
Abstract
Salmonella belonging to the Enterobacteriaceae family is commonly divided into Typhoidal and non-typhoidal group. Clinical manifestations can range from gastroenteritis, bacteremia or septicemia without gastrointestinal (GI) upset, enteric fever, chronic carriage to focal infections like osteomyelitis, meningitis, endocarditis etc. Conventional diagnostic tests like blood, stool, food sample culture have a long turnaround time. Chromogenic media modification reduces identification time. Apart from Widal test and ELISA, rapid point of care serological tests like Tubex TF, Typhidot are advantageous in basic set up but limited by modest accuracy in high burden settings. Apart from immunological assays, other diagnostic modalities include PCR, mass spectrophotometry, spectroscopy, optical phenotyping, biosensors etc. Rising drug resistance of Salmonella to first line Ampicillin, Chloramphenicol and Cotrimoxazole with increasing nonsusceptibility to fluoroquinolones and 3rd generation cephalosporin for past few decades is a public threat. Prevention and control measures include basic sanitation, safe water access, safe food handling, public education, physical, chemical, biocontrol methods, vaccination etc. Targeting the menace of antimicrobial resistance in Salmonella species needs a collaborative effort like One Health approach which optimizes the public health, animal and environmental health and reduce the dependency on antibiotics.
Keywords
- Salmonella
- typhoid
- paratyphoid
- non-typhoidal
- diagnosis
- drug resistance: prevention: control
1. Introduction
The bacterium
2. Taxonomy and nomenclature
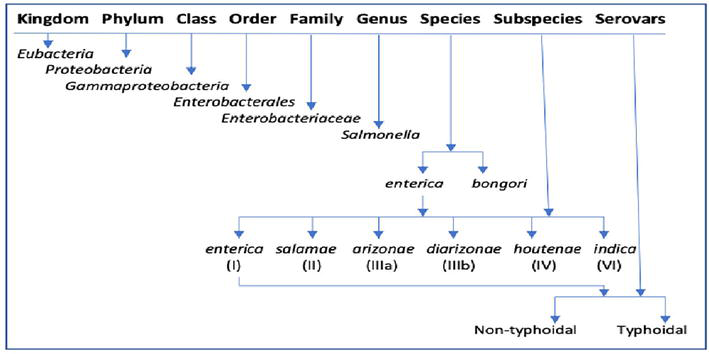
Figure 1.
Apart from this, further classification of
Antigenic formula usually begins with subspecies written in Roman letters followed by somatic O antigens, flagellar H antigens phase 1 and phase 2, followed by other results if present. A colon separates major antigens and comma separates components of one antigen. Various symbols appearing in the antigenic formula make it complicated for example, square brackets like [ ] means the O or H antigen in question may be present or absent, ( ) parentheses means antigen is weakly agglutinable, { } curly bracket means antigens are mutually exclusive, hyphen – means particular flagellar phase is absent and underlined O factor means the factor is coded by bacteriophage (lysogenic strain). For example, antigenic formulae for the
3. Disease transmission
Salmonellosis is mostly foodborne. Consumption of contaminated meat, dairy and eggs is the cause as the bacteria mostly resides in the gut of animals and birds, the major sources being pigs, cattle and poultry. Transmission through the consumption of contaminated produce like fruits, vegetables, dried seeds as well as cereals has also been documented [11]. Person-to-person spread occurs in enteric fever via ingestion of food and water contaminated with human feces as Typhoidal fever lacks animal reservoirs. Food handlers especially, asymptomatic carriers of the bacteria with poor hygiene, sanitation and lack of clean water contribute to the spread of the disease [12]. The history of Mary Mallon “Typhoid Mary” is synonymous with the spread of the disease. The Irish-born American cook was a healthy carrier who was found to be responsible for the disease in hundreds of people [13]. The infective dose of bacteria has been reported to be less in non-typhoidal disease to the tune of 103 whereas approximately 105 bacilli is the infective dose for enteric fever [14, 15]. Various factors exist which increase the susceptibility of the host to Salmonellosis. Children less than 5 years of age, elderly and immunocompromised cases are more susceptible. Hosts with conditions like achlorhydria, gastric surgery, inflammatory bowel disease, hemoglobinopathies, and impaired immune conditions like diabetes mellitus, AIDS, leukemia and lymphomas are more likely to get infected with a lower infective dose [14, 15, 16, 17].
