Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
The forested landscape of New England (USA) was dramatically altered by logging during the nineteenth and early twentieth centuries. Although the northern temperate forests of the region have largely regenerated, the streams and rivers remain impacted. The loss of terrestrial wood, organic material, and nutrient inputs during the forest regeneration period has affected habitat quality and biotic communities, most notably in small headwater streams. The same waterways are further impacted by now undersized stream crossings, mostly culverts associated with old infrastructure that alter hydrology and sediment transport; moreover, these culverts have created barriers to the movement of riverine organisms. We synthesize literature on headwater stream wood additions and culvert removal in North America and discuss observed patterns in organic matter, benthic macroinvertebrates, and Brook Trout (Salvelinus fontinalis) from before and after wood additions and stream-crossing enhancements in a previously logged watershed in New England. There were minimal changes to habitat and substrate two years after restoration efforts. However, streams with wood additions retained a higher density of rafted organic matter and had significantly higher benthic macroinvertebrate density. Additionally, two years after restoration, one year-old Brook Trout were significantly longer in restored streams than prior to restoration. Collectively, these results document a relatively rapid increase in organic matter retention, macroinvertebrates, and Brook Trout size, soon after restoration efforts.
Environmental Science and Policy, Center for the Environment, Plymouth State University, Plymouth, New Hampshire, USA
Amy M. Villamagna*
Environmental Science and Policy, Center for the Environment, Plymouth State University, Plymouth, New Hampshire, USA
Brigid C. O’Donnell
Biological Sciences, Center for the Environment, Plymouth State University, Plymouth, New Hampshire, USA
Tyson R. Morrill
Biological Sciences, Center for the Environment, Plymouth State University, Plymouth, New Hampshire, USA
Ben J. Nugent
New Hampshire Fish and Game Department, New Hampton, New Hampshire, USA
Jared B. Lamy
Biological Sciences, Center for the Environment, Plymouth State University, Plymouth, New Hampshire, USA
*Address all correspondence to: amvillamagna@plymouth.edu
1. Introduction
The forested landscape of the New England region of the United States was dramatically altered in the nineteenth and early twentieth century [1]. Old-growth forests were largely cleared, first for agriculture and then in response to the growing demand for wood products [2]. By 1839, New England and New York accounted for 41% of the nation’s timber harvest [3] and by the mid-1880s approximately 80% of New Hampshire forests had been cleared [4]. Since this time of dramatic disturbance, logging within riparian areas has decreased mature forested cover, increased stream temperatures and stream siltation, and reduced wood inputs that provide critical habitat [1, 5, 6, 7, 8, 9, 10, 11].
Allochthonous coarse particulate organic matter (CPOM), commonly in the form of autumn-shed leaves, supplies most of the energy to first order streams in eastern US deciduous forests [12, 13]. Retention of this energy in headwaters is often driven by large woody debris from riparian forests that trap CPOM and sediments, creating wood jams that operate as bioenergetic hotspots [8, 11, 14] and influence nutrient cycling, microbial populations, and macroinvertebrate densities locally and downstream [15, 16, 17, 18, 19, 20]. Retained CPOM can be transformed into dissolved organic matter (DOM) by leaching [21] or broken down into fine particulate organic matter (FPOM) by mechanical or biological processes. CPOM, DOM, and FPOM become the vectors for increased microbial and invertebrate biomass that influence food webs from the bottom up [22, 23, 24, 25]. The lack of such inputs has been shown to dramatically reduce invertebrate productivity [13, 26]. Wood jams also form pools, create protective overhead cover, and diversify habitat [27, 28, 29], providing fish in headwater streams with refuge from predation [30] and high flow events [31]. Several studies have suggested that wood additions benefit lotic fish populations [29, 32, 33, 34]. However, others suggest the impacts are lessened in headwater streams where boulder substrates and high gradients create habitat complexity [32, 35]. Headwater streams represent 79% of river length in the conterminous United States, and so clarifying the impacts of restoration is a worthwhile goal [36].
Although the northern temperate forests of the region have largely regenerated with the decrease in logging, the suppression of natural fires, and land conservation (e.g., the White Mountains National Forest), the waterways that crisscross the landscape remain impacted. Mature forests contribute less wood, organic material, and nutrients to streams and rivers than their old-growth counterparts [8, 37]. The absence of these terrestrial inputs has a documented effect on habitat quality, species assemblages, and species densities [1, 38, 39]. The same systems remain impacted by dams and stream crossings (mostly culverts) associated with the timber industry. The artificial water conduits of the intensive logging period were constructed for short-term economic gain, not long-term resilience. Although some stream crossings were replaced during the development of the modern automobile corridor, those that remain within the reforested areas are undersized for current hydrologic flows or poorly fit, resulting in altered hydrologic and sediment flows and substantial impediments to the movement of aquatic species [40, 41]. Culverts can alter sediment transport, organic matter retention, and hydrology [41]; some evidence shows culverts may negatively impact benthic macroinvertebrate communities [18, 42, 43, 44]. Impassable road-stream crossings also limit the ability of fish to thermoregulate, forage efficiently, and spawn effectively [45, 46, 47, 48].
Headwater streams maintain robust populations of Brook Trout (Salvelinus fontinalis) throughout their historic range in the eastern USA [49]. However, this species is highly vulnerable to habitat fragmentation and degradation within headwater streams [46, 50]. Brook Trout require high-quality and heterogeneous habitat to thermoregulate, avoid competition, access spawning habitat, and to find refuge during seasonal droughts and extreme flow events [47, 51, 52]. Therefore, habitat fragmentation and degradation attributed to impassable stream crossings and reduced woody inputs pose significant challenges to headwater Brook Trout populations and increase their risk of extirpation [47, 53, 54, 55, 56]. These threats are exacerbated by warming waters and the loss of suitable habitat [57, 58, 59, 60, 61].
Stream restoration efforts in historically logged watersheds often focus on wood additions [11], but documented responses across trophic levels are limited for headwater systems [32]. To date, few studies have documented a positive fish response to culvert removal or wood additions in headwater streams, and fewer still have tracked multitrophic responses to such restoration efforts [32, 62, 63, 64]. Researchers who study stream restoration efforts have emphasized a need for more monitoring before and after intervention, over longer time periods, and with detailed measurements of abiotic and biotic factors [29, 32, 33, 65]. While long and controlled experimental designs (e.g., Before-After-Control-Impact, or BACI) are generally preferred, the conditions for such studies are inherently rare given the limited and opportunistic nature of restoration (i.e., financial investment, time, and access to equipment).
Despite the prevalence of movement barriers and the lack of wood recruitment impacting streams, few studies have been published regarding Brook Trout populations in New Hampshire [49, 66, 67, 68]. Herein, we describe an adaptive sampling approach focused on evaluating early fine-scale abiotic and multitrophic responses to culvert removal and upstream wood additions in headwater streams within a watershed in central New Hampshire. We discuss patterns of wood accumulation, organic matter retention, benthic macroinvertebrates, and Brook Trout before and after restoration treatments which suggest rapid local and downstream responses across trophic levels. For a discussion of the impacts of habitat fragmentation and recent reconnection on the population genetic structure of wild Brook Trout in the watershed, we direct the reader to Lamy et al. [69]. In closing, we discuss the value of our work in the context of environmental management and conservation.
