Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
Evaluation of T cell clonality has been costly and/or time-consuming. The analysis of TCR β-chain constant region 1 (TRBC1) provides a simplified immunophenotypic assessment of T-cell clonality. Furthermore, due to the high variability of T-cell populations, there is a need for reliable and robust panels to sort normally from pathological T-cells. The CD27 and CD45RA phenotypic profiling strategy associated with the evaluation of TCRCBeta1 in the same cytometry tube is able to separate normal T Cell populations from clonal populations, gating clusters of cells according to their CD45RA x CD27 expression and then evaluate their TCRCBeta1 status. TCRCBeta1 marker is not only easily implemented in routine immunophenotyping but is also faster and much cheaper than the analysis of TCR-VBeta families either by PCR or by flow cytometry.
HEMOSC – Hematology and Hemotherapy Center of Santa Catarina, Laboratory Division - Cell Markers Laboratory, Florianopolis, SC, Brazil
Renata Kalfeltz
HEMOSC – Hematology and Hemotherapy Center of Santa Catarina, Laboratory Division - Cell Markers Laboratory, Florianopolis, SC, Brazil
*Address all correspondence to: mdanielahemosc@gmail.com
1. Introduction
Flow cytometry is a powerful tool for analyzing lymphoid subsets. This technology evolved from the simple quantification of TCD4 and TCD8 cells in HIV-positive patients to a broader comprehension of the lymphoid compartment. Recent publications describe impressive values as high as 85 different types of TCD4 and 45 types of TCD8.
The number of different T-cell phenotypes according to their functions makes the clonality assessment of T-cell populations even more challenging.
For a very long time, flow cytometry specialists all around the world waited for a so-to-speak “T cell kappa/lambda,” meaning a marker that could easily identify clonal T Cell populations with accuracy.
It turns out that the β-chain of T-cell receptor αβ (TCR) structure has a variant and a constant region. The variability of the constant region? It is region 1 or region 2 as simple as that.
Recently, screening of anti-TCR monoclonal antibodies revealed a high specificity of clone JOVI-1 for the TCR β-chain constant region 1 (TCRBC1) domain, providing an opportunity for a simplified immunophenotypic assessment of T-cell clonality. There are two genes associated with the β-chain constant region: TCRBC1 and TCRBC2. Each TCR (and therefore each T cell) irreversibly selects a TCR β-chain constant region encoded by either TCRBC1 or TCRBC2 for expression in a mutually exclusive manner, similar to the kappa and lambda immunoglobulin light chain utilization by B-cells. Therefore, normal TCRαβ T-cell populations are expected to exhibit comparable numbers of TCRBC1-positive and TCRBC2-positive subsets [1].
Since the vast majority of T-cell malignancies derive from the largely dominant TCRαβ T-cell subset, this approach is applicable to most scenarios where a neoplastic T-cell population is suspected.
2.1 Back to basics: a brief recap about the immune system
The cellular component of the innate (or natural) immune system consists of all the cells that lack immunologic memory, have phagocytic properties or release inflammatory mediators, such as neutrophils, eosinophils, monocytes, and natural killer cells.
The acquired (or adaptive) immune response is probably one of the most complex and advanced systems in known biology, with the ability to identify and memorize virtually any foreign antigen. It involves the proliferation of antigen-specific B and T cells, as well as their complex interactions.
While the innate response occurs to the same extent regardless of how many times the antigen is encountered, the acquired response improves with repeated exposure (1).
Both systems are deeply and beautifully intertwined: activated innate immune cells convey the information about the nature and origin of the antigen to the adaptive immune cells, which will elaborate the proper and specific response [2, 3, 4, 5].
Flow cytometry identifies the major types and subtypes of lymphocytes.
2.2 T cells
T cells are a heterogeneous group of short- and long-lived cells.
Under normal circumstances, the long-lived cells, typically contained within the naïve subset, are quiescent, remaining in a non-cycling state for months to years while awaiting encounter with cognate antigen [2, 3, 4, 5]. The short-lived cells are generally contained within the effector and memory subsets, undergoing variable levels of cell cycling in response to antigens encountered throughout the lifetime of the host.
