
Abstract
Recently, the role of the gut microbiome has become more prominent in gut-brain interactions. The microbiota-gut-brain axis homeostasis is responsible for our emotional behavior, stress response, and brain neurotransmitter balance. This bidirectional communication axis between the gut and the brain is influenced by the effect of the microbiome on the metabolic pathways of the host. Intestinal bacteria intervene directly in Trp metabolism, generating signaling molecules and specific metabolites with physiological effects on both the brain and the intestine. Trp is also metabolized under the influence of the microbiome and suffers three major pathways in the organism: The serotonin (5-HT), kynurenine, and indole pathway resulting in the production of neuro-active metabolites. This current chapter aims to cover the most recent data referring to the Trp metabolites and the microbiome-gut-brain axis in major gut disorders, such as irritable bowel syndrome (IBS), inflammatory bowel disease (IBD), and celiac disease. These diseases are frequently associated with mood disorders.
Keywords
- microbiome
- indole
- tryptophan
- kynurenine pathway
- serotonin
1. Introduction
Tryptophan (Trp) is an essential aromatic amino acid necessary for the normal growth and health of mammalian species [1]. The main source of Trp is the diet, however, small amounts can be produced by the gut microbiota,
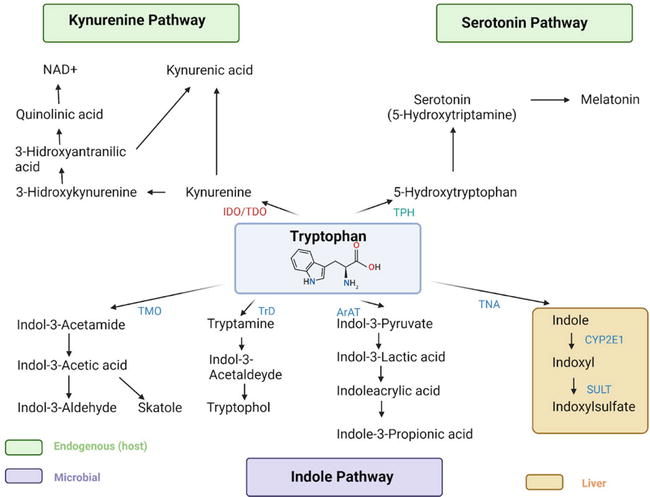
Figure 1.
The tryptophan metabolism. Tryptophan metabolism occurs via the kynurenine pathway or the serotonin pathway. Kynurenine is metabolized to kynurenic or quinolinic acid, which is then converted to nicotinamide adenine dinucleotide (NAD+). In the serotonin pathway, Trp is converted to 5-HTP in the enterochromaffin cells and enteric neurons and can be further metabolized to form melatonin. Microbiota metabolize Trp into indole and indole derivatives because they express different enzymes, resulting in end-products. Abbreviations: ArAT - aromatic amino acid aminotransferase; IDO - indoleamine 2,3-dioxygenase; NAD+- nicotinamide adenine dinucleotide; TDO - tryptophan 2,3-dioxygenase; TPH - tryptophan hydroxylase; TrD - tryptophan decarboxylase enzyme; TNA - tryptophanase; SULT - sulfotransferase. Created with BioRender.
Multiple studies demonstrated that the gut microbiota plays a key role in the gut-brain axis modulation by influencing the production of metabolites [10, 11].
Nowadays, microbiota located in the gut is regarded as a “virtual organ” outnumbering the human cells by a ratio of at least 10:1 [12]. However, this ratio was criticized by Sender et al. who suggested that the ratio is in fact 1:1 [13].
The intestinal microbiota is essential for normal brain development and for normal human behavior in adulthood, mainly based on the metabolism of Trp. This was demonstrated by studies on germ-free animals. These studies brought tremendous achievements in this regard, because it was demonstrated that the high Trp levels observed in germ-free mice decreased after colonization of their gut, indicating that microbiota plays a role in this decrease and also in the development of central serotonergic systems [14].
Some
The brain-gut axis is a bidirectional communication network between the gastrointestinal tract, the (ENS), and the (CNS), mediated primarily by serotonin. Serotonin plays a major role in the normal functioning of the gut-brain axis, acting as a neurotransmitter in both the CNS and the ENS [18].
Numerous sensations and functions are under the bidirectional (sensory and motor) control of vagal system: Nausea, fullness, visceral pain, peristalsis, sphincter control, etc. During the development of the enteric vagal innervation, the microbiota might influence the developing axons of the ENS [19]. The gut microbiota is apparently responsible for the maturation of the adult ENS through the release of 5-HT and activation of 5-HT4 receptors, indicating its involvement in the normal peristaltic function of the gut [20].
Trp metabolites, either of endogenous origin (kynurenine, kynurenic acid, and NAD+) or resulting from bacterial degradation (indole, indole propionic acid, indole acetic acid, skatole, and tryptamine), are involved in gut immune homeostasis and mucosal homeostasis, and in serotonergic signaling [21].
Trp metabolites produced by microbes play an important role in gut physiology. The Trp metabolite indole has anti-inflammatory activities and
Therefore, it is considered that dysbiosis is implicated in many functional disorders of the gut-brain axis, such as (IBS), irritable bowel disease (IBD), and certain neuropsychiatric disorders like autism or depression [25, 26, 27, 28].
In this chapter, we summarize the Trp metabolic pathways and we provide an in-depth description of their interrelation with the gut microbiota and their impact on the gut-brain axis in major gut disorders, such as IBS, IBD, and celiac disease (CeD).
2. Trp metabolic pathways
2.1 The serotonin pathway
The serotonin synthesis occurs primarily in the gastrointestinal tract (90%) and partially in the CNS (10%) starting from its precursor Trp [5]. The initial step in the serotonin pathway is represented by the Trp hydrolysis to 5-hydroxy Trp (5-HTP) via Trp hydroxylase 1 and 2 (TPH1/2). TPH1 is found in the periphery, while TPH2 is found in the CNS. 5-HTP is then rapidly decarboxylated by the aromatic L-amino decarboxylase (ALAAD) to 5-hydroxytryptamine (5-HT) known as serotonin [29].
5-HT is finally converted to melatonin in the pineal gland and the periphery or it is metabolized by monoamine oxidase (MAO) to 5-hydroxyindole acetaldehyde, which aldehyde dehydrogenase (ALDH) further oxidizes to 5-hydroxyindoleacetic acid (5-HIAA). The serotonin metabolite 5-HIAA is eliminated via urinary excretion [30].