The Beebe River watershed (USGS Hydrologic Unit Code: 01070010403; area = 20,187 acres/81.8 sq. km) is located in central New Hampshire (USA). This river is a tributary to the Pemigewasset River that flows south through New Hampshire into Massachusetts where it continues to the Atlantic Ocean. Like most forested areas in central and northern New Hampshire, the Beebe River watershed has a long history (>150 years) of logging, including the more selective timber harvests of the current era to accommodate power lines that run parallel to a section of the river’s mainstem. Indications of logging within the watershed were first documented on a map produced in 1860 when a completed road paralleled the Beebe River toward Sandwich and a sawmill was constructed on the upper river [70]. The ~42 km Beebe River Railroad was constructed between 1917 and 1921 [71]. The establishment of this access route initiated extensive logging in the region. Toward the end of WWI, virgin Red Spruce (Picea rubens) from the Beebe River area contributed one quarter of the timber used in construction of US military aircraft [72]. All the spruce was reportedly removed by 1920, but the railroad remained as one of three active routes through the White Mountains [71]. In 1923, a fire, likely attributed to forest slash ignited railside, burned more than 14 sq. km (3500 acres) of the Beebe headwater forest [72]. Much of the regional softwood had been harvested and other large expanses burned, while patches of hardwood forests remained. Logging continued throughout the region for 40–50 years, depleting the area of usable timber. As a result, the current forest stand age largely ranges between 80 and 100 years old [4].
Between 1977 and the early 2000s, ownership changed several times and the property was broken into smaller parcels [73], including 1960 ha purchased by the US Forest Service to expand the White Mountain National Forest. In 2014, The Conservation Fund (TCF), a US-based land conservation organization, purchased 22 sq. km (5441 acres), containing 27 percent of the Beebe River watershed, and led stream restoration and land protection efforts (in cooperation with Redstart Forestry in 2017, the town of Sandwich, NH in 2017, and the town of Campton, NH in 2018). Such efforts included the removal and replacement by steel and plank bridges of five undersized culverts and bridges that impeded stream crossings. This restoration was funded by the TCF’s Working Forest Fund as well as the U.S. Department of Agriculture’s Regional Conservation Partnership Program (RCPP). The RCPP was an initiative of the 2014 Farm Bill and was funded from the U.S. Forest Service’s Forest Legacy Program through the Land and Water Conservation Fund; this enabled TCF to permanently protect the land with working forest conservation easements that maintain public recreational access in perpetuity. In total, TCF reconnected nearly six miles of wild Brook Trout spawning and rearing habitat before selling the easements in 2023 (The Conservation Fund 2023).
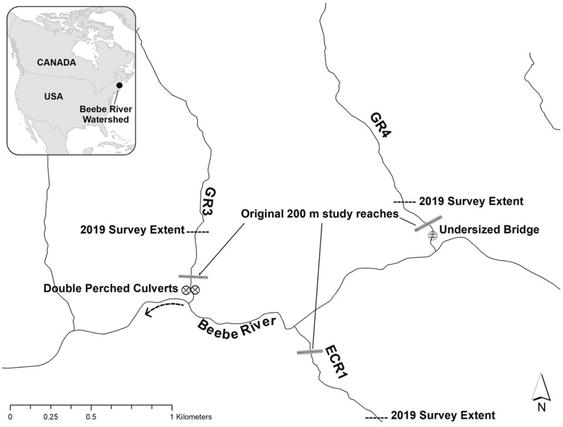
The study described herein arose as an opportunity to document the legacy effects of logging on the watershed and monitor the effects of restoration efforts. Data collection was conducted through a partnership between Plymouth State University, the NH Fish and Game Department, and the Pemigewasset Chapter of Trout Unlimited. Three tributaries to the Beebe River, denoted as GR3, GR4, and ECR1 (Figure 1) were the focus of this work. These streams were selected for this study based on their similarity in channel width (2–3 m), gradient (4–10% within each), substrate (boulder dominant; Table 1), canopy cover, proximity to one another (Figure 1), but also differences relating to the extent of habitat fragmentation, degradation, and restoration efforts. Mean daily maximum water temperatures in July ranged from 18.2°C to 20.6°C in GR3 (wood and culvert), 18.0°C to 21.0°C in GR4 (wood), and 17.9°C to 19.5°C in ECR1 (untreated) between 2016 and 2019. Mean daily average water temperatures in July ranged from 15.6°C to 18.0°C in GR3, 16.4°C to 18.2°C in GR4, and 16.3°C to 17.1°C in ECR1 between 2016 and 2019.
Figure 1.
Map of three study streams within the Beebe River watershed in central New Hampshire (USA). GR3 had two perched culverts replaced by a bridge and wood additions. GR4 had a passable but undersized bridge replaced by new bridge with wood additions. ECR1 was not affected by restoration efforts.
ECR1
GR3
GR4
Year
‘17
‘18
‘19
‘17
‘18
‘19
‘17
‘18
‘19
B & C (%)
62
45
65
55
56
70
76
57
88
L & S (%)
36
38
28
44
38
25
24
43
11
F (%)
2
18
6
1
6
6
0
1
2
P (%)
36
33
40
45
42
34
31
28
44
G (%)
10
4
5
21
29
7
22
28
7
R (%)
13
31
14
12
11
30
23
11
26
C (%)
41
32
41
22
18
30
24
33
22
P:R ratio
2.8
1.1
2.9
3.8
3.8
1.1
1.3
2.5
1.7
Table 1.
Substrate and habitat attributes over years (2017–2019) and across the three 200 m study reaches.
B & C: Boulder and Cobble; L & S: Large and Small Gravel; F: Fines; P: Pool; R: Riffle; C: Cascade; and P:R: Pool to Riffle.
A historic logging road parallels the Beebe River and intersects the south-facing study streams (GR3 and GR4) and a similar road intersects the north-facing study stream (ECR1) at a point 500 m upstream from the confluence with the mainstem. Up until 2017, the stream crossing at GR3 contained two perched culverts that were deemed impassable to Brook Trout, while GR4 contained a fully passable bridge that was a pinch point during high stream flow events. In ECR1, a perched culvert remains at the road-stream crossing 300 m above our original 200 m study reaches (Figure 1); however, ECR1 is considered the reference or untreated stream because the culvert was not removed and wood was not added over the study period. In August 2017, double perched culverts in GR3 and an undersized bridge over GR4 were replaced with steel stringer bridges that are fully passable by Brook Trout. After the 2017 sampling season, one to two trees per 100 m of stream were added upstream of our original 200 m study reaches in GR3 and GR4. GR3 and GR4 in a perpendicular orientation, with one to two locations having crisscrossed logs and several branches to accumulate leaves and create debris dams, according to federal protocols [74]. In August 2018, four trees were added to GR3 and one tree to GR4 within 100 m of the stream-river confluence. The 2018 wood additions occurred almost entirely downstream of benthic macroinvertebrate sampling. Therefore, we consider the August 2018 wood additions to have minimal influence on our results and attribute the observed responses to upstream wood additions. In summary, GR3 was treated with wood additions and culvert removal, GR4 was treated with only wood additions, and ECR1 was untreated.