2.3 Sorting out different types of T-cells: the CD45RA and CD27 strategy
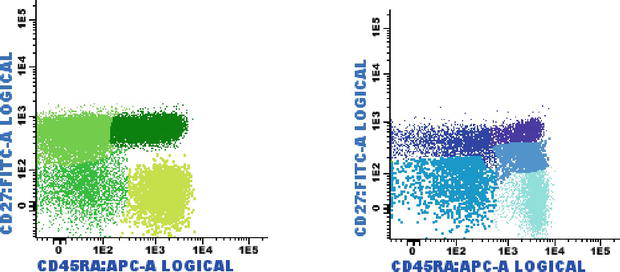
The major T Cell populations divide into naive T cells (those that have not yet contacted foreign antigens), effector T cells (that will disappear once the antigen is eliminated), and memory T cells (that may survive for years and may be easily reactivated if the same antigen appears again) (3). The naive and memory cells can be distinguished by the expression of different versions of the CD45 molecule: naive cells express CD45RA, and memory cells express CD45RO (1); these antigens can help identify different T cell subsets: naive (CD27+ CD45RA+), central/transitional memory (CM/TM CD27+ CD45RA negative), effector memory (EM CD27 negative and CD45RA negative) and terminally differentiated (TD CD27 negative CD45RA+) (Figure 1) [6].
Figure 1.
T-cell subsets based on the expression of CD27 x CD45RA: TCD4+ naïve (dark green), central memory/transitional memory (CM/TM; bright green), effector memory (EM; green) and terminally differentiated (TD; light green). TCD8+ naïve (purple), CM/TM (dark blue), EM (pale blue), and TD (turquoise). Some effector CD8+ T-cells showed dim CD27 positivity (blue).
The naive cells are T-cells yet to confront the enemy.
Effector T cells recognize antigens in lymphoid organs (central) or in peripheral non-lymphoid tissues (terminal) and are activated to perform functions that are responsible for the elimination of microorganisms and, in disease states, tissue damage.
Memory T cells that are generated by T cell activation are long-lived cells with a greater capacity to react against the antigen. After the T cell response subsides, there are many more memory cells of the corresponding clone than immature T cells that existed before the response. These memory cells respond quickly to subsequent encounters with the same antigen and generate new effector cells that eliminate it.
2.4 TCR
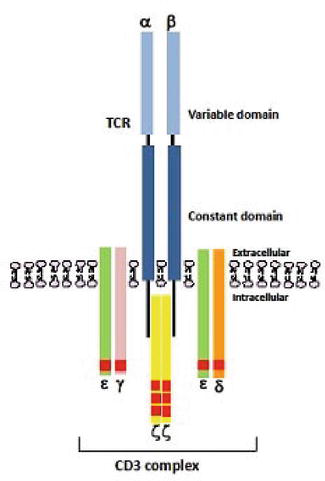
T cell receptor is a membrane-bound heterodimer consisting of two polypeptide chains (α/β or γ/δ) and is associated with cytoplasmic protein CD3. During T-cell development, rearrangement of VJ and VDJ (variable (V), diversity (D), joining (J), and constant (C) region) genes of α/β and γ/δ TCR chains provides the molecular basis for the vast diversity of the T-cell recognition repertoire (Figure 2) [6].
Figure 2.
Schematic representation of a TCR heterodimer consisting of an alpha (α) and beta (β) polypeptide chain, with each polypeptide containing a constant and a variable region.
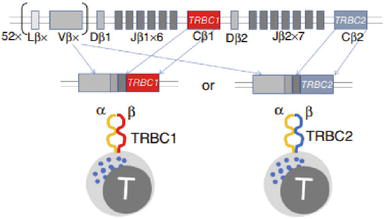
Only one type of β chain can be expressed on an αβ T cell and therefore all cells in a clonal T cell population express the same unique TCR (Figure 3).
Figure 3.
TCRCBeta1 structure and its V(D)J rearrangements [7].
