Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
Blastocystis is a very common gastrointestinal protozoan globally distributed; it colonizes humans and non-humans, and in some communities, it reaches prevalences of up to 100%. Blastocystis is transmitted through the fecal-oral route, contaminated food and water, and close contact with animals. There are 34 known subtypes of Blastocystis, and subtypes 1 to 4 (ST1–ST4) are the most common in humans. It should be remembered that its pathogenicity is controversial since some studies have shown that Blastocystis is more prevalent in healthy individuals; who have greater diversity and richness of the intestinal microbiota; other studies suggest that Blastocystis infections occur in individuals with intestinal dysbiosis. In America and Africa, a high incidence of ST1 and ST2 is observed in rural areas. Recent data indicate that Blastocystis is linked with specific gut microbiota profiles and health indicators. Convincing information and tools that distinguish asymptomatic colonization from infection in children have yet to be demonstrated. Although this protozoan can cause disease under certain circumstances, but the attention of Blastocystis may change, as the frequency of Blastocystis subtypes in children may vary depending on the geographic area and local health conditions.
Faculty of Health Sciences, University Juarez of State of Durango, Gómez Palacio, Durango, México
Claudia Muñoz Yañez
Faculty of Health Sciences, University Juarez of State of Durango, Gómez Palacio, Durango, México
*Address all correspondence to: janethguangorenagomez@gmail.com
1. Introduction
A complex and diverse population of microorganisms lives in the human intestine: bacteria, fungi, viruses, archaea, and protozoans [1]. Blastocystis spp. is a very common protozoan; it is a Stramenopile or Heterokonta Eukaryote [2], and it has different forms: vacuolar, avacuolar, granular, cystic, and amoeboid [3]. It colonizes a wide diversity of species in the world, such as humans and non-humans; approximately one billion individuals in the world are infected by this protozoan [4]. It is known that this protist is one of the most prevalent; in some communities, it reaches prevalence of up to 100% [5]. The fecal-oral route Blastocystis, through contaminated food and water and close contact with animals, which are the best-known transmission sources for Blastocystis [6, 7]. Studies have reported that it can cause diarrhea accompanied by abdominal pain, dizziness, vomiting, and weight loss, among others [8, 9]. Blastocystis infection is mainly associated with nonspecific gastrointestinal symptoms, both acute and chronic [10].
It has been identified 34 subtypes of Blastocystis [11, 12, 13], but only 17 are recorded, subtypes one to subtype nine are found in humans (ST1–ST9), and subtypes one to four (ST1–ST4) are the most frequent [14, 15]. It is worth noting that there are studies where Blastocystis is more prevalent in healthy individuals [16] who have a greater diversity and richness of the intestinal microbiota [17] and have even been postulated as a viable probiotic agent [18]. Other research suggests that Blastocystis infection occurs in individuals with intestinal dysbiosis and irritable bowel syndrome [19, 20]. The clinical significance of Blastocystis is still inconclusive [18, 21, 22]. All this dissimilarity may be due to the diversity of coexisting subtypes [15].
In America, a high incidence is observed for ST1 and ST2 [23]. In Mexico, higher prevalence was reported for ST1 (51%) [24] in 2023. A global prevalence of 44.0% was reported in school-age children in a rural area, with a prevalence of 56.5% for ST1, followed by ST2 (26.3%) and ST3 (19.7%). In Colombian children, a study carried out in 2021 reported a global prevalence of 58.2% of Blastocystis, and the first identification of Subtype 16 in humans was made [13]. Also, previously in Colombia, a study in 2015 found that subtype 3 is associated with urticaria in Argentine patients and that allele a134 predominated in patients with symptoms [25]. Later, in 2017, another study in the pediatric population found no statistical association between Blastocystis infection and subtypes with variables such as sex, age, symptoms or sociodemographic stratification [26]. Another study in Panama in apparently healthy school-age children from rural areas in 2020 found a global prevalence of Blastocystis of 74.2%, and the prevalence of ST1 was 42.2% [27].
Some African countries have reported a high prevalence of Blastocystis infection, although data on the prevalence and distribution of Blastocystis subtypes still needs to be available. On this continent, a multicenter study was carried out among certain villages in Senegal. An essential variation in prevalence was observed between the villages, 51.7–100% in this study; additionally, the first report of subtypes 10 and 14 in humans was made [28].
