Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
Oxidative stress is an imbalance between oxidation and antioxidant responses caused by the accumulation of free radicals in the body. Cells produce free radicals that cause oxidative damage such as aging, atherosclerosis, arthritis, cancer, and neurological diseases. Antioxidants are substances that can neutralize free radicals, thereby reducing the damage to the body. Antioxidants are classified as synthetic and natural antioxidants. Natural products, mainly extracted from some medicinal plants, have strong antioxidant activity and low toxicity and side effects. Moreover, antioxidant activities are also detected in probiotics. Therefore, this chapter summarized the roles of antioxidants and then suggested antioxidants from natural sources have good prospects in the prevention and treatment of various diseases related to oxidative stress. In this study, antioxidant activities were mentioned in plants such as Basella alba, Cistanche sp., and representative probiotics such as Bifidobacterium breve ATCC 15700, Lactobacillus rhamnosus PN04, and Lactococcus lactis PN05 which could serve in DNA damage protection and cytotoxicity oriented for cancer treatment. With the results obtained in these medicinal herbs and probiotics, the antioxidant activities of more and more medicinal herbs and probiotics should be exploited in health care and pharmaceutical product development.
School of Biotechnology, International University, Hochiminh City, Vietnam
Vietnam National University, Hochiminh City, Vietnam
*Address all correspondence to: nhktu@hcmiu.edu.vn
1. Introduction
Cells maintain reactive oxygen species (ROS) homeostasis through various systems, such as the superoxide dismutase (SOD) system, catalase (CAT) system, and glutathione peroxidase (GSH-Px) system [1]. Antioxidants are a group of substances that help trap and neutralize free radicals, thereby reducing the damage to the body caused by free radicals. Additionally, antioxidants play a protective role against several diseases including inflammation and cancer caused by oxidative stress [2]. Antioxidants are classified as synthetic antioxidants and general natural antioxidants. However, synthetic antioxidants sometimes have toxicity and side effects, especially with the long-term use [3]. The exploration of natural antioxidants that are safe and free of side effects has become a hot spot in the past few years. Some natural products, mainly extracted from some medicinal plants and microorganisms, have strong antioxidant activity and low toxicity and side effects [3]. For this reason, antioxidants from natural sources have good application prospects in the prevention and treatment of various diseases related to oxidative stress. Due to reducing the chemical drugs’ negative effects, herbal medicines have gained their attractiveness in recent years. The natural sources contain vast components having antioxidant activities which are essential in cancer prevention, damaged DNA protection, and preventing lipid peroxidation [4]. For example, phenylethanoid glycosides (PhGs) in medicinal herbs act as a hydrogen donor to reductive radicals, allowing them to scavenge free radicals and regulate antioxidant enzymes involved in in vivo free radical metabolism [5].
Oxidation affects humans causing many diseases. Importantly, DNA is easy to spoil, leading to health problems in humans. DNA, the genetic material of all living organisms on the planet, is often influenced by a variety of variables, including exogenous and endogenous agents, which may alter its activity inside cells and ultimately lead to dramatic changes in living animals. The main exogenous factors can include UV light, ionizing radiation, toxic chemicals, and so on, whereas the primary endogenous factor is commonly caused by ROS [6]. The endogenous agents include superoxide anion, hydroxyl (OH) radicals, and hydrogen peroxide radicals. Furthermore, endogenous agent-caused damage occurs more often than exogenous agent-caused harm. If the damage is not addressed or corrected, it may lead to mutations and other health issues including aging, cancer, diabetes, reperfusion, cataracts, and lung failure.
Because of these, the therapeutic approach to alternative conventional medicine has expanded, such that increased research focused on the use of natural metabolites produced by medicinal herbs and lactic acid bacteria.
Life needs oxygen element for many biological activities. When oxygen damages the cells, oxidation occurs. Cells use oxygen to produce energy and free radicals. Reactive oxygen species (ROS) and reactive nitrogen species (RNS) are the by-products produced by cellular redox processes. These reactive species play dual roles in humans toxically and beneficially. At low or moderate levels, reactive species have beneficial effects on cellular and immune redox signaling function; however, high concentrations cause oxidative stress which can produce harmful substances, leading to spoiling cell function and structure. The toxic properties may be free radicals [6]. Free radicals play a role in the aging process. Free radicals cause oxidative stress, involving cardiovascular, neurological, and other human disorders. Antioxidants are our important defense against free radicals. ROS species such as superoxide, hydroxyl, and hydrogen peroxide are equivalent to the reduced oxygen levels. Oxidation stress is due to unhealthy lifestyles, exposure to chemicals, pollution, smoking, drugs, illness and sol. Antioxidants from plants, animals, and mineral sources have been shown to be beneficial to human health and effectively reduce the incidence of free radicals. Antioxidants are also involved in reducing free radicals, improving the normal function, and treating those diseases. Low levels of antioxidants or low levels of antioxidants in the blood increase the risk of diseases such as cancer. So, antioxidants in a healthy diet and natural antioxidants are now being recognized for protecting health from oxidative stress. Oxidative stress is through four important steps including membrane lipid peroxidation, protein oxidation, DNA damage, and disorders in reduction cellular equivalent, leading to cell destruction and changes in signaling pathways. Consequently, these mechanisms are implicated in the pathogenesis of diseases such as mitochondrial diseases in which cells cannot produce energy, leading to death [7]. Every biological molecule presenting in our body is at risk of free radicals. Such damaged cellular molecules may impair cellular function or may result in cell death. Lipids in the membranes of subcellular organelles are very susceptible to free radical damage. Lipid peroxidation leads to the generation of large amounts of toxic substances. Lipid peroxidation in cell membranes can damage the cell membranes by disrupting fluidity and permeability. Lipid peroxidation begins by removing hydrogen atoms from the methylene group, leading to the formation of a single electron on the carbon atom or carbon radical. Proteins are also susceptible to free radicals. Free radicals cause the destruction of many proteins and loss of enzyme activities. Protein hydroperoxide can be generated by ROS/RNS-induced protein oxidation, forming radicals. Although most oxidized proteins are essentially nonfunctional and eliminated quickly, some may accumulate gradually over time and thus contribute to aging-related damage as well as various diseases [5].
