Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
Our team is growing all the time, so we’re always on the lookout for smart people who want to help us reshape the world of scientific publishing.
Home >
Books >
Herbs and Spices - New Perspectives in Human Health and Food Industry [Working Title]
Open access peer-reviewed chapter - ONLINE FIRST
Immune-Enhancing Medicinal Plants: Are they a One Health, One Welfare Actor?
Written By
Aurel Vasiu, Vasile Cozma, Anamaria Cozma-Petruț, Mihai Băieș, Marina Spînu, Emӧke Páll, Diana Olah, Carmen Dana Șandru, Gheorghiță Duca, Köbölkuti Lorand and Gabriel Gati
Submitted: 20 February 2024Reviewed: 22 February 2024Published: 08 April 2024
Medicinal plants have represented accessible and highly bioavailable remedies in traditional therapeutic and preventive practices of numerous populations worldwide. Veterinary treatments based on medicinal plants are also widespread, mainly targeting the control or prevention of parasitic diseases. Scientific support of the immune-stimulating efficacy of plants or their extracts in animals is less documented. The immunological activity of alcoholic plant extracts was investigated in numerous animal classes, starting from Pisces, through Reptilia and Aves and reaching Mammalia, envisaging their effects on innate and adaptive cell-mediated immunity, which the authors mean to share in this chapter, also providing a comparison of variable reactivity within and between the classes.
Faculty of Veterinary Medicine, University of Agricultural Sciences and Veterinary Medicine, Cluj-Napoca, Romania
Vasile Cozma
Faculty of Veterinary Medicine, University of Agricultural Sciences and Veterinary Medicine, Cluj-Napoca, Romania
Anamaria Cozma-Petruț
Faculty of Pharmacy “Iuliu Haţieganu”, Department of Bromatology, Hygiene, Nutrition, University of Medicine and Pharmacy, Cluj-Napoca, Romania
Mihai Băieș
Faculty of Veterinary Medicine, University of Agricultural Sciences and Veterinary Medicine, Cluj-Napoca, Romania
Marina Spînu*
Faculty of Veterinary Medicine, University of Agricultural Sciences and Veterinary Medicine, Cluj-Napoca, Romania
Institute of Research and Development for Montanology, Cristian, Sibiu, Romania
Emӧke Páll
Faculty of Veterinary Medicine, University of Agricultural Sciences and Veterinary Medicine, Cluj-Napoca, Romania
Institute of Research and Development for Montanology, Cristian, Sibiu, Romania
Diana Olah
Faculty of Veterinary Medicine, University of Agricultural Sciences and Veterinary Medicine, Cluj-Napoca, Romania
Carmen Dana Șandru
Faculty of Veterinary Medicine, University of Agricultural Sciences and Veterinary Medicine, Cluj-Napoca, Romania
Institute of Research and Development for Montanology, Cristian, Sibiu, Romania
Gheorghiță Duca
Institute of Research and Development for Montanology, Cristian, Sibiu, Romania
Köbölkuti Lorand
Faculty of Veterinary Medicine, University of Agricultural Sciences and Veterinary Medicine, Cluj-Napoca, Romania
Gabriel Gati
Environmental Health Center, Cluj-Napoca, Romania
School of Public Health, College of Political, Administrative and Communication Sciences, Babeș-Bolyai University, Cluj-Napoca, Romania
*Address all correspondence to: marina.spinu@gmail.com
1. Introduction
Medicinal plants have long served as readily available and highly bioavailable remedies in the traditional therapeutic and preventive approaches of various global populations. For centuries, a plethora of natural phytochemical compounds derived from plants have demonstrated medicinal properties, aiding in healing and disease prevention [1]. The application of phytotherapy in veterinary treatments is also widespread, primarily focused on managing or preventing parasitic and infectious diseases. There are multiple factors contributing to the growing interest in herbal medications within the field of veterinary medicine. One significant factor is the widely held belief in the population that medicinal plants are not only effective but also safer when compared to synthetic compounds. Additionally, a key driver is the economic aspect, given that herbal treatments often prove to be more cost-effective than traditional interventions [2, 3]. Phytotherapeutic remedies offer a viable alternative for treating organic livestock, eliminating the need for synthetic drugs. However, there is comparatively limited scientific documentation supporting the immune-stimulating effectiveness of plants or their extracts in animals.
Medicinal plants, rich in various bioactive components, have found widespread application in the development and synthesis of drugs, such as aspirin. The field of natural product chemistry has expanded globally, aiming to explore increasingly potent and cost-effective drugs with minimized side effects [4, 5, 6]. Medicinal plants have the capability to regulate the innate immune system by boosting the activity of protease inhibitors and lytic enzymes within immune cells and molecules, enabling them to respond effectively against invading pathogens [4, 7, 8]. The bioactive compounds commonly found in aromatic plants typically exist in mixtures, predominantly composed of phenolics and terpenes, which are chemically identified by their aromatic rings [9].
Phytogenics sourced from the Lamiaceae family, formerly called Labiatae [10] are widely used plant-based additives in livestock and aquaculture [11, 12, 13, 14]. The Lamiaceae family, consisting of over 7000 species grouped into approximately 240 genera, is widely distributed around the globe, exhibiting diverse heights and habitats [10, 15]. Members of this botanical family display diverse morphological characteristics, appearing as herbs, herbaceous plants, shrubs, or tree species [16]. Within them, numerous aromatic and medicinal plants are found, widely utilized across traditional and modern medicine, the food industry, and cosmetics [17, 18]. Extracts prepared from these plants yield bioactive compounds utilized for their growth promotion, antimicrobial, antioxidant, antitumoral, immunostimulant, anti-inflammatory, antiviral, antifungal, insecticidal, sedative, antiangiogenic, neuroprotective, and carminative potential [10, 12, 19, 20]. Phenolics and terpenes represent a category of volatile bioactive compounds derived from plants, possessing medicinal and biotechnological significance. Phenolic compounds, including monoterpenes such as carvacrol and its isomer thymol, have been subject to extensive study [12, 21, 22]. These compounds serve as the primary constituents in essential oils extracted from a variety of aromatic plants belonging to the Lamiaceae family. The most well-known members of this family include a variety of aromatic spices like thyme, mint, oregano, basil, sage, savory, rosemary, self-heal, hyssop, lemon balm, and some others with more limited use [23]. Animal feeds can be enhanced by incorporating natural additives such as oregano (Origanum vulgare L) and thyme (Thymus vulgaris-L). These aromatic herbs, rich in bioactive compounds, not only contribute to the flavor profile of animal products but also offer potential health benefits. Research suggests that certain components in oregano and thyme, such as phenolics and terpenes, may have antimicrobial, antioxidant, and immunomodulatory properties, making them valuable supplements in animal nutrition. Including these herbs in animal diets can be a holistic approach to promoting both palatability and potential health advantages in livestock and poultry.