2.2 The kynurenine pathway
Only 5% of the Trp is utilized as a substrate for the production of 5-HT. The remaining fraction of Trp enters the kynurenic pathway [31]. The Trp degradation arises primarily in the liver via the Trp 2,3-dioxygenase (TDO) and partially extrahepatic in the brain and the gastrointestinal tract via the indoleamine 2,3-dioxygenase (IDO). TDO and IDO are enzymes induced by glucocorticoids and pro-inflammatory cytokines and convert Trp to kynurenine as a first step in the kynurenine pathway [32]. Therefore, infections and chronic stress can reduce the amount of Trp that is converted to serotonin having a negative effect on mental health and the neuroendocrine stress axis [33, 34, 35].
After this initial step, kynurenine can be catabolized by three different enzymes.
Kynurenine aminotransferase (KAT) converts kynurenine into kynurenic acid (KA). KA is an N-methyl-D-aspartate (NMDA) receptor antagonist and α7 nicotinic acetylcholine receptor and it exhibits immunosuppressive, anti-oxidative, and neuroprotective roles [31].
Part of kynurenine can also be converted to 3-hydroxykynurenine (3-HK) by the kynurenine-3-monooxygenase (KMO). 3-HK is further converted to 3-hydroxyanthranilic (3-HAA) via the enzyme kynureninase. Both 3-HK and 3-HAA are neurotoxic and oxidative mediators [31]. 3-HK has been shown to induce endogenous oxidative stress and neuronal cell apoptosis and has been linked to a series of neurodegenerative disorders [36]. 3-HK can further be converted to xanthurenic acid (XA). 3-HAA can be converted to quinolinic acid (QA), which is further catabolized into nicotinamide adenine dinucleotide (NAD+) and niacin. KA and QA are both neuroactive inflammatory mediators [5]. Studies have found a link between high levels of QA and numerous neurological disorders such as anxiety, depression, Huntington’s disease, and Alzheimer’s disease [37, 38, 39]. In addition, kynureninase converts kynurenine to anthranilic acid (AA), a metabolite generally accepted as biologically inactive [40].
2.3 The indole pyruvate pathway
Most of the available Trp comes from dietary intake via foods such as nuts, seeds, milk, cheese, or tuna, and the majority of Trp is absorbed in the small intestine. However, the fraction that remains unabsorbed serves as a substrate for the gut microbes [5]. Through microbial degradation, Trp is metabolized into indole and indole derivatives via the indole pathway.
Several microbes in the gut express enzymes that use Trp to generate indole and other indole catabolites [5]. The microbial species involved in the indole pathway have been summarized in recent reviews [41, 42]. Trp is converted to indole via the enzyme Trp identified in gut bacteria such as
The first step in the degradation of Trp to indole-3-propionic acid (IPA) is the conversion to indole-3-pyruvic acid (IPYA) via the aromatic amino acid decarboxylase, followed by the conversion to ILA and anholocyclic acid (IA), and finally IPA. In addition, through different enzymes gut microbes degrade Trp to indole-3-aldehyde (IAId) and indole-3-acetaldehyde (IAAId) [5].
3. Trp metabolites and gastrointestinal disorders
3.1 Irritable bowel syndrome
IBS is a functional gastrointestinal disorder characterized by chronic abdominal pain associated with altered bowel habits [44]. It is the most commonly diagnosed gastrointestinal disorder, representing a significant social problem [45]. Visceral hypersensitivity accounts for the symptoms of chronic pain that patients with IBS report to their physicians. Pain perception might be influenced by the gut microbiota, which participates in pain signaling [46]. Species like
The stimulation of receptors in the gut wall sends signals via the spinal cord to the brain [48]. Specific mediators such as serotonin and kinins have been associated with the visceral pain observed in IBS patients along with the increased spinal cord excitability as a result of the NMDA receptor [49, 50, 51].
The role of Trp metabolites in IBS has been demonstrated by Fitzgerald et al. when comparing patients with IBS to healthy controls. They found that IBS patients’ blood had more kynurenine than the control group, and that there was a positive link between the severity of IBS symptoms and the kynurenine/tryptophan (Kyn/Trp) ratio [52]. A shift in Trp metabolism toward kynurenine production has also been observed by Han et al. along with a link between a Trp dysregulated metabolism and the severity of IBS depression [53].
Several other authors have observed increased levels of serotonin in patients with IBS compared to healthy controls. By comparing serum 5-HT levels under fasting conditions in patients with IBS and healthy controls Moskwa et al. demonstrated that serotonin levels are increased in both patients with diarrhea predominant and constipation predominant IBS [54]. However, Atkinson et al. reported increased levels of 5-HT in patients with diarrhea predominant IBS in both fasting and fed conditions, whereas patients with constipation predominant IBS did not show increased levels of 5-HT after meal ingestion. The results of this study suggest that reduced serotonin reuptake characterizes patients with diarrhea predominant IBS [55]. Other studies have demonstrated that serotonin levels are not significantly different in patients with IBS compared to controls. However, an altered serotonin metabolism appears to be implicated, with decreased 5-HIAA levels significantly lower 5-HIAA/5-HT ratio [56].
The mechanisms underlying altered plasma serotonin levels in IBS remain incompletely understood. However, in addition to modulating psychological and behavioral processes through the CNS, serotonin, and its metabolites are also known to play important modulatory roles in the ENS, co-regulating intestinal secretion, motility, and visceral perception [57]. One study demonstrated an association between increased plasma 5-HT levels in patients with IBS and the presence of
The 5-HT mucosal levels also play an important role. Existing studies demonstrated that the 5-HT levels are lower in IBS patients compared to the healthy controls and it is associated with a low expression of the re-uptake transporters for TPH1 and serotonin [59]. According to the studies, there might be differences in the 5-HT levels between IBS subtypes, since in IBS-C the levels are low in the colonic mucosa, and increased for IBS-D, respectively [60].