3. Antioxidant activities in DNA damage protection and cytotoxicity
3.1 Medicinal herbs
3.1.1 Phytochemicals served as antioxidants
Antioxidants are found in many vegetables, fruits, grains, tea, legumes, nuts, herbs and so on. Antioxidants may provide protection against diseases caused by free radicals. Phytochemicals or plant components are the main sources of antioxidants. The majority of these phytochemicals are redox-active molecules; therefore, they work to maintain a redox balance that is defined as an antioxidant. Phytochemicals are often classified as primary or secondary metabolites. The primary metabolites are sugars, amino acids, proteins, purines, and pyrimidines of nucleic acids, chlorophyll, etc., while secondary components include alkaloids (derived from amino acids), terpenes (a group of lipids), and phenolics (derived from carbohydrates). Plants produce an extremely impressive range of antioxidant compounds such as carotenoids, flavonoids, cinnamic acid, benzoic acid, folic acid, ascorbic acid, tocopherols, and tocotrienols to prevent oxidation of sensitive substrates. The plant-based antioxidants are believed to have better biological effects than synthetic ones because the plant components are part of the physiological processes of living flora, and thus, they are believed to have better compatibility with the human body. In this chapter, the plant extracts were studied for antioxidant activities. These plants have been used as daily vegetables in Vietnam.
3.1.2 Representative medicinal herbs
3.1.2.1 Basella alba (B. alba)
To clarify the roles of antioxidants, the study focused on the daily vegetable that can be used as a medicinal herb named Basella alba (B. alba). B. alba is one of the typical vegetables in Vietnam (Figure 1). B. alba is a good source of fiber, vitamins A, B, and C, thiamine, riboflavin, niacin, minerals such as calcium, magnesium, and iron, and several antioxidants. It also contains essential amino acids such as arginine, isoleucine, leucine, lysine, threonine, and tryptophan and a low percentage of soluble oxalates. B. alba has saponins, peptides, and phenolic compounds. B. alba leaves have anthocyanins, flavonoids, and phenolic acids which are good sources of phytochemicals and are also a potential source of antioxidants [8]. Because this vegetable is prepared in Vietnamese soup, the aqueous extract was performed. The extract was not soluble in methanol and ethanol; therefore, the antioxidant activities of the vegetable were determined by applying Fe3+ reducing power assay, reducing the power of the cucurbits which was determined by the previous method [9]. The reducing capacity of the polyphenolic extract of the B. alba leaves was measured by its ability to transform Fe3+ to Fe2+ at various concentrations (10, 25, 50, 75, 100, and 125 μg/mL). The results revealed that the reducing activity significantly increased (R2 = 0.9916), with the increased concentration at 250 μg/mL. The reductive capacity of a compound depends on the presence of reductones. The reducing power of the reference compound (gallic acid) was higher than the B. alba aqueous extract.
Figure 1.
Basella alba and antioxidant activity test.
3.1.2.2 Cistanches sp.
Cistanches belong to the family Orobanchaceae, and based on different chemical positions and genetic variations, the accepted genus Cistanche includes C. deserticola, C. tubulosa, C. sinensis, C. salsa, and C. salsa var. Albiflora [10].
C. derserticola can produce PhGs which belong to a group of natural compounds with many pharmacological properties, including antibacterial, antiviral, anticancer, anti-inflammatory, neuroprotective, antioxidant, and hepatoprotective activities. Individual PhGs have been shown to reduce apoptosis of neurons induced by various agents in in vitro and in vivo tests. Therefore, individual PhGs may have promising options for treating common neurodegenerative diseases including dementia and Parkinson’s disease [11]. Polysaccharides include 10 simple sugar bioactive components in Cistanche species including cinnamic acid, beta-glucopyranose, and hydroxyphenyl ethyl moieties, connected by ester and glycosidic bonds. Polysaccharides are responsible for enhancing immune function and fighting cancer. Furthermore, the free radical scavenging ability of polysaccharides that function as singlet and triplet oxygen scavengers, peroxide decomposers, enzyme inhibitors, and activators [12]. PhGs, and polysaccharides are also found in other plants which are considered as antioxidant sources. Moreover, Cistanche and others also contain ginsenosides including triterpene saponins and steroid glycosides called ginseng compounds. The common ginsenosides are Rb1, Rb2, Rg3, Rh2, Rh3, Rg1, Rg2, and Rh1. Typically, C. derserticola has ginsenosides (Rb1) and apigenin which are antioxidants and were detected by thin layer chromatography (TLC) (Figure 2).
Figure 2.
C. derserticola extracts checked for ginsenoside Rb1 (A) and apigenin (B).
They play a pivotal role in strong antioxidant effects in animal models and cell cultures. For instance, ginsenoside Rb1 exerts its antioxidant activity through two major mechanisms: chemical scavenging of certain types of ROS and a receptor-mediated genomic effect on antioxidant protein expression, such as SOD [13]. Secondarily, ginseng saponins have been shown to stimulate the formation of blood vessels and improve blood circulation in the brain, thereby enhancing memory and cognitive ability [14]. Additionally, ginsenoside Rg1 inhibits platelet activation via the inhibition of the ERK pathway and attenuates arterial thrombus formation in vivo [15].