In the summers of 2017–2019, we conducted habitat and benthic macroinvertebrate surveys once a season beginning at the stream-river confluence and continuing upstream to 200 m, including side channels (hereafter referred to as the “original 200 m reach”). We conducted Brook Trout surveys throughout the same reaches in July of 2016–2019. Surface areas within the 200 m reach varied over time with changes in water level and side channels (GR3: 371–587 m2, GR4: 444–687 m2, ECR1: 473–532 m2). All surveys were extended in 2019 to span the full upstream extent of Brook Trout (GR3 culvert and wood = 650 m, GR4 wood = 450 m, and ECR1 untreated = 900 m). Natural vertical barriers (≥1 m hydraulic jump) delimited the upstream extent of Brook Trout in the study streams. This expansion of our survey areas was intended to capture the full scope of available benthic macroinvertebrates and habitat for Brook Trout following changes in upstream habitat.
3.1 Habitat surveys
We surveyed stream habitat following a modified version of NH Fish and Game’s Rapid Habitat Assessment (New Hampshire Fish and Game, personal communication) and measured wood following TWF Monitoring Program: Large Woody Debris Survey [75]. Habitat and wood surveys were conducted annually in late July to mid-August, during base flow conditions (determined by USGS station #01016500). Habitat assessments included characterization of habitat unit type, habitat unit dimensions (length, width, average depth), channel characteristics (wetted and bankfull width), dominant substrate type, stream gradient, hydraulic jump height, stream bank stability, and riparian forest type. Surveys of large wood included characterization of tree species, wood origin, wood decay state, and wood dimensions. Large wood included any piece of wood that was ≥1 m length and ≥10 cm diameter [76] and located at least partially within the bankfull prism, as defined by water levels assumed during bankfull flow [77]. The locations and dimensions of smaller organic material moved by flowing water (i.e., rafted organic matter [ROM]) within the bankfull prism were recorded and associated with measured wood [78, 79]. We summarized relative changes in large wood density, dry and wet ROM, habitat availability, and substrate type in focal streams over time (2017–2019). Water temperature in the streams was continuously monitored every 30 minutes with HOBO sensors (U22-0001; Onset Computer Corp., Bourne Massachusetts) distributed longitudinally to measure thermal variability within and across streams [78, 79].
3.2 Benthic macroinvertebrate surveys
We collected benthic macroinvertebrates in June of 2017, 2018, and 2019 using Surber samplers and D-frame nets (sample area = 0.09 m2, net mesh size = 500 μm) from randomly selected riffles and pools in proportion to their occurrence (i.e., count) throughout the original 200 m study reaches. Six to seventeen D-frame samples were collected from the reaches in 2017 and 2018. To increase the efficiency of sampling and identification in 2019, we pooled seven samples from each stream in the field (Table 2). Macroinvertebrates were stored in 70% ethanol and transported to the lab for family-level identification.
Density and biomass of benthic macroinvertebrates in original 200 m reaches by stream, year, and order: GR3 (wood and culvert), GR4 (wood), and ECR1 (untreated).
Seven samples from the original 200 m were pooled in the field in 2019.
To test for significant changes in total and order-level benthic macroinvertebrate densities within a given stream between 2017 and 2018, we used a Pairwise Wilcoxon test with a Benjamini-Hochberg (BH) p-value adjustment. Considering samples were grouped by sampling zone in the field before laboratory identification in 2019, we conducted a one-sample T-test with a BH p-value adjustment to compare 2019 to 2018 and 2017. We adopted a non-parametric approach because the data did not conform to parametric assumptions and non-parametric methods are considered suitable, if not better, for environmental data that is not normally distributed [80, 81]. Dry weight biomass was estimated for each year using random sub-samples at the family-level and summarized at the order-level (Table 2). Family-level pooled samples were dried in the winter of 2019 in an oven (Precision Econotherm Lab Oven, Thermo Scientific) with forced ventilation for 16 h at 105°C, cooled for 24 h, and weighed with an electronic balance [82].
3.3 Brook Trout surveys
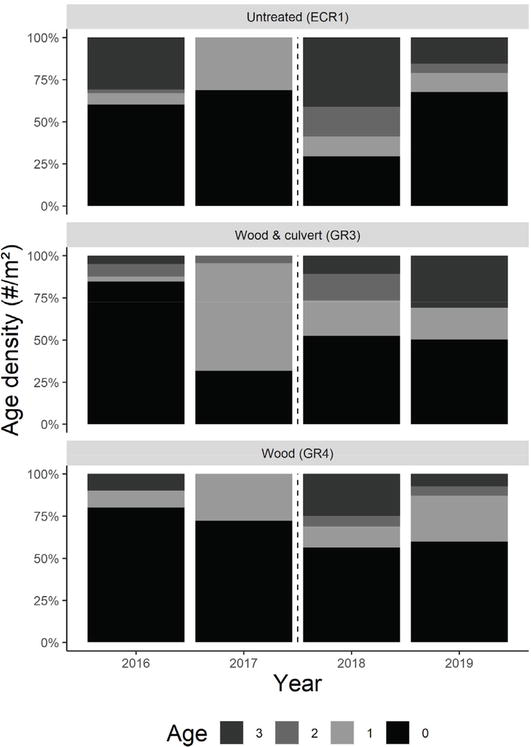
We surveyed fish using two-pass depletion methods [83] with Smith-Root model LR-24 DC (Vancouver, WA, U.S.A.) backpack electrofishing units at 600–800 volts in July of 2016–2019 to ensure age 0 (young of year) Brook Trout had reached catchable size. In addition to Brook Trout, we occasionally captured Rhinichthys atratulus (Blacknose Dace) and Rhinichthys cataractae (Longnose Dace) near the stream-river confluence. Non-target species were returned to the stream without measurement. Estimated Brook Trout capture probabilities were calculated from multi-pass removal sampling as described in Carle and Strub [84] and averaged between 84 and 100% across sampling events, with an overall average estimated capture probability of 95%. High capture probability justified our use of two-pass depletion methods. Upon capture, the location of each Brook Trout was recorded to ensure fish were returned to the same subsection after data collection. All fish were measured for weight (g) and total length (mm) and each untagged Brook Trout that was ≥ 60 mm total length and ≥ 2g was sampled for scales above the lateral line and posterior to the dorsal fin [85]. Scales were brought back to the lab for age estimation, which enabled us to assess and compare length at specific ages over time. Brook Trout scale samples were dry mounted between two glass microscope slides and viewed under 10X magnification to identify periods of winter growth and summer growth [85]. Age was determined with knowledge of the sampling month, but without knowledge of fish length or location. Thirty percent of age assignments were reviewed by a third party. For the few discrepancies noted between assessors, scales were reassessed. The age structure of Brook Trout captured in July from the original 200 m reach of each tributary was compared over time using a two-sample Kolmogorov-Smirnov test (Figure 2).
Figure 2.
Proportional abundance of Brook Trout by age, based on July captures in the original 200 m reaches in GR3 (wood and culvert), GR4 (wood), and ECR1 (untreated) from 2016 to 2019. Hashed lines mark when habitat restoration efforts occurred.
Biomass and density values were calculated by summing the individual weights and number of captured Brook Trout in each tributary and standardized by area. We used a Pairwise Wilcoxon Rank Sum test with a BH p-value adjustment to compare Fulton’s Condition Factor [86], total length of 1 year old (hereafter, age 1) Brook Trout, and density and biomass of Brook Trout within streams over time. We constrained our total length analyses to age 1 Brook Trout because this class was the most abundant in our samples (Figure 2), and younger individuals are more responsive to prey availability [87].