4. Pathogenesis and virulence factors
The bacilli colonize the ileum and colon by attaching to the epithelial cells (M cells) by fimbriae. They enter through its epithelial lining by interacting with apical microvilli. Bacteria-mediated endocytosis occurs when the bacilli trigger the formation of ruffles on the cell membrane of M cells leading to the engulfment of the bacteria in vesicles. These vacuoles containing
5. Clinical features
Infections caused by
5.1 Enteric fever
Enteric fever is caused by
5.2 Non-typhoidal gastroenteritis
Non-typhoidal
5.3 Invasive non-typhoidal Salmonella (iNTS) infections
The characteristic of iNTS infections is bacteremia by NTS without any focus of infection or gastroenteritis [35]. The most common iNTS strains are Choleraesuis, Dublin, Enteritidis, Typhimurium, Bovismorbificans, Arizona, and Gallinarum etc. Risk factors for bacteremia with other extra-intestinal manifestations include malnutrition, haemolytic anemia (Sickle cell anemia, Thalassemia), T-cell immunosuppression (AIDS, Hodgkin Lymphoma, Steroid Use, Transplantation, Chemotherapy), Schistosomiasis, Chronic granulomatous disease, mutations or acquired auto-immunity compromising IL-12, IFNγ function etc. [36]. The virulence determinants of iNTS are reported to be SPI-1 & SPI-2 responsible for an invasion via T3SS expression, survival of bacteria in macrophages and systemic manifestations;
5.4 Carrier state
Carriers are important from the public health point of view as they are the major cause of outbreaks. Temporary carriers are the cases who continue to shed Typhoid bacteria up to 3 months after disease resolution. It is seen in approximately 10% of cases. In chronic carriers, approximately 2–5% of cases fail to fully clear the infection even after a year of resolution of the disease [38]. Chronic carriers act as a reservoir of infection in the community as they intermittently shed the bacteria in stool or urine. Long-term survival of bacteria in carriers is due to the breach of the epithelial barrier, evasion of the host immune system and the forming of a niche in the host body. Mechanical barrier due to the sphincter of Oddi along with the flushing inability of bacteriostatic bile by the host is associated with the long-term carriage of the pathogen [39]. In contrast to Typhoidal
6. Epidemiology
World Health Organization reported Typhoid infections affecting approximately 9 million people with over 110,000 deaths as of 2019. Typhoid was mostly reported from the developing nations of Southeast Asia, Africa, the Eastern Mediterranean and the Western Pacific Region [2]. The European countries and USA reported a low number of cases approximately less than 10 per 100,000 population annually with mostly travel-related history to enteric fever endemic countries [40]. Low-incidence countries like Israel noted causative agents of enteric fever to be
Non-typhoidal
7. Detection
7.1 Culture based techniques
Even though culture-based methods are time-consuming, it is still the gold standard in diagnosis. Stools and suspected food products are the most commonly tested clinical samples. Apart from stool, other clinical samples are blood, body fluids, pus, bone marrow, duodenal aspirate, urine, punch biopsy samples of rose spots etc. [51]. The two most commonly done diagnostic tests for enteric fever are culture and Widal test. Bacterial culture depends on the site, age group, antimicrobial intake and stage of illness. Though blood culture is done commonly, the sensitivity is lower and variable (40–80%) as compared to bone marrow culture with a higher sensitivity of more than 80%. Also, blood culture sensitivity is higher in the first week as compared to subsequent weeks of infection and also gets significantly affected with antibiotics intake unlike bone marrow culture [52]. Blood and sterile body fluids can be cultured in blood culture bottles which can be conventional blood culture or automated blood culture bottles (Bactec/BacTAlert) which are further subcultured onto MacConkey agar. Culture methodology is different for contaminated samples like stool or food samples. The fecal samples on day 0 are cultured onto selective media like Xylose Lysine Deoxycholate (XLD) along with an enrichment broth like Selenite F Broth and incubated at 37°C for 18–24 hours. The enrichment broth is further subcultured onto selective media on Day 1. Enrichment broths like Selenite cystine broth, Gram Negative broth, and Tetrathionate broth are required for the selective enrichment of
7.2 Non-culture-based techniques
Immunological assays: The Widal test is a routinely used serological test in high-burden countries. It is an agglutination test quantifying antibodies against lipopolysaccharide O and flagellar H antigen. Though very convenient for low resource settings for its simplicity, the pitfall of this test primarily lies in the need for cautious interpretation of the results, limited sensitivity, specificity and difficulty distinguishing current and past infections [54]. Single unpaired tests showed poor sensitivity and specificity, especially in the early course of infection. Paired serological testing improves diagnostic accuracy but is limited in its clinical utility [51, 54]. Unlike the conventional Widal agglutination test, newer techniques in the immunological assay are based on the principle of
Molecular Assays: Molecular techniques broadly use the principle of DNA/RNA detection of specific pathogens by target or signal amplification or non-amplification methods [49]. PCR is an excellent tool for pathogen detection. The most commonly used PCR targets are flagellum genes like f
Mass Spectrometry-Based and Spectroscopy Method
Optical phenotyping methods like BARDOT (Bacterial Rapid Detection using Optical Scattering Technology) and BISLD (Bacteria Identification System by Light Diffraction) are available. The principle of the light scattering phenomenon is used where scattering images of colonies are compared with reference image libraries [69]. SELA-BARDOT technology uses on plate screening technique with laser which generates individual scatter signatures of multiple pathogens grown on SEL agar and provides results in real time [70]. This optical phenotyping method is non-destructive, has less turnaround time and is economical due to no requirement for any probes or reagents [70]. BISLD (Bacteria Identification System by Light Diffraction) is an automated system for bacterial colony identification. Converging spherical waves on the colonies obtain Fresnel diffraction patterns along with 2-D transmission coefficient maps in order to identify multiple species of bacteria [71].
Biosensors: Biosensor is a compact analytical device which provides information on analytes with the help of a biological recognition element and a transducer [72]. The advantages of biosensors over conventional diagnostic techniques are their higher sensitivity, specificity, portability potential and rapid results [49, 72, 73]. The three main components of a biosensor are a bio receptor, a transducer and an electronic system. The role of the electronic system component in biosensor devices is signal amplification, processing data and displaying the output [49]. Bio receptors are the analytes which recognize the target and the commonly used bioreceptors in
8. Antimicrobial treatment and drug resistance
Enteric fever was conventionally treated by first-line antibiotics consisting of ampicillin, chloramphenicol and cotrimoxazole. Strains resistant to these first-line antibiotics were termed as Multi-Drug Resistant (MDR)
With vaccine coverage,
AMR is rising in NTS due to the heavy use of antibiotics in animal husbandry. Clinically NTS diarrhea does not need antibiotics unless invasive in nature, elderly, children or immunocompromised case. AMR rates jumped from 20% in the 1990s to 70% in high-income countries by the 2000s according to surveillance data [93]. Of the prevalent serotypes,
AMR in
9. Prevention and control
Prevention and control of
Effective food biosecurity measures to prevent contamination by
Physical interventions for control of Salmonella with regards to food safety that have been evaluated alone or in combination with other methods include ultrasound, UV, high-pressure processing, irradiation, pulsed light, plasma, thermal processing, high-intensity pulsed electric fields, oscillating magnetic fields etc. [119, 120, 121]. Chemical decontamination methods include the use of biocides like chlorine-based chemicals, chlorine alternatives (e.g. organic acids, peracetic acid), gas treatments (e.g. ozone, chlorine dioxide) etc. [120, 121, 122]. A biocide is a formulation containing an active substance to destroy or control any harmful organism to human or animal health by means other than physical or mechanical action [123]. Biocides for disinfection purposes can be divided into oxidizing (e.g. chlorine, chlorine dioxide, iodine, peroxides, peracetic acid etc.) and non-oxidizing groups (e.g. quaternary ammonium compounds, amphoterics, aldehydes, phenolic compounds, biguanides, and acid anionic agents etc) [122, 124]. However, there are greater implications regarding the widespread use of antimicrobial biocide like the development of biocide resistance, dermal or respiratory exposure effects, environmental threat due to persistence in aquatic systems etc. [122].
