES elements allow the initiation of translation in a cap-independent manner, i.e., ribosomes bind internally without scanning the 5′ UTR of the transcript ensuring co-expression of genes before and after the IRES [90]. Most IRESs require IRES trans-acting factors (ITAFs) to function in addition to several canonical IFs [91]. These cis-acting RNA regulatory elements “hijack” ribosomes of the host cell, redirecting them towards the production of viral proteins [92, 93]. Viral IRESs have been classified into four different types based on their secondary structure and, in turn, their mechanism of initiation: Type I (enteroviruses/rhinoviruses), Type II (cardio-/aphthoviruses), Type III (flaviviruses e.g. HCV) and Type IV (dicistroviruses e.g. CrPV) [94, 95]. It has been shown that the efficiency of translation initiated from different IRES elements differs significantly [96, 97, 98].
Most IRES-based vectors developed up to now use picornavirus IRESs (e.g. EMCV and PV), based on the strong efficiency of such IRESs in transient transfection, compared to cellular IRESs [99, 100]. In mRNAs transcribed from bicistronic constructs, the first cistron is translated by a cap-dependent scanning mechanism while translation of the second cistron depends on the presence of IRES in the intercistronic space. The advantages of IRES-mediated systems include: complete separation of the coupled genes; lack of fusion proteins which may adversely affect the activity of the proteins; the relative expression of different genes can be “fine-tuned” by varying the strength of the IRES applied to each gene. IRES sequences, however, have limitations: (i) the large size (~500 bp) can reduce the packaging capacity in viral vectors [101], (ii) expression of the downstream gene can be as much as 10 fold lower than the upstream gene [102, 103, 104], (iii) recognition of different IRES elements varies with cell type, species, and even cell stress state [91, 97] and lastly (iv) the risk of homologous recombination and competition among different IRESs for translation factors [105].
Nevertheless, different IRES-based vectors including plasmids, adeno-associated virus (AAV)-derived and lentiviral vectors have been used successfully in pre-clinical as well as in clinical gene therapy protocols (reviewed in [106]). In plants, IRES-based constructs have been used to engineer pest resistance in tobacco [107], enhance salt and/or drought tolerance in potato [108] and produce carotenoids in rice endosperm [104]. However, compactness in length and their ability to allow the concatenated genes to be translated at equivalent levels both in vivo and in vitro has seen 2A peptides gain popularity over traditional IRES elements. Adenovirus [109], AAV [110], retrovirus [111], lentivirus [112] and plasmid [64] expression vectors incorporating 2A peptide sequences have been used in a variety of biotechnology and biomedical applications (reviewed in [113, 114, 115]).
7.1 Heterologous expression of multi-gene biosynthetic pathways in yeast
A particular advantage of using 2A co-expression technology is the ability to consistently express all the required genes in the same cell. For example, in biosynthetic pathways involving a number of separate enzymes it is critical they are all expressed in the same cell. If this is not the case, instead of progressing from reactants to products a breakdown in the pathway could lead to a build-up of volatile intermediates. Enzymatic systems requiring several cofactors have been reconstructed, by means of 2A peptides, both in Pichia pastoris and S. cerevisiae [48, 116, 117]. Significant advantages of the P. pastoris expression system include post-translational modifications such as glycosylation, proper protein folding and secretion (by Kex2 as signal peptidase) of recombinant proteins [118]. In the early days, environmental stress conditions imposed on the host presented a potential bottleneck in terms of recombinant protein production and secretion. Particular attention has been paid to the role of glycine betaine (GB) synthesis in abiotic stress resistance [119]. The GB pathway is a two-step oxidation from choline and betaine aldehyde, catalysed by choline monooxygenase (CMO) and betaine aldehyde dehydrogenase (BADH), respectively. The genes encoding the two enzymes of GB synthesis in the halophyte Suaeda salsa were cloned and fused with the F2A in a single ORF. The fused genes “CMO-2A-BADH” transformed in P. pastoris were expressed successfully and the polyprotein was cleaved to each functional protein [CMO-2A] and [BADH]. The recombinant yeasts were more tolerant to salt, methanol and high temperature stresses [120].
Biobased manufacturing for the production of fuels has become increasingly popular as the serious effects of global climate change become apparent. Using a viral 2A sequence, P. pastoris was engineered to produce the biofuels isobutanol and 3-methyl-1-butanol (3M1B, isoamyl alcohol). Upregulation of the endogenous L-valine biosynthetic pathway (PpIlv5, PpIlv3, PpIlv6, PpIlv2) and heterologous expression of the keto-acid degradation pathway (LlkivD, ScADH7) led to an engineered strain capable of producing isobutanol [121]. Specifically, the strain overexpressing all six genes produced a 100-fold improvement over the levels observed in the strains overexpressing only the two keto acid degradation pathway genes. In a follow on study, engineered strains overexpressing the keto-acid degradation pathway as well as the upper portion of the amino acid L-valine and L-leucine (PpLeu1, PpLeu2, PpLeu6) biosynthetic pathways were able to produce the platform chemical 3M1B [122]. Compared to the predominant biofuel ethanol, the advantages of 3M1B can be summarised as higher energy density, better miscibility with fossil fuels, lower water solubility, and greater applicability to conventional engines. The potential of this system is vividly demonstrated by the expression of nine genes from a single polycistronic transcript based on T2A peptides—presently, the construct with the highest number of genes expressed in a coordinated fashion. This expression strategy resulted in P. pastoris strains producing the target products of both the carotenoid (CrtE, CrtB, CrtI,CrtY) and violacein (vioC, vioB, vioE, vioD, vioA) biosynthesis pathways [47].
To date, a variety of studies have examined the use of multicomponent enzymatic systems based on the 2A approach in the baker’s yeast S. cerevisiae [123, 124, 125, 126, 127, 128]. The utility of 2A peptides has been demonstrated in several metabolic engineering applications, such as the production of C-glucosylflavones or β-carotene. Flavonoids are important phytochemical compounds with a range of medical benefits including powerful activities as antioxidants. In rice plants, two enzymes are responsible for the biosynthesis of flavone-C-glycosides, the cytochrome P450 flavanone 2-hydroxylase (F2H) and C-glucosyltransferase (CGT). While pathway engineering in tobacco resulted in minor C-glycoside formation, yeast transformed with the [F2H-F2A-CGT] polyprotein produced high concentrations of 2-hydroxynaringenin which could be converted chemically to the “nature-identical” plant product [129]. “Carotenoids” is a generic term used to designate a diverse group of pigments widely distributed in nature. Chemical synthesis of the flavour and fragrance (apo-) carotenoid β-ionone is economically impractical. For de novo production of apocarotenoids in yeast, three β-carotene biosynthesis genes from the carotenoid-producing ascomycete Xanthophyllomyces dendrorhous (crtI, crtE and crtYB) were co-expressed in S. cerevisiaefrom a single T2A polycistronic construct. To enable β-ionone production, a carotenoid-cleavage dioxygenase from raspberry (RiCCD1) was co-expressed in the β-carotene producing strain - a novel microbial production system for a fruit flavour compound [130].
