
Abstract
The latest advances in basic and clinical research on the main components of the platelet serotonergic system are presently reviewed. These components consist of serotonin (5-HT), enzymes that participate in 5-HT metabolism, the serotonin transporter (SERT), and 5-HT1A, 5-HT2A, 5-HT3, and 5-HT4 receptors (each with their corresponding mechanism of intracellular transduction). An additional focus is on related biomarkers or drugs for the diagnosis or treatment of the pathophysiology of diverse disorders such as depression, anxiety, hemorrhagic dengue, coagulopathy generated by COVID-19, myocardial infarction, and preeclampsia. The drugs analyzed include serotonin reuptake inhibitors and serotonergic drugs that act on 5-HT receptors. Through the platelet serotonergic system, serotonergic drugs not only interact with the central nervous system but also may participate in coagulation, vascular permeability, and peripheral vascular resistance, which has many implications. Finally, perspectives are offered for future research on biomarkers and new therapeutic targets.
Keywords
- platelets
- serotonin
- pathophysiology
- serotonin transporter
- 5-HT1A
- 5-HT2A
1. Introduction
Scientific advances in biomedical research have allowed for the discovery of numerous factors in the human organism related to ligand-receptor interactions and intracellular signaling mechanisms. The new insights have provided a deeper comprehension of the interrelationship of diverse complex physiological systems on a molecular and systemic level. For example, recent studies on the platelet serotonergic system have created new opportunities based on a better understanding the pathophysiology of severe dengue, coagulopathy associated with COVID-19, myocardial infarction, preeclampsia, and other diseases.
Platelets are anucleate blood cells with a biconvex discoid structure and a diameter of 2–4 μm. They develop from megakaryocytes of the bone marrow and are released into the bloodstream through projections of pseudopodia known as proplatelets [1]. Each megakaryocyte is able to produce ~1000 to 3000 platelets as a result of multiple divisions [2]. Platelets contain 5–8 mitochondria that synthesize various proteins involved in platelet activation and apoptosis [3]. Unactivated platelet cells have an average circulation lifespan of 8–10 days before being eliminated by the spleen. People generally have 150–400 × 109 platelets per liter of peripheral blood [1].
A key molecule of the platelet serotonergic system is serotonin (5-HT), which belongs to the family of indolamines. It is composed of an indole ring hydroxylated at position 5 and an ethylamine lateral chain [4]. Apart from 5-HT, other important components of the platelet serotonergic system are the serotonin transporter (SERT) responsible for the recapture of 5-HT, a broad range of serotonin membrane receptors, and enzymes that participate in 5-HT metabolism [5]. The elements of the platelet serotonergic system are located in the central nervous system (CNS) [6] enterochromaffin cells, and diverse tissues (e.g., pancreatic, pulmonary, hepatic, and hematopoietic) [7, 8, 9]. This composition of the serotonergic system regulates most of the physiological functions of the organism that have a role in the maintenance of homeostasis.
At the beginning of the 20th century, platelets were thought to be the only source of 5-HT in mammals. About 100 years ago, Dr. Vittorio Erspamer was looking for substances capable of causing the contraction of smooth muscle. In 1930, he identified such a compound, which he called enteramine, in the gastric mucosa of rabbits. Almost two decades later (in 1948), a vasoconstrictive substance, denominated serotonin, was isolated from human serum in the laboratory of Dr. Irving Page. The structure of serotonin was established as 5-HT in 1949, and in 1952 it was confirmed that enteramine and serotonin are the same compound [7].
Regarding the biosynthesis of 5-HT, the first step is the conversion of L-tryptophan (L-Trp) to 5-hydroxytryptophan (5-HTP), catalyzed by the rate-limiting enzyme tryptophan hydroxylase. Subsequently, 5-HTP is transformed into 5-HT by another enzyme, aromatic L-amino acid decarboxylase (4). Additionally, L-Trp can be metabolized in the kynurenine pathway to nicotinamide, a fundamental component in the formation of two molecules involved in important redox reactions in cell metabolism: nicotinamide adenine dinucleotide (NAD+) and nicotinamide adenine dinucleotide phosphate (NADP+). The majority of 5-HT in the human organism is produced and stored in the gastrointestinal tract (Figure 2). From this location, it enters the bloodstream and by means of SERT is rapidly internalized into platelets [5, 10] (5), then stored in the dense granules of these cells. Only a small fraction of 5-HT (<1%) remains free in plasma [11].