Of all the flavonoids, apigenin (4′,5,7-trihydroxyflavone) is one of the most widely distributed flavonoids in the plant kingdom, and one of the most studied phenolics [16]. Apigenin can be found in significant amounts in vegetables, herbs, plants, and plant-based beverages. Like other flavonoids, apigenin reported numerous functional activities such as antioxidant properties [17], anti-hyperglycemic [18], anti-inflammatory, and (in myocardial ischemia) anti-apoptotic effects [19]. Therefore, the various biological activities of apigenin and several methods of extraction from natural sources, or also with the use of modern extractive approaches, are further analyzed.
ROS/RNS interferes with DNA, leading to oxidative damage. DNA is vulnerable to free radicals such as the hydroxyl group (∙OH). The C4-C5 double bond of pyrimidine is very sensitive to hydroxyl group attack, which generates a series of oxidative pyrimidine destruction products, such as thymine glycol, uracil glycol, urea residue, 5-hydroxydeoxyuridine, 5-hydroxydeoxycytidine, hydantoin, and others. Likewise, purines are also susceptible to the hydroxyl group, leading to the formation of 8-hydroxydeoxyguanosine (8-OHdG), 8-hydroxydeoxyadenosine, formamidopyrimidines, and purine products of oxidation. Free radical attack also caused the activation of the enzyme poly(ADP-ribose) synthetase; thus, DNA fragmentation and programmed cell death occur. This process is due to exhausting NAD+ levels in the cells, thereby disrupting the electron transport chain function. Free radicals such as the hydroxyl group react with carbohydrates. Plant extracts gave antioxidant activities. Based on this activity, the study evaluated the ability of the extract for the protection of DNA from UV/H2O2 effects.
3.1.3 DNA damage prevention and cytotoxicity
Different amounts of dry extracts showed similar protection levels, while there was no band of DNA exposed in UV/H2O2 (Figure 3). Figure 3 shows the loss of DNA under UV effect, but still remained in plant treatments.
Figure 3.
DNA damaged protection of C. derserticola extracts. (A) DNA before and after UV treatment; (B) DNA protection in extracts.
DNA damaged by UV was recovered because ginsenoside accelerated nucleotide excision repair (NER). NER is the important pathway that mammals can use to eliminate DNA lesions [20]. Rg2 contributed to p53 upregulation, but the mode of activation had not been reported [21]. Upregulation of p53 caused posttranslational modification that includes targeting murine double minute 2 (MDM2). MDM2 is an E3 ubiquitin ligase. MDM2 has oncogenic activities that might be dependent and in-dependent on p53. MDM2 was increased in many kinds of human cancer. Another protein also involves p53 that is wild-type p53-induced phosphatase 1 (WIP1) [22]. Moreover, the antioxidant property of ginseng was also reported to be involved in cell cycle arrest. Ginsenosides arrest the cell cycle via the c-Myc/p53 pathway [23]. The Myc gene is one of the oncogenes relating a nuclear protein involved in p53 expression and cell cycle. It is also reported that ginsenoside Rg3 reduces DNA damage by alkylating agents in normal cells [24]. In this study, tubulin was selected as the target protein to investigate different mechanisms in cell cycle arrest. Beside these tumor proteins, O6-methylguanine-DNA methyl transferase (MGMT) was also used to study on the interaction with ligands. Ligand-protein complexes were investigated on potential inhibition on MDM2, a primarily negative regulator of p53 and MGMT.
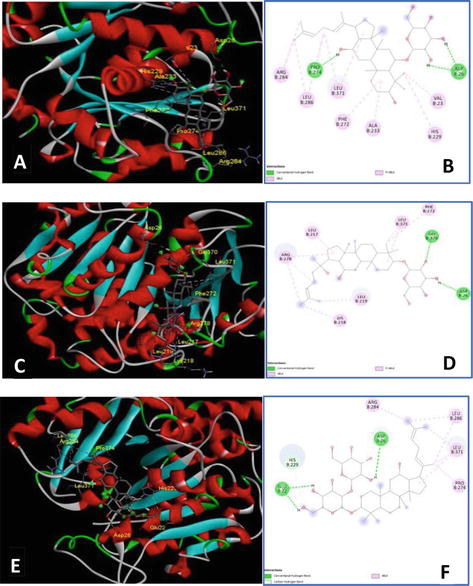
Additionally, tubulin beta class I (TUBB) protein belonging to globular protein is the major component of the cytoskeleton. It takes part in many important processes including cell mobility, cellular transport, and DNA segregation (i.e., mitosis). Residues involved in the inhibitor-binding site of TUBB validated by taxol molecule are Val23, Asp26, Leu217, Leu219, His229, Leu230, Ala233, Ser236, Phe272, Pro274, Leu275, Thr276, Ser277, Arg278, Pro260, Arg369, Gly370, and Leu371. Among them, the residues Pro274, Leu275, Thr276, Ser277, and Arg278 are claimed to be important for the taxol molecule. The selected constituents showed interaction to maximum two out of five aforementioned residues. Docking results showed that all the ligands mainly displayed hydrophobic interaction with the active residues. Only four ginsenosides could display hydrogen bonds with Asp26, mimicking the interaction between Taxotere and TUBB (Table 1, Figure 4). Ginsenoside Rh4 had the lowest binding energy as it showed three H-bonds with both Asp26 and Pro274. Despite occupying four H-bonds including Asp26 and Thr276, ginsenoside Rg3 was ranked 6 as it lacked hydrophobic interaction to stabilize. Due to the limit of hydrogen bonds, PN constituents showed weak interaction with TUBB featuring protein-ligand complexes that may be reversible. This confirmed PN compounds only temporarily halt the cell cycle allowing DNA to repair.