7.2 Engineering carotenoid biosynthesis in plants
Conventional methods to “stack” transgenes in plants include sexual crossing, sequential re-transformation and single/multiple plasmid co-transformation [131]. These strategies, however suffer from the inherent weakness that co-expression of the heterologous proteins is unreliable. 2A/2A-like constructs have been successfully used to co-express β- and δ-zein proteins to improve the nutritional value of tobacco plants [132]; mpi (maize proteinase inhibitor) and pci (potato carboxypeptidase inhibitor) for insect and pathogen resistance in rice [133] and Baccillus thuringiensis (Bt) crystal proteins, Cry1Ab and Cry2Ab to delay insect resistance to Bt toxins in rice [134]. In a recent study, 11 genes were successfully introduced into transgenic maize to rebuild the anthocyanin biosynthesis pathway (ZmBz1, ZmBz2, ZmC1 and ZmR2) by combining the advantages of a bidirectional promoter (BDP) and 2A linker peptides [135]. Carotenoids have attracted interest not only as a source of pigmentation but also for their beneficial effects on human health—since most animals are unable to synthesise carotenoids, they need to obtain them from food and dietary supplements. To alleviate vitamin A deficiency (VAD) through food intake, biofortified “Golden Rice” (Oryza sativa, GR) was engineered via two gene cassettes for the expression of the phytoene synthase (Psy) and carotene desaturase (CrtI) genes to produce β-carotene (provitamin A) (GR1 [136], GR2 [137]). Among GR varieties, “Korean Golden Rice” was developed using the recombinant PAC gene that linked the Capsicum Psy and Pantoea CrtI genes (PAC, Psy-F2A-CrtI) [104]. To increase the levels of carotenoids, the DNA sequences of these two carotenoid biosynthetic genes originating from heterologous sources, Capsicum annum for Psy and Pantoea ananatis for CrtI, were codon optimised for rice plants (pstPAC) [138]. In a separate study, Kim et al. genetically manipulated the carotenoid biosynthetic pathway in Korean soybean using an efficient Agrobacterium-mediated transformation method and the PAC recombinant gene to produce transformants whose β-carotene levels exceeded those reported in transgenic rice plants [139].
Zeaxanthin, capsanthin, canthaxanthin and astaxanthin are high-value carotenoids used industrially as colourants and feed supplements. Their antioxidant properties orchestrate important pathways in animals, including immuno/photo protection and oxidative stress modulation [140]. In both prokaryotes and eukaryotes, Astaxanthin is derived from β-carotene by hydroxylation and ketolation reactions catalysed by β-carotene hydroxylase (BCH) and β-carotene ketolase (BKT), respectively. In early studies of ketocarotenoid production, both genes from a marine bacterium (Paracoccus sp) were expressed simultaneously as a F2A polyprotein in both tobacco and tomato plants [141]. β-Carotene can typically be increased by the introduction of one or two transgenes, however producing diverse carotenoids beyond β-carotene (e.g. zeaxanthin and ketocarotenoids) typically requires the simultaneous engineering of a greater number of transgenes [142]. To produce downstream carotenoids, Ha and colleagues adopted three steps of the pathway to produce Zeaxanthin—a BCH expression cassette, under the control of the rice globulin promoter, combined with a PAC gene giving rise to a single T-DNA vector (pB-PAC; Bch and Psy-2A-CrtI). Astaxanthin was produced by the introduction of four steps (PSY, CRTI, BCH and BKT) using a single T-DNA vector (pBAK-PAC) with two cassettes for two bicistronic genes (BAK, Bch-F2A-Bkt and PAC, Psy-F2A-CrtI). The single T-DNA vectors resulted in the accumulation of zeaxanthin and astaxanthin in the endosperm of the transgenic rice seeds [143]. In a later study, bi-, tri and quadcistronic 2A systems for co-expression of these four genes were tested in transgenic rice seeds. To appease public concerns, three highly active 2A variants not found in mammalian viruses, T2A20, IMNV2A130 and IMNV2A230 were used for the coordinated expression of multiple transgenes. This study demonstrated successful expression of two, three and four genes resulting in stepwise biosynthesis of β-carotene, zeaxanthin, adonixanthin and astaxanthin [144].
7.3 The broad utility of 2A and 2A-like sequences
Previous and more recent works employed the 2A approach to assemble biosynthetic pathways for heterologous expression in filamentous fungi. For bioproduct synthesis, the P2A peptide was employed to reconstitute synthesis of β-lactam antibiotics by moving the genes for the complete penicillin biosynthesis pathway from Penicillium chrysogenum (pcbAB, pcbC, and penDE) to the fungal host Aspergillus nidulans [145]. A similar approach was also used by Hoefgen and colleagues to produce the psychotropic mushroom alkaloid psilocybin by heterologous expression of the entire biosynthetic gene cluster (psiH-psiD-psiK-psiM) in A. nidulans [49]. A 2A peptide-based bicistronic protein expressing platform was used to monitor transgene expression (Cel7A, cellobiohydrolase) in the cellulase producing fungus Trichoderma reesei using eGFP as a marker (Cel7A-F2A-eGFP or eGFP-F2A-Cel7A) [38]. To date, 2A peptides have been used in various artificial reporter polyprotein systems comprising chloramphenicol acetyl-transferase (CAT), GUS and fluorescent proteins (FPs, e.g. GFP, RFP, YFP) in various cell types, as well as targeting to various subcellular localisations in plant [52, 146] and animal cells [56, 59]. Targeted endonucleases including zinc finger nucleases (ZFN), transcription activator-like effector nucleases (TALENs) and clustered regularly interspaced short palindromic repeats (CRISPRs) are increasingly being used for genome editing (GE) in numerous cell types and species. GE by using 2A peptide coupled co-expression of nuclease and fluorescent proteins combined with fluorescence-activated cell sorting (FACS) can aid selective enrichment of transfected cells. This approach to achieve higher genome editing efficiencies works well for these three major nucleases [147, 148]. To expand the experimental toolkit, antibiotic (hygromycin) and magnetic H-2KK cell selection methods have been developed to enrich transfected cells with ZFN/TALEN-induced mutations (2A-HygroR-eGFP and eGFP-2A-H-2Kk) [149]. In GE, the two colour “Traffic Light Reporter” (TLR; eGFP-T2A-mCherry) developed by Scharenberg and colleagues has proved invaluable for evaluating the two major DNA repair pathways, nonhomologous end-joining (NHEJ) and homologous recombination (HDR) [150]. Developing new methodologies governing CRISPR-Cas activity using 2A sequences include genetic switches [151], logic gates [152], and gene regulation circuits [153].
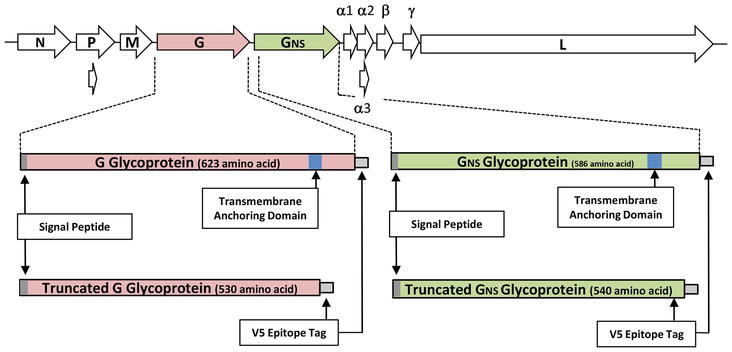
New types of cancer treatment via gene therapy promise to revolutionise management of the disease. Here, 2A is used to co-express the T-cell receptor TCRα and TCRβ chains (in transformed patient T-cells: ex vivo gene therapy) targeted against cancer antigens. Use of 2A sequences ensures both subunits of the receptor can be co-expressed at similar levels, minimising side effects. A second major impact in human health has been in the rapidly expanding field of regenerative medicine and transplantation. 2A has played a pivotal role in the co-expression of the multiple transcription factors required to produce induced pluripotent stem cells (iPSCs)—one can now make patient-specific stem cells relatively simply to avoid immune response problems and/or organ rejection (reviewed in [115]). Many other biotechnological applications that depend on the co-expression of genes use 2A/2A-like sequences e.g. the production of antibodies and antigens that can be used in vaccine production [64, 154, 155]. Bovine ephemeral fever virus (BEFV) is an economically important pathogen that causes an acute febrile illness of cattle and water buffalo in many parts of the world (Africa, the Middle East, Australia and Asia). Various studies have been conducted to develop an efficient vaccine for BEFV, including live attenuated, subunit and recombinant vaccines [156]. Although experimental and commercial BEFV vaccines have been developed, these suffer from either genetic instability, low efficacy, the need for multiple doses, or, are simply too expensive under field conditions. The 14.9 kb (−ve) ssRNA genome of BEFV contains 10 genes in the order 3′-N-P-M-G-GNS-α1-α2-β-γ-L-5′ (Figure 4) [157]. G protein, as a class I transmembrane surface glycoprotein, is the target of neutralising antibodies and a prime candidate for subunit vaccine design [158]. In a recent study, we developed an antigen expression system to express recombinant G/GNS glycoproteins with the transmembrane anchor domain deleted, enabling truncated proteins to be secreted into tissue culture media after mammalian cell transfection [65]. The V5 epitope tag was genetically fused to the C-termini of the proteins for protein detection and purification of expressed proteins from the cell media (Figure 4). Furthermore, a genetic fusion of each of the virus glycoproteins with GFP via T2A was used for live-cell fluorescence microscopy. In this “proof-of-principle” study, truncated G glycoprotein was detected throughout the exocytic pathway and secreted efficiently from BHK cells into the cell media. In a follow-on study we evaluated the immunogenicity of a secreted, C-terminally truncated form of bovine viral diarrhoea virus (BVDV) E2 glycoprotein in mice [66]. BVDV is the causative agent of one of the most widespread and economically important virus infections in cattle [159]. Glycoprotein E2 plays a key role in host cell immune responses to viral infection [160]. Here, truncated E2 glycoprotein purified from cell media was found to induce both humoral and cellular immune responses in BALB/c mice. This antigen expression system provides both a simple purification protocol along with a feasible strategy for further, large-scale, production of vaccines.