Immunosuppressive diseases present a significant risk to the well-being and efficiency of livestock, including pigs and poultry, leading to financial losses for producers. These diseases can be initiated by diverse factors, including viral, bacterial, and parasitic infections, as well as stressors related to the environment and management, such as insufficient nutrition or overcrowding [1]. These illnesses can weaken the animal’s immune system, making it more vulnerable to secondary infections and reducing its ability to fight against pathogens [24].
Understanding the mechanisms of immune processes and improving the possibilities for their control are particularly important [25]. The immune system comprises an intricate network of both innate and adaptive components, possessing an exceptional ability to adapt and respond to a wide array of challenges [26].
The inherent physiological reactions of all living organisms against invading pathogens represent fundamental host responses. Prokaryotes, in particular, protect themselves through the utilization of restriction enzymes and clustered regularly interspaced palindromic repeats (CRISPRs), enabling the degradation of foreign pathogens attempting to invade [27].
The training and development of key components in the host’s innate and adaptive immune system are significantly influenced by the microbiome [26, 28, 29, 30]). The emergence of specific branches within the immune system, especially those linked to adaptive immunity [30], has occurred alongside the evolution of a complex microbiota [26]. The close interplay between the microbiome and the immune system implies that enhancing the resident microbiome, whether through pre/probiotic supplements or appropriate interventions to prevent the disruption of microbial communities, can strengthen immune defenses [28, 31].
Some of the best known immunologically active plant extracts are the lectins, such as phytohemagglutinin M or P (American pokeweed, Fam. Phytolaccaceae, Genus Phytolacca) or concanavalin A obtained from Canavalia ensiformis (L.) DC. (jack-bean, Fam. Fabaceae, Genus Canavalia), as a mitogen to prompt T-lymphocyte cell division, stimulating proliferation by causing an increase in cAMP [32].
Similarly, the nettle family, Urticaceae, include a numerous number of well-known and useful plants, including the genus Urtica, Boehmeria, Pipturus, and Debregeasia (about 2625 species, 53 genera). Urtica dioica L., as one of the best-known representatives, contains numerous compounds, including phenols, sterols, fatty acids, alkaloids, flavonoids, lignans, and terpenoids. Ther plant is used for its antioxidant, anti-inflammatory, antimicrobial, antiparasitic, protective for cardiovascular, nervous and digestive systems, antidiabetic and antiaging properties [33].
The immunological activity of alcoholic plant extracts was investigated in numerous animal classes, starting from Pisces, through Reptilia and Aves and reaching Mammalia, envisaging their effects on both innate and adaptive immune system branches, which the authors mean to share in this chapter, also providing a comparison of variable reactivity within and between the classes.
2. Plant active principles and innate cell-mediated immune response in fish
The immune system in fish, similar to higher vertebrates, includes both the innate and adaptive immune systems [5, 34]. The defense system of fish bears numerous similarities to the immune system of higher vertebrates, being well developed and integrated. In fish, the season and temperature changes are two factors that strongly influence both the immune response and antibody synthesis [35]. Although significant progress has been made in recent years regarding the immune system in fish, it is necessary to expand research on characterizing defense cell populations and their mechanisms of action. The thymus, spleen, kidneys, and the ontogenetic lymphoid process are the main lymphoid organs in fish [36]. Immunity in fish, like in other animal categories, represents their ability to cope with the attack of specific pathogens, whether they are viruses, bacteria, or parasites [37]. The environmental temperature significantly affects the immune response of fish, as they are among the most primitive organisms that possess an adaptive immune system consisting of lymphocytes, immunoglobulins, major histocompatibility complex (MHC), and, last but not least, T-cell receptors (TCR) [36]. According to studies, pathogens on the body surface or those that enter the immune organisms can be destroyed or only attenuated. On the other hand, it seems that the state of immunity is what limits the possibility of attachment of ectoparasites or intestinal parasites [38]. Researchers believe that humoral factors are present in tissues, internal fluids, and the mucus layer, including lysozyme, alexin, interferon, and properdin. Fish produce lysozyme, lectins, complement, and acute-phase proteins (C-reactive proteins, amyloid A and P). The complement system intervenes against pathogens and inflammatory processes, playing a crucial role in both humoral and cellular immunity [39]. The presence of complement in the tegument mucus has been highlighted, providing it the ability to act as a primary defense barrier for the organism [40].
In several fish species, the use of monoclonal antibodies has allowed the identification of the two types of lymphocytes, namely B lymphocytes and T lymphocytes [34]. According to studies by Scapigliati et al. [41], few recognize peripheral T cells, and a majority of these antibodies are directed against immunoglobulins and B cells that carry Ig on their surface.
Due to limited data on the ontogeny and differentiation of lymphoid cells in fish, researchers have employed monoclonal antibodies for the superior phenotypic characterization of these lymphocytes and various cellular subpopulations [42].
Knowledge of the degree of development of both the immune system and its functions play a crucial role. The organization and functioning of the immune system in all its complexity are directly influenced by the phylogenetic development of the organism [26]. The presence of suitable receptors and effectors is what determines both the functioning and accuracy of the immune response [43, 44].
Phagocytosis in fish represents one of the main innate immune mechanisms targeting immediate defense. The in vitro testing of phagocytosis can not only reveal the antimicrobial reactivity of Pisces but also help in identifying cellular responses to environmental compound, thus testing the effects of plant chemical compounds on fish phagocytes. Farmed fish (Onchorhyncus mykis) are exposed to farming technology (housing, feeding, nests, grouping, anti-parasitic and other treatments, etc.) and temperature changes in the aquatic habitat due to microclimate changes, which will presumably induce stress and impaired leukocyte function. Under these circumstances, we hypothesized that an immune-modulating treatment should stimulate the innate cell-mediated immune response. Mint, present on the banks of the rivers where trout species naturally dwell, containing as biologically active ingredients volatile oils (comprising menthol, menthone, menthyl acetate, pinene, limonene, cineole, and flavonoids such as rutin and rosmarinic acid) could alleviate the immune suppression induced by thermal stress in farmed rainbow trout.