The 5-HT secreted into plasma by platelets binds to many different membrane receptors, which are classified into seven families designated as 5-HT1–7. There are distinct subtypes for each family, denoted by adding a letter to the name (e.g., 5-HT1A, or 5-HT receptor type 1, subtype A) [12, 13]. Among the subtypes of 5-HT receptors (5-HTRs) on the platelet membrane, those most commonly studied are 5-HT1A [14, 15], 5-HT2A [16], 5-HT3 [17] and 5-HT4 [18]. They will hereafter be discussed along with their ligands and mechanisms of signal transduction.
Platelets participate in the defense against pathogens [19] as well as in the exacerbation of autoimmune diseases [20, 21]. For example, in the presence of certain infectious agents or inflammatory stimuli, activated platelets trigger the innate and adaptive immune response by releasing granules containing 5-HT, β-defensin [22, 23] prostaglandins, and other inflammatory mediators. 5-HT released into plasma stimulates receptors with the capacity to promote vasoconstriction and therefore increase peripheral vascular resistance [24], which is characteristic of a broad range of hypertensive pathologies [25, 26].
In summary, platelets may be regarded as circulating neurons with a complex communication network that enables them to interact with cells in many types of tissues to regulate diverse physiological processes. Hence the platelet serotonergic system acts as an axis of autocrine and paracrine communication [27] capable of mediating inflammation, blood pressure, and the immune response.
These insights into the platelet serotonergic system imply advantages, disadvantages, and challenges in relation to medical applications. Considering the platelet serotonergic system as the common denominator for the CNS and diverse peripheral tissues (e.g., in the gastrointestinal tract, the hematopoietic system, and the cardiovascular apparatus), platelets can be viewed as “circulating mirrors” of neurons and innate immune cells [28, 29]. That is, the activity of the platelet serotonergic system appears to be a reflection of what occurs in peripheral tissues during various pathological processes: neuropsychological disorders (e.g., stress, depression, anxiety, and epilepsy), coagulopathies associated with infectious diseases [e.g., dengue [30] and COVID-19 [31], and hypertensive disorders such as preeclampsia [24] and acute myocardial infarction [32]. For instance, the aforementioned neuropsychological disorders involve increased permeability of the blood-brain barrier, which gives rise to a greater filtration of platelets towards the CNS. In the brain, platelets release 5-HT and other chemical mediators, exacerbating neural electrical activity. In cases of epilepsy, this release of 5-HT could increase the severity of convulsions [33]. Thus, compounds capable of slowing the course of platelet activation might be useful for treating epilepsy. A deeper understanding of the platelet serotonergic system should allow for the development of new biomarkers for diagnosis and new drugs (e.g., agonists or antagonists of 5-HTRs) with enhanced therapeutic activity.
Regarding the disadvantages of the close relation between the role of the platelet serotonergic system in the CNS and peripheral tissues, drugs targeting the brain to treat anxiety, depression, and epilepsy might also act on the periphery and cause adverse effects. For example, platelet function can be affected by serotonin reuptake inhibitors (SRIs) such as fluoxetine [34] and valproic acid. The latter is an agonist of 5-HT2A used to treat epilepsy [35] and may under certain conditions produce thrombocytopenia [36].
A challenge for future research is the design and development of drugs able to interact selectively with the serotonergic system in the CNS or peripheral tissues. Moreover, it is crucial to gain insights into the dynamics of platelet activity leading to the release of 5-HT into plasma in order to better comprehend the kinetics of 5-HT in the pathophysiology of severe dengue, high blood pressure, preeclampsia, and acute myocardial infarction.