Rank
Ligands
Protein target
Binding energy value (kcal/mol)
Residues involved in interaction
1
Ginsenoside Rh4
TUBB
−8.12
Val23, Asp26, His229, Ala233, Phe272, Pro274,Arg284, Leu286, and Leu371
2
Ginsenoside Rh2
TUBB
−6.34
Asp26, Leu217, Lys218, Leu219, Phe272, Arg278,Gly370, and Leu371
3
Ginsenoside Rg5
TUBB
−6.17
Glu22, Asp26, His229, Pro274, Arg284, Leu286, and Leu371
4
Panaxydol
TUBB
−6.0
Phe272, Leu275, Thr276, Pro360, Arg369, Gly370, and Leu371
5
Falcarindiol
TUBB
−5.99
Leu217, His229, Leu230, Pro274, Leu275, Thr276,Arg284, Leu286, and Leu371
Phe272, Pro274, Thr276, Pro360, Leu371, and Gly370
8
Panaxynol
TUBB
−4.36
Leu217, His229, Leu230, Ala233, Phe272, Leu275,Ser277, and Arg278
Table 1.
Binding energy of compounds docked into tubulin beta protein.
Figure 4.
Best docking conformations and interactions between TUBB (shown in colored cartoon mode) and ginsenoside Rh4, Rh2, and Rg5 (gray stick), respectively. The 3D illustration demonstrated the interaction of TUBB with ginsenoside Rh4 (A), Rh2 (C), and Rg5 (E). 2D diagrams depicted interaction types of ginsenoside Rh4 (B), Rh2 (D), and Rg5 (F) and the active sites of TUBB.
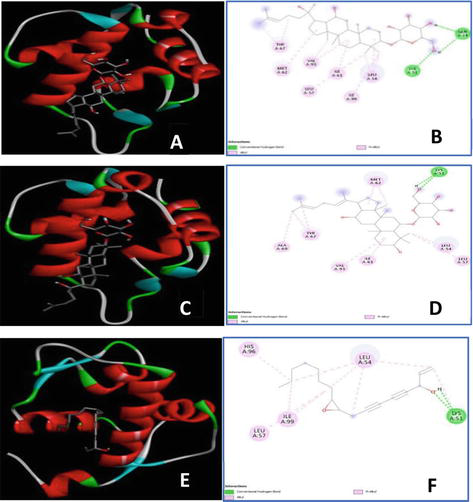
MGMT, commonly known as the suicide repair enzyme, plays a pivotal role in maintaining genome stability. MGMT repairs DNA-alkylated lesions at the O6 position of guanine. Such lesions may lead to DNA double-strand breaks or DNA mismatch resulting in mutations. The binding pocket of this enzyme is located at Tyr114, Pro138, Pro140, Cys145, Val148, Asn157, Tyr158, Ser159, and Lys165. The residue Cys145 is considered an important site for the occurrence of alkyl transfer. Only quercetin and the ligand group of polyacetylene could form hydrogen bonds with Cys145, whereas the ginsenoside showed H-interaction mainly with Ser159. Plus, the ginsenosides Rh2 and Rh4 were ranked second and third, respectively, but they only occupied one or two orientations with key residues, leaving most of the pocket empty. Although quercetin possessed the best binding energy, it created an unfavorable connection with Val128.
MDM2 is a regulator suppressing the expression of p53 protein [25]. At homeostasis, p53 is maintained at a low level via the MDM2-p53 complex (Table 2, Figure 5). This complex is mediated by hydrophobic interaction via three key residues of p53, namely, Phe19, Trp23, and Leu26. Inhibition of this negative regulator leads to the stability of p53 by mimicking p53 hydrophobic interaction with MDM2. Residues of MDM2 involved in the p53-MDM2 complex are Leu54, Leu57, Ile61, Met62, Gln72, His73, Val75, Val93, His96, Ile99, and Tyr100. MDM2 binding pocket can be divided into one large and main pocket Phe19/Trp23 and the subpocket Leu26. One ring of Rh2 formed hydrophobic bonds with Leu54, Leu57, Ile61, Met62, Val93, and Ile99 occupying pocket Phe19/Trp23. It also showed an H-interaction with Gln24 and a hydrophobic bond with Ile99 which was similar to Leu26 subpocket. Ginsenoside Rh4 ranked the second displayed linkage and interactions to the main Phe19/Trp23 pocket but left the subpocket Leu26 mostly empty (no interactions were found with Ile99). All the ginsenosides, flavonoids, and polyene ligands displayed strong affinity to MDM2; however, as this is an in silico test, further in vitro and in vivo experiments need to be performed to confirm this mode of interaction.
Rank
Ligands
Protein target
Binding energy value (kcal/mol)
Residues involved in interaction
1
Ginsenoside Rh2
MDM2
−8.16
Gln24, Lys51, Leu54, Leu57, Ile61, Met62,Tyr67, Val93, and Ile99
Binding energy of compounds docked into murine double minute 2 protein.
Figure 5.
Best docking conformations and interactions between MDM2 (shown in colored cartoon mode) and ginsenoside Rh2, Rh4, and panaxydol (gray stick), respectively. The 3D illustration demonstrated the interaction of MGMT with ginsenoside Rh2 (A), Rh4 (C), and panaxydol (E). 2D diagrams depicted interaction types of ginsenosideRh2 (B), Rh4 (D), panaxynol (F) and the active sites of MDM2.
From the results in Tables 1 and 2 and Figures 4 and 5, ginsenosides were predicted to interact with the oncogenes (MDM2 and TUBB). Then, via antioxidant activities of ginsenosides, these genes will suppress p53 and cell cycle [23, 24]. The prediction was also performed with MGMT to help with the effect of ginsenosides in DNA damage prevention and cancer treatment. Together with the roles of medicinal herbs in antioxidation relating to cancer treatment toward, probiotics are also discussed.