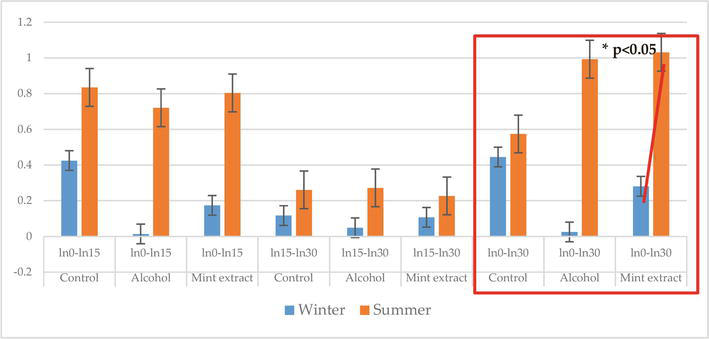
In an in vitro experiment using Mentha piperita L. (Order Lamiales, Fam. Lamiaceae, Genus Mentha) alcoholic extract prepared in concordance with the provision of the German Pharmacopeia in a carbon particle inclusion test [45], we demonstrated (Figure 1) that temperature of the season significantly influenced the leukocyte numbers and subpopulations; therefore, the immune cell-stimulating activity of the alcoholic mint due to its active principles proved to be a temperature-boosted process, which probably stimulated both the monocytes and the heterophiles, more intensely during summer than during winter (p < 0.05) [46].
Figure 1.
The phagocytic activity induced by the alcoholic mint extract in fish: There was a statistically significant increase (p < 0.05) in the carbon particle engulfment activity over the 0 to 30-minute in vitro testing period in summer (columns in orange) versus winter (blue columns), which indicated a potential stress-alleviating activity of the plant. Phagocytosis was measured during three periods of incubation 0 to 15 min and 15 to 30 min and overall activity from 0 to 30 min.
The mint alcoholic extract was used relying on no harmful effects reported in the literature and in dosages defined in previous in vitro studies, (unpublished data), in an amount of 20 μL/2 mL (v/v) [46].
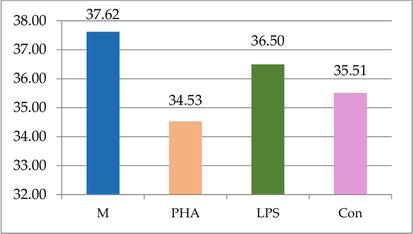
Similarly, the in vitro lymphocyte blast transformation test offers valuable information on how the adaptive immune system is ready to fight infections, by contact between the lymphoid cells and lectins, PHA and ConA. Testing the capacity of fish leukocytes to respond in an in vitro whole blood system (Figure 2) [45] to stimulation with lectins indicated that there were no significant differences between the mitogens used (PHA, ConA, and bacterial lipopolysaccharide), but their efficacy in fish was poor.
Figure 2.
The in vitro cell-mediated response to plant and bacterial lectins in fish: PHA and ConA induce non-significantly lower activation of fish lymphocytes when compared to bacterial lectin LPS (when compared to control culture M, there was a 8.2% decrease in PHA-treated variant, while the lectins induced only a 2.9% - LPS and 5.6% - PHA decrease, respectively).
3. Reptilia and their immune reactivity to plant extracts
The immune system of reptiles has been studied to a much-reduced extent compared to that of mammals, birds, or even fish and amphibians, probably due to their reduced economic impact. Further, the possibility of working with a high number of these individuals is minimal, for reasons such as their conservation and protection, the difficulty of keeping them in captivity, and so on [47]. Still, several research have investigated the ability of the immune system of reptiles to respond to antigenic challenges indicating the presence of agglutinating and neutralizing antibodies to various bacteria and viruses; all responses appear to be T-lymphocyte dependent [48]. In snakes, the immune response can vary depending on a number of extrinsic factors, such as nutritional status, environmental temperature, population density, season, and other ecological factors, while stress caused by capture or transport, various infections, and metabolic imbalances can also diminish immune processes [49]. It appears that the kinetics and amplitude of the humoral immune response correlated with the development of lymphoid tissues are affected by seasonal variations [50]. Secondary immune response has also been highlighted in some reptiles. Antibody-secreting cells have been demonstrated in the spleen, bone marrow, and peripheral blood but not in other lymphoid tissues such as the thymus or the cloacal complex [48]. Two classes of immunoglobulins IgM and a non-IgM (IgY)—the homolog of IgA found in mammals—have been described [51], the syntheses of which are temperature dependent [52, 53, 54, 55, 56]. Immunoglobulin classes of IgA, IgD, and IgE have not been identified in reptiles [57].
Cell-mediated immunity has been investigated by means of spleen cell transplantation and skin allografts and xenografts [53, 54, 58]. It must be taken into account that the conditions encountered by snakes in laboratories are not similar to those in nature (in terms of temperature conditions, day-night cycles, microhabitat, food, etc.); these “immuno-ecological” considerations can profoundly affect the “performance” of the immune system [47].
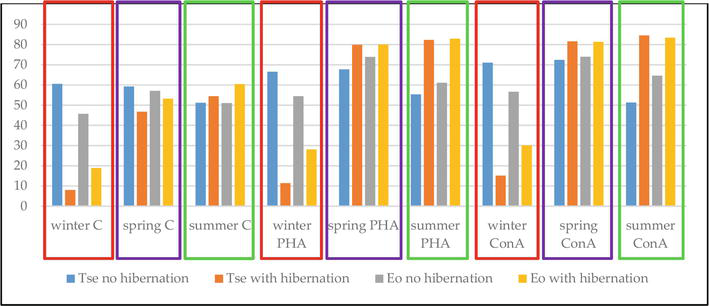
Testing the influence of lectins on two different species of turtles, Trachemys scripta elegans and Emys orbicularis (Figure 3), revealed a temperature-dependent response, with similar patterns in both species for the same season. Both lectins similarly increased the lymphocyte responses to the lectins when comparing the same season, while significant differences were observed due to the presence or absence of hibernation and between winter versus spring/summer (p < 0.001)(Köbölkuti, unpublished data).
In snakes (Vipera berus, Vipera amodytes), the PHA exerted a non-significantly different stimulation, still being useful to evaluate the response potential of the cell-mediated immunity [59]. No statistically significant differences were recorded between cultivation variants (Figure 4) or species, while the animals were hibernating, at an incubation temperature of 20°C, but there were differences between species (p = 0.002 in V. berus and p = 0.007 in V. ammodytes).
Figure 4.
Stimulation indices recorded by the in vitro blast transformation test for Viperinae lymphocyte growth: no significant differences between the variants during hibernation (there was a slight increase of 12.12% in the PHA co-cultivated variant versus untreated control).
Thus, it was concluded that the lifecycle rather than the species/variety influences the immune response to plant lectins in Viperinae.