2. Transport and receptors of 5-HT in platelets
SERT, a member of the family of transporters of solutes dependent on Na+/Cl6 (the solute carrier 6, or SLC6), controls intra- and extracellular concentrations of 5-HT [37, 38]. Its main function is to modulate peripheral signaling and homeostasis [39]. Given that 5-HT is a protonated molecule under physiological conditions, it is incapable of crossing the lipidic bilayer of cell membranes. Thus, the proper balance of its absorption into platelets and release from the same depends on the regulation of bidirectional transport carried out by SERT [37]. As the plasma level of 5-HT increases, SERT expression is upregulated on the platelet membrane and vice versa. Hence, regulating SERT expression is a plausible strategy for controlling plasma 5-HT levels [40].
In contrast, SRIs (e.g., fluoxetine and sertraline) prescribed to patients diagnosed with severe depression have been tested in animal models to assess their possible adverse effects. The principal clinical and biochemical changes are related to hemostasis, which is linked to the concentration of 5-HT in platelets. The use of SRIs has been reported to induce both low [41, 42] and high concentrations of 5-HT [43]. This discrepancy can be explained by the utilization of different drug schemes, assay methodologies, and/or techniques for the determination of 5-HT.
According to a study by Linder et al., veins (unlike arteries) do not have functional SERT. Nevertheless, they are capable of absorbing a significantly greater quantity of 5-HT than arteries, suggesting that their capture of 5-HT, independent of SERT [38] involves other monoamine transporters (e.g., the norepinephrine transporter and the dopamine active transporter) that are similar to SERT in their mechanism of absorption, function, and regulation [37, 44]. Further research is needed on the location and function of SERT and other monoamine transporters in the cardiovascular system in order to explore possible pharmacological targets for the treatment of vascular disorders.
The effects of 5-HT are mediated by specific 5-HTRs [45]. The 5-HT2A (type 2, subtype A) receptor is best characterized on the platelet membrane and in the cells of vascular smooth muscle tissue [46]. After binding with 5-HT2A, 5-HT is released into the bloodstream, resulting in the activation of nearby platelets and greater platelet aggregation to form thrombi during the inflammatory response. Moreover, there is evidence of a vasoconstrictor effect elicited by the activation of 5-HT2A receptors in cells of vascular smooth muscle cells, and a vasodilator effect stemming from the activation of 5-HT1B in endothelial cells through a mechanism dependent on nitric oxide (NO) [47].
Based on these findings, the antagonist sarpogrelate was tested on 5-HT2A receptors, observing a decreased serum concentration of 5-HT [48]. Such an effect could possibly diminish the formation of thrombi, thus reducing the risk of acute myocardial infarction. However, the search for a selective compound is complicated by the structural homology (46–50%) of 5-HT2A with the 5-HT2B and 5-HT2C subtypes [49]. Activation of 5-HT2B leads to the contraction of smooth muscle tissue of the stomach and muscular tissue in the human intestine. It also takes part in the relaxation of the endothelial tissue of veins [50, 51]. The 5-HT2C receptor has not been detected in the cardiovascular system and its activity is as yet unknown due to the lack of selective ligands [52].
The activation of 5-HT1A causes a drop in blood pressure and the heart rate [53], indicating a physiological role in the activation of the vagus nerve signaling the heart [54] and bladder [55]. The evaluation of antagonists of 5-HT1A has demonstrated a dose-dependent vasoconstriction, thus elevating the blood pressure and heart rate. It would then seem that selective agonists of 5-HT1A should be able to lower blood pressure, reduce the activity of the sympathetic nervous system, and increase stimulation of the vagus nerve [56].
In a similar sense, cardiopulmonary reflexes have been examined in relation to the brain and peripheral 5-HT1A receptors, which are known to modulate the recapture of 5-HT. Altered expression of 5-HT1A has been found on the platelet membrane of patients with major depressive disorder, with more severe depression associated with an overexpression of the receptor and decreased activity of 5-HT in platelets [14].
There are few reports on the function of 5-HT1B/1D subtypes. A study based on a rat model suggests that their activation may produce vasodilation and a drop in blood pressure [47, 52].