One of the novel biocontrol measures includes the use of bacteriophages (phages). Bacteriophages are viruses that infect and replicate in bacterial cells to create new progeny by using the host machinery [125]. The potential of phages against the rise of antibiotic-resistant
Phytobiotics are plant extracts like herbs and spices which prevent food spoilage from contaminants, increase food shelf life and improve fortified food quality. In animals, phytobiotics owing to their antibacterial effects and microbiota modulation are known to improve carcass quality, decrease pathogenic bacteria proliferation, stimulate the immune system etc. [131, 132]. Bioactive compounds in phytobiotics also alter bacterial cytoplasmic membranes and genetic machinery, disrupt iron uptake pathways etc. [133]. Some examples of herbs and spices used against
Prebiotics, probiotics, synbiotics and postbiotics help prevent gut colonization by
Another alternative method that can target bacteria is nanoparticles. Nanoparticles are ultrafine colloidal particles ranging from 1 to 100 nm in diameter [142]. The antibacterial mechanisms of nanoparticles are still unclear but the current theories are induction of oxidative stress, metal ion release and non-oxidative pathways [142]. Various types of nanoparticles against the control of
Tackling drug-resistant
10. Conclusion
The disease burden caused by
Alternatives to antibiotics are the need of the hour to combat rising antimicrobial resistance in
It is of vital importance to commit to research and development so that efficient and advanced methods can quickly detect antibiotic-resistant
Conflict of interest
The authors declare no conflict of interest.
References
- 1.
World Health Organization (WHO). Salmonella (Non-Typhoidal) Factsheets. 2018. Available from: https://www.who.int/news-room/fact-sheets/detail/salmonella-(non-typhoidal) [Accessed: December 28, 2023] - 2.
World Health Organization (WHO). Salmonella Typhoid Factsheets. 2023. Available from: https://www.who.int/news-room/fact-sheets/detail/typhoid#:~:text=Typhoid%20fever%20is%20a%20life,and%20spread%20into%20the%20bloodstream [Accessed: December 28, 2023] - 3.
Ryan MP, O'Dwyer J, Adley CC. Evaluation of the complex nomenclature of the clinically and veterinary significant pathogen Salmonella . BioMed Research International. 2017;2017 :3782182 - 4.
Hardy A. Food, hygiene, and the laboratory. A short history of food poisoning in Britain, circa 1850-1950. Social History of Medicine. 1999; 12 (2):293-311 - 5.
MacKenzie KD, Palmer MB, Köster WL, White AP. Examining the link between biofilm formation and the ability of pathogenic Salmonella strains to colonize multiple host species. Frontiers in Veterinary Science. 2017;4 :138 - 6.
Popoff MY, Bockemühl J, Gheesling LL. Supplement 2001 (no. 45) to the Kauffmann–White scheme. Research in Microbiology. 2003; 154 (3):173-174 - 7.
Brenner FW, Villar RG, Angulo FJ, Tauxe R, Swaminathan B. Salmonella nomenclature. Journal of Clinical Microbiology. 2000;38 :2465-2467 - 8.
Issenhuth-Jeanjean S, Roggentin P, Mikoleit M, Guibourdenche M, De Pinna E, Nair S, et al. Supplement 2008-2010 (no. 48) to the White–Kauffmann–Le Minor scheme. Research in Microbiology. 2014; 165 :526-530 - 9.
Grimont PA, Weill F-X. Antigenic formulae of the Salmonella serovars. 9th Ed. Paris, France: WHO Collaborating Centre for Reference and Research onSalmonella ; 2007 - 10.
Trüper HG. Judicial Commission Of The International Committee On Systematics Of Prokaryotes. The type species of the genus Salmonella Lignieres 1900 is Salmonella enterica (ex Kauffmann and Edwards 1952) Le Minor and Popoff 1987, with the type strain LT2T, and conservation of the epithet enterica in Salmonella enterica over all earlier epithets that may be applied to this species. Opinion 80. International Journal of Systematic and Evolutionary Microbiol. Jan 2005; 55 (1):519-520 - 11.
Ehuwa O, Jaiswal AK, Jaiswal S. Salmonella , food safety and food handling practices. Food. 2021;10 (5):907 - 12.
Kirchhelle C, Pollard AJ, Vanderslott S. Typhoid-from past to future. Clinical Infectious Diseases [Internet]. 2019; 69 (Suppl 5):S375-S376 - 13.
Marineli F, Tsoucalas G, Karamanou M, Androutsos G. Mary Mallon (1869-1938) and the history of typhoid fever. Annals of Gastroenterology. 2013; 26 (2):132-134 - 14.
Bronze MS, Greenfield RA, editors. Biodefence Principles and Pathogens. Greenfield Norfolk, United Kingdom: Horizon Bioscience; 2005 - 15.
Ryan KJ, Ray CG, editors. Sherris Medical Microbiology: An Introduction to Infectious Disease. 4th ed. New York: McGraw-Hill; 2004 - 16.
Giannella RA. Salmonella. In: Baron S, editor. Medical Microbiology. 4th ed. Galveston (TX): University of Texas Medical Branch at Galveston; 1996. p. 21 - 17.
Pucciarelli MG, García-Del Portillo F. Salmonella intracellular lifestyles and their impact on host-to-host transmission. Microbiology Spectrum. 2017; 5 (4):10.1128 - 18.
Sabbagh SC, Forest CG, Lepage C, Leclerc JM, Daigle F. So similar, yet so different: Uncovering distinctive features in the genomes of Salmonella enterica serovars Typhimurium and Typhi. FEMS Microbiology Letters. 2010;305 (1):1-13 - 19.