4. Aves: a new evolutionary approach to improved immune responsiveness
In birds, in contrast to mammals, one of the primary lymphoid organs is the bursa of Fabricius, a gut-associated lymphoid tissue [60], characterized by 12–15 folds containing follicles, an organ designed for the maturation of B lymphocytes, with lymph nodes being absent [61]. The medullary portion of the follicles develops between embryonic days 11 and 13, while the cortical region’s development begins around the time of hatching and concludes by the end of the second week of life. However, regarding the immune system, the differences between birds and mammals are not limited to this aspect [62]. Due to the lack of cross-reactivity for the three classes of immunoglobulins, namely IgA, IgM, and IgG, distinct names have been recommended, despite the immunoglobulin structures of birds being similar to those of mammals. Additionally, in birds, some immune-competent cells, such as T lymphocytes that lack receptors for erythrocytes, exhibit differences compared to those in mammals. Since the immune response in birds is conditioned by a set of factors different from those encountered in mammals, there is a need for organs, effectors, and regulatory mechanisms to ensure the appropriate amplitude of the immune response. Within the bursa of Fabricius, where B lymphocytes undergo differentiation, researchers have established that the fundamental mechanism of the immune response in birds closely resembles that in mammals [63]. Bursa-dependent B cells exclusively handle antibody production in birds. The chicken bursa serves as a valuable animal model for exploring B cell differentiation and function due to its distinct role in B cell development and its convenient anatomical accessibility on specific days [61, 64]. Although there is currently a wealth of data regarding the complexity of effector immune mechanisms in birds, many aspects related to the mode of action, especially lymphocytic interaction, in assessing the exact role of bursa of Fabricius, and elucidating the evolution of the B lymphocyte, remain not fully understood [65]. It is possible that the unresolved aspects concerning the biology of avian lymphocytes may be due to the lack of suitable markers, such as the formation of E rosettes, which could serve as a basis for preparative procedures [66]. The antibodies found in birds, especially in chickens, do not activate the human complement system and do not react with rheumatoid factor, nor with murine anti-IgG antibodies or human or bacterial Fc receptors, unlike those in mammals [66, 67]. Among the immune effectors found in birds are T lymphocytes, B lymphocytes, macrophages, natural killer cells (NK cells), cytotoxic T lymphocytes, and antigen-presenting cells (APC). Research has highlighted the presence of two functionally distinct cell types in birds, analogous to those in mice, namely TH (helper T cells) and TS (suppressor T cells). These cells can interact with both T and B lymphocytes [61, 68]. In contrast to other organisms, in birds, activated B lymphocytes at the plasmocyte stage produce three types of antibodies: IgG/IgY (serum/yolk), IgM, and IgA [69]. In birds, antibody-dependent cellular cytotoxicity (ADCC) is the ability of an adherent cell with phagocytic capacity, non-T, non-B, equipped with Fc receptors, to exhibit activity against syngeneic or allogeneic erythrocytes. Antigen-presenting cells are activated under the influence of molecular factors and cytokines [70]. In the absence of antigenic stimulation, the transformation of the immune system appears to follow a well-established pattern [71].
Urtica dioica (stinging nettle), a plant well known for the biological properties of its phenolic compounds, could be considered as a natural alternative source for possible applications in industrial areas, such as food/feed, cosmetics, and also phytomedicine. Its anti-oxidative capacity was considered the foundation of its biological activities, which still depends on the habitat and external factors that influence its composition.
In a study aiming to investigate the effect of pollution on the efficacy of the active compound of the stinging nettle [72], changes in body weight, total leukocyte numbers (TL), and delayed type hypersensitivity (DTH) were monitored in 28 day-old chickens (n = 12/group) injected subcutaneously twice (days 0 and 7), with 0.5 ml of alcoholic nettle extracts harvested from both unpolluted and polluted areas, against untreated and 700 alcohol-treated controls. No significant changes were recorded for the weight gain/period between groups from polluted and unpolluted areas. Nevertheless, the total leukocyte numbers decreased in the group from polluted areas, this change being reflected by the local hypersensitivity test (wattle test), the size of the infiltration being diminished by the extract from the unpolluted area.
Investigations of the in vitro effects of lectins in adult hens vaccinated against Newcastle disease (day 0, day 14) during a three-week experiment indicated that the reactivity of the birds increased gradually versus PHA and ConA toward the end of the experiment, with a more pronounced stimulation capacity of the Canavalia ensiformis (L.) DC. than that of Phytolacca americana L. (unpublished data, Table 1).
Rec.
Control
PHA
LPS
Con
Day 0
69.27 ± 13.31
64.20 ± 81.90
68.66 ± 10.67
67.12 ± 12.79
Day 9
59.57 ± 11.20
52.99 ± 9.09
54.87 ± 11.1
56.21 ± 7.28
Day 21
67.37 ± 6.11
70.93 ± 12.73
77.23 ± 21.2
76.22 ± 12.23
Table 1.
Stimulation indices in hens subjected in vitro to antigenic stimulation: The increase in the LPS activity when compared to that of PHA and ConA indicated an increased activity of B lymphocytes.
Rec = sampling on days 0, 9, and 21; Control = untreated variant; PHA = phytohaemagglutinin M of Phytolacca americana L.; LPS = lipopolysaccharide of bacterial cell wall; Con = concanavaline A of Canavalia ensiformis (L.) DC. Values are expressed as arithmetic means ± standard error.
A comparison of reactivity in Pisces, Reptilia, and Aves to the lectins of Canavalia ensiformis (L.) DC. and Phytolacca americana L. indicated a gradual increase paralleling the phylogenetic progress of the species.
In mammals, the initial defense mechanism consists of physical barriers such as the skin and mucosal membranes [73, 74]. The epithelial cells from respiratory, gastrointestinal, and urogenital tracts not only create mechanical barriers but also release various antimicrobial factors, such as antimicrobial peptides and defensins, with a crucial role in the innate immune response [74, 75]. Other vital cellular components of the innate immune system in cattle include neutrophils, NK cells, dendritic cells (DC), gamma delta T cells, mucosa-associated invariant, macrophages, and granulocytes [74, 76].
The development of the immune system commences early in gestation across all mammalian species. As the fetus grows, the immune system undergoes a multitude of changes, with cells emerging and adopting specialized functions [77, 78]. The maturation of the immune system in cattle coincides with embryonic development and subsequently progresses during fetal development, leading to an increasingly intricate immune response to antigenic stimulation. The development of the immune system in calves takes place early during the fetal period. Both sides of the immune response, cellular and humoral, develop in parallel [79].