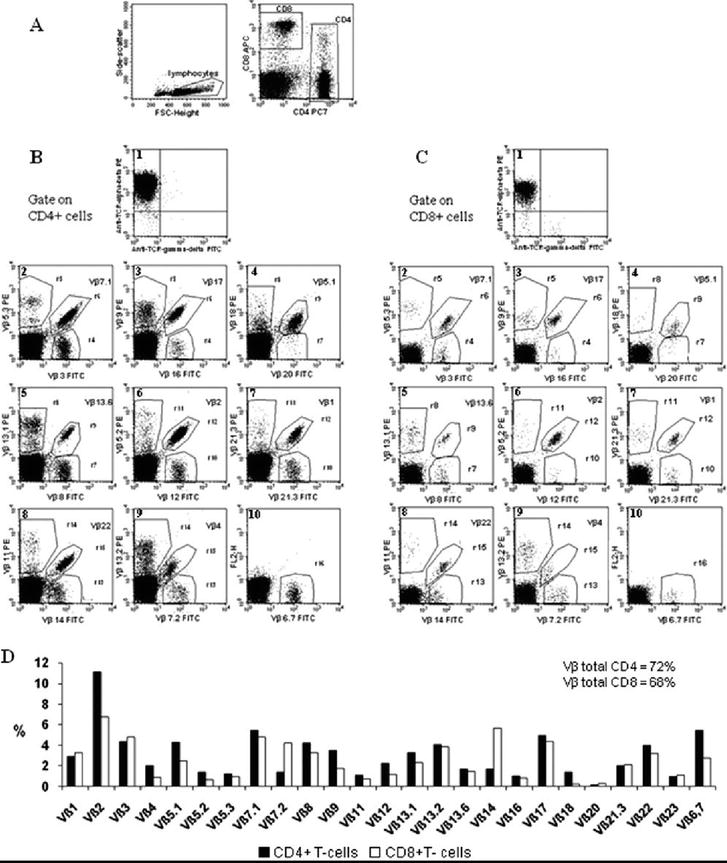
Flow cytometry-based analysis of TCR-β variable region families, employing a set of 25 monoclonal antibodies, allows for the characterization and enumeration of approximately 85% of the total human T-cell repertoire and is a useful target to track clonal expansions.
However, TCR-VBeta families analysis is expensive and time-consuming. TCR-Vβ repertoire analysis by flow cytometry is labor-intensive, costly, difficult to interpret, and of limited sensitivity (Figure 4). In addition, although next-generation sequencing (NGS) of rearranged TCRs has been an alternative tool with greater resolution and analytic sensitivity, it is highly complex, costly, and not usually available in routine diagnostic practice.
Lymphocytes are the unique cells in the body with clonally expressed antigen receptors, each specific for a different antigenic determinant. Each T lymphocyte clone expresses antigen receptors with a unique specificity, which is different from the specificities of the receptors on other clones.
Thus, there are millions of clones of lymphocytes in the body, allowing the recognition and response to millions of foreign antigens.
The activation of lymphocytes follows a series of sequential steps that begin with the synthesis of new proteins necessary for many of the subsequent changes. The immature cells then begin to proliferate, resulting in an increased size of antigen-specific clones, a process called clonal expansion. In some infections, the number of T cells infected by the microorganism can increase more than 50,000 times. This rapid clonal expansion of specific lymphocytes is necessary to keep pace with the ability of microorganisms to rapidly replicate [8, 9].
The rearrangement of antigen receptor genes is a key event in the development of lymphocytes and is responsible for the generation of this diverse repertoire. Each T lymphocyte clone produces an antigen receptor with a unique antigen-binding structure in a genetic process similar to the production of surface immunoglobulins on B lymphocytes. The ability to generate these extremely diverse repertoires does not require an equally large number of genes of different antigen receptors; otherwise, much of the human genome would be dedicated to encoding a large number of TCR molecules.
The genes that encode the different antigen receptors of B and T lymphocytes are generated by the rearrangement, in each lymphocyte, of different gene segments of the variable region (V) with gene segments of diversity (D) and junction (J). This specialized process of rearranging genes at specific locations is called V(D)J recombination.
In αβ T cells, the TCR β chain is the first to be rearranged.
With such a complex genetic mechanism involved in the development of the TCR, it is no wonder that something eventually goes wrong.