Figure 4.
Genome structure of BEFV and synthesis of the regions encoding the G and GNS glycoproteins. Structural organisation of the BEFV genome shown as arranged in negative sense. The BEFV genome encodes five structural proteins including a nucleoprotein (N), a polymerase-associated protein (P), a matrix protein (M), a large RNA-dependent RNA polymerase (L) and a glycoprotein (G) spanning the viral envelope and a non-structural glycoprotein (GNS) followed by a series of “accessory” proteins. Regions encoding the full-length G and GNS proteins (boxed areas) are shown, together with the signal peptide (grey shaded areas) and the transmembrane anchoring domains (blue shaded areas). Regions encoding the C-terminally truncated (transmembrane anchoring domains deleted) forms of G and GNS proteins (boxed areas) are shown, together with the signal peptide (grey shaded areas) and the C-terminal V5 epitope tag (white boxes).
A particularly useful aspect of the ribosome skipping mechanism is the possibility of differentially targeting the proteins produced from a single gene. For example, Park and colleagues used three 2A peptides (F2A, T2A and E2A) to simultaneously express four distinct fluorescent protein variants targeted to subcellular compartments such as mitochondria, nuclei, endosomes, and membranes for real-time monitoring of cellular dynamics in HeLa cells [161]. To facilitate studies of subcellular Ca2+ signalling in insulin-secreting pancreatic beta cells, particularly, signalling between the ER and mitochondria, Jeyarajan et al. [162] designed a plasmid that contained the sequences of two different Ca2+ reporter molecules, G-CEPIA-er and R-CEPIA3-mt, separated by T2A. Successful segregation of the two probes to their respective organelles allowed simultaneous measurement of changes in free Ca2+ to be made using live cell imaging in real time. 2A peptides were also used for the co-expression and differential subcellular targeting of two fluorescent marker proteins in insect cell culture systems (Drosophila melanogaster S2 cells) and Drosophila suzukii cell lines [163]. All four 2A peptides tested (F2A31, F2A32, T2A, DCV2A) showed comparable activity in cell lines, leading to the production of independent upstream and downstream proteins that were directed to the nucleus or membrane by a C-terminal nuclear localisation signal (NLS) on the upstream protein and a poly-lysine/CAAX membrane anchor on the downstream protein.
In prokaryotes proteins with linked functions (e.g. the formation of macromolecular structures, biochemical pathways) can be expressed in a co-ordinated manner by encoding multiple proteins as polycistronic mRNAs: a single RNA is transcribed from different genes organised—concatenated—into operons. This is only possible since prokaryotic ribosomes initiate translation by binding individual coding regions within the (polycistronic) mRNA. In contrast, eukaryotes initiate translation only at a single site—the 7meG “cap” structure at the 5′end of mRNA. Here, expression of proteins is achieved by the co-ordinated transcription of the individual genes located at different sites throughout the genome. The model of one gene one enzyme/one gene one polypeptide developed in the 1940s was substantially modified in the late 1960s from studies on the biogenesis of poliovirus proteins and insulin. In both these cases it was proposed proteins were formed by post-translational processing of a single precursor (poly)protein [164, 165, 166, 167]. Encoding multiple proteins in the form of a polyprotein is a common, if not ubiquitous, strategy of viruses with positive stranded (mRNA sense) genomes. Such polyproteins are proteolytically “processed” by virus-encoded proteinases, although host-cell proteinases may also be utilised in processing virus polyproteins.
Viruses have evolved many different mechanisms to both maximise the coding capacity of their genomes and to generate multiple, different, proteins from a single ORF—many involving manipulating or modifying the host-cell translational apparatus [2]. In the case of 2A-mediated ribosome skipping, the strategy has been very widely used in biotechnology and biomedicine in the generation of transgenic cell lines/organisms as outlined above. Being able to concatenate genes in this manner has simplified and accelerated the whole process of producing transgenics: reducing, or even eliminating, the need for re-iterative rounds of transformation/selection required to introduce “traits” based upon multiple gene expression. The range of applications is immense, spanning humans, animals, plants and fungi. To conclude, the study of FMDV polyprotein processing has provided a valuable tool that has allowed researchers to create “artificial” polyproteins such that multiple, different, proteins can be generated from a single (poly)cistronic construct—driven by a single promoter. Furthermore, since 2A-mediated recoding occurs co-translationally, products can be targeted to different combinations of sub-cellular sites or secreted from the cell. Over 1500 research publications (https://www.st-andrews.ac.uk/ryanlab/Index.html) and many patent/patent applications (https://www.freepatentonline.com/login.html) stand testament to the influence 2As have had on co-expression technology—this is not the end, this is not even the beginning of the end, this is just perhaps the end of the beginning.
The long term support of the Wellcome Trust and the Biotechnology and Biological Sciences Research Council (BBSRC) is gratefully acknowledged. The University of St Andrews is a charity registered in Scotland no. SCO13532.