Daigle F. Typhi genes expressed during infection or involved in pathogenesis. Journal of Infection in Developing Countries. 2008; 2 (6):431-437 - 20.
Smith JL. The role of gastric acid in preventing foodborne disease and how bacteria overcome acid conditions. Journal of Food Protection. 2003; 66 :1292-1303 - 21.
Marcus SL, Brumell JH, Pfeifer CG, Finlay BB. Salmonella pathogenicity islands: Big virulence in small packages. Microbes and Infection. 2000;2 (2):145-156 - 22.
Amavisit P, Lightfoot D, Browning GF, Markham PF. Variation between pathogenic serovars within Salmonella pathogenicity islands. Journal of Bacteriology. 2003;185 (12):3624-3635 - 23.
Van Asten AJA, Van Dijk JE. Distribution of “classic” virulence factors among Salmonella spp. FEMS Immunology and Medical Microbiology. 2005;44 (3):251-259 - 24.
Monack DM, Mueller A, Falkow S. Persistent bacterial infections: The interface of the pathogen and the host immune system. Nature Reviews Microbiology. 2004; 2 :747-765 - 25.
Guiney DG, Fierer J. The role of the spv genes in Salmonella pathogenesis. Frontiers in Microbiology. 2011;2 :129 - 26.
Röder J, Hensel M. Presence of SopE and mode of infection result in increased Salmonella -containing vacuole damage and cytosolic release during host cell infection bySalmonella enterica. Cellular Microbiology. 2020;22 :e13155 - 27.
Eng S-K, Pusparajah P, AbMutalib N-S, Ser H-L, Chan K-G, Lee L-H. Salmonella : A review on pathogenesis, epidemiology and antibiotic resistance. Frontiers in Life Science. 2015;8 (3):284-293 - 28.
Eckmann L, Kagnoff MF. Cytokines in host defense against Salmonella . Microbes and Infection. 2001;3 (14-15):1191-1200 - 29.
Ahmad Bhat K, Manzoor T, Ahmad Dar M, Farooq A, Ahmad Allie K, Majeed Wani S, et al. Salmonella infection and pathogenesis [internet]. In: Enterobacteria. London, UK: IntechOpen; 2022 - 30.
Darby J, Sheorey H. Searching for Salmonella . Australian Family Physician. 2008;37 (10):806-810 - 31.
Centre for Disease Control and Prevention. Typhoid and Paratyphoid Fever. 2023. Available from: https://www.cdc.gov/typhoid-fever/health-professional.html#:~:text=Confusion%2C%20delirium%2C%20and%20intestinal%20perforation,10%20days%20for%20paratyphoid%20fever [Accessed: January 02, 2024] - 32.
Parry CM, Hien TT, Dougan G, White NJ, Farrar JJ. Typhoid fever. The New England Journal of Medicine. 2002; 347 :1770-1782 - 33.
Kuvandik C, Karaoglan I, Namiduru M, Baydar. Predictive value of clinical and laboratory findings in the diagnosis of the enteric fever. The New Microbiologica. 2009; 32 :25-30 - 34.
Hohmann EL. Nontyphoidal salmonellosis. Clinical Infectious Disease. 2001; 15 (32):263-269 - 35.
Saphra I, Winter JW. Clinical manifestations of salmonellosis in man; an evaluation of 7779 human infections identified at the New York Salmonella Center. The New England Journal of Medicine. 1957; 256 (24):1128-1134 - 36.
Fierer J. Invasive non-typhoidal Salmonella (iNTS) infections. Clinical Infectious Diseases. 2022; 75 (4):732-738 - 37.
Agbor TA, McCormick BA. Salmonella effectors: Important players modulating host cell function during infection. Cellular Microbiology. 2011; 13 (12):1858-1869 - 38.
Gunn JS, Marshall JM, Baker S, Dongol S, Charles RC, Ryan ET. Salmonella chronic carriage: Epidemiology, diagnosis, and gallbladder persistence. Trends in Microbiology. 2014; 22 (11):648-655 - 39.
Gonzalez-Escobedo G et al. Chronic and acute infection of the gall bladder by Salmonella Typhi: Understanding the carrier state. Nature Reviews Microbiology. 2011;9 :9-14 - 40.
Cooke FJ, Day M, Wain J, Ward LR, Threlfall EJ. Cases of typhoid fever imported into England, Scotland and Wales (2000-2003). Transactions of the Royal Society of Tropical Medicine and Hygiene. 2007; 101 :398-404 - 41.
Meltzer E, Yossepowitch O, Sadik C, Dan M, Schwartz E. Epidemiology and clinical aspects of enteric fever in Israel. The American Journal of Tropical Medicine and Hygiene. 2006; 74 :540-545 - 42.
Wain J, Hendriksen RS, Mikoleit ML, Keddy KH, Ochiai RL. Typhoid fever. Lancet. 2015; 385 (9973):1136-1145 - 43.
Crump JA. Progress in typhoid fever epidemiology. Clinical Infectious Diseases. 2019; 68 (Suppl 1):S4-S9 - 44.
Breiman RF, Cosmas L, Njuguna H, Audi A, Olack B, Ochieng JB, et al. Population-based incidence of typhoid fever in an urban informal settlement and a rural area in Kenya: Implications for typhoid vaccine use in Africa. PLoS One. 2012; 7 (1):e29119 - 45.
Gao XY, Tang QY, Liu FF, Song Y, Zhang ZJ, Chang ZR. Epidemiological characteristics of typhoid fever and paratyphoid fever in China, 2004-2020. Zhonghua Liu Xing Bing Xue Za Zhi. 2023; 44 (5):743-750 - 46.
Balasubramanian R, Im J, Lee JS, Jeon HJ, Mogeni OD, Kim JH, et al. The global burden and epidemiology of invasive non-typhoidal Salmonella infections. Human Vaccines & Immunotherapeutics. 2019;15 (6):1421-1426 - 47.
Ao TT, Daugla DM, Toralta J, Ngadoua C, Fermon F, Page A-L, et al. Global burden of invasive nontyphoidal Salmonella disease. 2010(1). Emerging Infectious Diseases. 2015; 21 (6):941-949 - 48.