3.2 Probiotics
3.2.1 Probiotic concepts
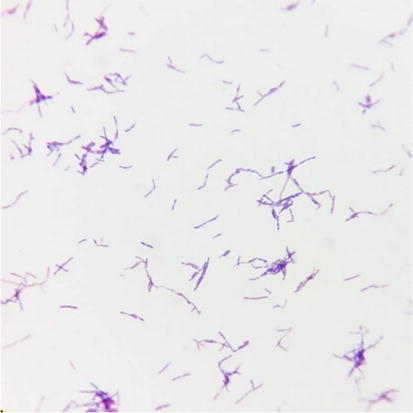
Lactic acid bacteria are important organisms honored for their fermentative capability as well as their health and nutritive benefits. They produce colorful composites similar to organic acids, diacetyl, hydrogen peroxide, and bacteriocin or bactericidal proteins during lactic restlessness. The lactic acid bacteria used commonly are Bifidobacterium strains. The Bifidobacterium genus belongs to the phylum Actinobacteria, class Actinobacteria, order Bifidobacteriales, and family Bifidobacteriaceae [26]. A more detailed description of Bifidobacterium genus is identified as a genus of Gram-positive bacteria, nonmotile [27]. Cells often stain irregularly with methylene blue and often branched anaerobic bacteria species (Figure 6), except Bifidobacterium psychrophilum, Bifidobacterium scardovii, and Bifidobacterium tsurumi tense which can tolerate O2 [28]. To date, there are 80 species (sub-species) in the genus Bifidobacterium, and this number is growing every year [29].
Figure 6.
The morphology of Bifidobacterium breve ATCC 15700 after gram-staining.
To date, bifidobacteria are regulated as dietary supplements and specific products. These species are also called as “generally recognized as safe” (GRAS) microorganisms [30]. A number of clinical studies claim that Bifidobacterium breve and other species (sub-species) in the genus Bifidobacterium can induce multiple health-promoting effects on its host through a variety of metabolites, including vitamins, polyphenols, conjugated linoleic acid, and short-chain fatty acids (SCFAs), proteins, or peptides [31].
Besides, the genera Bifidobacterium, Lactobacillus, Leuconostoc, Pediococcus, Enterococcus, and so on were used in food fermentation [32]. Only a few strains of Lactobacillus are used mainly in pseudomembrous colitis prevention such as Lactobacillus acidophilus. They produce acid, aroma, exopolysaccharides, proteins, and a variety of metabolites [32]. Therefore, these bacteria can involve antioxidant activity, DNA damage, and cancer prevention in a certain.
To find out the point of view, the study showed the antioxidant activity, DNA damage, and cancer prevention of Bifidobacterium breve ATCC 15700, Lactobacillus rhamnosus PN04, and Lactococcus lactis PN05 as representatives besides medicinal plants.
3.2.2 Representative probiotics
This chapter gave the research as an illustration of lactic acid bacteria benefits, especially the antioxidant activities involving DNA damage protection and cytotoxicity toward anticancer activity. Table 3 shows the survival rate of Saccharomyces cerevisiae (S. cerevisiae) compared to negative control sample (untreated sample). Based on results, hydroxyurea (HU) has a significant effect at a concentration of 100 mM. Besides, incubation time had influence on the protection ability of B. breve protein extracts. All extracts provided better DNA protection when better DNA protection when mixed with S. cerevisiae for 1–2 hours before treatment with H2O2 – HU complex. However, longer incubation time did not change the percentage of survival of S. cerevisiae. Bifidobacterium could prevent DNA damaged by hydroxyurea via antioxidant activity. The study performed in S. cerevisae which could play in vivo test which can be replaced for working in mice. The study also did in vitro test on DNA as Table 3. As a result, plant and LAB could prevent DNA damage via antioxidant agents.
Components
% Survival
CE (20 mg/mL)
H2O2 (1 mM) + HU (40 mM)
H2O2 (1 mM) + HU (100 mM)
Case 1
(hH)1
78.1003 ± 1.4394
54.2366 ± 1.4394
(hH)2
61.5917 ± 1.9819
47.5754 ± 0.79
(hH)3
67.8872 ± 2.8716
41.8788 ± 2.9496
Case 2
(hH)1E
67.214 ± 0.6169
53.8313 ± 0.0353
(hH)2E
69.2042 ± 0.2447
45.266 ± 0.3657
(hH)3E
68.4472 ± 2.5178
38.3967 ± 1.35927
Case 3
(hHE)1
68.1475 ± 1.2307
65.5729 ± 1.4192
(hHE)2
67.9151 ± 0.273
59.2573 ± 0.6829
(hHE)3
69.9547 ± 2.7488
69.5909 ± 1.1288
Case 4
E(hH)1
79.5539 ± 2.7549
68.9468 ± 0.9499
E(hH)2
74.7305 ± 0.7937
68.8953 ± 0.7768
E(hH)3
67.5318 ± 1.5285
40.75504 ± 0.217604735
Table 3.
Survival percentage of Saccharomyces cerevisiae in oxidative stress and antioxidant effects.
Case 1: S. cerevisae was treated in hydroxyurea (HU) and H2O2 in 1, 2, and 3 hours. Case 2: extracts were added into yeast after adding H2O2 – HU (100mM) and incubated at 1 h named (hH)1; 2 h named (hH)2; and 3 h named (hH)3. Case 3: extract, HU, and H2O2 were added in yeast culture and incubated in 1 h, 2 h, and 3 h. Case 4: extract was incubated with yeast for 1, 2, and 3 h and then added with HU and H2O2 followed more incubation.