3.2 Inflammatory bowel diseases
Inflammatory bowel diseases (IBDs) are a group of chronic inflammatory disorders of the gastrointestinal tract, represented by Crohn’s disease (CD) and ulcerative colitis (UC). IBDs represent a healthcare problem globally, with increasing incidence and prevalence in the Western civilization [61]. Although diarrhea and abdominal pain dominate the clinical presentation of patients with CD and UC, there are major differences between the two disorders. In UC the inflammation is limited to the colonic mucosal layer, it involves the rectum in almost all of the cases and it is extended to the proximal part of the colon in a continuous fashion [62]. On the other hand, in CD the inflammation occurs in all the layers of the gut wall and it can involve any part of the gastrointestinal tract. The CD is typically characterized by segments of normal appearing bowel interrupted by areas of disease [63].
The pathogenesis of IBDs is not yet completely understood, however, genetic, environmental, and immunologic factors and an altered gut microbiota have been demonstrated to play key roles in the development of IBDs [63, 64]. Recent studies suggest that IBD is possibly caused by the improper functionality of the immune system, but also by a pronounced dysbiosis accompanied by modifications in its metabolome [65, 66]. Adult patients with CD showed decreased amounts of
When comparing healthy volunteers to patients with IBD, researchers have observed that IBD patients had lower Trp levels in both serum and fecal samples [69, 70].
Experimental animal studies demonstrated that Trp is responsible for the expression of secretory immunoglobulin A (sIgA) and β-defensins [71]. A decreased level of Trp is followed by a production of inflammatory interleukins in the immune cells, such as IL-10 and transforming growth factor-β (TGF-β) [72]. It was demonstrated that IBD has an early onset if a functional deficient mutation in the IL-10 gene is present [73].
IBDs are known to be characterized by a pro-inflammatory state with increased levels of pro-inflammatory cytokines, such as tumor necrosis factor-α (TNF-α), interleukin-12 (IL-12), interleukin-17 (IL-17), and IFN-gamma [74]. These cytokines can enhance the kynurenine pathway by inducing IDO expression, thus leading to higher levels of Trp metabolites [31]. A meta-analysis of the transcriptome and a systematic review of the metabolome demonstrated that patients with IBD exhibit decreased Trp absorption, enhanced kynurenine pathway, altered indole pathway along, and altered indole pathway with decreased Trp plasma levels. In addition, the authors identified increased expression levels of aryl hydrocarbon receptor (AhR), which is a ligand-activated transcription factor [75]. AhR plays a key role in maintaining gut homeostasis (Figure 2). The activated Trp-AhR pathway modulates cell renewal and turnover, induces expression of IL-10 receptors and tight junctions in intestinal epithelial cells, thus promoting intestinal barrier integrity and helping to regulate tissue regeneration [76].
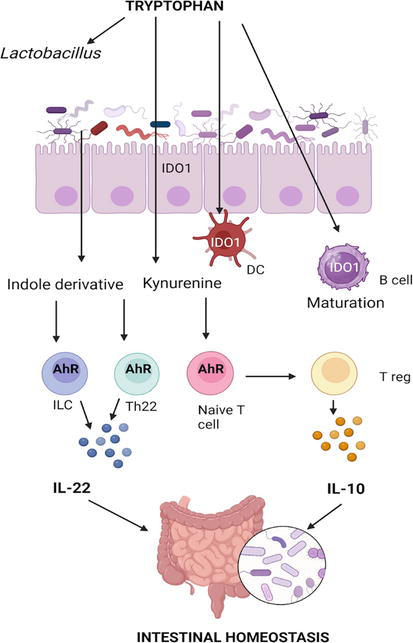
Figure 2.
The role of tryptophan in intestinal homeostasis. Kynurenine and indole derivatives (indole-3-aldehyde, and indole-3-acetic acid) bind to AhR on immune cells and stimulate the production of Il-10 and IL-22. AhR- aryl hydrocarbon receptor; DC - dendritic cell; IDO1 - Indoleamine 2,3 dioxygenase-1; ILC - innate lymphoid cells. Created with BioRender.
Several Trp metabolites, such as kynurenine, 5-HT, XA, and KA have the capacity to bind to AhR and activate it leading to expression of cytokines such as interleukin-22 (IL-22) and IL-17 [22].
One study showed that the Trp metabolites XA and KA have been negatively correlated to inflammation not only in colitis mice models but also in humans with IBD. Lower XA and KA levels have been linked to clinical, biological, and endoscopic markers of disease activity and to a higher expression of pro-inflammatory cytokines. The authors demonstrated that XA and KA have an anti-inflammatory role and that increasing XA and KA levels could represent a promising new therapeutic approach for IBD patients [77].
Moreover, the gut microbiota plays a key role in the kynurenine pathway metabolism and IBD pathophysiology due to its ability to metabolize Trp via the indole pathway. In other words, dysbiosis can alter the kynurenine pathway and promote inflammation in IBD [78]. Recent studies have identified a number of substances that could regulate Trp metabolism. Ginsenoside Rg1, a natural compound, showed promising results in colitis model mice, alleviating inflammation by regulating several microbiota metabolic pathways, including reducing the levels of Trp metabolites [79]. Fucose administration increased the levels of Trp-producing
The involvement of the kynurenine pathway in inflammation has been linked to the development and severity of IBD. Thus, targeting different parts of this metabolic pathway could represent a new treatment strategy for treating IBD. Diets rich in foods that contain Trp and dietary supplements containing Trp can be beneficial since there are studies demonstrating Trp rich diets are associated with a decrease in inflammation, while Trp deficient diets have been linked to exacerbation of colitis [83, 84].
3.3 Celiac disease
CeD is an immune-mediated inflammatory disease of the small intestine caused by sensitivity to dietary gluten affecting genetically predisposed individuals. The global prevalence of CeD is estimated to be around 1% based on serologic testing [85, 86]. Epidemiological studies have reported a prevalence of around 1:70 to 1:300 when biopsy verification was added to serologic tests [87]. Both environmental (dietary gluten) and genetic factors contribute to the pathogenesis of CeD. The high frequency of intrafamilial incidence and the strikingly close association with the human leukocyte antigen (HLA) DR3-DQ2 and/or DR4-DQ8 gene locus provide evidence for the genetic basis of CeD [88, 89]. As early as 1964, the association between CeD and altered Trp metabolism was demonstrated by measuring the urinary levels of several Trp metabolites. The authors reported increased urinary levels of Trp metabolites and low vitamin B6 levels [90]. The role of the gut microbiota was also demonstrated in both earlier and more recent studies and increased abundance of some bacterial strains such as
Dysbiosis is apparently responsible for CeD with the onset in adult life and for the cases that do not respond well to a gluten-free diet [98, 99].