Galanis E, Lo Fo Wong DM, Patrick ME, Binsztein N, Cieslik A, et al. Web-based surveillance and global Salmonella distribution, 2000-2002. Emerging Infectious Diseases. 2006; 12 :381-388 - 49.
Awang MS, Bustami Y, Hamzah HH, Zambry NS, Najib MA, Khalid MF, et al. Advancement in Salmonella detection methods: From conventional to electrochemical-based sensing detection. Biosensors (Basel). 2021;11 (9):346 - 50.
Wang M, Zhang Y, Tian F, Liu X, Du S, Ren G. Overview of rapid detection methods for Salmonella in foods: Progress and challenges. Food. 2021;10 (10):2402 - 51.
Andrews JR, Ryan ET. Diagnostics for invasive Salmonella infections: Current challenges and future directions. Vaccine. 2015; 33 :C8-C15 - 52.
Gilman RH, Terminel M, Levine MM, Hernandez-Mendoza P, Hornick RB. Relative efficacy of blood, urine, rectal swab, bone-marrow, and rose-spot cultures for recovery of Salmonella Typhi in typhoid fever. Lancet. 1975; 1 :1211-1213 - 53.
Carroll KC, Pfaller MA, Landry ML, McAdam AJ, Patel R, Richter SS, et al., editors. Manual of Clinical Microbiology. 12th ed. Washington, D.C.: ASM Press; 2019 - 54.
Dutta S, Sur D, Manna B, Sen B, Deb AK, et al. Evaluation of new-generation serologic tests for the diagnosis of typhoid fever: Data from a community-based surveillance in Calcutta, India. Diagnostic Microbiology and Infectious Disease. 2006; 56 :359-365 - 55.
Procop GW, Koneman EW. Koneman’s Color Atlas and Textbook of Diagnostic Microbiology. 7th ed. Baltimore, MD: Wolters Kluwer; 2016 - 56.
Adhikari A, Rauniyar R, Raut PP, Manandhar KD, Gupta BP. Evaluation of sensitivity and specificity of ELISA against Widal test for typhoid diagnosis in endemic population of Kathmandu. BMC Infectious Diseases. 2015; 15 :523 - 57.
Fadeel MA, House BL, Wasfy MM, Klena JD, Habashy EE, Said MM, et al. Evaluation of a newly developed ELISA against Widal, TUBEX-TF and Typhidot for typhoid fever surveillance. Journal of Infection in Developing Countries. 2011; 5 (3):169-175 - 58.
House D, Chinh NT, Diep TS, Parry CM, Wain J, Dougan G, et al. Use of paired serum samples for serodiagnosis of typhoid fever. Journal of Clinical Microbiology. 2005; 43 (9):4889-4890 - 59.
Thiha A, Ibrahim F. A colorimetric enzyme-linked immunosorbent assay (ELISA) detection platform for a point-of-care dengue detection system on a lab-on-compact-disc. Sensors (Basel). 2015; 15 (5):11431-11441 - 60.
De La Rica R, Stevens MM. Plasmonic ELISA for the ultrasensitive detection of disease biomarkers with the naked eye. Nature Nanotechnology. 2012; 7 :821-824 - 61.
Thorns CJ, McLaren IM, Sojka MG. The use of latex particle agglutination to specifically detect Salmonella Enteritidis. International Journal of Food Microbiology. 1994; 21 :47-53 - 62.
Thriemer K, Ley B, Menten J, Jacobs J, van den Ende J. A systematic review and meta-analysis of the performance of two point of care typhoid fever tests, Tubex TF and Typhidot, in endemic countries. PLoS One. 2013; 8 :e81263 - 63.
Beig FK, Ahmad F, Ekram M, Shukla I. Typhidot M and Diazo test vis-à-vis blood culture and Widal test in the early diagnosis of typhoid fever in children in a resource poor setting. The Brazilian Journal of Infectious Diseases. 2010; 14 (6):589-593 - 64.
Najib MA, Mustaffa KMF, Ong EBB, Selvam K, Khalid MF, Awang MS, et al. Performance of immunodiagnostic tests for typhoid fever: A systematic review and meta-analysis. Pathogens. 2021; 10 (9):1184 - 65.
Ricke SC, Kim SA, Shi Z, Park SH. Molecular-based identification and detection of Salmonella in food production systems: Current perspectives. Journal of Applied Microbiology. 2018; 125 (2):313-327 - 66.
Bell RL, Jarvis KG, Ottesen AR, McFarland MA, Brown EW. Recent and emerging innovations in Salmonella detection: A food and environmental perspective. Microbial Biotechnology. 2016; 9 :279-292 - 67.
Pandian S, Lakshmi SA, Priya A, Balasubramaniam B, Zaukuu J-LZ, Durgadevi R, et al. Spectroscopic methods for the detection of microbial pathogens and diagnostics of infectious diseases—An updated overview. PRO. 2023; 11 (4):1191 - 68.
Sun J, Xu X, Feng S, Zhang H, Xu L, Jiang H, et al. Rapid identification of Salmonella serovars by using Raman spectroscopy and machine learning algorithm. Talanta. 2023; 253 :123807 - 69.
Singh AK, Bettasso AM, Bae E, Rajwa B, Dundar MM, Forster MD, et al. Laser optical sensor, a label-free on-plate Salmonella enterica colony detection tool. MBio. 2014; 5 :e01019-e01013 - 70.
Abdelhaseib MU, Singh AK, Bhunia AK. Simultaneous detection of Salmonella enterica, Escherichia coli and Listeria monocytogenes in food using a light scattering sensor. Journal of Applied Microbiology. 2019; 126 (5):1496-1507 - 71.
Suchwalko A, Buzalewicz I, Wieliczko A, Podbielska H. Bacteria species identification by the statistical analysis of bacterial colonies Fresnel patterns. Optics Express. 2013; 21 (9):11322-11337 - 72.
Kirsch J, Siltanen C, Zhou Q , Revzin A, Simonian A. Biosensor technology: Recent advances in threat agent detection and medicine. Chemical Society Reviews. 2013; 42 (22):8733-8768 - 73.