clustered regularly interspaced palindromic repeats – associated protein 9
CrPV
cricket paralysis virus
DBM
diamond back moth
DBMIV
diamondback moth iflavirus
DCV
Drosophila C virus
DERSV
duck egg-reducing syndrome virus
DNA
deoxyribonucleic acid
EoPV
ectropis oblique picorna-like virus
EMCV
encephalomyocarditis virus
eIFs
eukaryotic initiation factors
ER
endoplasmic reticulum
ERAV
equine rhinitis A virus
ERBV-1
equine rhinitis B virus 1
FACS
fluorescence-activated cell sorting
FMDV
foot-and-mouth disease virus
FPs
fluorescent proteins
GE
genome editing
GFP
green fluorescent protein
GR
golden rice
GUS
glucuronidase
HACKing
highly efficient and accessible system by cracking genes into the genome
HCV
hepatitis C virus
ICC
immunocytochemistry
IMNV
infectious myonecrosis virus
IP
immunoprecipitation
iPSCs
induced pluripotent stem cells
IRES
internal ribosome entry site
ITAFs
IRES trans-acting factors
mRNA
messenger RNA
NLS
nuclear localisation signal
OMRV
Omono River virus
ORF
open reading frame
OpbuCPV18
Operophtera brumata cypovirus-18
PnPV
Perina nuda picorna-like virus
PTV-1
porcine teschovirus-1
PV
poliovirus
RFP
red fluorescent protein
RtPV
Rattus tanezumi parechovirus
RNA
ribonucleic acid
SRP
signal recognition particle
TALEN
transcription activator-like effector nucleases
TaV
Thosea asigna virus
TCR
T-cell receptor
TEV
tobacco etch virus
TLR
Traffic Light Reporter
ToV-TJ
Tianjin totivirus
tRNA
transfer RNA
VAD
vitamin A deficiency
WP-LV
Wenzhou picorna-like virus
WCP
Wuhan carp picornavirus
YFP
yellow fluorescent protein
ZFN
zinc finger nucleases
References
1.Gesteland RF, Atkins JF. Recoding: Dynamic reprogramming of translation. Annual Review of Biochemistry. 1996;65:741-768
2.Ryan MD, Luke GA. Translation of viral proteins. In: Bamford, Zuckerman M, editors. Encyclopedia of Virology. 4th ed. Vol. 1. London, New York: Elsevier; 2021. pp. 444-459. DOI: 10.1016/8978-0-12-814515-9.00119-3
3.Donnelly MLL, Luke GA, Mehrotra A, Li X, Hughes LE, Gani D, et al. Analysis of the aphthovirus 2A/2B polyprotein “cleavage” mechanism indicates not a proteolytic reaction, but a novel translational effect: A putative ribosomal “skip”. Journal General Virology. 2001;82:1013-1025
4.Atkins JF, Wills NM, Loughran G, Wu CY, Parsawar K, Ryan MD, et al. A case for “StopGo”: reprogramming translation to augment codon meaning of GGN by promoting unconventional termination (Stop) after addition of glycine and then allowing continued translation (Go). RNA. 2007;13:803-810
5.Brown JD, Ryan MD. Ribosome “skipping”: “Stop-carry on” or “StopGo” translation. In: Atkins JF, Gesteland RF, editors. Recoding: Expansion of Decoding Rules Enriches Gene Expression. New York: Springer; 2010. pp. 101-122
6.Yang X, Cheng A, Wang M, Jia R, Sun K, Pan K, et al. Structures and corresponding functions of five types of picornaviral 2A proteins. Frontiers in Microbiology. 2017;8:1373. DOI: 10:3389/fmicb.2017.01373
7.Ryan MD, Belsham GJ, King AM. Specificity of enzyme-substrate interactions in foot-and-mouth disease virus polyprotein processing. Virology. 1989;173(1):35-45
8.Belsham GJ. Distinctive features of the foot-and-mouth disease virus, a member of the picornavirus family; aspects of virus protein synthesis, protein processing and structure. Progress in Biophysics and Molecular Biology. 1993;60:241-260
9.Ryan MD, Drew J. Foot-and-mouth disease virus 2A oligopeptide mediated cleavage of an artificial polyprotein. The EMBO Journal. 1994;13:928-933
10.Sharma P, Yan F, Doronina VA, Escuin-Ordinas H, Ryan MD, Brown JD. 2A peptides provide distinct solutions to driving stop-carry on translational recoding. Nucleic Acids Research. 2012;40(7):3143-3151
11.Donnelly MLL, Gani D, Flint M, Monaghan S, Ryan MD. The cleavage activities of aphthovirus and cardiovirus 2A proteins. Journal of General Virology. 1997;78:13-21
12.Ryan MD, King AM, Thomas GP. Cleavage of foot-and-mouth disease virus polyprotein is mediated by residues located within a 19 amino acid sequence. Journal of General Virology. 1991;72:2727-2732
13.Varshavsky A. The N-end rule. Cell. 1992;69(5):725-735. ISSN: 0092-8674
14.de Felipe P, Ryan MD. Targeting of proteins derived from self-processing polyproteins containing multiple signal sequences. Traffic. 2004;5:616-626. DOI: 10.1111/j.1398-9219. 2004.00205.x
15.Donnelly MLL, Hughes LE, Luke GA, Mendoza H, ten Dam E, Gani D, et al. The “cleavage” activities of foot-and-mouth disease virus 2A site-directed mutants and naturally occurring “2A-like” sequences. Journal of General Virology. 2001;82:1027-1041
16.Luke GA, de Felipe P, Lukashev A, Kallioinen SE, Bruno EA, Ryan MD. The occurrence, function, and evolutionary origins of “2A-like” sequences in virus genomes. Journal of General Virology. 2008;89(4):1036-1042
17.Doronina VA, Wu C, de Felipe P, Sachs MS, Ryan MD, Brown JD. Site-specific release of nascent chains from ribosomes at a sense codon. Molecular and Cellular Biology. 2008;28(13):4227-4239
19.Gao Z, Zhou J, Zhang J, Ding Y, Liu Y. The silent point mutations at the cleavage site of 2A/2B have no effect on the self-cleavage activity of 2A of foot-and-mouth disease virus. Infection, Genetics and Evolution. 2014;28:101-106
20.Ryan MD, Donnelly MLL, Lewis A, Mehrotra AP, Wilkie J, Gani D. A model for nonstoichiometric, co-translational protein scission in eukaryotic ribosomes. Bioorganic Chemistry. 1999;27:55-79
21.Kjær J, Belsham GJ. Selection of functional 2A sequences within foot-and-mouth disease virus; requirements for the NPGP motif with a distinct codon bias. RNA. 2018;24(1):12-17. DOI: 10.1261/rna.063339.117
22.Hahn H, Palmenberg AC. Mutational analysis of the encephalomyocarditis virus primary cleavage. Journal Virology. 1996;70:6870-6875
23.Roulston C, Luke GA, de Felipe P, Ruan L, Cope J, Nicholson J, et al. “2A-like” signal sequences mediating translational recoding: A novel form of dual protein targeting. Traffic. 2016;17(8):923-939
24.Luke GA, Ryan MD. The protein coexpression problem in biotechnology and biomedicine: Virus 2A and 2A-like sequences provide a solution. Future Virology. 2013;8(10):983-996
25.Novoa EM, Ribas de Pouplana L. Speeding with control: Codon usage, tRNAs, and ribosomes. Trends in Genetics. 2012;28:574-581
26.Mauro VP, Chappell SA. A critical analysis of codon optimization in human therapeutics. Trends in Molecular Medicine. 2014;20:604-613
27.Szymczak AL, Workman CJ, Wang Y, Vignali KM, Dilioglou S, Vanin EF, et al. Correction of multi-gene deficiency in vivo using a single “selfcleaving” 2A peptide-based retroviral vector. Nature Biotechnology. 2004;22(5):589-594
29.Wang X, Tian X, Marchisio MA. Logic circuits based on 2A peptide sequences in the yeast Saccharomyces cerevisiae. ACS Synthetic Biology. 2003;12:224-237
30.Tan Y, Liang H, Chen A, Guo X. Coexpression of double or triple copies of the rabies virus glycoprotein gene using a “self-cleaving” 2A peptide-based replication-defective human adenovirus serotype 5 vector. Biologicals. 2010;38:586-593
31.Yang S, Cohen CJ, Peng PD, Zhao Y, Cassard L, Yu Z, et al. Development of optimal bicistronic lentiviral vectors facilitates high-level TCR gene expression and robust tumor cell recognition. Gene Therapy. 2008;15(21):1411-1423