Because the Ig and TCR genes are sites of multiple DNA recombination events in B and T cells, and because these sites become active for transcription after recombination, genes from other loci can be abnormally translocated to these loci and, as a result, may be abnormally transcribed. In B and T lymphocyte tumors, oncogenes are often translocated to the Ig or TCR gene loci. These chromosomal translocations are often accompanied by an accentuated transcription of oncogenes and are one of the factors that promote the development of lymphoid tumors [8].
2.6 Recognizing clonal T-cells
Most of the cases of T-cell diseases are easily spotted because of a subpopulation prevalence (TCD4+, TCD8+, T double-positive, or T double-negative). If there is no obvious dominance of a particular T-cell subtype, phenotypic aberrations, most commonly the dim expression of CD3 and dim/negative expression of CD7, maybe the clues.
2.7 Combining the CD45RA and CD27 strategy with TCRCBeta1: separating wheat from chaff
The CD27 and CD45RA phenotypic profile strategy associated with clonality assessment is able to sort out the normal populations from the clonal populations, especially since all clonal populations usually show additional phenotypic aberrancies.
TCRCBeta1, as a strategy to assess T-cell clonality by the addition of a single anti-TRBC1 antibody to a diagnostic flow cytometry T-cell panel, was introduced in 2020 [1, 7, 10, 11, 12].
Clonality detection using TRBC1 should always be performed with multiple other T-cell antigens, sorting out different T-cell subsets and separating neoplastic from benign T-cells. Eight to ten-color flow cytometry panels based on previously published data should be used in immunophenotyping.
Thresholds for percentages of TRBC1-positive events greater than 85% or less than 15% might be defined as clonality in current literature.
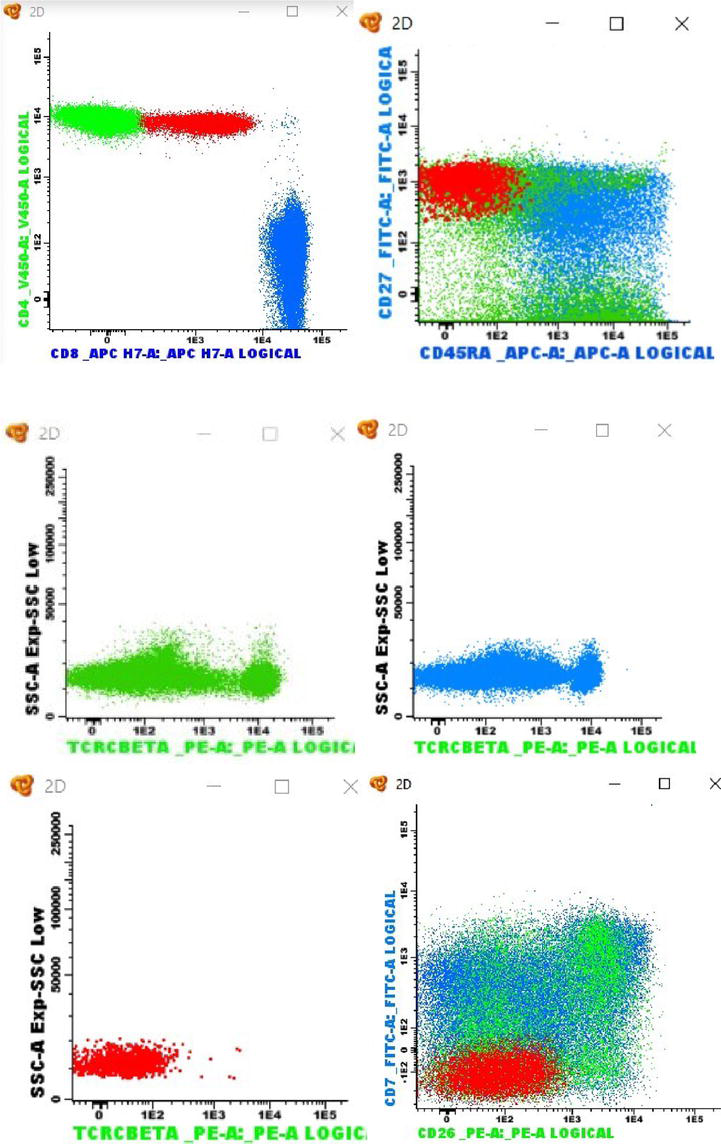
In our laboratory (data not published), to study CD27 and CD45RA phenotypic profile strategy associated with clonality assessment, TCRCBeta1 (clone JOVI-1) was added to EuroFlow’s CLPD-T Tube 2 [13] (CD27, CD45RA, CD8, CD16 and CD56, CD4, CD3, CD45) in the PE channel. The combination of CD27 and CD45RA with TCRCBeta1 proved to be an excellent tool for identifying abnormal T-cell populations, even in small percentages (Figure 5).