Shen Y, Xu L, Li Y. Biosensors for rapid detection of Salmonella in food: A review. Comprehensive Reviews in Food Science and Food Safety. 2021; 20 (1):149-197 - 74.
Park KS. Nucleic acid aptamer-based methods for diagnosis of infections. Biosensors & Bioelectronics. 2017; 102 :179-188 - 75.
Silva NFD, Magalhães JMCS, Freire C, Delerue-Matos C. Electrochemical biosensors for Salmonella: State of the art and challenges in food safety assessment. Biosensors and Bioelectronics. 2018; 99 :667-682 - 76.
Khansili N, Rattu G, Krishna PM. Label-free optical biosensors for food and biological sensor applications. Sensors and Actuators B: Chemical. 2018; 265 :35-49 - 77.
Wang L, Wang R, Chen F, Jiang T, Wang H, Slavik M, et al. QCM-based aptamer selection and detection of Salmonella Typhimurium. Food Chemistry. 2017; 221 :776-782 - 78.
Yue X, Sun J, Yang T, Dong Q , Li T, Ding S, et al. Rapid detection of Salmonella in milk by a nuclear magnetic resonance biosensor based on the streptavidin-biotin system and O-carboxymethyl chitosan target gadolinium probe. Journal of Dairy Science. 2021; 104 (11):11486-11498 - 79.
Lu Y, Shi Z, Liu Q. Smartphone-based biosensors for portable food evaluation. Current Opinion in Food Science. 2019; 28 :74-81 - 80.
Riu J, Giussani B. Electrochemical biosensors for the detection of pathogenic bacteria in food. TrAC Trends in Analytical Chemistry. 2020; 126 :115863 - 81.
Britto CD, Wong VK, Dougan G, Pollard AJ. A systematic review of antimicrobial resistance in Salmonella enterica serovar Typhi, the etiological agent of typhoid. PLoS Neglected Tropical Diseases. 2018;12 :e0006779 - 82.
Levine MM, Simon R. The gathering storm: Is untreatable typhoid fever on the way? MBio. 2018; 9 (2):e00482-18 - 83.
Hirose K, Hashimoto A, Tamura K, et al. DNA sequence analysis of DNA gyrase and DNA topoisomerase IV quinolone resistance-determining regions of Salmonella enterica serovar Typhi and serovar Paratyphi A. Antimicrobial Agents and Chemotherapy. 2002;46 :3249-3252 - 84.
Dahiya S, Sharma P, Kumari B, Pandey S, Malik R, Manral N, et al. Characterisation of antimicrobial resistance in Salmonellae during 2014-2015 from four centres across India: An ICMR antimicrobial resistance surveillance network report. Indian Journal of Medical Microbiology. 2017;35 :61-68 - 85.
Saha SK, Talukder SY, Islam M, Saha S. A highly ceftriaxone-resistant Salmonella Typhi in Bangladesh. The Pediatric Infectious Disease Journal. 1999;18 :387 - 86.
Ali Shah SA, Nadeem M, Syed SA, Fatima Abidi ST, Khan N, Bano N. Antimicrobial sensitivity pattern of Salmonella Typhi: Emergence of resistant strains. Cureus. 2020; 12 (11):e11778 - 87.
Chatham-Stephens K, Medalla F, Hughes M, et al. Emergence of extensively drug-resistant Salmonella Typhi infections among travelers to or from Pakistan-United States, 2016-2018. Morbidity and Mortality Weekly Report. 2019; 68 :11-13 - 88.
Hooda Y, Sajib MSI, Rahman H, et al. Molecular mechanism of azithromycin resistance among typhoidal Salmonella strains in Bangladesh identified through passive pediatric surveillance. PLoS Neglected Tropical Diseases. 2019;13 :e0007868 - 89.
Rowe B, Ward LR, Threlfall EJ. Multidrug-resistant Salmonella Typhi: A worldwide epidemic. Clinical Infectious Diseases. 1997; 24 :S106-S109 - 90.
Qamar FN, Yousafzai MT, Dehraj IF, Shakoor S, Irfan S, Hotwani A, et al. Antimicrobial resistance in Typhoidal Salmonella: Surveillance for enteric fever in Asia project, 2016-2019. Clinical Infectious Diseases. 2020; 71 (Suppl 3):S276-S284 - 91.
Hasan R, Zafar A, Abbas Z, Mahraj V, Malik F, Zaidi A. Antibiotic resistance among Salmonella enterica serovars Typhi and Paratyphi A in Pakistan (2001-2006). Journal of Infection in Developing Countries. 2008; 2940 :289-294 - 92.
Saha S, Sajib MSI, Garrett D, Qamar FN. Antimicrobial resistance in Typhoidal Salmonella: Around the world in 3 days. Clinical Infectious Diseases. 2020; 71 (Suppl 2):S91-S95 - 93.
Lin-Hui S, Chiu C-H, Chu C, Ou JT. Antimicrobial resistance in nontyphoid Salmonella serotypes: A global challenge. Clinical Infectious Diseases. 2004;39 (4):546-551 - 94.
Crump JA, Medalla FM, Joyce KW, Krueger AL, Hoekstra RM, Whichard JM, et al. Antimicrobial resistance among invasive nontyphoidal Salmonella enterica isolates in the United States: National antimicrobial resistance monitoring system, 1996 to 2007. Antimicrobial Agents and Chemotherapy. 2011; 55 :1148-1154 - 95.
Fey PD, Safranek TJ, Rupp ME, Dunne EF, Ribot E, Iwen PC, et al. Ceftriaxone-resistant Salmonella infection acquired by a child from cattle. The New England Journal of Medicine. 2000; 342 (17):1242-1249 - 96.
Threlfall EJ, Fisher IS, Berghold C, Gerner-Smidt P, Tschäpe H, Cormican M, et al. Antimicrobial drug resistance in isolates of Salmonella enterica from cases of salmonellosis in humans in Europe in 2000: Results of international multi-centre surveillance. Euro Surveillance. 2003; 8 (2):41-45 - 97.