32.Wang Y, Wang F, Wang R, Zhao P, Xia Q. 2A self-cleaving peptide-based multi-gene expression system in the silkworm Bombyx mori. Scientific Reports. 2015;5(5):16273. DOI: doi.org/10.1038/srep16273
33.Wang Y, Wang F, Xu S, Wang R, Chen W, Hou K, et al. Optimization of a 2A self-cleaving peptide-based multigene expression system for efficient expression of upstream and downstream genes in silkworm. Molecular Genetics and Genomics. 2019;294(4):849-859
34.Minskaia E, Ryan MD. Protein coexpression using FMDV 2A: Effect of “linker” residues. BioMed Research International Article. 2013;ID 291730. DOI: 10.1155/2013/291730
35.Minskaia E, Nicholson J, Ryan MD. Optimisation of the foot-and-mouth disease virus 2A co-expression system for biomedical applications. BMC Biotechnology. 2013;13:67. Available from: http://www.biomedcentral.com/1472-6750/13/67
36.Ho SC, Bardor M, Li B, Lee JJ, Song Z, Tong YW, et al. Comparison of internal ribosome entry site (IRES) and furin-2A (F2A) for monoclonal antibody expression level and quality in CHO cells. PLoS One. 2013;8:e63247
37.Liu Z, Chen O, Wall JBJ, Zheng M, Zhou Y, Wang L, et al. Systematic comparison of 2A peptides for cloning multi-genes in a polycistronic vector. Scientific Reports. 2017;7:2193
38.Subramanian V, Schuster LA, Moore KT, Taylor LE, Baker JO, Vander Wall TA, et al. A versatile 2A peptide-based bicistronic protein expressing platform for the industrial cellulase producing fungus, Trichoderma reesei. Biotechnology for Biofuels. 2017;10:34. DOI: 10.1186/s13068-017-0710-7
39.Jiao X, Sun W, Zhang Y, Lin X, Zhang Q , Wang Q , et al. Exchanging the order of carotenogenic genes linked by porcine teschovirus-1 2A peptide enable to optimize carotenoid metabolic pathway in Saccharomyces cerevisiae. Royal Society of Chemistry Advances. 2018;8:34967
40.Ryan MD, Luke GA, Hughes LE, Cowton VM, ten Dam E, Li X, et al. The aphtho-and cardiovirus “primary” 2A/2B polyprotein “cleavage”. In: Semler BL, Wimmer E, editors. Molecular Biology of Picornaviruses. Washington DC: ASM Press; 2002. pp. 213-223
41.Doronina VA, de Felipe P, Wu C, Sharma P, Sachs MS, Ryan MD, et al. Dissection of a co-translational nascent chain separation event. Biochemical Society Transactions. 2008;36(4):712-716
42.Machida K, Mikami S, Masutani M, Mishima K, Kobayashi T, Imataka H. A translation system reconstituted with human factors proves that processing of encephalomyocarditis virus proteins 2A and 2B occurs in the elongation phase of translation without eukaryotic release factors. Journal of Biological Chemistry. 2014;289(46):31960-31971
43.Kim JH, Lee SR, Li L-H, Park H-J, Park J-H, Lee KY, et al. High cleavage efficiency of a 2A peptide derived from porcine teschovirus-1 in human cell lines, zebrafish and mice. PLOS One. 2011;6(4):e18556. DOI: 10.1371/journal.pone.0018556
44.Chng J, Wang T, Nian R, Lau A, Hoi KM, Ho SC, et al. Cleavage efficient 2A peptides for high level monoclonal antibody expression in CHO cells. MAbs. 2015;7:403-412. DOI: 10.1080/19420862.2015.1008351
45.Zhu X, Ricci-Tam C, Hager ER, Sgro AE. Self-cleaving 2A peptides for expression of multiple genes in Dictyostelium discoideum. PLoS One. 2023;18(3). DOI: 10.1371/journal.pone.0281211
46.de Felipe P, Hughes LE, Ryan MD, Brown JD. Co-translational, intraribosomal cleavage of polypeptides by the foot-and-mouth disease virus 2A peptide. The Journal of Biological Chemistry. 2003;13:11441-11448
47.Geier M, Fauland P, Vogl T, Glieder A. Compact multi-enzyme pathways in P. pastoris. Chemical Communications. 2015;51:1643-1646
48.Souza-Moreira TM, Navarrete C, Chen X, Zanelli CF, Valentini SR, Furlan M, et al. Screening of 2A peptides for polycistronic gene expression in yeast. FEMS Yeast Research. 2018;18(5). DOI: 10.1093/femsyr/foy036
49.Hoefgen S, Lin J, Fricke J, Stroe MC, Mattern DJ, Kufs JE, et al. Facile assembly and fluorescence-based screening method for heterologous expression of biosynthetic pathways in fungi. Metabolic Engineering. 2018;48:44-51
50.Rasala BA, Chao S-S, Pier M, Barrera DJ, Mayfield SP. Enhanced genetic tools for engineering multigene traits into green algae. PLoS One. 2014;9(4):e94028. DOI: 10.1371/journal.pone.0094028
51.Koh HG, Kang NK, Kim EK, Jeon S, Shin S-E, Lee B, et al. Advanced multigene expression system for Nannochloropsis Salina using 2A self-cleaning peptides. Journal of Biotechnology. 2018;278:39-47
52.Burén S, Ortega-Villasante C, Ötvös K, Samuelsson G, Bakó L, Villarejo A. Use of the foot-and-mouth disease virus 2A peptide co-expression system to study intracellular protein trafficking in Arabidopsis. PLoS One. 2012;7(12):eS1973. DOI: 10.1371/journal.pone.0051973
53.Zhang B, Rapolu M, Kumar S, Gupta M, Liang Z, Han Z, et al. Coordinated protein co-expression in plants by harnessing the synergy between an intein and a viral 2A peptide. Plant Biotechnology Journal. 2017;15:718-728. DOI: 10.1111/pbi.12670
54.Provost E, Rhee J, Leach SD. Viral 2A peptides allow expression of multiple proteins from a single ORF in transgenic zebrafish embryos. Genesis. 2007;45(10):625-629
55.Jeong I, Kim E, Seong JY, Park HC. Overexpression of Spexin 1 in the dorsal habenula reduced anxiety in zebrafish. Frontiers in Neural Circuits. 2019;13:1-8. DOI: doi.org/10.3389/fncir2019.00053
56.Grabert K, Sehgal A, Irvine KM, Wollscheid-Lengeling E, Ozdemir DD, Stables J, et al. A transgenic line that reports CSFIR protein expression provides a definitive marker for the mouse mononuclear phagocyte system. The Journal of Immunology. 2020;205(11):3154-3166. DOI: 10.4049/jimmunol.2000835
57.El Amrani A, Barakate A, Askari BM, Li X, Roberts AG, Ryan MD, et al. Coordinate expression and independent subcellular targeting of multiple proteins from a single transgene. Plant Physiology. 2004;135:16-24
58.Yan J, Wang H, Xu Q , Jain N, Toxavidis V, Tigges J, et al. Signal sequence is still required in genes downstream of “autocleaving” 2A peptide for secretary or membrane-anchored expression. Analytical Biochemistry. 2010;399:144-146
59.Trichas G, Begbie J, Srinivas S. Use of the viral 2A peptide for bicistronic expression in transgenic mice. BioMed Central BMC Biology. 2008;6:40. ISSN: 1741-7007
60.Kumagai H, Matsuura T, Kato Y, Watanabe H. Development of a bicistronic expression system in the branchiopod crustacean Daphnia magna. Genesis. 2017;55:e23083. DOI: 10.1002/dvg.23083
61.Mukherjee M, Wang ZQ. A well-characterized polycistronic-like gene expression system in yeast. Biotechnology and Bioengineering. 2023;120:260-271
62.Fang J, Qian JJ, Yi S, Harding TC, Tu GH, VanRoey M, et al. Stable antibody expression at therapeutic levels using the 2A peptide. Nature Biotechnology. 2005;23(5):584-590
63.François IEJA, van Hemelrijck W, Aerts AM, Wouters PFJ, Proost P, Broekaert WF, et al. Processing in Arabidopsis thaliana of a heterologous polyprotein resulting in differential targeting of the individual plant defensins. Plant Science. 2004;166:113-121. ISSN: 0168-9452