Recent studies have demonstrated that the gut microbiota of CeD patients has a reduced capacity of converting Trp into metabolites capable of activating the AhR, leading to intestinal inflammation [97, 98]. The correlation between decreased AhR activity and CeD is supported by several authors [99, 100, 101, 102, 103]. One of the mechanisms by which beneficial bacteria could alleviate CeD is by modulating the AhR pathway. Studies on rodent models demonstrated that treating mice with lactobacilli strains known to produce AhR ligands such as
4. The relationship between Trp metabolites, related gastrointestinal disease, and weight loss
Serotonin, a neurotransmitter derived from Trp, plays a role in regulating mood and appetite. However, the relationship between serotonin levels and weight loss is complex and not fully understood. While serotonin itself is not directly linked to weight loss, its influence on appetite and mood can indirectly impact weight management [18].
The relationship between serotonin and weight is complex and can involve various factors. Serotonin is known to contribute to feelings of satiety and fullness [104, 105]. Low serotonin levels have been associated with an increased appetite and cravings for carbohydrate-rich foods [106]. Some individuals may engage in emotional eating to boost serotonin levels, especially during periods of stress or low mood [107].
Changes in serotonin levels can influence mood, and mood disturbances may affect eating habits. For example, people with depression may experience changes in appetite, leading to weight gain or loss [108]. Certain medications, such as selective serotonin reuptake inhibitors (SSRIs), which are commonly prescribed for the treatment of depression and anxiety, can affect serotonin levels. Some individuals may experience weight changes as a side effect of these medications [108].
The link between gastrointestinal diseases and weight loss has been studied from several points of view and will be detailed in the following paragraphs as stated in the literature.
IBD can affect the absorption of nutrients in the intestines [109]. Chronic inflammation and damage to the intestinal lining can impair the body’s ability to absorb essential nutrients, leading to malabsorption. This malabsorption can result in weight loss and nutritional deficiencies [110].
Symptoms of IBD, such as abdominal pain, diarrhea, and nausea, can contribute to a reduced appetite. Individuals with IBD may experience discomfort or pain during or after eating, leading to a decrease in food intake and subsequent weight loss [110].
The chronic inflammation associated with IBD can increase the body’s metabolic rate. This increased demand for energy, coupled with malabsorption, can contribute to weight loss, even when individuals are not intentionally restricting their calorie intake [111].
The inflammation and other factors associated with IBD can lead to changes in body composition, including muscle wasting and loss of fat tissue. This can contribute to overall weight loss and weakness [111]. The severity of IBD symptoms can vary, and individuals may experience periods of flare-ups with more intense symptoms. During flare-ups, the impact on weight loss may be more pronounced [112]. It’s important to note that weight loss in IBD is often unintentional and can be a sign of underlying disease activity [113]. Managing IBD involves a multidisciplinary approach, including medical treatment, nutritional support, and lifestyle modifications.
IBS is different from IBD, and it is essential to distinguish between the two conditions. Unlike IBD (Crohn’s disease and ulcerative colitis), IBS is a functional gastrointestinal disorder characterized by symptoms such as abdominal pain, bloating, and changes in bowel habits without evidence of inflammation or damage to the digestive tract [60]. In general, IBS is not typically associated with significant weight loss. Weight loss is usually not a primary symptom of IBS, and any changes in body weight are more likely to be subtle or unintentional. However, there are a few indirect ways in which IBS might be linked to changes in weight [114].
Individuals with IBS may make changes to their diet to manage symptoms, such as avoiding certain trigger foods. This modification in dietary habits could potentially impact calorie intake and, in some cases, lead to weight changes [115].
Some people with IBS may experience symptoms such as diarrhea or abdominal pain that can affect their appetite or eating patterns. While this might not necessarily lead to significant weight loss, it can contribute to fluctuations in body weight [116]. IBS symptoms are known to be influenced by stress and psychological factors. Stress-related changes in eating patterns or behaviors may indirectly impact weight [117, 118]. It’s important to emphasize that IBS is a chronic condition, and its symptoms can vary widely among individuals. While weight loss is not a primary characteristic of IBS, other gastrointestinal conditions or non-gastrointestinal factors could contribute to changes in body weight.
CeD is an autoimmune disorder characterized by an intolerance to gluten, a protein found in wheat, barley, and rye [119]. The ingestion of gluten triggers an immune response in individuals with CeD, leading to inflammation and damage to the lining of the small intestine. This damage can result in malabsorption of nutrients, affecting various aspects of health, including body weight [119].
The relationship between CeD and weight loss is often observed due to a number of factors. The damage to the small intestine in CeD impairs the absorption of essential nutrients, including proteins, fats, carbohydrates, vitamins, and minerals. This malabsorption can lead to nutrient deficiencies, contributing to weight loss [110].
Individuals with CeD may experience symptoms such as abdominal pain, bloating, diarrhea, and nausea. These symptoms can affect appetite and food intake, leading to reduced caloric consumption and subsequent weight loss. Chronic diarrhea is a common symptom of CeD. Frequent bowel movements can result in fluid loss and contribute to weight loss [120]. The inflammation in the small intestine associated with CeD can increase the body’s metabolic rate, causing an elevated demand for energy. This increased metabolic activity, combined with malabsorption, can lead to weight loss [119]. In severe cases of untreated CeD, the body may start to break down muscle tissue for energy, contributing to muscle wasting and additional weight loss [121].
It’s important to note that while weight loss is a common symptom of CeD, not everyone with the condition experiences this. Some individuals with CeD may have atypical or minimal symptoms, making diagnosis challenging.
The treatment for CeD involves strict adherence to a gluten-free diet, which typically leads to the resolution of symptoms and the restoration of a healthy body weight. Early detection and adherence to a gluten-free diet can help manage symptoms and prevent complications associated with CeD [122].