Crump JA, Barrett TJ, Nelson JT, Angulo FJ. Reevaluating fluoroquinolone breakpoints for Salmonella enterica serotype Typhi and for non-Typhi Salmonellae. Clinical Infectious Diseases. 2003; 37 (1):75-81 - 98.
Mølbak K, Baggesen DL, Aarestrup FM, Ebbesen JM, Engberg J, Frydendahl K, et al. An outbreak of multidrug-resistant, quinolone-resistant salmonella enterica serotype typhimurium DT104. The New England Journal of Medicine. 1999; 341 (19):1420-1425 - 99.
Su LH, Chiu CH, Kuo AJ, Chia JH, Sun CF, Leu HS, et al. Secular trends in incidence and antimicrobial resistance among clinical isolates of Salmonella at a University Hospital in Taiwan, 1983-1999. Epidemiology and Infection. 2001; 127 (2):207-213 - 100.
Chiu CH, Su LH, Chu C, Chia JH, Wu TL, Lin TY, et al. Isolation of Salmonella enterica serotype choleraesuis resistant to ceftriaxone and ciprofloxacin. Lancet. 2004; 363 (9417):1285-1286 - 101.
Boerlin P, Reid-Smith RJ. Antimicrobial resistance: Its emergence and transmission. Animal Health Research Reviews. 2008; 9 :115-126 - 102.
Khan M, Shamim S. Understanding the mechanism of antimicrobial resistance and pathogenesis of Salmonella enterica serovar Typhi. Microorganisms. 2022;10 (10):2006 - 103.
Riyaaz AAA, Perera V, Sivakumaran S, de Silva N. Typhoid fever due to extended spectrum β- lactamase-producing Salmonella enterica serovar Typhi: A case report and literature review. Case Reports in Infectious Diseases. 2018;2018 :4610246 - 104.
Coipan CE, Westrell T, van Hoek AHAM, Alm E, Kotila S, Berbers B, et al. Genomic epidemiology of emerging ESBL-producing Salmonella Kentuckybla CTX-M-14b in Europe. Emerging Microbes & Infections. 2020;9 (1):2124-2135 - 105.
Robicsek A, Jacoby GA, Hooper DC. The worldwide emergence of plasmid-mediated quinolone resistance. The Lancet Infectious Diseases. 2006; 6 (10):629-640 - 106.
Acheampong G, Owusu M, Owusu-Ofori A, et al. Chromosomal and plasmid-mediated fluoroquinolone resistance in human Salmonella enterica infection in Ghana. BMC Infectious Diseases. 2019;19 :898 - 107.
Pavelquesi SLS, de Oliveira Ferreira ACA, Rodrigues ARM, de Souza Silva CM, Orsi DC, da Silva ICR. Presence of tetracycline and Sulfonamide resistance genes in Salmonella spp.: Literature review. Antibiotics (Basel). 2021;10 (11):1314 - 108.
Thai TH, Hirai T, Lan NT, Shimada A, Ngoc PT, Yamaguchi R. Antimicrobial resistance of Salmonella serovars isolated from beef at retail markets in the north Vietnam. The Journal of Veterinary Medical Science. 2012; 74 (9):1163-1169 - 109.
Deekshit VK, Kumar BK, Rai P, Srikumar S, Karunasagar I, Karunasagar I. Detection of class 1 integrons in Salmonella Weltevreden and silent antibiotic resistance genes in some seafood-associated nontyphoidal isolates of Salmonella in south-west coast of India. Journal of Applied Microbiology. 2012; 112 (6):1113-1122 - 110.
Kumar S, Ghosh RS, Iyer H, Ray A, Vannice K, MacLennan C, et al. Typhoid in India: An age-old problem with an existing solution. The Journal of Infectious Diseases. 2021; 224 (Supplement_5):S469-S474 - 111.
Mastroeni P, Chabalgoity JA, Dunstan SJ, Maskell DJ, Dougan G. Salmonella : Immune responses and vaccines. Veterinary Journal. 2001;161 :132-164 - 112.
Sears KT, Galen JE, Tennant SM. Advances in the development of Salmonella-based vaccine strategies for protection against Salmonellosis in humans. Journal of Applied Microbiology. 2021; 131 (6):2640-2658 - 113.
World Health Organization (WHO). Background Paper to SAGE on Typhoid Vaccine Policy Recommendations. Geneva: WHO; 2018 - 114.
Jin C, Gibani MM, Moore M, Juel HB, Jones E, Meiring J, et al. Efficacy and immunogenicity of a Vi-tetanus toxoid conjugate vaccine in the prevention of typhoid fever using a controlled human infection model of Salmonella Typhi: A randomised controlled, phase 2b trial. Lancet. 2017; 390 (10111):2472-2480 - 115.
Awad W, Ghareeb K. Some aspects of control of Salmonella infection in poultry for minimizing contamination in the food chain. World's Poultry Science Journal. 2014;70 (3):519-530 - 116.
Awuchi CG. HACCP, quality, and food safety management in food and agricultural systems. Cogent Food and Agriculture. 2023; 9 (1):2176280 - 117.
Mkangara M. Prevention and control of human Salmonella enterica infections: An implication in food safety. International Journal of Food Science. 2023;2023 :8899596 - 118.
Uelze L, Becker N, Borowiak M, et al. Toward an integrated genome-based surveillance of Salmonella enterica in Germany. Frontiers in Microbiology. 2021;12 :article 626941 - 119.
Jones FT. A review of practical Salmonella control measures in animal feed. Journal of Applied Poultry Research. 2011;20 :102-113 - 120.
Joint Food and Agriculture Organization (FAO)/World Health Organization (WHO) Expert Meetings on Microbiological Risk Assessment (JEMRA). Meeting Report. Prevention and Control of Microbiological Hazards in Fresh Fruits and Vegetables Part 4: Specific Commodities. 2023. Available from: https://www.who.int/publications/i/item/9789240077959 [Accessed: February 08, 2024] - 121.
Agirdemir O, Yurdakul O, Keyvan E, Sen E. Effects of various chemical decontaminants on Salmonella Typhimurium survival in chicken carcasses. Food Science and Technology. 2021; 41 (2):335-342 - 122.