64.Mizote Y, Masumi-Koizumi K, Katsuda T, Yamaji H. Production of an antibody fab fragment using 2A peptide in insect cells. Journal of Bioscience and Bioengineering. 2020;130(2):205-211
65.Lo Y-T, Tulloch F, Wu H-C, Luke GA, Ryan MD, Chu C-Y. Expression and immunogenicity of secreted forms of bovine ephemeral fever virus glycoproteins applied to subunit vaccine development. Journal of Applied Microbiology. 2021;131(3):1123-1135. DOI: 10.1111/jam.15044
66.Lo Y-T, Ryan MD, Luke GA, Chang W-C, Wu H-C. Immunogenicity of a secreted, C-terminally truncated, form of bovine viral diarrhea virus E2 glycoprotein as a potential candidate in subunit vaccine development. Scientific Reports. 2023;13:296. DOI: 10.1038/s41598-022-26766-y
67.de Felipe P, Luke GA, Hughes LE, Gani D, Halpin C, Ryan MD. E unum pluribus: Multiple proteins from a self-processing polyprotein. Trends in Biotechnology. 2006;24(2):68-75. ISSN: 0167-7799
68.de Felipe P, Luke GA, Brown JD, Ryan MD. Inhibition of 2A-mediated “cleavage” of certain artificial polyproteins bearing N-terminal signal sequences. Biotechnology Journal. 2010;5(2):213-223. ISSN: 1860-6768
69.Lengler J, Holzmüller H, Salmons B, Günzburg WH, Renner M. FMDV – 2A sequence and protein arrangement contribute to functionality of CYP2B1–reporter fusion protein. Analytical Biochemistry. 2005;343:116-124. DOI: 10.1016/j.ab.2005.05.004
70.Rothwell DG, Crossley R, Bridgeman JS, Sheard V, Zhang Y, Sharp TV, et al. Functional expression of secreted proteins from a bicistronic retroviral cassette based on foot-and-mouth disease virus 2A can be position dependent. Human Gene Therapy. 2010;21:1631-1637. DOI: 10.1089/hum.2009.197
71.Reinhardt A, Kagawa H, Woltjen K. N-terminal amino acids determine KL4F protein stability in 2A peptide-linked polycistronic reprogramming constructs. Stem Cell Reports. 2020;14:520-527. DOI: 10.1016/j.stemcr.2020.01.014
72.de Lima JGS, Lanza DCF. 2A and 2A-like sequences: Distribution in different virus species and applications in biotechnology. Viruses. 2021;13:2160. DOI: 10.3390/v13112160
73.de Lima JGS, Teixeira DG, Freitas TT, Lima JPMS, Lanza DCF. Evolutionary origin of 2A-like sequences in Totiviridae genomes. Virus Research. 2019;259:1-9
74.Su X, Shuo D, Luo Y, Pan X, Yan D, Li X, et al. An emerging duck egg-reducing syndrome caused by a novel picornavirus containing seven putative 2A peptides. Viruses. 2022;14:932. DOI: 10.3390/v14050932
75.Heras SR, Thomas MC, García-Canadas M, de Felipe P, García-Perez JL, Ryan MD, et al. L1Tc non-LTR retrotransposons from Trypanosoma cruzi contain a functional viral-like self-cleaving 2A sequence in frame with the active proteins they encode. Cellular and Molecular Life Sciences. 2006;63:1449-1460
76.Odon V, Luke GA, Roulston C, de Felipe P, Ruan L, Escuin-Ordinas H, et al. APE-type non-LTR retrotransposons of multicellular organisms encode virus-like 2A oligopeptide sequences, which mediate translational recoding during protein synthesis. Molecular Biology and Evolution. 2013;30(8):1955-1965
77.Luke GA, Pathania US, Roulston C, de Felipe P, Ryan MD. DxExNPGP – Motives for the motif. Recent Research Developments in Virology. 2014;9:25-42
78.Sun WW, Zhang B, Han Z, Kumar S, Gupta M. Non-viral 2A-like sequences for protein coexpression. Journal of Biotechnology. 2022;358:1-8
79.Osborn MJ, Panoskaltsis-Mortari A, McElmurry RT, Bell SK, Vignali DAA, Ryan MD, et al. A picornaviral 2A-like sequence-based tricistronic vector allowing for high-level therapeutic gene expression coupled to a dual-reporter system. Molecular Therapy. 2005;12:569-574
80.Daniels RW, Rossano AJ, Macleod GT, Ganetzky B. Expression of multiple transgenes from a single construct using viral 2A peptides in Drosophila. PLoS One. 2014;9(6):e100637. DOI: 10.1371/journal.pone.0100637
81.Diao F, White BH. A novel approach for directing transgene expression in Drosophila: T2A–Gal4 in-frame fusion. Genetics. 2012;190:1139-1144
82.Efimova VS, Isaeva LV, Orekhov PS, Bozdaganyan ME, Rubtsov MA, Novikova LA. Using a viral 2A peptide-based strategy to reconstruct the bovine P450scc steroidogenic system in S. cerevisiae. Bovine P450scc system expression using 2A peptides. Journal of Biotechnology. 2021;325:186-195
83.Yue Q , Meng J, Qiu Y, Yin M, Zhang L, Zhou W, et al. A polycistronic system for multiplexed and precalibrated expression of multigene pathways in fungi. Nature Communications. 2023;14:4267. DOI: 10.1038/s41467-023-40027-0
84.Pelletier J, Sonenberg N. Internal initiation of translation of eukaryotic mRNA directed by a sequence derived from poliovirus RNA. Nature. 1988;334:320-325
85.Jang SK, Krausslich HG, Nicklin MJ, Duke GM, Palmenberg AC, Wimmer E. A segment of the 5′ nontranslated region of encephalomycarditis virus RNA detects internal entry of ribosomes during in vitro translation. Journal Virology. 1988;62(8):2636-2643
86.Bieleski L, Talbot S. Kaposi’s sarcoma-associated herpesvirus vCyclin open reading frame contains an internal ribosome entry site. Journal Virology. 2001;75:1864-1869
87.Tahiri-Alaoui A, Smith LP, Baigent S, Kgosana L, Petherbridge LJ, Lambeth LS, et al. Identification of an intercistronic internal ribosome entry site in a Marek’s disease virus immediate-early gene. Journal Virology. 2009;83:5846-5853
88.Jackson RJ, Hellen CU, Pestova TV. The mechanism of eukaryotic translation initiation and principles of its regulation. Nature Reviews Molecular Cell Biology. 2010;11:113-127
89.Hinnebusch AG. The scanning mechanism of eukaryotic translation initiation. Annual Review of Biochemistry. 2014;83:779-812. DOI: 10.1146/annurev-biochem-060713-035802
90.Yamamoto H, Unbehaun A, Spahn CMT. Ribosomal chamber music toward an understanding of IRES mechanisms. Trends in Biochemical Sciences. 2017;42:655-668. DOI: 10.1016/j.tibs2017.06.002
91.Godet AC, David F, Hantelys F, Tatin F, Lacazette E, Garmy-Susini B, et al. IRES trans-acting factors, key actors of the stress response. International Journal of Molecular Science. 2019;20(4):924. DOI: 10.3390/ijms20040924
92.Jan E, Mohr I, Walsh D. A Cap-to-Tail guide to mRNA translation strategies in virus-infected cells. Annual Review of Virology. 2016;3:283-307. DOI: 10.1146/annurev-virology.100114-055014
94.Filbin ME, Kieft JS. Toward a structural understanding of IRES RNA function. Current Opinions in Structural Biology. 2009;19:267-276. DOI: 10.1016/sbi.2009.03.005
95.Johnson AG, Grosely R, Petrov AN, Puglisi JD. Dynamics of IRES-mediated translation. Philosophical Transactions of the Royal Society B: Biological Sciences. 2017;372:20160177. DOI: 10.1098/rstb.2016.0177
96.Borman AM, Bailly JL, Girard M, Kean KM. Picornavirus internal ribosome entry segments: Comparison of translation efficiency and the requirements for optimal internal initiation of translation in vitro. Nucleic Acids Research. 1995;23:3656-3663
97.Borman AM, Le Mercier P, Girard M, Kean KM. Comparison of picornaviral-IRES driven internal initiation of translation in cultured cells of different origins. Nucleic Acids Research. 1997;25:925-932