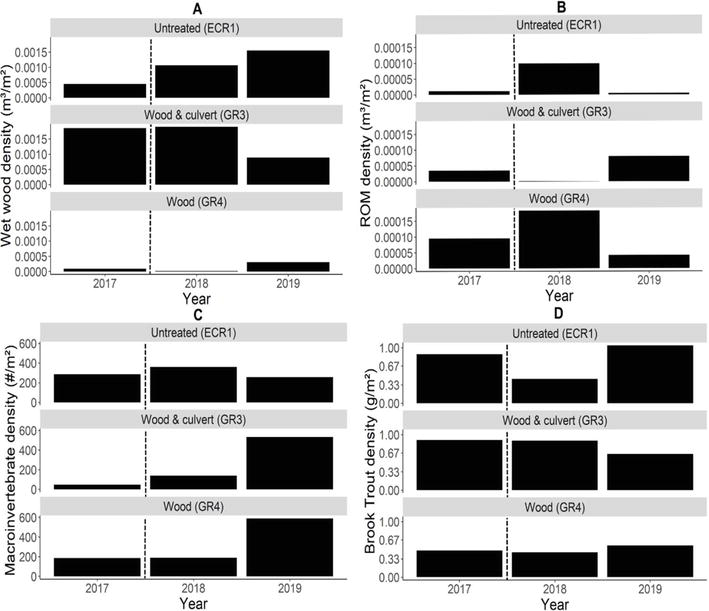
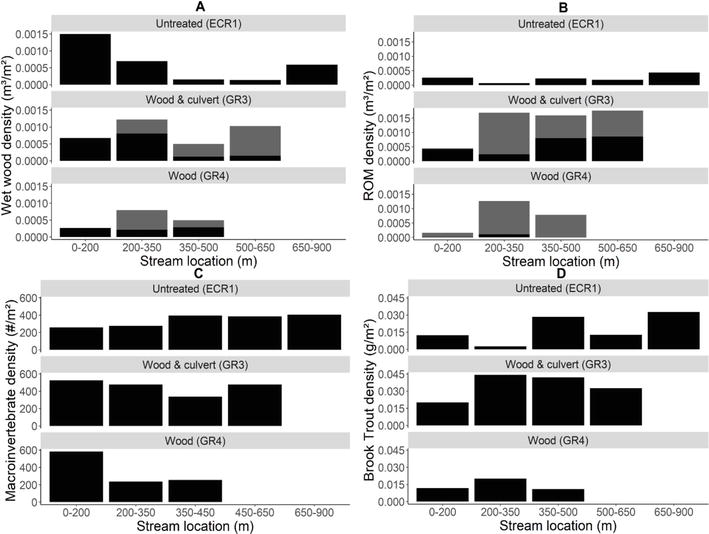
There were no notable changes in substrate composition or habitat types in the original 200 m reaches after perched culvert removal (GR3) or wood additions (GR3 and GR4), except immediate downstream of the culvert in GR3 where a large scour pool was replaced with a long riffle. Wet wood density (i.e., wood that is fully immersed in the stream) and ROM density in the original 200 m reaches varied over time, but there was no clear relationship with stream modifications in either case (Figure 3A and B). There were no consistent changes in habitat or substrate types 2 years after wood additions in the original 200 m reaches (Table 1), likely because less than 6% of total wood additions occurred there. Roughly half of the 2019 wet wood density across the full survey extent in GR3 and GR4 was associated with wood additions; however, ~94% of this added wood was situated upstream of the original 200 m reach (Figure 4A), and ROM was predominantly retained upstream of the original 200 m reach (Figure 4B). Moreover, GR3 held roughly four times and GR4 two and a half times greater ROM density than ECR1 in 2019. Of that ROM, 53% in GR3 and 95% in GR4 was associated with added wood (Figure 4B).
Figure 3.
Density of (A) wet wood, (B) rafted organic matter (ROM), (C) macroinvertebrates, and (D) Brook Trout in GR3 (wood and culvert), GR4 (wood), and ECR1 (untreated) from 2017 to 2019 in the original 200 m reaches. Hashed lines mark when habitat restoration efforts occurred.
Figure 4.
Density of (A) wet wood, (B) rafted organic matter (ROM), (C) macroinvertebrates, and (D) Brook Trout in 2019, following culvert removal and wood additions, in the extended survey reaches: 650 m in GR3 (wood and culvert), 450 m in GR4 (wood), and 900 m in ECR1 (untreated). In A and B, black indicates natural/existing wood and gray indicates added wood.
4.2 Benthic macroinvertebrates
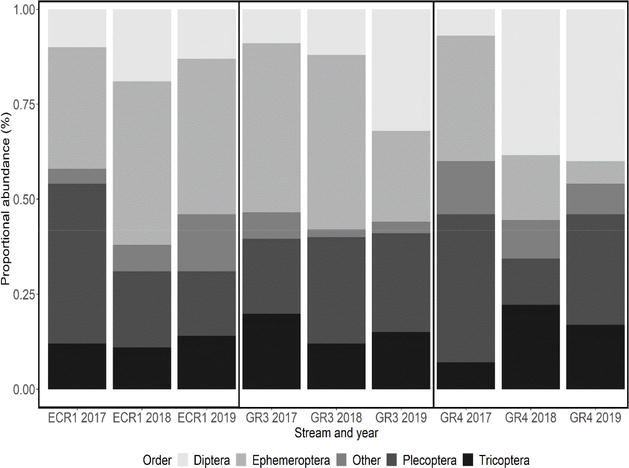
Macroinvertebrate density roughly tripled 1 year after perched culvert removal and wood additions in GR3 (P = 0.015; Figure 3C), then tripled again, 2 years after restoration efforts (P < 0.001; Figure 3C). These increases were not proportional across orders (Figure 5) but were significant for all orders after 2 years (P = 0.001–0.020; Table 2). In GR4, macroinvertebrate density did not significantly differ 1 year after wood additions (P = 0.903; Figure 3C) but tripled after 2 years (P < 0.001; Figure 3C), with significant increases observed across all orders (P = 0.000–0.039; Table 2), except Ephemeroptera (P = 0.116; Table 2). Meanwhile, in our untreated stream (ECR1), we did not observe a significant change in macroinvertebrate density 1 year after restoration efforts (P = 0.358; Figure 3C), or 2 years after restoration efforts in any of the orders (P = 0.173–1; Table 2). We observed an increase in benthic macroinvertebrate biomass between 2017 and 2019 in all streams, but these values were partially influenced by the presence of a few large individuals (Plecoptera and Odonata) that often accounted for half of the biomass in a sample (Table 2). After excluding very large individuals, the trends in macroinvertebrate biomass were similar with those for density.
Figure 5.
Proportional abundance of benthic macroinvertebrate orders sampled from 2017 to 2019 in the original 200 m reaches of GR3 (wood and culvert), GR4 (wood), and ECR1 (untreated).
Prior to restoration efforts, benthic macroinvertebrate density in GR3 was one third that of GR4. Given the proximity of these streams, their shared underlying geology, and similar channel morphology and temperature regimes, this large difference in density prior to restoration was unexpected. One year and 2 years after restoration efforts, GR3 and GR4 macroinvertebrate densities were roughly equivalent (Figure 3C); therefore, we suspect that the impassable culvert in GR3 may have constrained benthic macroinvertebrate density prior to restoration efforts. This observation is consistent with studies that have found significant differences in abundance and taxonomic richness between benthic macroinvertebrate communities residing in habitats upstream and downstream of culverts [18, 42, 43]. In addition, these findings align with a study that observed significant increases in invertebrate richness in headwater streams following culvert removals [44]. However, to date, no studies have reported significant differences in benthic macroinvertebrate abundances before and after culvert removal [88].