5. Future perspectives
To date, several studies focused on the importance of the gut microbiota, this “virtual symbiotic organ” that clearly interferes with many metabolic and immune functions in our organism. The human health and disease state are depending on the microbial metabolites resulted by host-microorganism or microorganism – microorganism interactions at gut level [107]. The complete understanding of the gut microbiota complexity and the possibility to manipulate the microbes as future therapeutic strategy will open new perspectives regarding our understanding of gastrointestinal and psychiatric pathologies. An appealing approach for the patients associating depression or anxiety that usually accompany IBS, IBD, or CeD is provided by the administration of probiotics and/or prebiotics [108]. Some other diseases involving the gut-microbiota-brain axis, such as anorexia nervosa apparently produced by changes in dietary intake and pre-morbid gut microbiota changes might benefit of the use of probiotics [109]. We mentioned here anorexia nervosa because there are many similarities between this eating disorder and the above-mentioned gastrointestinal diseases and mental health disorders [110].
The probiotics rich in
Acronyms and abbreviations
anthranilic acid | |
aromatic L-amino decarboxylase, | |
aldehyde dehydrogenase | |
aryl hydrocarbon receptor | |
Crohn’s disease | |
celiac disease | |
central nervous system | |
enteric nervous system | |
anholocyclic acid | |
indole-3-acetaldehyde | |
indole-3-aldehyde | |
inflammatory bowel disease | |
irritable bowel syndrome | |
indoleamine 2,3-dioxygenase | |
interleukin-8 | |
interleukin-12 | |
interleukin-17 | |
indole-3-lactic acid | |
indole-3-propionic acid | |
indole-3-pyruvic acid | |
kynurenic acid | |
kynurenine-3-monooxygenase | |
monoamine oxidase | |
nicotinamide adenine dinucleotide | |
N-methyl-D-aspartate | |
quinolinic acid | |
immunoglobulin A | |
tryptophan 2,3-dioxygenase | |
transforming growth factor-β | |
necrosis factor-α | |
tryptophan hydroxylase | |
ulcerative colitis | |
xanthurenic acid | |
3-hydroxyanthranilic acid | |
3-hydroxykynurenine | |
5-hydroxyindoleacetic acid | |
5-hydroxytryptamine | |
5-hydroxytryptophan |
References
- 1.
Xue C et al. Tryptophan metabolism in health and disease. Cell Metabolism. 2023; 35 (8):1304-1326 - 2.
Li G, Young KD. Indole production by the tryptophanase TnaA in Escherichia coli is determined by the amount of exogenous tryptophan. Microbiology (Reading). 2013;159 (Pt 2):402-410 - 3.
Friedman M. Analysis, nutrition, and health benefits of tryptophan. International Journal of Tryptophan Research. 2018; 11 :1178646918802282 - 4.
Liu M et al. Microbial tryptophan metabolism tunes host immunity, metabolism, and extraintestinal disorders. Metabolites. 2022; 12 (9):834 - 5.
Roth W et al. Tryptophan metabolism and gut-brain homeostasis. International Journal of Molecular Sciences. 2021; 22 (6):2973 - 6.
Ciorba MA. Kynurenine pathway metabolites: Relevant to vitamin B-6 deficiency and beyond. The American Journal of Clinical Nutrition. 2013; 98 :863-864 - 7.
Li S. Modulation of immunity by tryptophan microbial metabolites. Frontiers in Nutrition. 2023; 10 :1209613 - 8.
Gao K et al. Increasing carbohydrate availability in the hindgut promotes hypothalamic neurotransmitter synthesis: Aromatic amino acids linking the microbiota-brain axis. Journal of Neurochemistry. 2019; 149 (5):641-659 - 9.
Sherwin E, Dinan TG, Cryan JF. Recent developments in understanding the role of the gut microbiota in brain health and disease. Annals of the New York Academy of Sciences. 2018; 1420 (1):5-25 - 10.
Fröhlich EE et al. Cognitive impairment by antibiotic-induced gut dysbiosis: Analysis of gut microbiota-brain communication. Brain, Behavior, and Immunity. 2016; 56 :140-155 - 11.
De Vadder F et al. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell. 2014; 156 (1-2):84-96 - 12.
Bianconi E et al. An estimation of the number of cells in the human body. Annals of Human Biology. 2013; 40 (6):463-471 - 13.
Sender R, Fuchs S, Milo R. Are we really vastly outnumbered? Revisiting the ratio of bacterial to host cells in humans. Cell. 2016; 164 (3):337-340 - 14.
Clarke G et al. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Molecular Psychiatry. 2013; 18 (6):666-673 - 15.
Ozogul F, Kuley E, Ozogul Y, Ozogul I. The function of lactic acid bacteria on biogenic amines production by food-borne pathogens in arginine decarboxylase broth. Food Science and Technology Research. 2012; 18 (6):795-804 - 16.
Ma J, Kobayashi DY, Yee N. Role of menaquinone biosynthesis genes in selenate reduction by enterobacter cloacae SLD1a-1 and Escherichia coli K12. Environmental Microbiology. 2009;11 (1):149-158 - 17.
Clark A, Mach N. Exercise-induced stress behavior, gut-microbiota-brain axis and diet: A systematic review for athletes. Journal of the International Society of Sports Nutrition. 2016; 13 :43 - 18.
Jenkins TA et al. Influence of tryptophan and serotonin on mood and cognition with a possible role of the gut-brain Axis. Nutrients. 2016; 8 (1):56 - 19.
Ratcliffe EM, Farrar NR, Fox EA. Development of the vagal innervation of the gut: Steering the wandering nerve. Neurogastroenterology and Motility. 2011; 23 (10):898-911 - 20.
De Vadder F et al. Gut microbiota regulates maturation of the adult enteric nervous system via enteric serotonin networks. Proceedings of the National Academy of Sciences of the United States of America. 2018; 115 (25):6458-6463 - 21.
Rhee SH, Pothoulakis C, Mayer EA. Principles and clinical implications of the brain-gut-enteric microbiota axis. Nature Reviews. Gastroenterology & Hepatology. 2009; 6 (5):306-314 - 22.
Sun M et al. Tryptophan (Trp) modulates gut homeostasis via aryl hydrocarbon receptor (AhR). Critical Reviews in Food Science and Nutrition. 2020; 60 (10):1760-1768 - 23.
Ehrlich AM et al. Indole-3-lactic acid associated with bifidobacterium-dominated microbiota significantly decreases inflammation in intestinal epithelial cells. BMC Microbiology. 2020; 20 (1):357 - 24.
Meng D et al. Indole-3-lactic acid, a metabolite of tryptophan, secreted by Bifidobacterium longum subspecies infantis is anti-inflammatory in the immature intestine. Pediatric Research. 2020;88 (2):209-217 - 25.