Figure 5.
TCR-VBeta family repertoire (from cytometry part a 75A: 743751, 2009) [14].
2.8 Phenotypic features of T-cell chronic lymphoproliferative disorders (T-CLPD)
T-CLPD diagnosis is far from simple; almost 20 different entities are described in 2022 WHO’s edition (Table 1) [15]. According to the type of population restriction, the expression of CD4 and CD8 can be used to formulate a list of diagnostic possibilities and determine what additional information is required for further classification (Table 2) [16].
4th edition
5th edition
T-lymphoblastic leukemia/lymphoma, NOS
Unchanged
Early T-precursor lymphoblastic leukemia/lymphoma
Unchanged
Adult T-cell leukemia/lymphoma
Unchanged
Sezary syndrome
Unchanged
Primary cutaneous CD4-positive small or medium T-cell lymphoproliferative disorder
Nodal peripheral T-cell lymphoma with TFH phenotype
Nodal TFH cell lymphoma, NOS
Table 1.
The 5th edition of the World Health Organization classification of hematolymphoid tumors: Overview of changes and new additions to the classification of T-cell lymphomas.
Disease entities
Distinguishing phenotypic features
Additional diagnostic information
CD4+ CD8−
CTCL / Sézary syndrome
CD7(−) CD26(−) CD23+/−
Characteristic morphology and clinical presentation. HTLV-1(−)
*T-PLL
Usually lacks significant phenotypic aberrancy
80% t(14;14)(q11;q32) or inv.(14) (q11;q32). TCL1 expression
*Adult T-cell leukemia/lymphoma
CD7(−) CD25+ (uniform bright)
HTLV-1+ Endemic Japan and Caribbean
Anaplastic large cell lymphoma
Loss of many pan–T-cell antigens Strong uniform CD30+
Anaplastic morphology ALK gene rearrangement
Angioimmunoblastic
Aberrant phenotype. CK10+/−
Characteristic morphology
*Peripheral CL, NOS
Variable phenotype, often aberrant loss of CD5 and/or CD7
Diagnosis by exclusion of other distinct disease entities
CD4− CD8+
T-cell Large granular lymphocyte leukemia.
Frequent aberrant expression CD5 and/or CD7. Positive expression of NK markers
LGL morphology. Indolent course, associated with cytopenias
Subcutaneous panniculitis-like TCL
Usually only focal CD56, EBV(−), TCR α/β + Perforin+
Must be distinguished from lupus profundus
Hepatosplenic TCL
CD5(−) CD7+ CD16+/− CD56+ CD57(−) TIA-1+ Perforin(−) May be double-negative.
Often TCR γ/δ but may be TCR α/β, EBV(−). Frequent isochromosome 7q. Aggressive clinical course.
Table 2.
Flow cytometric approach to the diagnosis and classification of TCD4+ and TCD8+ lymphoid neoplasms (adapted from flow cytometric immunophenotyping for hematologic neoplasms, 16).
Thus, the immunophenotypic criteria that have been described as of value for the diagnosis of suspected T-CLPD include: (1) deletion of one or more pan-T molecules (CD7, CD5, CD2, and CD3); (2) expression of a molecule or a combination of two or more molecules not usually expressed by a particular maturational stage, and therefore designated as aberrant (CCR7, CD26, CD27, CD28, CD45RA, and CD45RO); (3) discordant expression of CD3 and TCR on the membrane, and (4) co-expression of CD4 and CD8 and/or the absence of expression of both molecules in an important proportion of T cells.
Such criteria, although useful in daily laboratory routine, are fallible, since a clear immunophenotype impairment has only been found in 60–70% of T-CLPD.