The classification of Blastocystis as pathogenic or non-pathogenic has been problematic since a direct causal link has not been demonstrated [22]. However, some explanations for its pathogenicity are the excretion of proteases and the presence of more than five parasites per field in the intestine, mainly causing an inflammatory process at lamina propria level. Also, the composition of the intestinal microbiota, the immune system, and the nutritional status of the host are some of the decisive factors for the presence of the parasite and its pathogenicity [29, 30]. It is known that Blastocystis and the intestinal microbiota cohabit in the intestinal tract of the host and can interact with each other [31, 32, 33, 34, 35, 36, 37, 38]. Bacteria are the most abundant in the intestinal microbiota, and two main phyla are present: Firmicutes and Bacteroidetes (representing 80%). At lower levels, Proteobacteria, Actinobacteria, Fusobacteria, and Verrucomicrobia are present [39, 40]. Recent data suggest that Blastocystis is associated with specific gut microbiota profiles and health indices. Stensvold et al. (2022) used metabarcoding analyses to identify variations in fecal microbiota diversity between Blastocystis-carrying and non-carrying individuals. Alpha diversity was considerably higher in Blastocystis carriers. Subjects with Blastocystis-positive stools had gut microbiomes associated with eubiosis, while in individuals with negative feces for Blastocystis, the intestinal microbiome was similar to those associated with dysbiosis [41]. Importantly, clinical data were not obtained from the subjects from whom feces were obtained, so the role of Blastocystis as a potential modulator of the intestinal microbiota needs to be known with greater scrutiny.
More sensitive tools that differentiate asymptomatic colonization from infection in children have yet to be demonstrated. Although the parasite can cause disease under certain circumstances, the focus on Blastocystis may shift from a clinical to a public health perspective [42]. On the other hand, the frequency of Blastocystis subtypes in children may vary depending on the geographic area and local health conditions. Since in developing countries, drinking water and hygiene are scarce, in rural communities, contact with animals and children is more frequent, and it is a population that is more vulnerable to infection through the anus-oral route.
It was described for the first time in 1912 [43] by Alexeieff, who named it considered it a yeast and named it Blastocystis enterocola; Brumpt described the parasite and named it Blastocystis hominis [44]. Zierd et al. classified it as a parasite, and using ribosomal DNA sequences, Blastocystis was classified within the kingdom Chromista by Silberman. It was not until 1997 that Clark described the polymorphisms of this protozoan [45].
2.1 Taxonomy
Blastocystis belongs to the kingdom Chromista, Subkingdom Chromobiota, Subphylum Opalinata, class Blastocystea [46]. It is the main unicellular eukaryotic protozoan found in the intestine of mammals [47]. It presents characteristics of the protist kingdom with one or more nuclei, strict anaerobe, and sensitivity to O2. It measures 5–40 μm and reproduces mainly by binary fission.
2.2 Morphology
There are many variations between sizes and the form of presentation according to subtypes, but the primary peculiarity is the appearance of a central vacuole (occupies 90% of the cytoplasm) with metabolic and storage functions, easily observable after staining [48]. In humans, different forms of Blastocystis have been described including vacuolar, avacuolar, granular, multi vacuolar, cystic, and amoeboid form, which is more prevalent in symptomatic individuals [3].
It presents four well-differentiated morphological stages:
2.2.1 Vacuolar form
The form most easily identified in feces, they have a size of 5–15 μm and binary fission reproduction. The diagnosis is made based on vacuolar form. They have a central corpuscle or vacuole composed of lipids and carbohydrates with reserve functions, which compress the cell nucleus and cytoplasm [49].
2.2.2 Ameboid form
This form measures 10 μm, does not have a central body, but has 1 or 2 slow-moving pseudopodia. Cellular debris has been found inside, which suggests that it has a significant role in the nutrition of the microorganism [49].
2.2.3 Granular form
It has 1 to 4 nuclei and measures between 6 and 8 μm. This form is scarce, and three types of granules are distinguished: metabolic, reproductive, and lipidic [50].
2.2.4 Cyst form
They measure 3–10 μm and are ovoid or spherical; the cells include lipid and glycogen deposits and vacuoles. Generally, the isolated nuclei are binucleate. They survive approximately 1 month at room temperature; however, they are sensitive to disinfectants and extreme temperatures [49].
2.3 Life cycle
The life cycle is similar to most protists. Two types of cycles have been described: binary fission and autogamy for the formation of cysts that can be thick or thin-walled, which help in the transmission of the parasite. The avacuolar cell is present in the intestine, passing through the intestinal tract. After the disintegration of vesicles in the cytoplasm, the multi-vacuolar form is generated, which is covered by a thick cell wall. The cystic wall forms under the cell cover disintegrates, resulting in a cyst, which is the infectious cell of Blastocystis spp. Ingestion by a host completes the cycle; excystment is the final route of infection caused by exposure to gastric acid and intestinal enzymes [51].