We posit that increased ROM retention from wood additions in the upper reaches contributed to increased benthic macroinvertebrate density and possibly led to a bottom-up response following habitat restoration. Our observations of increased organic matter retention following wood additions in headwater streams corresponding with downstream increases in invertebrate secondary production are consistent with previous studies [25, 89, 90, 91].
The dramatic change in habitat and substrate within the area directly impacted by culvert removal and bridge installation may have influenced community composition and density. However, since we did not examine the post-restoration effects of culvert removal independent of wood additions in GR3, we cannot fully discount the role of other factors influencing benthic macroinvertebrates in these study streams. Future studies examining benthic macroinvertebrate functional feeding groups may provide more insight into culvert and wood impacts on stream abiotic processes [90].
4.3 Brook Trout
While we did not detect significant changes in Brook Trout population density (P = 0.243–0.438) (Figure 3D) or biomass (P = 0.280–0.480) in the original 200 m reaches in response to wood additions or culvert removal, previous studies have documented trout population increases one to 2 years after habitat modifications [62, 63, 92]. However, these same studies noted that rapid increases in trout density and biomass may be driven by immigration [62, 63, 92]. For example, Kratzer [63] noted an increase in Brook Trout abundance in treated stream sections 1 year after wood additions. This increase initially coincided with a decrease in biomass in nearby control sections, suggesting immigration from control sections into treated sections. However, within 2–4 years, biomass in control sections rebounded and then exceeded previous levels while treated sections maintained initial biomass increases, suggesting an overall increase in carrying capacity over time. A recent meta-analysis of salmonid responses to stream restoration efforts across 100 studies showed a significant increase in abundance and biomass but noted that monitoring time (<5 years) factors into variation in impacts following restoration [34].
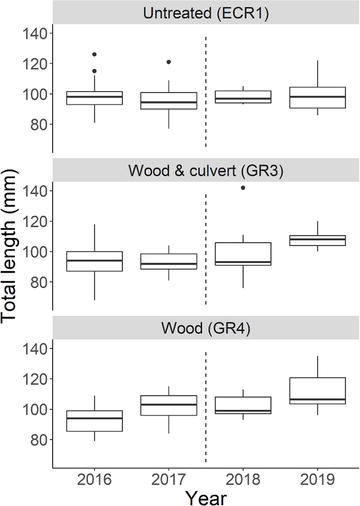
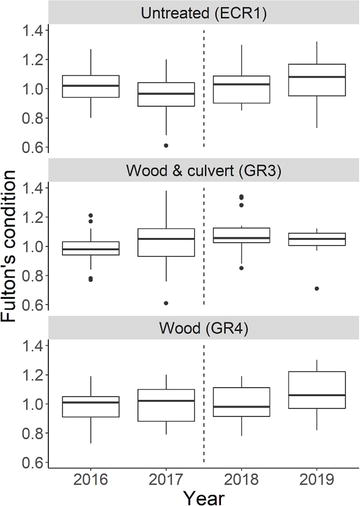
Despite no significant change in density or biomass, we detected a significant increase in age 1 Brook Trout length within the original 200 m reaches of treated streams 2 years after habitat modifications (GR3 culvert and wood: 2016–2019 P = 0.015, 2017–2019 P = 0.031; GR4 wood only: 2016–2019 P = 0.011; Figure 6). We found no significant difference in age 1 Brook Trout length in the untreated stream (ECR1 P = 0.433). Fulton’s condition did not differ within the original 200 m reaches over time (P = 0.17–0.99; Figure 7), with the exception of a significantly higher Fulton’s condition in GR3 in 2018 compared to 2016 (P = 0.038).
Figure 6.
Total length of age 1 Brook Trout captured in July in the original 200 m reaches in GR3 (wood and culvert), GR4 (wood), and ECR1 (untreated) from 2016 to 2019. The thick line indicates the median, boxes show the interquartile range. Hashed lines mark when habitat treatments occurred.
Figure 7.
Fulton’s condition for Brook Trout captured in July in the original 200 m reaches in GR3 (wood and culvert), GR4 (wood), and ECR1 (untreated) from 2016 to 2019. The thick line indicates the median, boxes show the interquartile range. Hashed lines mark when habitat treatments occurred.
Within our study, age 1 Brook Trout represented between 5 and 60% of the observed populations in July between 2016 and 2019 (Figure 2). We detected a significant length increase in age 1 Brook Trout 2 years after habitat modifications (Figure 6), without concomitant significant increases in population density and biomass within the original 200 m reaches (Figure 2D). Notably, increases in Brook Trout size have not been detected in previous wood addition studies [62]. We hypothesize that age 1 stream-dwelling Brook Trout were able to utilize increased benthic macroinvertebrate density for growth, which could explain observed increases in length. The hypothesis is supported by Keeley [93] who found that smaller Brook Trout rely more heavily on invertebrate prey than larger trout that become piscivorous. Additionally, previous studies have shown that headwater Brook Trout growth and population density are regulated by density dependence and macroinvertebrate production [94, 95]. Although we cannot factor out the possibility that some larger age 1 Brook Trout moved into GR3 and GR4 from the mainstem, GR4 has had no impediments to fish passage and our PIT tag mark-recapture data supports the contention that age 1 Brook Trout largely remain in their respective headwater streams during the sampling season [78, 79]. Ultimately, we anticipate that increased food availability and Brook Trout size may translate into higher fecundity, population density, and biomass over multiple generations.
Our results suggest that macroinvertebrates experienced a relatively rapid response to wood additions, but potential positive effects from culvert removal still need further exploration. Downstream of wood additions, we did not observe substantial short-term habitat changes or increases in Brook Trout density or biomass. However, we did observe significant increases in benthic macroinvertebrate density and age 1 Brook Trout length downstream of wood additions that retained the majority of ROM. Previous studies have shown that headwater Brook Trout populations are regulated by density dependence and invertebrate production [94, 95]. In light of this, we may see increases in Brook Trout biomass or density downstream of wood additions with continued years of monitoring.
Our results reflect short-term responses to restoration efforts in high gradient headwater streams of historically logged but still young forests where in-stream wood recruitment is limited. Broadly, researchers and managers should consider the ecological benefits of in-stream restoration efforts from a bottom-up perspective of habitat creation and energy capture toward increased secondary production of macroinvertebrates and fish [96]. While a long-term ecological monitoring plan that collects extensive data before and after manipulations across broad upstream and downstream perspectives is ideal [34, 90], it is rare given resource limitations and the opportunistic nature of many restoration endeavors. We were fortunate that our partners valued the data to inform decisions and invested in a pre-restoration assessment and post-restoration monitoring; however, given the resources available at the onset, we were limited in the extent of our monitoring. Early on our partners presented research and restoration plans widely, building community support for the endeavor. With an unanticipated infusion of funding in 2018, we were able to expand sampling efforts to capture the habitat characteristics associated with wood additions and track a suite of variables potentially driving our observed downstream responses. Rather than ignore the upstream extent of the focal streams to preserve the original reach of 200 m, we opted to expand our scope to better inform management and conservation decisions. It was through the consistent communication of our seasonal data collection efforts, observations and data analyses with our partners that prompted them to establish additional environmental protection by writing in a mandatory 15 m “no harvest” riparian buffer into the conservation easements that would conserve the property in perpetuity. To our knowledge, this was the first explicit insertion of such strict stream protection policy into a conservation easement in New Hampshire where the law states that no more than 50% of the basal area of trees can be cut within 15 m of a stream (Basal Area Law, 2019; RSA 227-J:9). This study, examined as a whole, demonstrates the importance of engaging collaborative and diverse partners (management, conservation, and scientific) in collaborative restoration and monitoring decisions and effective communicating scientific observations with non-scientific audiences to encourage data-driven conservation action.