Bettencourt Cota J, Vieira-Pinto M, Oliveira M. Biocide use for the control of non-Typhoidal Salmonella in the food-producing animal scenario: A primary food production to fork perspective [internet]. In: Salmonella-Perspectives for Low-Cost Prevention, Control and Treatment. London, UK: IntechOpen; 2024 - 123.
European Parliament, European Council. Regulation (EU) No 528/2012 of the European Parliament and of the Council of 22 May 2012 Concerning the Making Available on the Market and Use of Biocidal Products Text with EEA Relevance. 2012 - 124.
Fisher J. Types of disinfectant. In: Encyclopaedia of Food Science. Food Technology and Nutrition. Cambridge, Massachusetts, USA: Academic Press; 1993. pp. 1382-1385 - 125.
Zhou WY, Sun SF, Zhang YS, et al. Isolation and characterization of a virulent bacteriophage for controlling Salmonella Enteritidis growth in ready-to-eat mixed-ingredient salads. Journal of Food Protection. 2021;84 :1629-1639 - 126.
Albino LA, Rostagno MH, Hungaro HM, Mendonça RC. Isolation, characterization, and application of bacteriophages for Salmonell a spp. biocontrol in pigs. Foodborne Pathogens and Disease. 2014;11 :602-609 - 127.
Islam MS, Zhou Y, Liang L, Nime I, Liu K, Yan T, et al. Application of a phage cocktail for control of Salmonella in foods and reducing biofilms. Viruses. 2019;11 (9):841 - 128.
Meile S, Du J, Dunne M, Kilcher S, Loessner MJ. Engineering therapeutic phages for enhanced antibacterial efficacy. Current Opinion in Virology. 2022; 52 :182-191 - 129.
Khan MAS, Rahman SR. Use of Phages to treat antimicrobial-resistant Salmonella infections in poultry. Veterinary Sciences. 2022;9 (8):438 - 130.
Schmelcher M, Loessner MJ. Bacteriophage endolysins: Applications for food safety. Current Opinion in Biotechnology. 2016; 37 :76-87 - 131.
Kuralkar P, Kuralkar SV. Role of herbal products in animal production—An updated review. Journal of Ethnopharmacology. 2021; 278 :114246 - 132.
Alagawany M, Elnesr SS, Farag MR, Abd El-Hack ME, Barkat RA, Gabr AA, et al. Potential role of important nutraceuticals in poultry performance and health—A comprehensive review. Research in Veterinary Science. 2021; 137 :9-29 - 133.
Kotzekidou P, Giannakidis P, Boulamatsis A. Antimicrobial activity of some plant extracts and essential oils against foodborne pathogens in vitro and on the fate of inoculated pathogens in chocolate. LWT-Food Science and Technology. 2008; 41 (1):119-127 - 134.
Ruvalcaba-Gómez JM, Villagrán Z, Valdez-Alarcón JJ, Martínez-Núñez M, Gomez-Godínez LJ, Ruesga-Gutiérrez E, et al. Non-antibiotics strategies to control Salmonella infection in poultry. Animals (Basel). 2022;12 (1):102 - 135.
Gibson GR, Hutkins R, Sanders ME, Prescott SL, Reimer RA, Salminen SJ, et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nature Reviews Gastroenterology & Hepatology. 2017; 14 :491-502 - 136.
Tran THT, Everaert N, Bindelle J. Review on the effects of potential prebiotics on controlling intestinal enteropathogens Salmonella andEscherichia coli in pig production. Journal of Animal Physiology and Animal Nutrition. 2018;102 (1):17-32 - 137.
Ranjha MMAN, Shafique B, Batool M, et al. Nutritional and health potential of probiotics: A review. Applied Sciences. 2021; 11 (23):11204 - 138.
Villagran-de la Mora Z, Nuño K, Olga V, Avalos H, Castro-rosas J, Carlos G, et al. Effect of a synbiotic mix on intestinal structural changes, and Salmonella Typhimurium andClostridium perfringens colonization in broiler chickens. Animals. 2019;9 :777 - 139.
Vinderola G, Sanders ME, Salminen S, Szajewska H. Postbiotics: The concept and their use in healthy populations. Frontiers in Nutrition. 2022; 9 :1002213 - 140.
Gingerich E, Frana T, Logue CM, Smith DP, Pavlidis HO, Chaney WE. Effect of feeding a postbiotic derived from Saccharomyces Cerevisiae fermentation as a preharvest food safety hurdle for reducingSalmonella Enteritidis in the ceca of layer pullets. Journal of Food Protection. 2021;84 (2):275-280 - 141.
Wang HT, Yu C, Hsieh YH, Chen SW, Chen BJ, Chen CY. Effects of albusin B (a bacteriocin) of Ruminococcus Albus 7 expressed by yeast on growth performance and intestinal absorption of broiler chickens-its potential role as an alternative to feed antibiotics. Journal of the Science of Food and Agriculture. 2011;91 :2338-2343 - 142.
Wang L, Hu C, Shao L. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. International Journal of Nanomedicine. 2017; 12 :1227-1249 - 143.
Desin TS, Köster W, Potter AA. Salmonella vaccines in poultry: Past, present and future. Expert Review of Vaccines. 2013;12 (1):87-96 - 144.
Allafchian A, Vahabi MR, Jalali SAH, Mahdavi SS, Sepahvand S, Farhang HR. Design of green silver nanoparticles mediated by Ferula ovina Boiss. Extract with enhanced antibacterial effect. Chemical Physics Letters. 2022;791 :article 139392 - 145.
McEwen SA, Collignon PJ. Antimicrobial resistance: A one health perspective. Microbiology Spectrum. 2018; 6 (2). DOI: 10.1128/microbiolspec.arba-0009-2017 - 146.
Chernov VM, Chernova OA, Mouzykantov AA, Lopukhov LL, Aminov RI. Omics of antimicrobials and antimicrobial resistance. Expert Opinion on Drug Discovery. 2019; 14 (5):455-468