2.4 Transmission
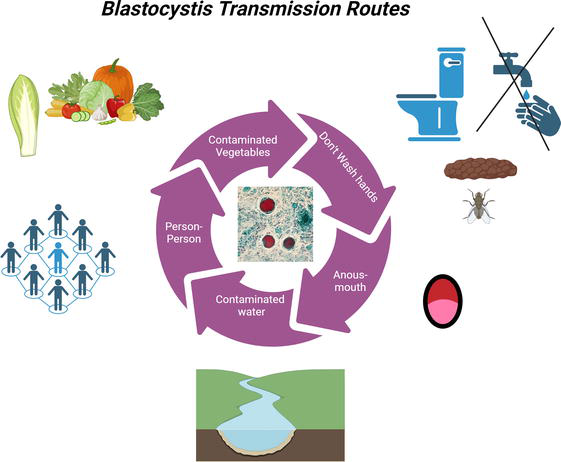
Blastocystis is found in the feces of mammals (humans and primates), as well as amphibians, birds, livestock, and reptiles [52]; its exposure is usually incidental, and the transmission of fecal-oral has been reported as the primary form of transmission. Recent studies suggest that it is present in contaminated water, although exposure to these factors is usually insufficient for infection. Among the risk factors associated with Blastocystis infection are overcrowding, same-sex relationships, contaminated water, low educational level, travelers, living with infected animals, and poor hygiene (Figure 1) [53].
Figure 1.
Blastocystis spp. transmission routes. Blastocystis can be transmitted through fecal-oral routes, such as water and food contaminated with infected feces. It can also spread from person to person through direct contact with contaminated fecal matter.
2.5 Pathogenicity
The pathophysiology of Blastocystis is not entirely known; it is believed to lie mainly in apoptosis, degradation of transmembrane proteins that results in exaggeration of intestinal permeability, induction of pro-inflammatory cytokines and the regulation of nitric oxide [3, 48].
2.5.1 Blastocystis and immune system
Blastocystis spp. can degrade immunoglobulin A (An antibody present in mucosal secretions, predominating in tears, saliva, and gastrointestinal secretions) [48]. Although it is generally accepted that Blastocystis is not an invasive pathogen and cannot phagocytose the host microbiota, pathogenicity depends on the subtype and the Blastocystis proteases [54]; for example, one study described that hydrolases alter the colonic mucus layer.
In vivo, studies with rats have shown that the microorganism’s proteases stimulate the production of Interleukin 8 (IL-8) by colon epithelial cells through mechanisms dependent on nuclear factors, which are responsible for fluid loss and intestinal inflammation in affected individuals. Therefore, it is believed that these enzymes produce hyperplasia of the goblet cells and cecal mucosa in infected rats [49]. It was also experimentally demonstrated that Blastocystis spp. can trigger increased cell proliferation in human colorectal cancer, causing interferon-gamma (IFN γ) and p53 expression deregulation. The subtype with the most pathogenic relationship is ST3, which increases the proliferation of cancer cells by decreasing the apoptosis of these cells and severe inflammatory reactions [55]. Regarding other classes of enzymes, the gene encoding the beta-1,3-galactosyltransferase (b1,3GalT), was acquired laterally by this protozoan and was duplicated several times in its genome; the encoding proteins are involved in molecular mimicry, which helps Blastocystis to camouflage [56].
Blastocystis has been reported in immunocompromised patients, and those under stress since the overproduction of oxygen and hydroxyl free radicals causes oxidation of lipids and proteins, resulting in poor functioning of the body’s tissues and deficient antibody production [57]. Blasto-Ag (an antigen produced by B. hominis) is responsible for the downregulation of peripheral blood cells, and this inhibitory effect is more significant in asymptomatic individuals. In the infection, it is expected to find low levels of IgA with high levels of immunoglobulin G (predominant antibody in cerebrospinal fluid, peritoneal fluid, and blood, which responds to the presence of viruses and bacteria), associated with the overproduction of eosinophils (which suppress infection). In this context, there are studies of children with leukemia where immunoglobulin M (the first antibody produced to fight infections found in blood and lymph) and IgG are high compared to IgA [58].
However, it should be noted that recent studies have identified Blastocystis as part of the microbiota, found in more than 50% of the healthy population, with a tendency to long-term colonization, with the same microorganism found 10 years later [56].
2.6 Subtypes
As previously mentioned, 34 subtypes of Blastocystis spp. have been identified. STs 1–4 are the most prevalent in 90% of carriers; these subtypes are found in 95% of patients with a single infection [14].
2.6.1 Distribution of subtypes by hosts
ST1 and ST2 are also found in other animal species, including monkeys, cows, chickens, pigs, rats, dogs, and nonhuman primates. ST3 has been found in nonhuman primates, pigs, and cattle. ST4 has been described in rodents and monkeys. The rarer subtypes in humans (ST5–8) are found most frequently in other hosts: ST5 is common in cattle, apes, pigs, rats, dogs, and Old World monkeys, while ST6 and ST7 are mainly found in birds and cattle (ST6). ST8 has been recognized in marsupials, different species of captive primates and their caretakers. These rarer subtypes in humans have been suggested to be of zoonotic derivation, and there is some confirmation that ST8 has been found in zookeepers of nonhuman primates [15, 59].