Since 1973, Schultz and colleagues [80] demonstrated that fetal peripheral blood lymphocytes are present starting from day 45, while cells carrying IgM are detected from day 59, and those carrying IgG emerge from day 135. The stage at which the immune system matures is highly significant, allowing the developing organism to distinguish between self and non-self antigens. This differentiation helps prevent undesired immune responses against its own components in the future. Therefore, from the age of 3 to 4 months of gestation onward, the fetus does not mount an immune response to either self or non-self antigens. This immunological tolerance is achieved through the T-mediated mechanism, avoiding immune reactions against self and non-self antigens that may mistakenly appear outside the first 3 months of gestation [80]. Through in vitro stimulation tests, we can assess cellular reactivity. By cultivating cells in the presence of polyclonal mitogens that specifically stimulate T lymphocytes, such as phytohemagglutinin (PHA) and concanavalin A, we can demonstrate both T lymphocyte reactivity and the integrity of intercellular cooperation pathways among antigen-presenting cells (APC), T lymphocytes, and B lymphocytes (APC-T-B). The cellular immune system of calves can respond to antigenic stimulation from birth. The immune system in calves matures progressively, evolving from conception to achieving full maturity around 6 months after birth; the response varies with age and the structure of the antigen [81]. Cellular effectors, both specific and nonspecific, play a significant role in defending the organism against infections [25]. They participate in defense mechanisms through processes like phagocytosis and pinocytosis, releasing lytic enzymes that contribute to the destruction of microorganisms. Cells with the ability to capture, ingest, and digest pathogens are actively involved in the phagocytic process [82]. Studies have demonstrated a notable increase in the blood concentration of γδ T lymphocytes, particularly in newborn calves, accounting for 25–30% of peripheral blood leukocytes. In contrast, adult cattle typically exhibit values ranging from 3 to 10% [83]. The epitheliochorial type of placenta found in ruminants does not allow the transplacental passage of immunoglobulins, resulting in calves being born agammaglobulinemic [84, 85]. Studies have indicated that starting from the fourth month of gestation, the fetal response to antigenic stimulation in cattle is possible, although the antibodies are IgM, specific to a primary response, and they disappear rapidly. Newborns exhibit a lower level of immune competence compared to adults, as a consequence of the transient presence of a high titer of endogenous corticosteroids during the perinatal period [86].
Similar to calves, piglets are agammaglobulinemic at birth. Their ability to survive is directly reliant on obtaining maternal immunity through the intake of colostrum and milk [87]. The protective function of maternal immunity involves various factors, particularly specific systemic humoral immunity.
The farmed subjects, representatives of the class as well as the complex structure and functionality of the mammalian immune system, trigger numerous studies targeting the value of alternative immune-stimulating therapies in those species.
In a study researching the adjuvant and immune cell-stimulating potential of certain plants from the perspective of farmed herbivores differing by their digestive physiology, alcoholic vegetal extracts were tested in vitro [88]. Romanian Spotted dairy cows, Angora goats, and Romanian draft horses were sampled for blood, and the samples were treated in vitro in the blast transformation test [45] with alcoholic extracts of Calendula officinalis L., Echinacea angustifolia L., and E. purpurea L. The growth indices were the lowest in goats (C. officinalis L.- 58.52 ± 10.02%, E. angustifolia L. - 50.06 ± 11.67%, E.purpurea L.-50.79 ± 10.98%) and higher in bovine (C. officinalis L.– 69.9 ± 2.65%, E.angustifolia L.- 74.9 ± 10.1%, p < 0.05) and increased toward E. angustifolia L. versus C. officinalis L.; further, the responses to C. officinalis L., E. angustifolia L. and E.purpurea L. were the most pronounced in horses. Thus, there were differences between the tested species, probably due to the differences in the anatomy as well as the physiology of their immunity.
It is well-known that the stress of various origins represents an important factor impeding on vaccination results in bovine. In a study aimed at investigating the anti-stress and immunological influence of the oral administration of a marigold tea (antioxidant activity 85.10 ± 5.21%) to adult bovine, vaccinated against multiple antigens with Cattle Master, it was shown that the stress levels constantly decreased in marigold tea-treated animals (1.09 ± 0.047 to 0.064 ± 0.02) and did not significantly vary in controls. The blast transformation test indicated lower SI% and a prolonged response to vaccination in the treated group (40.65 ± 14.91 to 79.82 ± 3.34 versus 50.62 ± 5.93 and 81.31 ± 2.25%) by the end of the experiment. It could be concluded that the marigold tea exerted a stress-lowering effect but prolonged the response to vaccination when orally administered as an infusion to adult bovine.
As mentioned before, the outcome of the host-pathogen interaction microbial aggression highly depends on immunity, and medicinal plants, available on the pastures, could enhance and strengthen the resistance against diseases.
Research was carried out on Mangalitza pigs (suckling, weaned piglets and sows) raised for tasty lean meat mainly on low-input farms to monitor the effects of plants readily available from local resources such as Calendula officinalis L., Satureja hortensis L., Allium sativum L., Coriandrum sativum L., and Cucurbita maxima L. An in vitro leukocyte blast transformation test was carried out by treating each 0.2 ml of the whole blood mixed with RPMI1640 culture medium (1:4 v/v) with 1.5 μL/0.2 mL of the selected alcoholic extracts (70°, v/v). The results indicated that statistically significant differences were present between the young age groups, suckling and weaned piglets (p = 0.017 to 0.000016) for all plants except marigold, for weaned piglets and sows (p = 0.0001–0.0359) for all plant extracts, while for suckling piglets/sows p = 0.0035 and p = 0.0461 were recorded for thyme and garlic, respectively. No toxic effects were present for the tested alcoholic extracts of Calendula officinalis L., Satureja hortensis L., Allium sativum L., Coriandrum sativum L., and Cucurbita maxima L. (unpublished data). This experiment underlined that the plant extracts used, well-known for their biological effects, impacted animals based on the age of the pigs and plant family, proving their immune-stimulating capacity [89].
Multiple beneficial biological effects of medicinal plants, underlined by millennia of ethno-pharmaceutical practices, prove to still hide part of their constructive potential. In animal farming and veterinary medicine, a support for One Health, One Welfare for consumers, medicinal plants could provide help, strengthening the fight in preserving both.
Further, the differentiated effect of plant extracts on different classes of animals, based on their phylogeny, impacted on individuals grounded on their level of immune development as well as age and raising technology, proving their immune-stimulating capacity allocated to their own taxonomy.
Use of plants in preserving animal health and welfare by stimulating immune functions as shown in the case of mint, marigold, pumpkin, summer savory, and garlic seems encouraging within the One Health, One Welfare scheme.
Numerous investigations, such as those described at the level of laboratory tests in the present work, provided proof that the biological activity of plant extracts was both animal and plant species dependent. Tailored treatment and feed supplementation schemes founded on the laboratory results could be beneficial in farmed animals as well, providing basis for sustainability of the enterprises at lower costs.
We acknowledge the financial support provided by the European Union’s Horizon 2020 research and innovation program, under grant agreement no. 816172, PPILOW and the Ministry of Research, Innovation and Digitization, CNCS -UEFISCDI, within PNCD III and ERANET Core Organic Co-fund ROAM Free #249/2021.
None of the authors have any existing or possible conflict of interest, including financial, personal or any other relationship which could influence their scientific work.