5-HT3 receptors are pentameric ion channels. They are controlled by a ligand consisting of five subunits generated by the alternative splicing of the RNA sequence of 5-HT3A-E [57]. Each subunit comprises ~450 amino acids and weighs an average of 50 kDa. These subunits are arranged around a central sodium, potassium, or calcium ion-conducting pore. In relation to other serotonin receptors, 5-HT3 has a distinct molecular structure and signaling pathway [58, 59]. Activation by 5-HT causes the ionic pore to open, and the entering current activates and subsequently desensitizes the receptor [60, 61]. 5-HT3 receptors are located in the spinal nucleus of the trigeminal nerve, the area postrema, and the solitary tract [62], areas of the CNS linked to emesis. They are also expressed on the surface of activated platelets and are associated with platelet aggregation. Hence, they likely contribute to thrombosis, although further investigation is necessary to clarify their physiological and pathophysiological role in platelet function [17, 63].
The affinity of 5-HT for 5-HT3 receptors in schizophrenic patients has been found to be four times greater than in healthy individuals. This affinity is related to the participation of isoform D of the amino acid serine (D-serine) in ligand-receptor binding [64]. Future research is needed on other possible structural and functional alterations in 5-HT3 receptors located on the platelets of schizophrenic patients in order to find new biomarkers and treatment targets. The two known therapeutic effects of targeting 5-HT3 receptors are related to nausea/emesis and irritable bowel syndrome. Regarding the former condition, ondansetron is an antagonist of 5-HT3 receptors that effectively relieves emesis induced by chemotherapy [65].
Regarding 5-HT4 receptors, the seven variants described in the literature differ in the sequence of the C-terminal segment. They have been detected at the central and peripheral level [65]. While their activation in the brain appears to influence long-term depression [66], in the intestine they can affect peristalsis, motility, and secretion [67]. 5-HT4 receptors are expressed on enterochromaffin cells in the gastrointestinal tract and on neurons of the enteric nervous system. The binding of 5-HT to these receptors facilitates the release of acetylcholine and the relaxation of the colon [68, 69], effects produced by the partial agonist tegaserod, which affects diverse physiological functions of the gastrointestinal tract [68, 70]. The expression level on platelet membranes is much lower for 5-HT4 than 5-HT2. Accordingly, there is an approximately 8-fold greater level of platelet mRNA encoding 5-HT2 than 5-HT4 receptors [18].
3. Serotonin receptor signaling pathways
The activation of 5-HTRs by 5-HT leads to the triggering of highly regulated signaling pathways capable of transmitting signals from the surrounding environment to the interior of the cell. Since signaling pathways elicit effector responses responsible for orchestrating many essential physiological processes of the human organism, they are a crucial factor in drug development. The most relevant signaling pathways activated by 5-HT are discussed hereafter, with the main focus on serotonin platelet receptors, and secondarily on those found on other types of cells.
Once released by the cells of the gastrointestinal tract, 5-HT is absorbed by SERT [71], thus allowing it to be dispersed into the bloodstream, captured by platelets, and transported to peripheral tissues. About 95% of 5-HT in the blood is stored in platelet dense granules, which also contain ATP, ADP, and Ca2+ [72, 73]. The various inhibitors of the capture of 5-HT by platelets include fluoxetine, sertraline, paroxetine, escitalopram, and citalopram (Figure 1). Although the specific mechanism of action of citalopram has not yet been defined, it is reportedly not through the inhibition of SERT.