Labus JS et al. Differences in gut microbial composition correlate with regional brain volumes in irritable bowel syndrome. Microbiome. 2017; 5 (1):49 - 26.
Pinto-Sanchez MI et al. Probiotic Bifidobacterium longum NCC3001 reduces depression scores and alters brain activity: A pilot study in patients with irritable bowel syndrome. Gastroenterology. 2017;153 (2):448-459.e8 - 27.
Strati F et al. New evidences on the altered gut microbiota in autism spectrum disorders. Microbiome. 2017; 5 (1):24 - 28.
Jiang H et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain, Behavior, and Immunity. 2015; 48 :186-194 - 29.
Höglund E, Øverli Ø, Winberg S. Tryptophan metabolic pathways and brain serotonergic activity: A comparative review. Frontiers in Endocrinology (Lausanne). 2019; 10 :158 - 30.
Badawy AA. Tryptophan metabolism: A versatile area providing multiple targets for pharmacological intervention. Egyptian Journal of Basic and Clinical Pharmacology. 2019; 9 - 31.
Sheibani M et al. Kynurenine pathway and its role in neurologic, psychiatric, and inflammatory bowel diseases. Molecular Biology Reports. 2023; 50 (12):10409-10425 - 32.
Takikawa O. Biochemical and medical aspects of the indoleamine 2,3-dioxygenase-initiated L-tryptophan metabolism. Biochemical and Biophysical Research Communications. 2005; 338 (1):12-19 - 33.
O’Farrell K, Harkin A. Stress-related regulation of the kynurenine pathway: Relevance to neuropsychiatric and degenerative disorders. Neuropharmacology. 2017; 112 (Pt B):307-323 - 34.
Dantzer R et al. From inflammation to sickness and depression: When the immune system subjugates the brain. Nature Reviews. Neuroscience. 2008; 9 (1):46-56 - 35.
Maes M et al. The inflammatory & neurodegenerative (I & ND) hypothesis of depression: Leads for future research and new drug developments in depression. Metabolic Brain Disease. 2009; 24 (1):27-53 - 36.
Okuda S et al. 3-hydroxykynurenine, an endogenous oxidative stress generator, causes neuronal cell death with apoptotic features and region selectivity. Journal of Neurochemistry. 1998; 70 (1):299-307 - 37.
Stone TW, Darlington LG. Endogenous kynurenines as targets for drug discovery and development. Nature Reviews. Drug Discovery. 2002; 1 (8):609-620 - 38.
Irwin MR, Miller AH. Depressive disorders and immunity: 20 years of progress and discovery. Brain, Behavior, and Immunity. 2007; 21 (4):374-383 - 39.
Schwarcz R. The kynurenine pathway of tryptophan degradation as a drug target. Current Opinion in Pharmacology. 2004; 4 (1):12-17 - 40.
Reyes Ocampo J et al. Kynurenines with neuroactive and redox properties: Relevance to aging and brain diseases. Oxidative Medicine and Cellular Longevity. 2014; 2014 :646909 - 41.
Roager HM, Licht TR. Microbial tryptophan catabolites in health and disease. Nature Communications. 2018; 9 (1):3294 - 42.
Su X, Gao Y, Yang R. Gut microbiota-derived tryptophan metabolites maintain gut and systemic homeostasis. Cells. 2022; 11 (15):2296 - 43.
Williams BB et al. Discovery and characterization of gut microbiota decarboxylases that can produce the neurotransmitter tryptamine. Cell Host & Microbe. 2014; 16 (4):495-503 - 44.
Gaspar B et al. The intestinal microbiome in humans: It's role for a healthy life and in the onset of disease. In: MIcrobiome - the Key for Human Health. London, UK: IntechOpen; 2024 - 45.
Drossman DA et al. AGA technical review on irritable bowel syndrome. Gastroenterology. 2002; 123 (6):2108-2131 - 46.
Cryan JF et al. The microbiota-gut-brain axis. Physiological Reviews. 2019; 99 (4):1877-2013 - 47.
Agus A, Planchais J, Sokol H. Gut microbiota regulation of tryptophan metabolism in health and disease. Cell Host & Microbe. 2018; 23 (6):716-724 - 48.
Arnold W. Pathophysiology of irritable bowel syndrome. 2023. Available from: https://www.uptodate.com/contents/pathophysiology-of-irritable-bowel-syndrome?search=irritable%20bowel%20syndrome&source=search_result&selectedTitle=3~150&usage_type=default&display_rank=3#H1 - 49.
Buéno L, Fioramonti J, Garcia-Villar R. Pathobiology of visceral pain: Molecular mechanisms and therapeutic implications. III. Visceral afferent pathways: A source of new therapeutic targets for abdominal pain. American Journal of Physiology. Gastrointestinal and Liver Physiology. 2000; 278 (5):G670-G676 - 50.
Faure C et al. Serotonin signaling is altered in irritable bowel syndrome with diarrhea but not in functional dyspepsia in pediatric age patients. Gastroenterology. 2010; 139 (1):249-258 - 51.
Willert RP et al. The development and maintenance of human visceral pain hypersensitivity is dependent on the N-methyl-D-aspartate receptor. Gastroenterology. 2004; 126 (3):683-692 - 52.
Fitzgerald P et al. Tryptophan catabolism in females with irritable bowel syndrome: Relationship to interferon-gamma, severity of symptoms and psychiatric co-morbidity. Neurogastroenterology and Motility. 2008; 20 (12):1291-1297 - 53.
Han L et al. Altered metabolome and microbiome features provide clues in understanding irritable bowel syndrome and depression comorbidity. The ISME Journal. 2022; 16 (4):983-996 - 54.
Moskwa A et al. Serum serotonin concentration and urine 5-hydroxyindole acetic acid excretion in patients with irritable bowel syndrome. Polski Merkuriusz Lekarski. 2007; 22 (131):366-368 - 55.
Atkinson W et al. Altered 5-hydroxytryptamine signaling in patients with constipation- and diarrhea-predominant irritable bowel syndrome. Gastroenterology. 2006; 130 (1):34-43 - 56.
Thijssen AY et al. Alterations in serotonin metabolism in the irritable bowel syndrome. Alimentary Pharmacology & Therapeutics. 2016; 43 (2):272-282 - 57.