References
1.Nabi F, Ahmed J, Tao W, Lu Q , Bhutto ZA, Qadir A, et al. An updated review on efficiency of penthorum chinense pursh in traditional uses, toxicology, and clinical trials. BioMed Research International. 2023:4254051. DOI: 10.1155/2023/4254051
2.Kubkomawa HI, Nafarnda DW, Tizhe MA, Daniel TK, Shua NJ, Ugwu CC, et al. Ethno-veterinary health management practices amongst livestock producers in Africa: A review. Advances in Agricultural Sciences. 2020;6:1-006
3.Romero B, Susperregui J, Sahagún AM, Diez MJ, Fernández N, García JJ, et al. Use of medicinal plants by veterinary practitioners in Spain: A cross-sectional survey. Frontiers in Veterinary Science. 2022;9:1060738. DOI: 10.3389/fvets.2022.10607384
4.Awaad AS, Al-Jaber NA. Antioxidant natural plant. Recent Progress in Medicinal Plants: Ethnomedicine: Source & Mechanism I. 2010;27:1e35
5.Awaad E, Awaad A. Role of medicinal plants on growth performance and immune status in fish. Fish & Shellfish Immunology. 2017;67:40-54. DOI: 10.1016/j.fsi.2017.05.034
6.Hassan BAR. Medicinal plants (importance and uses). Pharmaceutica Analytica Acta. 2013. DOI: 10.4172/2153-2435.1000e139
7.Sakai M. Current research status of fish immunostimulants. Aquaculture. 1999;172:63e92
8.Van Hai N. The use of medicinal plants as immunostimulants in aquaculture: A review. Aquaculture. 2015;446:88e96
9.Christaki E, Giannenas I, Bonos E, Florou-Paneri P. Chapter 2 - Innovative uses of aromatic plants as natural supplements in nutrition. In: Florou-Paneri P, Christaki E, Giannenas I, editors. Feed Additives. Cambridge, Massachusetts: Academic Press; 2020. pp. 19-34. DOI: 10.1016/B978-0-12-814700-9.00002-9
10.Ramos da Silva LR, Ferreira OO, Cruz JN, de Jesus Pereira Franco C, Oliveira Dos Anjos T, Cascaes MM, et al. Lamiaceae essential oils, phytochemical profile, antioxidant, and biological activities. Evidence-based Complementary and Alternative Medicine. 2021;2021:6748052. DOI: 10.1155/2021/6748052
11.Elumalai P, Kurian A, Lakshmi S, Faggio C, Esteban MA, Ringø E. Herbal immunomodulators in aquaculture. Reviews in Fisheries Science & Aquaculture. 2020;29(1):33-57. DOI: 10.1080/23308249.2020.1779651
12.Firmino JP, Galindo-Villegas J, Reyes-Lopez FE, Gisbert E. Phytogenic bioactive compounds shape fish mucosal immunity. Frontiers in Immunology. 2021;12:695973
13.Franz CM, Baser KHC, Hahn-Ramssl I. Chapter 3 - herbs and aromatic plants as feed additives: Aspects of composition, safety, and registration rules. In: Florou-Paneri P, Christaki E, Giannenas I, editors. Feed Additives. Cambridge, Massachusetts: Academic Press; 2020. pp. 35-56. DOI: 10.1016/B978-0-12-814700-9.00003-0
14.Napoli E, Siracusa L, Ruberto G. New tricks for old guys: Recent developments in the chemistry, biochemistry, applications and exploitation of selected species from the Lamiaceae Family. Chemistry & Biodiversity. 2020;17(3):e1900677. DOI: 10.1002/cbdv.201900677
15.Kallunki JA, Heywood VH. Flowering plants of the world. Brittonia. 1994;46:4. DOI: 10.2307/2806914
16.Harley RM. Checklist and key of genera and species of the Lamiaceae of the Brazilian Amazon. Rodriguésia. 2012;63(1):129-144
17.Nieto G. Biological activities of three essential oils of the Lamiaceae family. Medicines (Basel). 2017;4:63. DOI: 10.3390/medicines4030063
18.Sun J, Sun P, Kang C, Zhang L, Guo L, Kou Y. Chemical composition and biological activities of essential oils from six lamiaceae folk medicinal plants. Frontiers in Plant Science. 2022;13:919294. DOI: 10.3389/fpls.2022.919294
19.Luz TRSA, Leite JAC, de Mesquita LSS. Seasonal variation in the chemical composition and biological activity of the essential oil of Mesosphaerum suaveolens (L.) Kuntze. Industrial Crops and Products. 2020;153:112600
20.Ouakouak H, Benchikha N, Hassani A, Ashour ML. Chemical composition and biological activity of Mentha citrata Ehrh., essential oils growing in southern Algeria. Journal of Food Science and Technology. 2019;56(12):5346-5353
21.Kosakowska O, Węglarz Z, Pióro-Jabrucka E, Przybył JL, Kraśniewska K, Gniewosz M, et al. Antioxidant and antibacterial activity of essential oils and Hydroethanolic extracts of Greek oregano (O. Vulgare L. subsp. hirtum (link) Ietswaart) and common oregano (O. Vulgare L. subsp. vulgare). Molecules. 2021;26(4):988. DOI: 10.3390/molecules26040988
22.Mączka W, Twardawska M, Grabarczyk M, Wińska K. Carvacrol-a natural phenolic compound with antimicrobial properties. Antibiotics (Basel). 2023;12(5):824. DOI: 10.3390/antibiotics12050824
23.Uritu CM, Mihai CT, Stanciu GD, Dodi G, Alexa-Stratulat T, Luca A, et al. Medicinal plants of the family Lamiaceae in pain therapy: A review. Pain Research & Management. 2018;8:7801543. DOI: 10.1155/2018/7801543
24.Düpjan S, Dawkins MS. Animal welfare and resistance to disease: Interaction of affective states and the immune system. Frontiers in Veterinary Science. 2022;9:929805. DOI: 10.3389/fvets.2022.929805
25.Chaplin DD. Overview of the immune response. The Journal of Allergy and Clinical Immunology. 2010;125(2 Suppl. 2):S3-S23. DOI: 10.1016/j.jaci.2009.12.980
26.Belkaid Y, Hand TW. Role of the microbiota in immunity and inflammation. Cell. 2014;157(1):121-141. DOI: 10.1016/j.cell.2014.03.011
27.Dunin-Horkawicz S, Klaus KO, Lupas AN. Prokaryotic protein networks mediating innate immunity and apoptosis. Journal of Molecular Biology. 2014;426:1568-1582. DOI: 10.1016/j.jmb.2013.11.030
28.Di Sotto A, Vitalone A, Di Giacomo S. Plant-derived nutraceuticals and immune system modulation: An evidence-based overview. Vaccines (Basel). 2020;8(3):468. DOI: 10.3390/vaccines8030468