Based on next-generation sequencing [60], Blastocystis co-infection using suitable for subtyping (STS) and polymerase chain reaction (PCR) in fecal samples has been established. The mixed subtypes most frequently found are ST1 + ST3, ST1 + ST2, and ST2 + ST3—Figure 4. A case of co-infection by three different subtypes (probably from the same strain) was recently described. This assumption is supported by the polymorphism observed among the 38 sequences belonging to subtype 3 [44].
2.6.2 Prevalence of Blastocystis subtypes according to geographic region
Blastocystis spp. worldwide prevalence is around 25% in humans [45], with a marked superiority in some developing countries with an approximate prevalence of 55–100% [28], while in developed countries, the prevalence reported is 10–15%; this difference has been attributed to hygiene levels and the presence or absence of contact with animals and contaminated water and food [61]. Blastocystis colonizes between one and two million people globally and is one of humans and animals most common intestinal parasites. Humans are primarily colonized by subtypes 1–4, but the relative prevalence of these subtypes appears to differ substantially between regions [23]. According to reports, in Europe, ST1–4 are found more frequently and specifically in the North and South America, since a higher prevalence was found subtypes (ST1–3). The countries with greater variety of these subtypes are the United States, Brazil, and Colombia, with 14, 9 and 8 subtypes, respectively [59].
2.6.3 Differences between subtypes
Currently, the Blastocystis’s subtype is the most accepted approach to explain its pathogenicity Blastocystis [62]. Also, intrasubtypic variations have been observed, thus suggesting that not all strains of a particular subtype are pathogenic and that the subtype does not predict pathogenicity [50].
Subtype 3 (ST3) is one of the most common in humans, as well as the most frequently found in fecal samples from patients with both gastrointestinal and dermatological clinical manifestations (urticaria) [62]. About the pathogenicity of ST4, in a meta-analysis that included studies from Europe, Asia, Africa, and South America this subtype has a much higher global sequence conservation than the others. Furthermore, heat shock proteins (such as 0PHA3 and KOG3047, a ubiquitously expressed prefoldin-like chaperone) and cytosolic Ca2+ ion-dependent cysteine proteases (such as KOG0045, which is a cysteine-like peptidase similar to calpain) were found in the ST4 genomes that were not present in other ST genomes, which may represent virulence factors unique to ST4 [63]. The activities of cysteine proteases in ST4 and ST7 isolates have shown significant variations, which may be one of the reasons for the differences in their virulence [10]. Several studies worldwide have demonstrated that ST5 is derived from animals since it has been found in people who live in rural areas and have close contact with animals. In addition, poor hygiene plays a vital role in contagion [62, 64].
About subtype 7 (ST7), in vitro assays revealed that this subtype caused alterations in the intestinal epithelial barrier by altering binding proteins such as occludin and zonula occludens-1 (ZO-1) [38], which was associated with greater adhesiveness to intestinal epithelial cells and more significant cysteine protease activity than ST4. In the same study, histopathological results showed that mice infected with ST7 had more colon damage and ulceration than control mice [32]. Also, it has shown that ST7 in isolate H (with greater adhesiveness than other isolates) binds preferentially to colonic tissue concerning the cecum and terminal ileum. ST7 has also been associated with more significant colonization of other parasites [60]. Furthermore, comparing sensitivity and resistance to certain antibiotics showed that ST7 has greater resistance to metronidazole than ST4 whereas it has greater sensitivity to emetine than ST4 [61]. In ST7, the 0IZK7 Cystatin B has a potential role in parasitic cysteine protease function and inhibition of host proteases; this protein is also present in ST2 but not in ST1, ST3 and ST4 [62].
2.6.4 Distribution of subtypes in children from different countries
With the advent of new-generation sequencing (Figure 2), since 2020, more Blastocystis subtypes have been identified; for example, in North Thailand the child population in rural areas, which is a vulnerable group to poverty and poor sanitation conditions (Table 1). The authors suggest that the main transmission routes of Blastocystis to humans are soil and water [11]. Contrary to another study in the Thai-Myanmar Border, Ratchaburi Province (Thailand) in the same year, 2023 [69], in which the prevalence of Blastocystis was very low (3.35%). In Thailand, Blastocystis infection rates are 0.7–45.2% in school-age children, and the highest rates are found in young children living in orphan homes in Pathum Thani province; this is consistent with several studies that mention that the prevalence of Blastocystis is higher in developing countries where the prevalence reaches 100% [28]. This is the case in children in Senegal, where open defecation of the population of these regions is attributed to the lack of latrines and water. Conversely, data on the prevalence and distribution of Blastocystis subtypes remain scarce in Africa; however, a multicenter study in 2020 with a total of 731 stool samples collected from healthy children living in 10 villages in the region northwest of Senegal showed significant variation of Blastocystis infections between villages from 51.7 to 100%, the overall prevalence was 80.4% and mixed infections 23%, the possible potential sources of transmission in this population are suggested to be person-person transmission, zoonotic and transmission through contaminated water, also, this is the first report of subtype 10 and 14 in humans [28].