When comparing with case 1, the survival rate was significant higher. Furthermore, gradually increasing incubation time resulted in increased survival (53.8314 ± 0.0353% to 57.9562 ± 1.5168% when extracts were added and incubated at 1–2 hours after adding H2O2 – HU (100 mM) (case 2) and 65.5729 ± 1.4192% to 69.5909 ± 1.1288% when adding extracts and H2O2 – HU (100 mM) at the same time (case 3). For case 4, extract was added in yeast culture to protect the yeast survival capacity, and then the mixture was contacted with H2O2 – HU (40 mM and 100 mM). It seemed that the yeast survival rate was improved than case 1, case2, and case 3 during the 2-hour incubation. Especially, H2O2 – HU (100 mM) caused the lower survival rates, but case 4 could improve survival rate when comparing to cases 1 and 2. However, the survival rate was low in case 4 during the 3-hour incubation. Probably, any active component of bacteria was degraded by yeast during longer incubation. Actually, the incubation time of the extract in case 4 was longer than 3 h because the extract was incubated with yeast before and then prolonged 3 h incubation in HU and H2O2. Anyway, the bacterial extract could prevent the toxicity of HU and H2O2 by means of the antioxidant activity.
The protection of incentive cells by the expert under the effect of H2O2 – HU is harmonious with the compliances above, suggesting the possible part of the excerpt in the junking of hydrogen peroxide (H2O2) revolutionaries. This supports the excerpt’s antioxidant exertion by demonstrating its part in guarding cells from DNA dangerous chemicals that the release of OH- revolutionaries from H2O2 – HU complex is the main reason for converting DNA damage in cells [33, 34]. The protection of Bifidobacteria for prokaryotic and eukaryotic cells involved in numerous DNA mismatch form (DMR) protein. DMR’s top function is to ameliorate replication dedication by repairing replication-associated base-base mismatches. Modulating DNA recombination and easing DNA damage signaling are significant primary functions [35]. DMR blights are connected with cancer vulnerability, including heritablenon-polyposis colorectal cancer, resistance to certain chemotherapeutic agents, and anomalies in meiosis and sterility in mammalian systems [36]. Inactivation of DMR in cells causes sporadic malice in humans. In response to some forms of DNA damage, the DMR system is necessary for cell cycle arrest and/or programmed cell death. This capability was inactivated when exposed to high attention of H2O2 (1 mM). As a result, when adding HU− a medicine applied to treat neoplastic conditions but has a side effect of inhibits DNA conflation, S. cerevisiae vitality was being dropped. Former study showed that S. cerevisiae mutants lacking CuZnSOD have been shown to be hypersensitive to hypertonic medium and to parade elevated oxidative damage [37]. This study aimed to estimate the capability of the excerpt to recover damage to S. cerevisiae under stressors and to estimate the antioxidant capacity. S. cerevisiae came weak and unable of resistance to DNA mutagens called hydroxyurea (HU). Numerous exploration studies have concentrated on anticancer and antioxidative parcels as defensive adjuncts against a host of conditions [38]. Although colorful synthetic agents parade anticancer and antioxidative conditioning, pointing the agents which are not only precious, but stable and safe is important. Multitudinous reports have indicated that both LAB and investigated milk as anticancer goods [39]. Data from epidemiological and experimental studies have also indicated that the ingestion of certain LAB strains, or of fermented dairy products, might palliate the threat of certain type of cancers and inhibit the growth of excrescences. In an in vitro environment, the anticancer and antioxidative parcels of heat-killed (HK) cells and fractionations of HK cells of several Lactobacillus strains.
Lactic acid bacteria including Lactobacillus acidophilus and Lactobacillus casei to test the apoptosis of human cancer colon cell line [40, 41]. In recent years, there have been many studies using animal models, which have clearly demonstrated a protective effect of dietary supplements of LAB against colon tumor development [42]. The antioxidant activity of the extracts of the culture of Lactobacillus rhamnosus PN04 and Lactococcus lactis PN05 showed the DPPH (2,2′-diphenyl-1-picrylhydrazyl radical) scavenging assay [43]. The effect of two LAB strains to cancer cell lines is fairly similar. Indeed, the stationary phase produces the highest amount of antioxidant activity (Table 4). So, only culture supernatant and polysaccharide at this phase were performed test antitumor activity on HeLa and Hep G2 cells. Depending on the different effect mechanism to cancer cell lines, the cytotoxicity percentage of each sample is also different. For example, culture supernatant of Lactobacillus rhamnosus PN04 (64.40) was more effective than polysaccharide (58.78) on Hep G2 cells. However, there is a converse on HeLa cells with values of 46.99 and 86.16 respectively. Similarly, the culture supernatant of Lactococcus lactis PN05 (66.09) was more effective than polysaccharide (50.36) on Hep G2 cells. However, there is a converse on HeLa cells with values of 34.60 and 86.86, respectively. These results indicated that both culture supernatant and polysaccharide of Lactobacillus rhamnosus PN04 and Lactococcus lactis PN05 potently inhibited the viability of each cell line. Consequently, lactic acid bacteria can show cytotoxicity due to antioxidant activities. Lactic acid bacteria can protect DNA based on antioxidant action, leading to anticancer activity prospectively. The curious question is whether lactic acid bacteria can involve DNA repair/protection that makes lactic acid bacteria kill cancer cells, but not normal cells. The study brings out the important effects of antioxidant activities for health prevention that should be clarified more.
Growth phase
% DPPH scavenging activity of culture supernatant
Early exponential phase
93.17 ± 0.075
Late exponential phase
94.33 ± 0.117
Stationary phase
94.69 ± 0.075
Death phase
94.31 ± 0.075
Table 4.