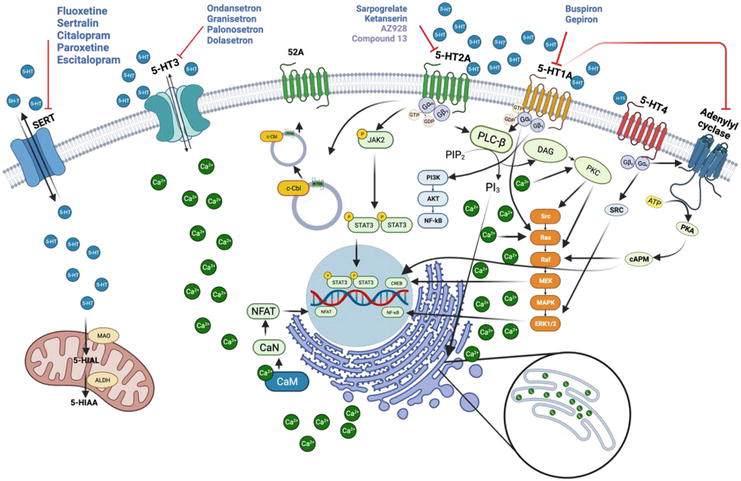
Figure 1.
The serotonin signaling pathway. Serotonin (5-HT), transported in and out of platelets by the serotonin transporter (SERT), triggers signaling by means of its receptors, including 5-HT1A, 5-HT2A, and 5-HT4 (which are G protein-coupled receptors, or GPCRs) as well as 5-HT3. The receptors illustrated on the platelet cell have significant functions in numerous cell types. Multiple signaling pathways (e.g., intracellular calcium release) are activated by these receptors. Signaling through such receptors has been associated with a variety of diseases. Drugs and compounds that interfere with the activity of 5-HT receptors have been shown to affect signaling, thus producing therapeutic and/or adverse effects. The blue circles represent 5-HT, the black arrows indicate stimulation, and the red blunt arrows portray inhibition [
3.1 5-HT1A receptors
5-HT1A receptors are coupled to the Gαi/o subunit of intracellular guanine nucleotide-binding proteins (G proteins), giving rise to the inhibition of adenylyl cyclase and the regulation of protein kinase A and cyclic adenosine monophosphate (PKA/cAMP) through descending pathways. Signaling triggered by these receptors increases the phosphorylation of extracellular signal-regulated kinases (ERKs) by means of a pathway dependent on phosphatidylinositol-3-kinase and serine/threonine kinase Akt (PI3K/Akt) (Figure 1) [74].
3.2 5-HT2A receptors
The binding of 5-HT to 5-HT2A receptors generates conformational changes that activate the Gαq subunit by the GDP/GTP exchange reaction. The resulting GTP activates the phospholipase C-β (PLC-β) enzyme by la dissociation of complex Gβγ [75]. The PLC-β enzyme splits the phospholipid phosphatidylinositol 4,5-bisphosphate (PIP2) into phosphatidylinositol diacylglycerol (DAG) and inositol triphosphate (IP3). DAG activates protein kinase C (PKC) on the membrane, while IP3 induces the release of Ca2+ from intracellular organelles (e.g., the endoplasmic reticulum). A high level of Ca2+ in the cytosol causes the formation of the Ca2+/calmodulin complex, which activates calcineurin phosphatase (CaN). This in turn gives rise to the translocation of the nuclear factor of activated T cells (NFAT) to the nucleus, where it plays a crucial role in the CNS and in neurological diseases [75]. In response to 5-HT binding, the 5-HT2A receptor can activate another signaling pathway by means of the phosphorylization of the Janus kinase Jak2. The latter undergoes auto-phosphorylization and at the same time triggers the phosphorylization of STAT3, resulting in its translocation to the nucleus [76]. The c-Cbl carboxyl terminus protein recycles 5-HT2A receptors (Figure 1) [77].
The inhibition of 5-HT2A receptors is of interest to limit platelet aggregation and as a consequence prevent adverse cardiovascular events. The main 5-HT2A inhibitors in clinical use today are ketanserin and sarpogrelate. Investigation is being carried out on small molecules such as compound AZ928, derived from 6-fluorobenzo [d]isoxazole. AZ928 has demonstrated a high affinity for the receptor and antiplatelet activity superior to that of sarpogrelate, with a favorable safety profile [78]. Other compounds have a nucleus of 4-phenylcyclohexane-5-spiro- and 5-methyl-5-phenyl-hidantoin bound to various fragments of aryl piperazine. One example is “compound 13”, reported to inhibit platelet aggregation in a manner comparable to ketanserin and with greater efficiency than sarpogrelate [79]. The antagonists of 5-HT2A are promising candidates for the future research on the development of new antiplatelet agents.