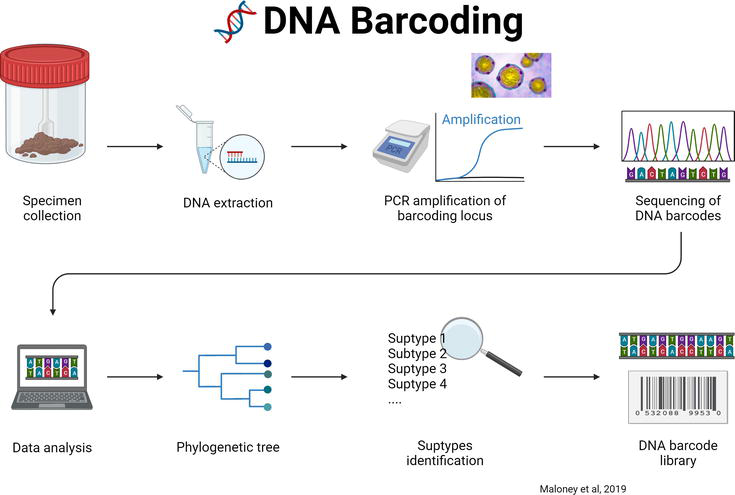
Figure 2.
Next generation sequencing method for the subtyping of Blastocystis. Next-generation sequencing (NGS) in the context of barcoding refers to applying high-capacity DNA sequencing technologies to identify and characterize multiple species, in this case, those of Blastocystis, using genetic barcodes. This involves the massive generation of DNA sequences from multiple samples simultaneously.
Sequence types of Blastocystis found in the populations of children studied.
n number of samples; % of sequenced samples.
In Latin America, in a study carried out in Colombia in a child population that attended daycare centers in Medellín, where the population was urban, the global prevalence of Blastocystis was 36.6% by microscopy and 58.2% by polymerase chain reaction (PCR); this is the first report of subtype 16 identification in humans [13]. In other studies in rural areas of Panama and Mexico, the prevalence of Blastocystis was of 74.2% and 44.0%, respectively. Additionally, in apparently healthy school-aged children in Panama, Blastocystis subtype infection was not associated with gastrointestinal symptoms except for diarrhea in these children [27]. In children from Mexico, the frequency of Blastocystis subtypes was related to ingesting contaminated foods such as sweets, snacks, and artisanal foods at food stalls [24]. As previously described, the child population in developing countries is the most affected by Blastocystis infection. The studies in the child population aim to understand the dynamics of transmission and its consequences for the health of this group. In this regard, the relationship between age and epidemiological and molecular characterization of Blastocystis infection is vital since the lack of information of long-term parasite stability in asymptomatic children makes it difficult to interpret the transmission and pathogenesis (Table 1).
2.6.5 Association of Blastocystis and the gut microbiota
The association of Blastocystis and the fecal microbiota in children has been little studied; recently a study with 57 samples of school-age children in Colombia showed that the composition of the intestinal bacterial community was not different between colonized children and those not colonized by Blastocystis; however, a greater microbial richness was observed in the fecal microbiota of children colonized by Blastocystis, Firmicutes was the most predominant phylum in both groups, and a more significant proportion of Bacteroidetes was found in children not colonized [36].
A previous study in 2016 with subjects aged 9 to 70 years found a higher relative abundance of Bacteroides in Blastocystis-negative samples and a low abundance of Prevotella and a higher abundance of the Clostridial group XIVa in Blastocystis-negative samples. Blastocystis is associated with fecal microbiota characterized by low relative abundances of Bacteroides, Clostridial group XIVa, and high levels of Prevotella [71].
The studies described above aim to understand the dynamics of the transmission of Blastocystis according to subtypes, and implications on health mainly in children. For this reason, it is crucial to characterize Blastocystis infection epidemiologically and molecularly to generate conclusive epidemiological information in developing countries. However, information on stability and epidemiology in asymptomatic children complicates the interpretation of the transmission and pathogenesis of the subtypes of this protozoan. A host can harbor more than two subtypes; however, the extent of subtype diversity within the host is poorly understood. New studies have emerged in which Blastocystis is considered a member of the healthy intestinal microbiota. This disparity is suggested to be due to the high variability between subtypes and host immunity, so it is necessary to know and evaluate the diversity of Blastocystis subtypes; this is crucial to understanding the epidemiology and sources of transmission to humans and, subsequently, depending on the patient’s symptoms, give appropriate treatment. It is crucial to investigate the alterations in the structure of the gut microbiota related to the presence of Blastocystis.