Antioxidant activities of Lactobacillus rhamnosus PN04 in culture phases.
Consequently, antioxidant activities of probiotics/lactic acid bacteria could involve in DNA damage protection. Many metabolites were produced in the bacteria; however, exopolysaccharides were studied most in cytotoxicity. This activity of lactic acid bacteria was like other species such as seaweeds [44]. It is undoubtable. Therefore, the lactic acid bacteria could be exploited for cancer therapy development by finding their antioxidants in the future.
Antioxidant conditioning can be detected in both medicinal plants and lactic acid bacteria. Antioxidant agents could prevent cytotoxicity and DNA damage. This chapter showed the antioxidant activities of daily vegetables such as Basella alba or Cistanche sp., a source of functional food together with a few representative probiotics such as Bifidobacterium breve ATCC 15700, Lactobacillus rhamnosus PN04, and Lactococcus lactis PN05 which could be served in DNA damage protection and cytotoxicity oriented for cancer treatment. The hydroxyl-free revolutionaries are known to beget damage to cellular DNA. UV photolysis of H2O2 is responsible for utmost oxidative damage to proteins and DNA. The study mentioned the representatives as the daily food or functional food. Thus, medicinal plants and probiotics for damaged DNA protection and cytotoxicity toward cancer treatment should be focused more. This chapter recommends that medicinal plants and lactic acid bacteria in health forestallment should be exploited more for DNA damage prevention and cancer treatment based on antioxidant activities.
1.He L, He T, Farrar S, Ji L, Liu T, Ma X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell Physiology and Biochemistry. 2017;44(2):532-553
2.Cao L, Xu X, Cao LL, Wang RH, Coumoul X, Kim SS, et al. Absence of full-length Brca1 sensitizes mice to oxidative stress and carcinogen-induced tumorigenesis in the esophagus and forestomach. Carcinogenesis. 2007;28:1401-1407
3.Mahmoud AM, Wilkinson FL, Sandhu MA, Dos Santos JM, Alexander MY. Modulating oxidative stress in drug-induced injury and metabolic disorders: The role of natural and synthetic antioxidants. Oxidative Medicine and Cellular Longevity. 2019;2019:3206401
4.Conrad M, Kagan VE, Bayir H, et al. Regulation of lipid peroxidation and ferroptosis in diverse species. Genes and Development. 2018;32(9-10):602-619
5.Knecht KT, Bradford BU, Mason RP, Thurman RG. In vivo formation of a free radical metabolite of ethanol. Molecular Pharmacology. 1990;38(1):26-30
6.Collins AR. Oxidative DNA damage, antioxidants, and cancer. BioEssays. 1999;21(3):238-246
7.Rahaman MM, Hossain R, Herrera-Bravo J, et al. Natural antioxidants from some fruits, seeds, foods, natural products, and associated health benefits: An update. Food Science & Nutrition. 2023;11(4):1657-1670
8.Deshmukh SA, Gaikwad DK. A review of the taxonomy, ethnobotany, phytochemistry and pharmacology of Basella alba (Basellaceae). Journal of Applied Pharmaceutical Sciences. 2014;4(01):153-165
9.Güder A, Korkmaz H. Evaluation of in-vitro antioxidant properties of hydroalcoholic solution extracts Urtica dioica L., Malva neglecta Wallr. and their mixture. Iranian Journal of Pharmaceutical Research. 2012;11(3):913-923
10.Song Y, Zeng K, Jiang Y, Tu P. Cistanches Herba, from an endangered species to a big brand of Chinese medicine. Medicinal Research Reviews. 2021;41(3):1539-1577
11.Huang J, Zhao D, Cui C, Hao J, Zhang Z, Guo L. Research progress and trends of phenylethanoid glycoside delivery systems. Food. 2022;11(5):769
12.Sui ZF, TingMin G, Liu B, Peng SW, Zhao ZL, Li L, et al. Water-soluble carbohydrate compound from the bodies of Herba Cistanches: Isolation and its scavenging effect on free radical in skin. Carbohydrate Polymers. 2011;85(1):75-79
13.Salekdeh GH, Siopongco J, Wade LJ, Ghareyazie B, Bennett J. Proteomic analysis of rice leaves during drought stress and recovery. PROTEOMICS: International Edition. 2022;2(9):1131-1145
14.Lü JM, Jiang J, Jamaluddin M, Liang Z, Yao Q , Chen C. Ginsenoside Rb1 blocks ritonavir-induced oxidative stress and eNOS downregulation through activation of estrogen receptor-beta and upregulation of SOD in human endothelial cells. International Journal of Molecular Sciences. 2019;20(2):294
15.Oliynyk S, Oh S. Actoprotective effect of ginseng: Improving mental and physical performance. Journal of Ginseng Research. 2023;37(2):144-166
16.Zhou Q , Jiang L, Xu C, et al. Ginsenoside Rg1 inhibits platelet activation and arterial thrombosis. Thrombosis Research. 2014;133(1):57-65
17.Salehi B, Venditti A, Sharifi-Rad M, Kręgiel D, Sharifi-Rad J, Durazzo A, et al. The therapeutic potential of apigenin. International Journal of Molecular Sciences. 2019;20(6):1305
18.Villa-Rodriguez JA, Kerimi A, Abranko L, et al. Acute metabolic actions of the major polyphenols in chamomile: An in vitro mechanistic study on their potential to attenuate postprandial hyperglycaemia. Scientific Reports. 2018;8(1):5471