This study was a collaborative effort among faculty and students at Plymouth State University, the New Hampshire Fish and Game Department (B. Nugent, D. Timmins, J. Magee), the Pemigewasset chapter of Trout Unlimited (#726), and The Conservation Fund. We thank numerous research assistants for their contribution to field surveys and lab analysis: M. Collins, A. Dunlap, D. Tarr, J. Carboneau, C. Barnes, K. Crowley, S. Bevier, S. Welch, J. Corbet, N. Roberts, S. Bevier, N. Scola, and G. Nyhan. We also thank E. Laflamme for statistical advice and comments on earlier drafts. Sampling was permitted through New Hampshire Fish and Game Department Scientific Licenses F2016-101, F2017-097, F2018-94, and F2019-102 and Plymouth State University IACUC approval. Financial support included funding from the National Fish and Wildlife Foundation “Bring Back the Natives” program, New Hampshire Water Resources Research Center through the US Geological Survey National Institutes for Water Resources Grants Program (Section 104 g) [grant number G16AP00070], Tourism, Environment, and Sustainable Societies Cluster and the Lovett Fund at Plymouth State University, graduate assistantship funding from Plymouth State University, and undergraduate research funding from the Center for the Environment and the Student Research and Creativity Fund at Plymouth State University.
References
1.Nislow KH, Lowe WH. Influences of logging history and riparian forest characteristics on macroinvertebrates and Brook Trout (Salvelinus fontinalis) in headwater streams (New Hampshire, U.S.A.). Freshwater Biology. 2006;51:388-397
2.Foster DR. Land-use history (1730-1990) and vegetation dynamics in Central New England, USA. Journal of Ecology. 1992;80(4):753-771
3.Williams M. Clearing the United States forests: Pivotal years 1810-1860. Journal of Historical Geography. 1982;8(1):12-28
4.Langley-Turnbaugh SJ, Keirstead DR. Soil properties and land use history: A case study in New Hampshire. Northeastern Naturalist. 2005;12(4):391-402
5.Hayes DB, Taylor WW, Drake MT, Marod SM, Whelan GE. The value of headwaters to brook trout (Salvelinus fontinalis) in the Ford River, Michigan, USA. In: Haigh MJ, Krecke J, Rajwar GS, Kilmartin MP, editors. Headwaters: Water Resources and Soil Conservation. Brookfield, Vermont: Balkeman, A. A; 1998. pp. 175-185
6.Marschall EA, Crowder LB. Assessing population responses to multiple anthropogenic effects: A case study with brook trout. Ecological Applications. 1996;1:152-167
7.Beschta RL, Bilby ER, Brown WG, Holtby LB, Hofstra DT. Stream temperature and aquatic habitat: Fisheries and forestry interactions. In: Salo EO, Cundy TW, editors. Streamside Management: Forestry and Fishery Interactions. Seattle, Washington USA: University of Washington, Institute of Forest Resources. Contribution 57; 1987. pp. 191-232
8.Keeton WS, Kraft CE, Warren DR. Mature and old-growth riparian forests: Structure, dynamics, and effects on Adirondack stream habitats. Ecological Applications. 2007;17(3):852-868
9.Macdonald JS, MacIsaac EA, Herunter HE. The effect of variable-retention riparian buffer zones on water temperatures in small headwater streams in sub-boreal forest ecosystems of British Columbia. Canadian Journal of Forest Research. 2003;33(8):1371-1382
10.Wheeler AP, Angermeier PL, Rosenberger AE. Impacts of new highways and subsequent landscape urbanization on stream habitat and biota. Reviews in Fisheries Science. 2005;13(3):141-164
11.Warren RD, Kraft EC, Keeton SW, Nunery SJ, Likens GE. Dynamics of wood recruitment in streams of the northeastern U.S. Forest Ecology and Management. 2009;258(5):804-813
12.Wallace JB, Whiles RM, Eggert S, Cuffney TF, Lugthart GJ, Chung K. Long-term dynamics of coarse particulate organic matter in three Appalachian mountain streams. Journal of the North American Benthological Society. 1995;14(2):217-232
13.Wallace JB, Eggert S, Meyer J, Webster J. Stream invertebrate productivity linked to forest subsidies: 37 stream-years of reference and experimental data. Ecology. 2015;96(5):1213-1228
14.Bilby RE, Likens GE. Importance of organic debris dams in the structure and function of stream ecosystems. Ecology. 1980;1980(61):1107-1113
15.Battin TJ, Kaplan LA, Findlay S, Hopkinson CS, Martin E, Packman AI, et al. Biophysical controls on organic carbon fluxes in fluvial networks. Nature Geoscience. 2008;1:95-100
16.Beckman ND, Wohl E. Carbon storage in mountainous headwater streams: The role of old-growth forest and logjams. Water Resource Research. 2014;50:2376-2393
17.Elosegi A, Díez J, Pozo J. Contribution of dead wood to the carbon flux in forested streams. Earth Surface Processes and Landforms. 2007;32:1219-1228
18.Khan B, Colbo MH. The impact of physical disturbance on stream communities: Lessons from road culverts. Hydrobiologia. 2008;600:229-235
19.Smock LA, Metzler GM, Gladden JE. Role of debris dams in the structure and functioning of low-gradient headwater streams. Ecology. 1989;70:764-775
21.Petersen RC, Cummins KW. Leaf processing in a woodland stream. Freshwater Biology. 1974;4:343-368
22.Rau GH, Anderson NH. Use of 13 C/12 C to trace dissolved and particulate organic matter utilization by populations of an aquatic invertebrate. Oecologia. 1981;48:19-21
24.Peters GT, Benfield EF, Webster JR. Chemical composition and microbial activity of seston in a southern Appalachian headwater stream. Journal of the North American Benthological Society. 1989;1:74-84
25.Entrekin SA, Rosi EJ, Tank JL, Hoellein TJ, Lamberti GA. Quantitative food webs indicate modest increases in the transfer of allochthonous and autochthonous C to macroinvertebrates following a large wood addition to a temperate headwater stream. Frontiers in Ecology and Evolution. 2020;8:114
26.Wallace JB, Eggert SL, Meyer JL, Webster JR. Multiple trophic levels of a forest stream linked to terrestrial litter inputs. Science. 1997;277(5322):102-104
27.Champoux O, Biron MP, Roy GA. The long-term effectiveness of fish habitat restoration practices: Lawrence Creek, Wisconsin. Annals of the Association of American Geographers. 2003;93(1):42-54
28.Opperman JJ, Merenlender AM, Lewis D. Maintaining wood in streams: A vital action for fish conservation. In: Division of Agriculture and Natural Resources. Davis, California, USA: University of California; 2006. DOI: 10.3733/ucanr.8157
29.Whiteway SL, Biron PM, Zimmermann A, Venter O, Grant JWA. Do in-stream restoration structures enhance salmonid abundance? A meta-analysis. Canadian Journal of Fisheries and Aquatic Sciences. 2010;67(5):831-841