Gershon MD, Tack J. The serotonin signaling system: From basic understanding to drug development for functional GI disorders. Gastroenterology. 2007; 132 (1):397-414 - 58.
Mujagic Z et al. Integrated fecal microbiome-metabolome signatures reflect stress and serotonin metabolism in irritable bowel syndrome. Gut Microbes. 2022; 14 (1):2063016 - 59.
Foley S et al. Impaired uptake of serotonin by platelets from patients with irritable bowel syndrome correlates with duodenal immune activation. Gastroenterology. 2011; 140 (5):1434-43.e1 - 60.
Saha L. Irritable bowel syndrome: Pathogenesis, diagnosis, treatment, and evidence-based medicine. World Journal of Gastroenterology. 2014; 20 (22):6759-6773 - 61.
M’Koma AE. Inflammatory bowel disease: An expanding global health problem. Clinical Medicine Insights. Gastroenterology. 2013; 6 :33-47 - 62.
Iancu MA et al. Revisiting the intestinal microbiome and its role in diarrhea and constipation. Microorganisms. 2023; 11 (9):2177 - 63.
Ruxandra Florentina I et al. Intestinal Microbiomics in physiological and pathological conditions. In: Vasudeo Z et al., editors. Advances in Probiotics for Health and Nutrition. London, UK, Rijeka: IntechOpen; 2023. p. Ch. 6 - 64.
Zhang YZ, Li YY. Inflammatory bowel disease: Pathogenesis. World Journal of Gastroenterology. 2014; 20 (1):91-99 - 65.
Ni J et al. Gut microbiota and IBD: Causation or correlation? Nature Reviews. Gastroenterology & Hepatology. 2017; 14 (10):573-584 - 66.
Imhann F et al. Interplay of host genetics and gut microbiota underlying the onset and clinical presentation of inflammatory bowel disease. Gut. 2018; 67 (1):108-119 - 67.
Barnich N, Denizot J, Darfeuille-Michaud A. E. coli -mediated gut inflammation in genetically predisposed Crohn's disease patients. Pathologie Biologie (Paris). 2013;61 (5):e65-e69 - 68.
Walker AW et al. High-throughput clone library analysis of the mucosa-associated microbiota reveals dysbiosis and differences between inflamed and non-inflamed regions of the intestine in inflammatory bowel disease. BMC Microbiology. 2011; 11 :7 - 69.
Lamas B et al. CARD9 impacts colitis by altering gut microbiota metabolism of tryptophan into aryl hydrocarbon receptor ligands. Nature Medicine. 2016; 22 (6):598-605 - 70.
Shiomi Y et al. GCMS-based metabolomic study in mice with colitis induced by dextran sulfate sodium. Inflammatory Bowel Diseases. 2011; 17 (11):2261-2274 - 71.
Liang H et al. Dietary l-tryptophan supplementation enhances the intestinal mucosal barrier function in weaned piglets: Implication of tryptophan-metabolizing microbiota. International Journal of Molecular Sciences. 2018; 20 (1):20 - 72.
Sorgdrager FJH et al. Tryptophan metabolism in inflammaging: From biomarker to therapeutic target. Frontiers in Immunology. 2019; 10 :2565 - 73.
Neurath MF. Cytokines in inflammatory bowel disease. Nature Reviews. Immunology. 2014; 14 (5):329-342 - 74.
Sanchez-Munoz F, Dominguez-Lopez A, Yamamoto-Furusho JK. Role of cytokines in inflammatory bowel disease. World Journal of Gastroenterology. 2008; 14 (27):4280-4288 - 75.
Wang S et al. The involvement of intestinal tryptophan metabolism in inflammatory bowel disease identified by a meta-analysis of the transcriptome and a systematic review of the metabolome. Nutrients. 2023; 15 (13):2886 - 76.
Rothhammer V, Quintana FJ. The aryl hydrocarbon receptor: An environmental sensor integrating immune responses in health and disease. Nature Reviews. Immunology. 2019; 19 (3):184-197 - 77.
Michaudel C et al. Rewiring the altered tryptophan metabolism as a novel therapeutic strategy in inflammatory bowel diseases. Gut. 2023; 72 (7):1296-1307 - 78.
O’Mahony SM et al. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behavioural Brain Research. 2015; 277 :32-48 - 79.
Cheng H et al. Ginsenoside Rg1 alleviates acute ulcerative colitis by modulating gut microbiota and microbial tryptophan metabolism. Frontiers in Immunology. 2022; 13 :817600 - 80.
Borisova MA et al. Fucose ameliorates tryptophan metabolism and behavioral abnormalities in a mouse model of chronic colitis. Nutrients. 2020; 12 (2):445 - 81.
Yan X et al. Fructooligosaccharides protect against OVA-induced food allergy in mice by regulating the Th17/Treg cell balance using tryptophan metabolites. Food & Function. 2021; 12 (7):3191-3205 - 82.
Yang W et al. Fuzhuan brick tea polysaccharide improved ulcerative colitis in association with gut microbiota-derived tryptophan metabolism. Journal of Agricultural and Food Chemistry. 2021; 69 (30):8448-8459 - 83.
Islam J et al. Dietary tryptophan alleviates dextran sodium sulfate-induced colitis through aryl hydrocarbon receptor in mice. The Journal of Nutritional Biochemistry. 2017; 42 :43-50 - 84.
Lanis JM et al. Tryptophan metabolite activation of the aryl hydrocarbon receptor regulates IL-10 receptor expression on intestinal epithelia. Mucosal Immunology. 2017; 10 (5):1133-1144 - 85.
Rubio-Tapia A et al. The prevalence of celiac disease in the United States. The American Journal of Gastroenterology. 2012; 107 (10):1538-1544; quiz 1537, 1545 - 86.
Choung RS et al. Prevalence and morbidity of undiagnosed celiac disease from a community-based study. Gastroenterology. 2017; 152 (4):830-839.e5 - 87.
Gujral N, Freeman HJ, Thomson AB. Celiac disease: Prevalence, diagnosis, pathogenesis and treatment. World Journal of Gastroenterology. 2012; 18 (42):6036-6059 - 88.
Liu E et al. Risk of pediatric celiac disease according to HLA haplotype and country. The New England Journal of Medicine. 2014; 371 (1):42-49 - 89.