98.Oumard A, Hennecke M, Hauser H, Nourbakhsh M. Translation of NRF mRNA is mediated by highly efficient internal ribosome entry. Molecular and Cellular Biology. 2000;20:2755-2759
99.Dirks W, Wirth M, Hauser H. Bicistronic transcription units for gene expression in mammalian cells. Gene. 1993;128:247-249
100.Müller P, Oumard A, Wirth D, Kröger A, Hauser H. Polyvalent vectors for coexpression of multiple genes. In: Schleef M, editor. Plasmids for Therapy and Vaccination. Weinheim, Germany: Wiley-VCH; 2001. pp. 119-137
101.Hellen C, Sarnow P. Internal ribosome entry sites in eukaryotic mRNA molecules. Genes and Development. 2001;15:1593-1612
102.Mizuguchi H, Xu Z, Ishii-Watabe A, Uchida E, Hayakawa T. IRES-dependent second gene expression is significantly lower than cap-dependent first gene expression in a bicistronic vector. Molecular Therapy. 2000;1:376-382. ISSN: 1525-0016
103.Martin P, Albagli O, Poggi MC, Boulukos KE, Pognonec P. Development of a new bicistronic retroviral vector with strong IRES activity. BMC Biotechnology. 2006;6:4
104.Ha S-H, Liang YS, Jung H, Ahn M-J, Suh S-C, Kweon S-J, et al. Application of two bicistronic systems involving 2A and IRES sequences to the biosynthesis of carotenoids in rice endosperm. Plant Biotechnology Journal. 2010;8:928-938. ISSN: 1467-7644
105.Douin V, Bornes S, Creancier L, Rochaix P, Favre G, Prats AC, et al. Use and comparison of different internal ribosome entry sites (IRES) in tricistronic retroviral vectors. BMC Biotechnology. 2004;4:16. DOI: 10.1186/1472-6750-4-16
106.Renaud-Gabardos E, Hantelys F, Morfoisse F, Chaufour X, Garmy- Susini B, Prats AC. Internal ribosome entry site-based vectors for combined gene therapy. World Journal of Experimental Medicine. 2015;5(1):11-20. ISSN: 2220-315X
107.Urwin P, Zubko E, Atkinson H. The biotechnological application and limitation of IRES to deliver multiple defence genes to plant pathogens. Physiological and Molecular Plant Pathology. 2002;61:103-108
108.El-Banna A, Hajiezaei MR, Wissing J, Ali Z, Vass L. Over-expression of PR-10a leads to increased salt and osmotic tolerance in potato cell cultures. Journal of Biotechnology. 2010;150:277-287
109.Garanina EE, Mukhamedshina YO, Salafutdinov II, Kiyasov AP, Lima LM, Reis HJ, et al. Construction of recombinant adenovirus containing picorna-viral 2A-peptide sequence for the co-expression of neuro-protective growth factors in human umbilical cord blood cells. Spinal Cord. 2016;54:423-430
110.Lewis JE, Brameld JM, Hill P, Barrett P, Ebling FJP, Jethwa PH. The use of a viral 2A sequence for the simultaneous over-expression of both the vgf gene and enhanced green fluorescent protein (eGFP) in vitro and in vivo. Journal of Neuroscience Methods. 2015;256:22-29
111.Browne EP. An interleukin-1 beta-encoding retrovirus exhibits enhanced replication in vivo. Journal of Virology. 2015;89:155-164
112.Peng Y, Yang T, Tang X, Chen F, Wang S. Construction of an inducible CRISPR/Cas9 system for CXCR4 gene and demonstration of its effects on MKN-45 cells. Cell Biochemistry and Biophysics. 2020;78:23-30
113.Luke GA. Translating 2A research into practice. In: Agbo EC, editor. Innovations in Biotechnology. Croatia: InTech; 2012. pp. 161-186. DOI: 10.5772/30091
114.Luke GA, Roulston C, Tilsner J, Ryan MD. Growing uses of 2A in plant biotechnology. In: D Ekinci editor. Biotechnology. Croatia: InTech; 2015. p. 165-193. DOI: 10.5772/59878
115.Luke GA, Ryan MD. Therapeutic applications of the “NPGP” family of viral 2As. Reviews in Medical Virology. 2018;28(6):1-12. DOI: 10.1002/rmv.2001
116.Schwarzhans J-P, Luttermann T, Geier M, Kalinowski J, Friehs K. Towards systems metabolic engineering in Pichia pastoris. Biotechnology Advances. 2017;35:681-710
117.Tian Y, Li Y, Zhao F, Meng C. Engineered Pichia pastoris production of fusaruside, a selective immunomodulator. BMC Biotechnology. 2019;19:37. DOI: 10.1186/s12896-019-0532-8
118.Karbalaei M, Rezaee SA, Farsiani H. Pichia pastoris: A highly successful expression system for optimal synthesis of heterologous proteins. Journal of Cell Physiology. 2020;235(9):5867-5881
119.Zulfiqar F, Ashraf M, Siddique KHM. Role of glycine betaine in the thermotolerance of plants. Agronomy. 2022;12:276. DOI: 10.3390/agronomy1202027
120.Wang S, Yao Q , Tao J, Qiao Y, Zhang Z. Co-ordinate expression of glycine betaine synthesis genes linked by the FMDV 2A region in a single open reading frame in Pichia pastoris. Applied Microbiology Biotechnology. 2007;77:891-899
121.Siripong W, Wolf P, Kusumoputri TP, Downes JJ, Kocharin K, Tanapongpipat S, et al. Metabolic engineering of Pichia pastoris for production of isobutanol and isobutyl acetate. Biotechnology for Biofuels. 2018;11:1. DOI: 10.1186/s13068-017-1003-x
122.Siripong W, Angela C, Tanapongpipat S, Runguphan W. Metabolic engineering of Pichia pastoris for production of isopentanol (3-Methyl-1-butanol). Enzyme and Microbial Technology. 2020;138:109557. DOI: 10.1016/j.enzmictec.2020.109557
123.Park M, Kang K, Park S, Kim YS, Ha S-H, Lee SW, et al. Expression of serotonin derivative synthetic genes on a single self-processing polypeptide and the production of serotonin derivatives in microbes. Applied Microbiology and Biotechnology. 2008;81:43-49
124.Cruz-Teran CA, Tiruthani K, Mischler A, Rao BM. Inefficient ribosomal skipping enables simultaneous secretion and display of proteins in Saccharomyces cerevisiae. ACS Synthetic Biology. 2017;6:2096-2107
125.Verba KA, Agard DA. Protein expression and purification of the Hsp90-Cdc37-Cdk4 kinase complex from Saccharomyces cerevisiae. Bio-Protocol. 2017;7(19):e2563
126.Grzeschik J, Hinz SC, Könning D, Pirzer T, Becker S, Zielonka S, et al. A simplified procedure for antibody engineering by yeast surface display: Coupling display levels and target binding by ribosomal skipping. Biotechnology Journal. 2017;12(2). DOI: 10.1002/biot.201600454
127.Rosowski S, Becker S, Toleikis L, Valldorf B, Grzeschik J, Demir D, et al. A novel one-step approach for the construction of yeast surface display fab antibody libraries. Microbial Cell Factories. 2018;17(1):3
128.Botman D, de Groot DH, Schmidt P, Goedhart J, Teusink B. In vivo characterisation of fluorescent proteins in budding yeast. Scientific Reports. 2019;9(1):2234
129.Brazier-Hicks M, Edwards R. Metabolic engineering of the flavone-C-glycoside pathway using polyprotein technology. Metabolic Engineering. 2013;16:11-20
130.Beekwilder J, van Rossom HM, Koopman F, Sonntag F, Buchhaupt M, Schrader J, et al. Polycistronic expression of β-carotene biosynthetic pathway in Saccharomyces cerevisiae coupled to β-ionone production. Journal of Biotechnology. 2014;192:383-392
131.Rajput R, Naik J, Misra P, Trivedi PK, Pandey A. Gene pyramiding in transgenic plant development: Approaches and challenges. Journal of Plant Growth Regulation. 2023;42:6038-6056. DOI: 10.1007/s00344-022-10760-9
132.Randall J, Sutton D, Ghoshroy S, Bagga S, Kemp JD. Co-ordinate expression of β- and δ- zeins in transgenic tobacco. Plant Science. 2004;167:367-372
133.Quilis J, López-García B, Meynard D, Guiderdoni E, Segundo BS. Inducible expression of a fusion gene encoding two proteinase inhibitors leads to insect and pathogen resistance in transgenic rice. Plant Biotechnology Journal. 2014;12:367-377