19.Lim R, Barker G, Wall CA, Lappas M. Dietary phytophenols curcumin, naringenin and apigenin reduce infection-induced inflammatory and contractile pathways in human placenta, foetal membranes and myometrium. Molecular Human Reproduction. 2013;19(7):451-462
20. Zhou Z, Zhang Y, Lin L, Zhou J. Apigenin suppresses the apoptosis of H9C2 rat cardiomyocytes subjected to myocardial ischemia-reperfusion injury via upregulation of the PI3K/Akt pathway. Molecular Medicine Reports. 2018;18(2):1560-1570
21.Cai BX, Jin SL, Luo D, Lin XF, Gao J. Ginsenoside Rb1 suppresses ultraviolet radiation-induced apoptosis by inducing DNA repair. Biological and Pharmaceutical Bulletin. 2009;32(5):837-841
22.Chung YH, Jeong SA, Choi HS, Ro S, Lee JS, Park JK. Protective effects of ginsenoside Rg2 and astaxanthin mixture against UVB-induced DNA damage. Animal Cells and Systems. 2018;22(6):400-406
23.Ladds M, Laín S. Small molecule activators of the p53 response. Journal of Molecular Cell Biology. 2019;11(3):245-254
24.He NW, Zhao Y, Guo L, Shang J, Yang XB. Antioxidant, antiproliferative, and pro-apoptotic activities of a saponin extract derived from the roots of panax notoginseng (Burk.) F.H. Journal of Medicinal Food. 2012;15(4):350-359
25.Li T, Sun W, Dong X, et al. Total ginsenosides of Chinese ginseng induces cell cycle arrest and apoptosis in colorectal carcinoma HT-29 cells. Oncology Letter. 2018;16(4):4640-4648
26.Zhang YH, Li HD, Li B, Jiang SD, Jiang LS. Ginsenoside Rg3 induces DNA damage in human osteosarcoma cells and reduces MNNG-induced DNA damage and apoptosis in normal human cells. Oncology Reports. 2014;31(2):919-925
27.Hidalgo-Cantabrana C, Delgado S, Ruiz L, Ruas-Madiedo P, Sánchez B, Margolles A. Bifidobacteria and their health-promoting effects. Microbiology Spectrum. 2017;5(3):10
28.Alessandri G, van Sinderen D, Ventura M. The genus bifidobacterium: From genomics to functionality of an important component of the mammalian gut microbiota running title: Bifidobacterial adaptation to and interaction with the host. Computational and Structural Biotechnology Journal. 2021;19:1472-1487
29.Whitman WB, Rainey F, Kämpfer P, Trujillo M, Chun J, DeVos P, et al., editors. Bergey’s Manual of Systematics of Archaea and Bacteria. 1st ed. Wiley; 2015
30.Turroni F, Duranti S, Milani C, Lugli GA, van Sinderen D, Ventura M. Bifidobacterium bifidum: A key member of the early human gut microbiota. Microorganisms. 2019;7(11):544
31.Petersen AB, Gniadecki R, Vicanova J, Thorn T, Wulf HC. Hydrogen peroxide is responsible for UVA-induced DNA damage measured by alkaline comet assay in HaCaT keratinocytes. Journal of Photochemistry and Photobiology. B, Biology. 2000;59(1-3):123-131
32.Wang Y, Wu J, Lv M, Shao Z, Hungwe M, Wang J, et al. Metabolism characteristics of lactic acid bacteria and the expanding applications in food industry. Frontiers in Bioengineering and Biotechnology. 2021;9:612285
33.Koziol S, Zagulski M, Bilinski T, Bartosz G. Antioxidants protect the yeast Saccharomyces cerevisiae against hypertonic stress. Free Radical Research. 2005;39(4):365-371
34.Fakruddin M, Hossain MN, Ahmed MM. Antimicrobial and antioxidant activities of Saccharomyces cerevisiae IFST062013, a potential probiotic. BMC Complementary and Alternative Medicine. 2017;17(1):64
35.Golla U, Bhimathati SS. Evaluation of antioxidant and DNA damage protection activity of the hydroalcoholic extract of Desmostachya bipinnata L. Stapf. Scientific World Journal. 2014;2014:215084
36.Garbacz K. Anticancer activity of lactic acid bacteria. Seminars in Cancer Biology. 2022;86(Pt 3):356-366
38.Li GM. Mechanisms and functions of DNA mismatch repair. Cell Research. 2008;18(1):85-98
39.Stojic L, Brun R, Jiricny J. Mismatch repair and DNA damage signalling. DNA Repair (Amst). 2004;3(8-9):1091-1101
40.Zhong L, Zhang X, Covasa M. Emerging roles of lactic acid bacteria in protection against colorectal cancer. World Journal of Gastroenterology. 2014;20(24):7878-7886
41.Baldwin C, Millette M, Oth D, Ruiz MT, Luquet FM, Lacroix M. Probiotic Lactobacillus acidophilus and Lactobacillus casei mix sensitize colorectal tumoral cells to 5-fluorouracil-induced apoptosis. Nutrition and Cancer. 2010;62(3):371-378
42.Kim SH, Moon JY, Lim YJ. Dietary intervention for preventing colorectal cancer: A practical guide for physicians. Journal of Cancer Prevention. 2022;27(3):139-146
43.Nguyen DT, Nguyen TH. Detection on antioxidant and cytotoxicity activities of exopolysaccharides isolated in plant-originated Lactococcus lactis. Biomedical and Pharmacology Journal. 2014;7(1):33-38
44.Pradhan B, Bhuyan PP, Ki JS. Immunomodulatory, antioxidant, anticancer, and pharmacokinetic activity of Ulvan, a seaweed-derived sulfated polysaccharide: An updated comprehensive review. Marine Drugs. 2023;21(5):300
Written By
Tu Nguyen
Submitted: 19 December 2023Reviewed: 12 January 2024Published: 08 March 2024