Pietzak MM et al. Stratifying risk for celiac disease in a large at-risk United States population by using HLA alleles. Clinical Gastroenterology and Hepatology. 2009; 7 (9):966-971 - 90.
Kowlessar OD, Haeffner LJ, Benson GD. Abnormal tryptophan metabolism in patients with adult celiac disease, with evidence for deficiency of vitamin B6. The Journal of Clinical Investigation. 1964; 43 (5):894-903 - 91.
Stĕpánková R et al. Changes in jejunal mucosa after long-term feeding of germfree rats with gluten. Scandinavian Journal of Gastroenterology. 1996; 31 (6):551-557 - 92.
Galipeau HJ et al. Intestinal microbiota modulates gluten-induced immunopathology in humanized mice. The American Journal of Pathology. 2015; 185 (11):2969-2982 - 93.
Ashorn S et al. Elevated serum anti-Saccharomyces cerevisiae, anti-I2 and anti-OmpW antibody levels in patients with suspicion of celiac disease. Journal of Clinical Immunology. 2008; 28 (5):486-494 - 94.
D’Argenio V et al. Metagenomics reveals dysbiosis and a potentially pathogenic N. flavescens strain in duodenum of adult celiac patients. The American Journal of Gastroenterology. 2016;111 (6):879-890 - 95.
Galipeau HJ, Verdu EF. The double-edged sword of gut bacteria in celiac disease and implications for therapeutic potential. Mucosal Immunology. 2022; 15 (2):235-243 - 96.
Nadal I et al. Imbalance in the composition of the duodenal microbiota of children with coeliac disease. Journal of Medical Microbiology. 2007; 56 (Pt 12):1669-1674 - 97.
Natividad JM et al. Impaired aryl hydrocarbon receptor ligand production by the gut microbiota is a key factor in metabolic syndrome. Cell Metabolism. 2018; 28 (5):737-749.e4 - 98.
Lamas B et al. Aryl hydrocarbon receptor ligand production by the gut microbiota is decreased in celiac disease leading to intestinal inflammation. Science Translational Medicine. 2020; 12 (566):eaba0624 - 99.
Dinallo V et al. Protective effects of aryl hydrocarbon receptor signaling in celiac disease mucosa and in poly I:C-induced small intestinal atrophy mouse model. Frontiers in Immunology. 2019; 10 :91 - 100.
Xu H et al. Gluten-sensitive enteropathy coincides with decreased capability of intestinal T cells to secrete IL-17 and IL-22 in a macaque model for celiac disease. Clinical Immunology. 2013; 147 (1):40-49 - 101.
Bodd M et al. HLA-DQ2-restricted gluten-reactive T cells produce IL-21 but not IL-17 or IL-22. Mucosal Immunology. 2010; 3 (6):594-601 - 102.
Li Y et al. Exogenous stimuli maintain intraepithelial lymphocytes via aryl hydrocarbon receptor activation. Cell. 2011; 147 (3):629-640 - 103.
Suwanai H et al. A defective Il15 allele underlies the deficiency in natural killer cell activity in nonobese diabetic mice. Proceedings of the National Academy of Sciences of the United States of America. 2010; 107 (20):9305-9310 - 104.
van Galen KA, Ter Horst KW, Serlie MJ. Serotonin, food intake, and obesity. Obesity Reviews. 2021; 22 (7):e13210 - 105.
Voigt JP, Fink H. Serotonin controlling feeding and satiety. Behavioural Brain Research. 2015; 277 :14-31 - 106.
Wurtman RJ, Wurtman JJ. Brain serotonin, carbohydrate-craving, obesity and depression. Obesity Research. 1995; 3 (Suppl 4):477s-480s - 107.
Grajek M et al. Prevalence of emotional eating in groups of students with varied diets and physical activity in Poland. Nutrients. 2022; 14 (16):3289 - 108.
Felton J et al. The relation of weight change to depressive symptoms in adolescence. Development and Psychopathology. 2010; 22 (1):205-216 - 109.
Balestrieri P et al. Nutritional aspects in inflammatory bowel diseases. Nutrients. 2020; 12 (2):372 - 110.
Montoro-Huguet MA, Belloc B, Domínguez-Cajal M. Small and large intestine (I): Malabsorption of nutrients. Nutrients. 2021; 13 (4):1254 - 111.
Roncoroni L et al. Nutrition in patients with inflammatory bowel diseases: A narrative review. Nutrients. 2022; 14 (4):751 - 112.
Sun Y et al. Stress triggers flare of inflammatory bowel disease in children and adults. Frontiers in Pediatrics. 2019; 7 :432 - 113.
Elsherif Y, Alexakis C, Mendall M. Determinants of weight loss prior to diagnosis in inflammatory bowel disease: A retrospective observational study. Gastroenterology Research and Practice. 2014; 2014 :762191 - 114.
Pickett-Blakely O. Obesity and irritable bowel syndrome: A comprehensive review. Journal of Gastroenterology and Hepatology (N Y). 2014; 10 (7):411-416 - 115.
Guo YB et al. Association between diet and lifestyle habits and irritable bowel syndrome: A case-control study. Gut Liver. 2015; 9 (5):649-656 - 116.
Harer KN. Irritable bowel syndrome, disordered eating, and eating disorders. Journal of Gastroenterology and Hepatology (N Y). 2019; 15 (5):280-282 - 117.
Hayes PA, Fraher MH, Quigley EM. Irritable bowel syndrome: The role of food in pathogenesis and management. Journal of Gastroenterology and Hepatology (N Y). 2014; 10 (3):164-174 - 118.
Qin HY et al. Impact of psychological stress on irritable bowel syndrome. World Journal of Gastroenterology. 2014; 20 (39):14126-14131 - 119.
Sharma N et al. Pathogenesis of celiac disease and other gluten related disorders in wheat and strategies for mitigating them. Frontiers in Nutrition. 2020; 7 :6 - 120.
Sabença C et al. Wheat/gluten-related disorders and gluten-free diet misconceptions: A review. Food. 2021; 10 (8):1765 - 121.
De Risi LL. Starving in the midst of plenty: Adult celiac disease. The American Journal of Nursing. 1970; 70 (5):1048-1053 - 122.
Simón E et al. The gluten-free diet for celiac disease: Critical insights to better understand clinical outcomes. Nutrients. 2023; 15 (18):4013