30.Harvey BC, Stewart AJ. Fish size and habitat depth relationships in headwater streams. Oecologia. 1991;87:336-342
31.Lewis SL. Physical factors influencing fish populations in pools of a trout stream. Transactions of the American Fisheries Society. 1969;98:14-19
32.Nagayama S, Nakamura F. Fish habitat rehabilitation using wood in the world. Landscape and Ecological Engineering. 2010;6:289-305
33.Roni P, Beechie TJ, Pess GR, Hanson KM. Wood placement in river restoration: Fact, fiction, and future direction. Canadian Journal of Fisheries and Aquatic Sciences. 2015;72(3):466-478
34.Foote KJ, Biron PM, Grant JW. Impact of in-stream restoration structures on salmonid abundance and biomass: An updated meta-analysis. Canadian Journal of Fisheries and Aquatic Sciences. 2020;9:1574-1591
35.Warren RD, Kraft EC. Brook trout (Salvelinus fontinalis) response to wood removal from high-gradient streams of the Adirondack Mountains (N.Y., U.S.A.). Canadian Journal of Fisheries and Aquatic Sciences. 2003;60(4):379-389
36.Colvin SA, Sullivan SMP, Shirey PD, Colvin RW, Winemiller KO, Hughes RM, et al. Headwater streams and wetlands are critical for sustaining fish, fisheries, and ecosystem services. Fisheries. 2019;44(2):73-91
37.Bilby RE, Ward JW. Characteristics and function of large woody debris in streams draining old-growth, clear-cut, and second-growth forests in southwestern Washington. Canadian Journal of Fisheries and Aquatic Sciences. 1991;48:2499-2508
38.Benke AC, Wallace JB. Influence of wood on invertebrate communities in streams and rivers. In: Gregory SV, Boyer KL, Gurnell AM, editors. The Ecology and Management of Wood in World Rivers. Symposium 37. Bethesda, MD: American Fisheries Society; 2003. pp. 149-177. DOI: 10.47886/9781888569568
39.Dolloff CA, Warren ML Jr. Fish relationships with large wood in small streams. In: Gregory SV, Boyer KL, Gurnell AM, editors. The Ecology and Management of Wood in World Rivers. Symposium 37. Bethesda, MD: American Fisheries Society; 2003. pp. 179-193. DOI: 10.47886/9781888569568
40.Poplar-Jeffers IO, Petty JT, Anderson JT, Kite SJ, Strager MP, Fortney RH. Culvert replacement and stream habitat restoration: Implications from brook trout Management in an Appalachian watershed, U.S.a. Restoration Ecology. 2009;17(3):404-413
41.Wellman JC, Daniel LC, Bradford SC. Long-term impacts of bridge and culvert construction or replacement on fish communities and sediment characteristics of streams. Journal of Freshwater Ecology. 2000;15(3):317-328
42.Peterson T. The effect of road culverts on the benthic macroinvertebrate community in wadeable lotic ecosystems. PSU McNair Scholars Online Journal. 2010;4:52-74
43.Gál B, Weiperth A, Farkas J, Schmera D. The effects of road crossings on stream macro-invertebrate diversity. Biodiversity and Conservation. 2020;29:729-745
44.Omoniyi GE, Piscart C, Pellan L, Bergerot B. Responses of macroinvertebrate communities to hydromorphological restoration of headwater streams in Brittany. Water. 2022;4:553
45.Kentaro M, Shoichiro Y. Effects of habitat fragmentation by damming on the persistence of stream-dwelling charr populations. Conservation Biology. 2002;16(5):1318-1323
46.Letcher BH, Nislow KH, Coombs JA, O’Donnell MJ, Dubreuil TL. Population response to habitat fragmentation in a stream-dwelling brook trout population. PLoS One. 2007;11:1-11
47.Petty JT, Hansbarger JL, Huntsman BM, Mazik PM. Brook trout movement in response to temperature, flow, and thermal refugia within a complex Appalachian riverscape. Transactions of the American Fisheries Society. 2012;141(4):1060-1073
48.Warren ML, Pardew MG. Road crossings as barriers to small-stream fish movement. Transactions of the American Fisheries Society. 1998;127:637-644
49.Hudy M, Thieling TM, Gillespie N, Smith EP. Distribution, status, and land use characteristics of subwatersheds within the native range of brook trout in the eastern United States. North American Journal of Fisheries Management. 2008;28(4):1069-1085
50.Kratzer JF, Warren DR. Factors limiting brook trout biomass in northeastern Vermont streams. North American Journal of Fisheries Management. 2013;33(1):130-139
51.Hoffman R, Dunham J. Fish-Movement Ecology in High-Gradient Headwater Streams: Its Relevance to Fish Passage Restoration through Stream Culvert Barriers. Virginia: US Geological Survey; 2007. OFR 2007-1140
52.Magoulick DD, Wilzbach MA. Are native brook charr and introduced rainbow trout differentially adapted to upstream and downstream reaches? Ecology of Freshwater Fish. 1998;7:167-175
53.Thompson DP, Rahel JF. Evaluation of artificial barriers in small Rocky Mountain streams for preventing the upstream movement of brook trout. North American Journal of Fisheries Management. 1998;18(1):206-210
54.Nathan LR, Smith AA, Welsh AB, Vokoun JC. Are culvert assessment scores an indicator of brook trout Salvelinus fontinalis population fragmentation? Ecological Indicators. 2018;84:208-217
55.Torterotot JB, Perrier C, Bergeron NE, Bernatchez L. Influence of forest road culverts and waterfalls on the fine-scale distribution of brook trout genetic diversity in a boreal watershed. Transactions of the American Fisheries Society. 2014;6:1577-1591
56.Wood DM, Welsh AB, Todd PJ. Genetic assignment of brook trout reveals rapid success of culvert restoration in headwater streams. North American Journal of Fisheries Management. 2018;5:991-1003
57.Battin J, Wiley MW, Ruckelshaus MH, Palmer RN, Korb E, Bartz KK, et al. Projected impacts of climate change on salmon habitat restoration. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:6720-6725
58.Durance I, Ormerod SJ. Climate change effects on upland stream macroinvertebrates over a 25-year period. Global Change Biology. 2007;13:942-957
59.Flebbe PA, Roghair LD, Bruggink JL. Spatial modeling to project southern Appalachian trout distribution in a warmer climate. Transactions of the American Fisheries Society. 2006;135(5):1371-1382
60.Hakala JP, Hartman KJ. Drought effect on stream morphology and brook trout (Salvelinus fontinalis) populations in forested headwater streams. Hydrobiologia. 2004;515:203-213
61.Ouellet V, Gibson EE, Daniels MD, Watson NA. Riparian and geomorphic controls on thermal habitat dynamics of pools in a temperate headwater stream. Ecohydrology. 2017;10(8):e1891
62.Gowan C, Fausch KD. Long-term demographic responses of trout populations to habitat manipulation in six Colorado streams. Ecological Applications. 1996;6:931-946
63.Kratzer JF. Response of brook trout biomass to strategic wood additions in the east branch Nulhegan River watershed, Vermont. North American Journal of Fisheries Management. 2018;38:1415-1422