29.Kogut MH, Lee A, Santin E. Microbiome and pathogen interaction with the immune system. Poultry Science. 2020;99(4):1906-1913. DOI: 10.1016/j.psj.2019.12.011
30.Zheng D, Liwinski T, Elinav E. Interaction between microbiota and immunity in health and disease. Cell Research. 2020;30:492-506. DOI: 10.1038/s41422-020-0332-7
31.Mezouar S, Chantran Y, Michel J, Fabre A, Dubus J-C, Leone M, et al. Microbiome and the immune system: From a healthy steady-state to allergy associated disruption. Human Microbiome Journal. 2018;10:11-20. DOI: 10.1016/j.humic.2018.10.001. ISSN 2452-2317
32.Parker CW. The role of intracellular mediators in the immune response. In: Cohen S, Pick E, Joost J, editors. Oppenheim, Biology of the Lymphokines. Amsterdam: Academic Press, Elsevier; 1979. pp. 541-583. DOI: 10.1016/B978-0-12-178250-4.50024-6
33.Taheri Y, Quispe C, Herrera-Bravo J, Sharifi-Rad J, Ezzat SM, Merghany RM, et al. Urtica dioica-derived phytochemicals for pharmacological and therapeutic applications. Evidence-based Complementary and Alternative Medicine: eCA. 2022:4024331. DOI: 10.1155/2022/4024331
34.Secombes C, Wang T. The innate and adaptive immune system of fish. Infectious Disease in Aquaculture Prevention and Control. 2001;231:3e68
35.Cerbu CG, Spînu M, Şandru CD, Vasiu A, Olah D, Brudașcă GF. Influence of the raising system on phagocytosis in the endangered Danube salmon (Hucho hucho). Global Ecology and Conservation. 2018;16:e00514
36.Viswanath K. Fish immune system and its nutritional modulation for preventive health care. Animal Feed Science and Technology. 2012;173(1-2):111-133
37.Buchmann K. Evolution of innate immunity: Clues from invertebrates via fish to mammals. Frontiers in Immunology. 2014;5:459. DOI: 10.3389/fimmu.2014.00459
38.Zapata A, Diez B, Cejalvo T, Gutiérrez-de Frías C, Cortés A. Ontogeny of the immune system of fish. Fish & Shellfish Immunology. 2006;20(2):126-136. DOI: 10.1016/j.fsi.2004.09.005
39.Yano T. Assays of hemolytic complement activity. In: Stolen JS, Fletcher TC, Anderson DP, Kaatari SL, Rowley AF, editors. Techniques in Fish Immunol. Vol. 2. USA: SOS Publications; 1992. pp. 131-141
40.Lambris JD. The chemistry, biology and phylogeny of C3 complement today. Complement profiles. Karger Basel. 1993;I:16-45
41.Scapigliati G, Romano N, Abelli L. Monoclonal antibodies in fish immunology: Identification, ontogeny and activity of T- and B-lymphocytes. Aquaculture. 1999;172:3-28
42.Castillo A, Sánchez C, Domínguez J, Kaatari SL, Villena AJ. Ontogeny of IgM and IgM-bearing cells in rainbow trout. Developmental and Comparative Immunology. 1993;17:419-424
43.Dozmorov IM, Dresser D. Immune system as a sensory system. International Journal of Biomedical Sciences. 2010;6(3):167-175
44.Wu L, Qin Z, Liu H, Lin L, Ye J, Li J. Recent advances on phagocytic B cells in teleost fish. Frontiers in Immunology. 2020;27(11):824. DOI: 10.3389/fimmu.2020.00824
45.Khokhlova IS, Spinu M, Krasnov BR, Degen AA. Immune response to fleas in a wild desert rodent: Effect of parasite species, parasite burden, sex of host and host parasitological experience. The Journal of Experimental Biology. 2004;207(Pt. 16):2725-2733. DOI: 10.1242/jeb.01090
46.Spînu M, Pall E, Niculae M, Vasiu A, Brudaşcă GF, Cerbu C, et al. Seasonal changes of phagocytic activity enhanced by an acoholic mint extract in cultured rainbow trout. IJPER. 2018;52(4s):s86-s90. DOI: 10.5530/ijper.52.4s.80
47.Jurd RD. In: Turner RJ, editor. Immunology: A Comparative Approach. John Wiley & Sons Ltd: England; 1994. pp. 137-147
48.Good RA, Papermaster BW. Ontogeny and phylogeny of adaptive immunity. Advances in Immunology. 1944;4:1-115
49.Cooper EL, Klempau AE, Zapata AG. Reptilian immunology. In: Gans C, Billett F, Maderson PFA, editors. Biology of the Reptilia. Vol. 14. New York: Wiley-Interscience; 1985. pp. 599-678
50.Frye FL. Biomedical and Surgical Aspects of Captive Reptile Husbandry. Florida: Krieger Publishing Company; 1991. pp. 101-149, 209-281
51.Salantino SK, Minton SA. Immune response of snakes. Copeia. 1973;3:504-515. Available from: https://www.jstor.org/stable/1443116
52.Ambrosius H. Immunoglobulins and antibody production in reptiles. In: Marchalonis JJ, editor. Comparative Immunology. Oxford, England: Blackwell; 1976. pp. 298-334
53.Borysenko M. Changes in spleen histology in response to antigenic stimulation in the snapping turtle, Chelydra sepentina. Journal of Morphology. 1976;149:243-264
54.Cooper EL. Skin transplant rejection in Apodan amphibians and Lacertan reptiles. American Zoologist. 1969;9:333
55.Karstad L. Influence of low body temperature on establishment of prolonged infections in animal reservoirs. In: Sci. Proc. 100th Ann. Meeting Amer. Vet. Med- Assoc. Chicago: American Veterinary Medical Association; 1963. pp. 186-193
57.Kollias GV. Immunological aspects of infectious disease in reptiles and amphibians. In: Hoff GL, Frye FL, Jacobson ER, editors. Diseases of Amphibians and Reptiles. New York NY: Plenum Press; 1984
58.Borysenko M, Cooper EL. Lymphoid tissue in the snapping turtle, Chelydra sepentina. Journal of Morphology. 1972;138:487-481
59.Kobolkuti L, Cadar D, Czirjak G, Niculae M, Kiss T, Sandru C, et al. The effects of environment and physiological cyclicity on the immune system of Viperinae. The Scientific World Journal. 2012:574867. DOI: 10.1100/2012/574867
60.Ekino S, Sonoda K. New insight into the origin of IgG-bearing cells in the bursa of Fabricius. International Review of Cell and Molecular Biology. 2014;312:101-137
61.Cheng J, Lei H, Xie C, Chen J, Yi X, Zhao F, et al. B lymphocyte development in the Bursa of Fabricius of young broilers is influenced by the gut microbiota. Microbiology Spectrum. 2023;11(2):e0479922. DOI: 10.1128/spectrum.04799-22