1.Alam A, Neish A. Role of gut microbiota in intestinal wound healing and barrier function. Tissue Barriers. 2018;6:1-21
2.Farré EM. The brown clock: Circadian rhythms in stramenopiles. Physiologia Plantarum. 2020;169:430-441
3.Tan TC, Suresh KG. Predominance of amoeboid forms of Blastocystis hominis in isolates from symptomatic patients. Parasitology Research. 2006;98(3):189-193. Epub ahead of print. DOI: 10.1080/01421590500312847
4.Scanlan PD, Stensvold CR. Blastocystis: Getting to grips with our guileful guest. Trends in Parasitology. 2013;29:523-529
5.El Safadi D, Gaayeb L, Meloni D, et al. Children of Senegal River Basin show the highest prevalence of Blastocystis sp. ever observed worldwide. BMC Infectious Diseases. 2014;14:1-11
6.Tan KSW. New insights on classification, identification, and clinical relevance of Blastocystis spp. Clinical Microbiology Reviews. 2008;21:639-665
7.Zou Y, Bin YW, Zou FC, et al. Molecular detection and subtype distribution of Blastocystis in farmed pigs in southern China. Microbial Pathogenesis. 2021;151:104751
8.Domínguez-Márquez MV, Guna R, Muñoz C, et al. High prevalence of subtype 4 among isolates of Blastocystis hominis from symptomatic patients of a health district of Valencia (Spain). Parasitology Research. 2009;105:949-955
9.Ramírez JD, Sánchez LV, Bautista DC, et al. Blastocystis subtypes detected in humans and animals from Colombia. México: Infection, Genetics and Evolution. 2014;22:223-228
10.Kumarasamy V. Blastocystis sp., parasite associated with gastrointestinal disorders: An overview of its pathogenesis, immune modulation and therapeutic strategies. Current Pharmaceutical Design. 2018;24:3172-3175
11.Jinatham V, Maxamhud S, Popluechai S, et al. Blastocystis one health approach in a rural community of Northern Thailand: Prevalence, subtypes and novel transmission routes. Frontiers in Microbiology. 2021;12:746340. Epub ahead of print. DOI: 10.3389/fmicb.2021.746340
12.Maloney JG, Molokin A, Seguí R, et al. Identification and molecular characterization of four new Blastocystis subtypes designated ST35-ST38. Microorganisms. 2023;11:46. Epub ahead of print. DOI: 10.3390/microorganisms11010046
13.Osorio-Pulgarin MI, Higuera A, Beltran-álzate JC, et al. Epidemiological and molecular characterization of Blastocystis infection in children attending daycare centers in medellín, Colombia. Biology (Basel). 2021;10:669. Epub ahead of print. DOI: 10.3390/biology10070669
14.Alfellani MA. Genetic diversity of Blastocystis in livestock and zoo animals. Protist. 2013;164:497-509
15.Skotarczak B. Genetic diversity and pathogenicity of Blastocystis. Annals of Agricultural and Environmental Medicine. 2018;25:411-416
16.Scanlan PD, Stensvold CR, Rajilić-Stojanović M, et al. The microbial eukaryote Blastocystis is a prevalent and diverse member of the healthy human gut microbiota. FEMS Microbiology Ecology. 2014;90:326-330
17.Lukeš J, Stensvold CR, Jirků-Pomajbíková K, et al. Are human intestinal eukaryotes beneficial or commensals? PLoS Pathogens. 2015;11:e1005039. Epub ahead of print. DOI: 10.1371/journal.ppat.1005039
18.Andersen LO, Stensvold CR. Blastocystis in health and disease: Are we moving from a clinical to a public health perspective? Journal of Clinical Microbiology. 2016;54:524-528
19.Nourrisson C, Scanzi J, Pereira B, et al. Blastocystis is associated with decrease of fecal microbiota protective bacteria: Comparative analysis between patients with irritable bowel syndrome and control subjects. PLoS ONE. 2014;9:1-9
20.Lepczyńska M, Dzika E, Kubiak K, et al. The role of Blastocystis sp. as an etiology of irritable bowel syndrome. Polish Annals of Medicine. 2016;23:57-60
21.Yakoob J, Jafri W, Beg MA, et al. Blastocystis hominis and Dientamoeba fragilis in patients fulfilling irritable bowel syndrome criteria. Parasitology Research. 2010;107:679-684
22.Audebert C, Even G, Cian A, et al. Colonization with the enteric protozoa Blastocystis is associated with increased diversity of human gut bacterial microbiota. Scientific Reports. 2016;6:1-11
23.Seyer A, Karasartova D, Ruh E, et al. Epidemiology and prevalence of Blastocystis spp. in North Cyprus. The American Journal of Tropical Medicine and Hygiene. 2017;96:1164-1170
24.Hidalgo-Gonzalez LA, Salgado-Lopez J, Pineda-Rodriguez SA, et al. Identification of Blastocystis sp. in school children from a rural Mexican village: Subtypes and risk factors analysis. Parasitology Research. 2023;122:1701-1707