The immunophenotypic profile of the neoplastic lymphoid T cell is already relatively well characterized in some entities, such as leukemia, chronic T-cell prolymphocytic leukemia, adult T-cell lymphoma/leukemia, and Tgamma/delta+ hepatosplenic lymphoma. In others, such as in angioimmunoblastic T lymphoma, the immunophenotypic is heterogeneous and complex, resulting from intra-tumoral heterogeneity [17].
3. Closing remarks: narrowing down diagnostic hypothesis
What we learned so far
Due to multiple DNA recombination events and highly active transcription, TCR gene loci are sites prone to the development of lymphoid tumors;
the majority of T-CLPD derive from TCR αβ;
the β-chain of TCR αβ structure has a variant and a constant region, and there are only two genes associated with the β-chain constant region: TCRBC1 and TCRBC2;
assessment of TCRBC1 by flow cytometry is a fast and easy method for establishing T-cell clonality in TCD4+, TCD8+, and double-positive T-CLPDs;
phenotypic evaluation of CD27 and CD45RA provides a normal background to sort out an abnormal T-cell population.
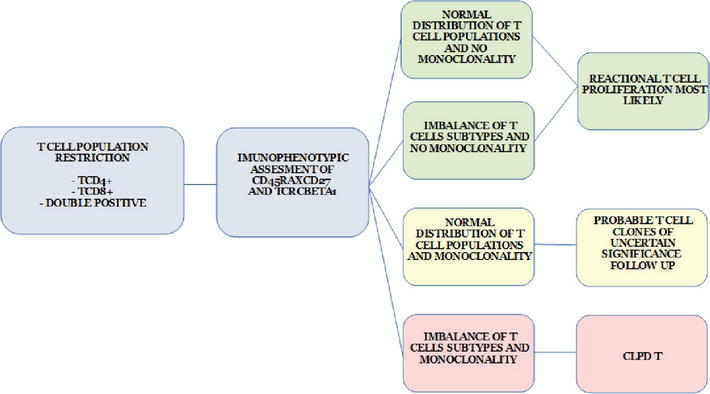
Based on all this information, flow cytometry facilities may develop a strategy for performing a CLPD-T diagnosis (or at least a diagnostic hypothesis) (Figure 6).
Figure 6.
Algorithm strategy for flow cytometric immunophenotyping for screening and classification of T-CLPD.
Once the T-CLPD is established, further flow cytometry evaluation is needed to determine any additional phenotypic features, such as lack of expression of T-cell markers, absence of co-stimulatory molecules, stage of maturation arrest, and expression of NK antigens [18].
With all the information in hand, clinical presentation (leukemic, extranodal, and cutaneous) and pathology will conclude the diagnosis.
The TCRCBeta1 marker is not only easily implemented in routine immunophenotyping but is also faster and much cheaper than the analysis of TCR-VBeta families either by PCR or flow cytometry.
The incorporation of a future TCRCBeta2 in the panel will refine and consolidate the T Cell clonality assessment and provide an almost definitive threshold between reactive and pathological T cells.