62.Davison F. The importance of the avian immune system and its unique features. In: Avian Immunology. Amsterdam: Elsevier, Academic Press; 2014. pp. 1-9
63.Salvante KG. Techniques for studying integrated immune function in birds. The Auk. 2006;123(2):575-586
64.Funk PE, Palmer JL. Dynamic control of B lymphocyte development in the bursa of Fabricius. Archivum Immunologiae et Therapiae Experimentalis (Warsz). 2003;51:389-398
65.Oláh I, Felföldi B, Benyeda Z, Nagy N, Kovács T. The morphology and differentiation of stromal cells in the cortex of follicles in the bursa of Fabricius of the chicken. Anatomical Record (Hoboken, NJ. 2007). 2022;305(11):3297-3306. DOI: 10.1002/ar.24893
66.Sharma JM. The structure and function of the avian immune system. Acta Veterinaria Hungarica. 1997;45(3):229-238
67.Romito M, Viljoen GJ, Du Plessis DH. Eliciting antigen-specific egg-yolk IgY with naked DNA. BioTechniques. 2001;31(3) 670, 672, 674-5
68.Yamamoto Y, Komatsu T, Suzuki Y. Localization of neuropeptides in endocrine cells of the chicken thymus. The Journal of Veterinary Medical Science 1997;59(7):601-603
69.Warr GW, Magor KE, Higgins DA. IgY: Clues to the origins of modern antibodies. Immunology Today. 1995;16(8):392-398
70.Murtaugh MP, Foss DL. Inflammatory cytokines and antigen presenting cell activation. Veterinary Immunology and Immunopathology. 2002;87(3-4):109-121
71.Carter JH. The immune system as a model for pattern recognition and classification. Journal of the American Medical Informatics Association. 2000;7(1):28-41. DOI: 10.1136/jamia.2000.0070028
72.Vasiu A, Sandru C, Spinu M, Ungureanu E, Marian F, Popescu S, et al. Environmental pollution degree changes the biological activity of nettle plant extracts in chickens. Current Perspectives on Medicinal and Aromatic Plants. 2023;6(2):84-89. DOI: 10.38093/cupmap.1337889
73.Rodríguez-Habibe I, Celis-Giraldo C, Patarroyo ME, Avendaño C, Patarroyo MA. A comprehensive review of the immunological response against foot-and-mouth disease virus infection and its evasion mechanisms. Vaccines (Basel). 2020;8(4):764. DOI: 10.3390/vaccines8040764
74.Vlasova AN, Saif LJ. Bovine immunology: Implications for dairy cattle. Frontiers in Immunology. 2021;29(12):643206. DOI: 10.3389/fimmu.2021.643206
75.Hans M, Madaan HV. Epithelial antimicrobial peptides: Guardian of the oral cavity. International Journal of Peptide. 2014;2014:370297. DOI: 10.1155/2014/370297
76.Burrell CJ, Howard CR, Murphy FA. Innate immunity. In: Fenner and White’s Medical Virology. Amsterdam: Elsevier; 2017. pp. 57-64. DOI: 10.1016/B978-0-12-375156-0.00005-9
77.Cortese VS. Neonatal immunology. The Veterinary Clinics of North America. Food Animal Practice. 2009;25(1):221-227. DOI: 10.1016/j.cvfa.2008.10.003
79.Barrington GM, Parish SM. Bovine neonatal immunology. The Veterinary Clinics of North America. Food Animal Practice. 2001;17(3):463-476. DOI: 10.1016/s0749-0720(15)30001-3
80.Sordillo LM, Aitken SL. Impact of oxidative stress on the health and immune function of dairy cattle. Veterinary Immunology and Immunopathology. 2009;128(1-3):104-109. DOI: 10.1016/j.vetimm.2008.10.305
81.Chase CC, Hurley DJ, Reber AJ. Neonatal immune development in the calf and its impact on vaccine response. The Veterinary Clinics of North America. Food Animal Practice. 2008;24(1):87-104. DOI: 10.1016/j.cvfa.2007.11.001
82.Rosales C, Uribe-Querol E. Phagocytosis: A fundamental process in immunity. BioMed Research International. 2017;2017:9042851. DOI: 10.1155/2017/9042851
83.Guzman E, Hope J, Taylor G, Smith AL, Cubillos-Zapata C, Charleston B. Bovine γδ T cells are a major regulatory T cell subset. Journal of Immunology. 2014;193(1):208-222. DOI: 10.4049/jimmunol.1303398
84.Bragg R, Macrae A, Lycett S, Burrough E, Russell G, Corbishley A. Prevalence and risk factors associated with failure of transfer of passive immunity in spring born beef suckler calves in Great Britain. Preventive Veterinary Medicine. 2020;181:105059. ISSN 0167-5877
85.Gabriël S, Geldhof P, Phiri IK, Cornillie P, Goddeeris BM, Vercruysse J. Placental transfer of immunoglobulins in cattle infected with Schistosoma mattheei. Veterinary Immunology and Immunopathology. 2005;104(3-4):265-272. DOI: 10.1016/j.vetimm.2004.12.010
86.Fair T. The contribution of the maternal immune system to the establishment of pregnancy in cattle. Frontiers in Immunology. 2015;6:7. DOI: 10.3389/fimmu.2015.00007
87.Salmon H, Berri M, Gerdts V, Meurens F. Humoral and cellular factors of maternal immunity in swine. Developmental & Comparative Immunology. 2009;33(3):384-393. ISSN 0145-305X. DOI: 10.1016/j.dci.2008.07.007
88.Vasiu A, Sandru C, Pall E, Ungureanu E, Marian F, Popescu S, et al. Host and plant taxonomy as influential factors in controling in vitro specific cell-mediated response in herbivores. Current Perspectives on Medicinal and Aromatic Plants. 2023a;6(2):78-83. DOI: 10.38093/cupmap.1337829
89.Spînu M, Rusu LA, Niculae M, Pall E, Vasiu A, Olah DI, et al. In vitro age dependent cell-mediated responses to antimicrobial plant extracts in free range Mangalitza pigs. In: The 6th International Mediterranean Symposium on Medicinal and Aromatic Plants (MESMAP-6), 16th to 19th April 2020, Izmir, Selcuk (Ephesus), Turkey
Written By
Aurel Vasiu, Vasile Cozma, Anamaria Cozma-Petruț, Mihai Băieș, Marina Spînu, Emӧke Páll, Diana Olah, Carmen Dana Șandru, Gheorghiță Duca, Köbölkuti Lorand and Gabriel Gati
Submitted: 20 February 2024Reviewed: 22 February 2024Published: 08 April 2024