25.Casero RD, Mongi F, Sánchez A, et al. Blastocystis and urticaria: Examination of subtypes and morphotypes in an unusual clinical manifestation. Acta Tropica. 2015;148:156-161
26.Ramírez JD, Flórez C, Olivera M, et al. Blastocystis subtyping and its association with intestinal parasites in children from different geographical regions of Colombia. PLoS ONE. 2017;12:1-13
27.Perea M, Vásquez V, Pineda V, et al. Prevalence and subtype distribution of Blastocystis sp. infecting children from a rural community in Panama. Parasite Epidemiology and Control. 2020;9:1-8
28.Khaled S, Gantois N, Ly AT, et al. Prevalence and subtype distribution of Blastocystis sp. in Senegalese school children. Microorganisms. 2020;8:1-17
29.Lepczyńska M, Dzika E. The influence of probiotic bacteria and human gut microorganisms causing opportunistic infections on Blastocystis ST3. Gut Pathogens. 2019;11:1-11
31.Stensvold CR, van der Giezen M. Associations between gut microbiota and common luminal intestinal parasites. Trends in Parasitology. 2018;34:369-377
32.Gentekaki E, Curtis BA, Stairs CW, et al. Extreme genome diversity in the hyper-prevalent parasitic eukaryote Blastocystis. PLoS Biology. 2017;15:1-42
33.Forsell J, Bengtsson-Palme J, Angelin M, et al. The relation between Blastocystis and the intestinal microbiota in Swedish travellers. BMC Microbiology. 2017;17:1-9
34.Kodio A, Coulibaly D, Koné AK, et al. Blastocystis colonization is associated with increased diversity and altered gut bacterial communities in healthy Malian children. Microorganisms. 2019;7:1-11
35.Tito RY, Chaffron S, Caenepeel C, et al. Population-level analysis of Blastocystis subtype prevalence and variation in the human gut microbiota. Gut Microbiota. 2019;68:1180-1189
36.Castañeda S, Muñoz M, Villamizar X, et al. Microbiota characterization in Blastocystis-colonized and Blastocystis-free school-age children from Colombia. Parasit Vectors. 2020;13:521. Epub ahead of print. DOI: 10.1186/s13071-020-04392-9
37.Nieves-Ramírez ME, Partida- Rodríguez O, Laforest-Lapointe I, et al. Asymptomatic intestinal colonization with protist Blastocystis is strongly associated with distinct microbiome ecological patterns. mSystems. 2018;3:1-18
38.Yason JA, Liang YR, Png CW, et al. Interactions between a pathogenic Blastocystis subtype and gut microbiota: In vitro and in vivo studies. Microbiome. 2019;7:1-13
39.Foster JA, Rinaman L, Cryan JF. Stress & the gut-brain axis: Regulation by the microbiome. Neurobiology of Stress. 2017;7:124-136
40.García Peña C, Álvarez Cisneros T, Quiroz Baez R, et al. Microbiota and aging. A review and commentary. Archives of Medical Research. 2017;48:681-689
41.Stensvold CR, Sørland BA, Berg RPKD, et al. Stool microbiota diversity analysis of Blastocystis-positive and Blastocystis-negative individuals. Microorganisms. 2022;10:1-6
42.Deng L, Wojciech L, Gascoigne NRJ, et al. New insights into the interactions between Blastocystis, the gut microbiota, and host immunity. PLoS Pathogens. 2021;17:1-15
43.Yoshikawa H, Nagano I, Wu Z, et al. Genomic polymorphism among Blastocystis hominis strains and development of subtype-specific diagnostic primers. Molecular and Cellular Probes. 1998;12:153-159
44.Poirier P, Meloni D, Nourrisson C, et al. Molecular subtyping of Blastocystis spp. using a new rDNA marker from the mitochondria-like organelle genome. Parasitology. 2014;141:670-681
45.Zhan T, He S, Liu T, et al. A novel genotype screening and phylogenetic analysis of Blastocystis hominis based on EF-1α. International Journal of Clinical and Experimental Pathology. 2017;10:8314-8323
46.Sierra RL, Muñoz SGD, Lora SFM, et al. Identificación de subtipos de Blastocystis sp. asociados a fuentes de transmisión en una zona rural del departamento del Quindío. Revista de la Asociación Colombiana de Ciencias Biológicas. 2023;35:113-127
47.Stensvold CR, Traub RJ, von Samson-Himmelstjerna G, et al. Blastocystis: Subtyping isolates using pyrosequencing™ technology. Experimental Parasitology. 2007;116:111-119
48.Ajjampur SSR, Tan KSW. Pathogenic mechanisms in Blastocystis spp.—Interpreting results from in vitro and in vivo studies. Parasitology International. 2016;65:772-779