64.Sweka JA, Hartman KJ. Effects of large woody debris addition on stream habitat and brook trout populations in Appalachian streams. Hydrobiologia. 2006;559:363-378
65.Rubin Z, Kondolf GM, Rios-Touma B. Evaluating stream restoration projects: What do we learn from monitoring? Water. 2017;9(3):174
66.Kelson SJ, Kapuscinski AR, Timmins D, Ardren WR. Fine-scale genetic structure of brook trout in a dendritic stream network. Conservation Genetics. 2015;16(1):31-42
67.Nislow KH, Lowe WH. Influences of logging history and stream pH on brook trout abundance in first-order streams in New Hampshire. Transactions of the American Fisheries Society. 2003l;132(1):166-171
68.Warren DR, Likens GE, Buso DE, Kraft CE. Status and distribution of fish in an acid-impacted watershed of the northeastern United States (Hubbard brook, NH). Northeastern Naturalist. 2008;15(3):375-390
69.Lamy JB, O'Donnell BC, Villamagna AM, Morrill TR, Nugent BJ, Hoekwater JC. Genetic analysis reveals a complex mosaic of admixture in brook trout in a historically fragmented watershed. North American Journal of Fisheries Management. 2023;43(4):947-961
70.Old Maps of New Hampshire. Grafton County NH Maps [Internet]. 2009. Available from: http://www.old-maps.com/nh/nh_CoGrafton.htm [Accessed: December 10, 2023]
71.Logging in Lincoln: Beebe River [Internet]. 2021. Available from: https://logginginlincoln.com/Beebe_River.html [Accessed: December 10, 2023]
72.Gove B. Logging Railroads along the Pemigewasset River. Littleton NH, USA: Bondcliff Books; 2006. 166 p
73.Hagan JM, Irland LC, Whitman AA. Changing timberland ownership in the Northern Forest and implications for biodiversity. Manomet Center for Conservation Sciences, Report # MCCS-FCP2005-1. 2005. Brunswick, Maine, 25 p. Available from: https://www.iatp.org/sites/default/files/181_2_78270.pdf
74.USDA. Practical Guide for Strategic Wood Additions (SWA) to Streams in Northern New England (ME, NH and VT). 2018. Natural Resources Conservation Services. Available from: https://efotg.sc.egov.usda.gov/api/CPSFile/22821/395_VT_PS_Stream_Habitat_Improvement_and_Management-SWA_Practical_2020
75.Schuett-Hames D, Pleus AE, War J, Fox M, Light J. TFW monitoring program method manual for the large woody debris survey. Washington State Dept. of Natural Resources Timber, Fish and Wildlife Agreement. 1999. TFW-AM9-99-004. DNR #106. Available from: https://www.dnr.wa.gov/publications/fp_tfw_am9_99_004.pdf
76.Richmond DA, Fausch KD. Characteristics and function of large woody debris in subalpine rocky mountain streams in northern Colorado. Canadian Journal of Fisheries and Aquatic Sciences. 1995;52(8):1789-1802
77.Overton KO, Wollrab SP, Roberts BC, Radko MA. R1/R4 (Northern/Intermountain Regions) fish and fish habitat standard inventory procedures handbook. 1997. Gen. Tech. Rep. INT-GTR-346. Ogden, UT: U.S. Department of Agriculture, Forest Service, Intermountain Research Station. 73 p. DOI: 10.2737/INT-GTR-346
78.Morrill T. Wild Brook Trout (Salvelinus fontinalis) Demographics and Movement in the Presence of Undersized Road Crossings in Headwater Streams in Central New Hampshire [Thesis]. Plymouth: Plymouth State University; 2019
79.Hoekwater J. Brook Trout (Salvelinus fontinalis): Wood Additions and Culvert Removal in Headwater Streams: A Trophic Cascade Perspective [Thesis]. Plymouth: Plymouth State University; 2020
80.Mumby PJ. Statistical power of non-parametric tests: A quick guide for designing sampling strategies. Marine Pollution Bulletin. 2002;44(1):85-87
81.Riaz M, Mahmood T, Arslan M. Non-parametric versus parametric methods in environmental sciences. Bulletin of Environmental Studies. 2016;1:128-130
82.Leuven RSEW, Brock TCM, van Druten HAM. Effects of preservation on dry and ash-free dry weight biomass of some common aquatic macroinvertebrates. Hydrobiologia. 1985;127:151-159
83.Zippin C. The removal method of population estimation. The Journal of Wildlife Management. 1958;22(1):82-90
84.Carle FL, Strub MR. A new method for estimating population size from removal data. Biometrics. 1978;34:621-630
85.Schneider JC, Laarman PW, Gowing H. Manual of fisheries survey methods II: with periodic updates. Michigan Department of Natural Resources: Fisheries Special Report, 25. 2000. Ann Arbor, MI
86.Carlander KD. Handbook of Freshwater Fishery Biology. Dubuque, Iowa: Brown; 1950. p. 281
87.Utz R, Hartman KJ. Density-dependent individual growth and size dynamics of central Appalachian brook trout (Salvelinus fontinalis). Canadian Journal of Fisheries and Aquatic Sciences. 2009;66:1072-1080
88.Frankiewicz P, Radecki-Pawlik A, Wałęga A, Łapińska M, Wojtal-Frankiewicz A. Small hydraulic structures, big environmental problems: Is it possible to mitigate the negative impacts of culverts on stream biota? Environmental Reviews. 2021;29(4):510-528
89.Flores L, Giorgi A, González JM, Larranaga A, Díez JR, Elosegi A. Effects of wood addition on stream benthic invertebrates differed among seasons at both habitat and reach scales. Ecological Engineering. 2017;106:116-123
90.Al-Zankana A, Matheson T, Harper D. Adding large woody material into a headwater stream has immediate benefits for macroinvertebrate community structure and function. Aquatic Conservation: Marine and Freshwater Ecosystems. 2021;4:930-947
91.Deane A, Norrey J, Coulthard E, McKendry DC, Dean AP. Riverine large woody debris introduced for natural flood management leads to rapid improvement in aquatic macroinvertebrate diversity. Ecological Engineering. 2021;163:106197
92.White LS, Charles G, Fausch KD, Josh GH, Saunders WC, Rosenfeld J. Response of trout populations in five Colorado streams two decades after habitat manipulation. Canadian Journal of Fisheries and Aquatic Sciences. 2011;68(12):2057-2063
93.Keeley ER. Demographic responses to food and space competition by juvenile steelhead trout. Ecology. 2001;82:1247-1259
94.Grossman G, Ratajczak R, Wagner C, Petty J. Dynamics and regulation of the southern brook trout (Salvelinus fontinalis) population in an Appalachian stream. Freshwater Biology. 2010;55(7):1494-1508
95.Waters FT. Annual production by a stream brook charr population and by its principal invertebrate food. Environmental Biology of Fishes. 1982;7(2):165-170
96.Layman CA, Rypel AL. Secondary production is an underutilized metric to assess restoration initiatives. Food Webs. 2020;25:e00174
Written By
Joshua C. Hoekwater, Amy M. Villamagna, Brigid C. O’Donnell, Tyson R. Morrill, Ben J. Nugent and Jared B. Lamy
Submitted: 14 January 2024Reviewed: 14 March 2024Published: 26 April 2024