134.Zhao Q , Liu M, Tan M, Gao J, Shen Z. Expression of Cry1Ab and Cry2Ab by a polycistronic transgene with a self-cleavage peptide in rice. PLoS One. 2014;9(10):e10006. DOI: 10.1371/journal.pone.0110006
135.Liu X, Yang W, Mu B, Li S, Zhou X, Zhang C, et al. Engineering of “Purple Embryo Maize” with a multigene expression system derived from a bidirectional promoter and self-cleaving 2A peptides. Plant Biotechnology Journal. 2018;16:1107-1109
136.Ye X, Al-Babili S, Klöti A, Zhang J, Lucca P, Beyer P, et al. Engineering the provitamin A (beta-carotene) biosynthetic pathway into (carotenoid-free rice) endosperm. Science. 2000;287(5451):303-305. DOI: 10.1126/science.287.5451.303
137.Paine JA, Shipton CA, Chaggar S, Howells RM, Kennedy MJ, Vernon G, et al. Improving the nutritional value of golden rice through increased pro-vitamin A content. Nature Biotechnology. 2005;23:482-487
138.Jeong YS, Ku H-K, Kim JK, You MK, Lim S-H, Kim J-K, et al. Effect of codon optimization on the enhancement of the β-carotene contents in rice endosperm. Plant Biotechnology Reports. 2017;11:171-179
139.Kim M-J, Kim JK, Kim HJ, Pak JH, Lee J-H, Kim D-H, et al. Genetic modification of the soybean to enhance the β-carotene content through seed-specific expression. PLoS One. 2012;7(10):e48287. DOI: 10.1371/journal.pone.0048287
140.Saini RK, Keum Y-S, Daglia M, Rengasamy KRR. Dietary carotenoids in cancer chemoprevention and chemotherapy: A review of emerging evidence. Pharmacological Research. 2020;157:104830
141.Ralley L, Enfissi EMA, Misawa N, Schuch W, Bramley PM, Fraser PD. Metabolic engineering of ketocarotenoid formation in higher plants. The Plant Journal. 2004;39:477-486
142.Zhu C, Zhu C, Naqvi S, Capell T, Christou P. Metabolic engineering of ketocarotenoid biosynthesis in higher plants. Archives of Biochemistry and Biophysics. 2009;483:182-190
143.Ha S-H, Kim JK, Jeong YS, You M-K, Lim S-H, Kim J-K. Stepwise pathway engineering to the biosynthesis of zeaxanthin, astaxanthin and capsanthin in rice endosperm. Metabolic Engineering. 2019;52:178-189
144.Jeong YS, Ku H-K, Jung Y-J, Kim JK, Lee KB, Kim J-K, et al. 2A-linked bi-, tri-, and quad-cistrons for the stepwise biosynthesis of β-carotene, zeaxanthin, and ketocarotenoids in rice endosperm. Metabolic Engineering Communications. 2021;12:e00166
145.Unkles SE, Valiante V, Mattern DJ, Brakhage AA. Synthetic biology tools for bioprospecting of natural products in eukaryotes. Chemistry & Biology. 2014;21:502-508
146.Samalova M, Fricker M, Moore I. Ratio-metric fluorescence-imaging assays of plant membrane traffic using polyproteins. Traffic. 2006;7:1701-1723
147.Ding Q , Lee YK, Schaefer EA, Peters DT, Veres A, Kim K, et al. A TALEN genome-editing system for generating human stem cell-based disease models. Cell Stem Cell. 2013;12(2):238-251. DOI: 10.1016/j.stem.2012.11.011
148.Duda K, Lonowski LA, Kofoed- Nielsen M, Ibarra A, Delay CM, Kang Q , et al. High-efficiency genome editing via 2A-coupled co-expression of fluorescent proteins and zinc finger nucleases or CRISPR/Cas9 nickase pairs. Nucleic Acids Research. 2014;42(10):e84. DOI: 10.1093/nar/gku251
149.Kim H, Kim M-S, Wee G. Magnetic separation and antibiotics selection enable enrichment of cells with ZFN/TALEN-induced mutations. PLoSONE. 2013;8(2):e56476. DOI: 10.1371/journa.pne.0056476
150.Certo MT, Ryu BY, Annis JE, Garibov M, Jarjour J, Rawlings DJ, et al. Tracking genome engineering outcome at individual DNA breakpoints. Nature Methods. 2011;8(8):671-676
151.Moore R, Spinhirne A, Lai MJ, Preisser S, Li Y, Kang T, et al. CRISPR-based self-cleaving mechanism for controllable gene delivery in human cells. Nucleic Acids Research. 2015;43(2):1297-1303. DOI: 10.1093/nar/gku1326
152.Hofmann A, Falk J, Prangemeier T, Happel D, Köber A, Christmann A, et al. A tightly regulated and adjustable CRISPR-dCas9 based AND gate in yeast. Nucleic Acids Research. 2019;47(1):509-520. DOI: 10.1093/nar/gky1191
153.Nakamura M, Srinivasan P, Chavez M, Carter MA, Dominguez AA, La Russa M, et al. Anti-CRISPR-mediated control of gene editing and synthetic circuits in eukaryotic cells. Nature Communications. 2019;10(1):194. DOI: 10.1038/s41467-018-08158x
154.Arevalo-Villalobos JI, Govea- Alonso DO, Baňuelos-Hernández B, González-Ortega O, Zarazúa S, Rosales-Mendoza S. Inducible expression of antigens in plants: A study focused on peptides related to multiple sclerosis immunotherapy. Journal of Biotechnology. 2020;318:51-56
155.Lin Y, Hu CC, Liao JT, Lee YL, Huang YW, Lin NS, et al. An effective way of producing fully assembled antibody in transgenic tobacco plants by linking heavy and light chains via a self-cleaving 2A peptide. Frontiers in Plant Science. 2018;9:1-11
156.Walker PJ, Klement E. Epidemiology and control of bovine ephemeral fever. Veterinary Research. 2015;46:124. DOI: 10.1186/s13567-015-0262-4
157.Walker PJ, Byrne KA, Cybinski DH, Doolan DL, Wang Y. Proteins of bovine ephemeral fever virus. Journal of General Virology. 1991;72:67-74
158.Almasi S, Bakhshesh M. Antigenic variation of bovine ephemeral fever viruses isolated in Iran, 2012-2013. Virus Genes. 2019;55:654-659. DOI: 10.1007/s11262-019-01688-6
159.Richter V, Lebl K, Baumgartner W, Obritzhauser W, Käsbohrer A, Pinior B. A systematic worldwide review of the direct monetary losses in cattle due to bovine viral diarrhoea virus infection. The Veterinary Journal. 2017;220:80-87. DOI: 10.1016/j.tvjl.2017.01.005
160.Donis RO, Corapi W, Dubovi EJ. Neutralizing monoclonal antibodies to bovine viral diarrhoea virus bind to the 56K and 58K glycoprotein. Journal of General Virology. 1988;69:77-86
161.Park K, Jeong J, Chung BH. Live imaging of cellular dynamics using a multi-imaging vector in single cells. Chemical Communications. 2014;50:10734
162.Jeyarajan S, Zhang IX, Arvan P, Lentz SI, Satin LS. Simultaneous measurement of changes in mitochondrial and endoplasmic reticulum free calcium in pancreatic beta cells. Biosensors. 2023;13:382. DOI: 10.3390/bios13030382
163.Schwirz J, Yan Y, Franta Z, Schetelig MF. Bicistronic expression and differential localization of proteins in insect cells and Drosophila suzukii using picornaviral 2A peptides. Insect Biochemistry and Molecular Biology. 2020;119:103324
164.Summers DF, Maizel JV. Evidence for large precursor proteins in poliovirus synthesis. Proceedings of the National Academy of Sciences USA. 1968;59:966-971. DOI: 10.1073/pnas.59.3.966
165.Grant PT, Reid KB. Biosynthesis of an insulin precursor by cod islet tissue and the occurrence of intermediates in the conversion to insulin. Biochemical Journal. 1968;109(3):32P. DOI: 10.1042/bj1090032pa
166.Chance RE, Ellis RM, Bromer WW. Porcine proinsulin: Characterization and amino acid sequence. Science. 1968;161(3837):165-167. DOI: 10.1126/science.161.3837.165
167.Tung AK, Yip CC. The biosynthesis of insulin and “proinsulin” in foetal bovine pancreas. Diabetologia. 1968;4:68-70
Written By
Garry A. Luke and Martin D. Ryan
Submitted: 06 February 2024Reviewed: 08 February 2024Published: 10 April 2024