49.del Coco VF, Molina NB, Basualdo JA, et al. Blastocystis spp.: Advances, controversies and future challenges. Revista Argentina de Microbiología. 2017;49:110-118
50.Jeremiah S, Parija S. Blastocystis: Taxonomy, biology and virulence. Tropenmedizin und Parasitologie. 2013;3:17
51.Popruk S, Adao DE V, Rivera WL. Epidemiology and subtype distribution of Blastocystis in humans: A review. Infection, Genetics and Evolution. 2021;95:1-14
52.Mohamed RT, El-bali MA, Mohamed AA, et al. Subtyping of Blastocystis sp. isolated from symptomatic and asymptomatic individuals in Makkah, Saudi Arabia. Parasites & Vectors. 2017;10:1-7
53.Basak S, Rajurkar MN, Mallick SK. Detection of Blastocystis hominis: A controversial human pathogen. Parasitology Research. 2014;113:261-265
54.Andersen LOB, Bonde I, Nielsen HBHB, et al. A retrospective metagenomics approach to studying Blastocystis. FEMS Microbiology Ecology. 2015;91:1-9
55.Kurt Ö, Doğruman Al F, Tanyüksel M. Eradication of Blastocystis in humans: Really necessary for all? Parasitology International. 2016;65:797-801
56.Stensvold CR, Clark CG. Current status of Blastocystis: A personal view. Parasitology International. 2016;65:763-771
57.Eme L, Gentekaki E, Curtis B, et al. Lateral gene transfer in the adaptation of the anaerobic parasite Blastocystis to the gut. Current Biology. 2017;27:807-820
58.Chandramathi S, Suresh K, Sivanandam S, et al. Stress exacerbates infectivity and pathogenicity of Blastocystis hominis: In vitro and in vivo evidences. PLoS ONE. 2014;9:1-11
59.Jiménez PA, Jaimes JE, Ramírez JD. A summary of Blastocystis subtypes in North and South America. Parasites & Vectors. 2019;12:1-9
60.Maloney JG, Molokin A, Santin M. Next generation amplicon sequencing improves detection of Blastocystis mixed subtype infections. Infection, Genetics and Evolution. 2019;73:119-125
61.Mohamed AM, Ahmed MA, Ahmed SA, et al. Predominance and association risk of Blastocystis hominis subtype I in colorectal cancer: A case control study. Infectious Agents and Cancer. 2017;12:1-8
62.Cakir F, Cicek M, Yildirim IH. Determination the subtypes of Blastocystis sp. and evaluate the effect of these subtypes on pathogenicity. Acta Parasitologica. 2019;64:7-12
63.Beghini F, Pasolli E, Truong TD, et al. Large-scale comparative metagenomics of Blastocystis, a common member of the human gut microbiome. ISME Journal. 2017;11:2848-2863
64.Yañez CM, Hernández AM, Sandoval AM, et al. Prevalence of Blastocystis and its association with Firmicutes/Bacteroidetes ratio in clinically healthy and metabolically ill subjects. BMC Microbiology. 2021;21:1-11
65.Sari IP, Benung MR, Wahdini S, et al. Diagnosis and identification of Blastocystis subtypes in primary school children in Jakarta. Journal of Tropical Pediatrics. 2018;64:208-214
66.Cinek O, Polackova K, Odeh R, et al. Blastocystis in the faeces of children from six distant countries: Prevalence, quantity, subtypes and the relation to the gut bacteriome. Parasites & Vectors. 2021;14:1-16
67.Semra Güreser A, Comba A, Karasartova D, et al. Detection of Blastocystis subtypes in children with functional abdominal pain and celiac disease in Çorum, Turkey. Iranian Journal of Parasitology. 2021;17:296-305
68.Abu A, Sutthikornchai C, Mahittikorn A, et al. Prevalence and subtype distribution of Blastocystis isolated from school-aged children in the Thai-Myanmar Border, Ratchaburi Province, Thailand. International Journal of Environmental Research and Public Health. 2022;20:1-9
69.McCain A, Gruneck L, Popluechai S, et al. Circulation and colonisation of Blastocystis subtypes in schoolchildren of various ethnicities in rural northern Thailand. Epidemiology and Infection. 2023;151:e85. Epub ahead of print. DOI: 10.1017/S0950268823000596
70.Tapia-Veloz E, Gozalbo M, Guillén M, et al. Prevalence and associated risk factors of intestinal parasites among schoolchildren in Ecuador, with emphasis on the molecular diversity of Giardia duodenalis, Blastocystis sp. and Enterocytozoon bieneusi. PLoS Neglected Tropical Diseases. 2023;17:1-21
71.O’Brien Andersen L, Karim AB, Roager HM, et al. Associations between common intestinal parasites and bacteria in humans as revealed by qPCR. European Journal of Clinical Microbiology and Infectious Diseases. 2016;35:1427-1431
Written By
Janeth Oliva Guangorena-Gómez and Claudia Muñoz Yañez
Submitted: 13 February 2024Reviewed: 19 February 2024Published: 10 April 2024