References
1.Shi M, Jevremovic D, Otteson GE, Timm MM, Olteanu H, Horna P. Single antibody detection of T-cell receptor αβ Clonality by flow cytometry rapidly identifies mature T-cell neoplasms and monotypic small CD8-positive subsets of uncertain significance. Cytometry Part B (Clinical Cytometry). 2020;98B:99-107
2.Appay V, van Lier RAW, Sallusto F, Roederer M. Phenotype and function of human T lymphocyte subsets: Consensus and issues. Cytometry Part A. 2008;73A:975-983
3.Purnama C, Camous X, Larbi A. An overview of T cell subsets and their potential use as markers of immunological aging. International Trends in Immunity. 2013;1(4):21-32
4.Mahnke YD, Brodie TM, Sallusto F, Roederer M, Lugli E. The who’s who of T-cell differentiation: Human memory T-cell subsets. European Journal of Immunology. 2013;43:2797-2809. DOI: 10.1002/eji.201343751
5.Bretschneider I, Clemente MJ, Meisel C, Guerreiro M, Streitz M, Hopfenmuller W, et al. Discrimination of T-cell subsets and T-cell receptor repertoire distribution. Immunologic Research. Jan 2014;58(1):20-27. DOI: 10.1007/s12026-013-8473-0
6.van der Burg M, Kalina T, Perez-Andres M, Vlkova M, Lopez-Granados E, Blanco E, et al. The EuroFlow PID orientation tube for flow Cytometric diagnostic screening of primary Immunodeficiencies of the lymphoid system. Frontiers in Immunology. March 2019;10(246):1-11. DOI: 10.3389/fimmu.2019.00246
7.Maciocia PM, Wawrzyniecka PA, et al. Targeting the T cell receptor β-chain constant region for immunotherapy of T cell malignancies. Nature Medicine. 2017;23:1416–1423. DOI: 10.1038/nm.4444
8.Abbas AK, Lichtman AH, Pillai S. Cellular and Molecular Immunology. 8th ed. Philadelphia: Elsevier Saunders; 2015
9.Delves PJ, Roitt IM. Advances in immunology. The immune system. The New-England Journal of Medicine. 2000;343(1):37-49. DOI: 10.1056/NEJM200007063430107
10.Shi M, Olteanu H, Jevremovic D, He R, Viswanatha D, Corley H, et al. T-cell clones of uncertain significance are highly prevalent and show close resemblance to T-cell large granular lymphocytic leukemia. Implications for laboratory diagnostics. Modern Pathology. 2020;33:2046-2057. DOI: 10.1038/s41379-020-0568-2
11.Horna P, Shi M, Jevremovic D, Craig FE, Comfere NI, Olteanu H. Utility of T-cell receptor β constant Chain-1 expression in the diagnosis of peripheral blood involvement by cutaneous T-cell lymphoma. The Journal of Investigative Dermatology. Apr 2021;141(4):821-829.e2. DOI: 10.1016/j.jid.2020.09.011
12.Illingworth A, Wong A, Devitt K, Li W, Furtado FM, Oak J, et al. ICCS Quality and Standards Committee TRBC1 Validation - Practical Guidance. 8 Aug 2023:1-19. Reviewed and approved by: Robert Durso, Regeneron Pharmaceuticals; Lorraine Liu, Vancouver Children’s Hospital; David Ng, University of Utah/ARUP Laboratories; Nina Rolf, Vancouver Children’s Hospital
13.Kalina T, Flores-Montero J, van der Velden VHJ, Martin-Ayuso M, Bottcher S, Ritgen M, et al. JJM van Dongen and a Orfao on behalf of the EuroFlow consortium, EuroFlow standardization of flow cytometer instrument settings and immunophenotyping protocols. Leukemia. 2012;26:1986-2010. DOI: 10.1038/leu.2012.122
14.Salameire D, Le Bris Y, Fabre B, Fauconnier J, Solly F, Pernollet M, et al. Efficient characterization of the TCR repertoire in lymph nodes by flow cytometry. Cytometry Part A. 2009;75A:743751
15.Alaggio R, Amador C, Anagnostopoulos I, et al. The 5th edition of the World Health Organization classification of Haematolymphoid tumours: Lymphoid neoplasms. Leukemia. 2022;36:1720-1748. DOI: 10.1038/s41375-022-01620-2
16.Craig FE, Foon KA. Flow cytometric immunophenotyping for hematologic neoplasms. Blood. 2008;111(8):3941-3967
17.Lima M, Almeida A, Santos AH, Teixeira MA, Algueró ML, Queirós ML, et al. Immunophenotypic analysis of the TCR-Vbeta repertoire in 98 persistent expansions of CD3(+)/TCR-alphabeta(+) large granular lymphocytes: Utility in assessing clonality and insights into the pathogenesis of the disease. The American Journal of Pathology. 2001;159:1861-1868
18.Jevremovic D, Olteanu H. Flow cytometry applications in the diagnosis of T/NK-cell lymphoproliferative disorders. Cytometry Part B. 2019;96B:99-115
Written By
Maria Daniela Holthausen Perico and Renata Kalfeltz
Submitted: 02 February 2024Reviewed: 12 February 2024Published: 14 March 2024