3.3 5-HT3 receptors
Pentameric complexes have a central conducting pore permeable to cations (e.g., Na+, Ca2+, or K+) [80, 81, 82]. The activation of these receptors in platelets brings about a rapid rise in the level of cytosolic calcium, known to play a fundamental role in platelet activation and aggregation [63]. Contrarily, resveratrol reduces the release of stored Ca2+ ions and at the same time inhibits the entrance of Ca2+ into platelets, thus inhibiting platelet aggregation prompted by thrombin (Figure 2) [83]. Further investigation is required to clarify the mechanism of the resveratrol-induced decrease in the level of Ca2+ ions in order to explore the feasibility of administering the drug to generate an effect opposite to the response of the activated receptor. The antagonists of 5-HT3 receptors include ondansetron, granisetron, tropisetron, palonosetron, dolasetron, and azasetron (Figure 1) [57].
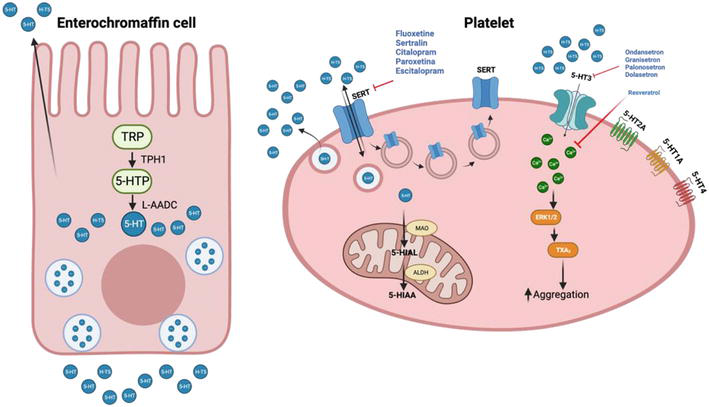
Figure 2.
The peripheral serotonergic system. Left: Synthesis of 5-HT in enterochromaffin cells, which transform tryptophan (TRP) into 5-hydroxytryptophan (5-HTP) through the enzyme tryptophan hydroxylase 1 (TPH1). Subsequently, 5-HTP is converted into 5-hydroxytryptamine (5-HT) by aromatic L-amino acid decarboxylase (L-AADC). Right: Platelets are activated by various serotonin receptors on their membrane (5-HT1A, 5-HT2A, 5-HT3, and 5-HT4). The storage of 5-HT in platelets takes place by means of the serotonin transporter (SERT), while the metabolism of 5-HT occurs through the enzyme monoamine oxidase (MAO). The latter transforms 5-HT into 5-hydroxy-indoleacetaldehyde (5-HIAL), which is oxidized by NAD + -dependent aldehyde dehydrogenase (ALDH) to 5-hydroxy-indoleacetic acid (5-HIAA). The blue circles represent 5-HT, black arrows indicate stimulation, and red blunt arrows portray inhibition. (modified from Ning Liu, et al., 2021) [
3.4 5-HT4 receptors
5-HT4 receptors have been identified on human platelets (as illustrated in Figure 1 and 2) [18]. Because of being coupled to G proteins, 5-HT4 receptors promote the production of cAMP and PKA by stimulating the activation of adenylyl cyclase. These receptors are also able to trigger metabolic pathways involving the activation of the protein kinase enzyme (MAPK) by nitrogen. In primary neurons, there is a potent but transitory activation of the ERK pathway, which operates independently of PKA. The principal route for the activation of neuronal ERKs by 5-HT4 receptors is dependent on Src protein-tyrosine kinase [84].
4. The platelet serotonergic system as a mirror of changes in stress, anxiety, depression, and epilepsy
In relation to the hypothesis of the link between the two locations of the platelet serotonergic system (the brain and peripheral tissues), recent preclinical studies have shown a significant reduction in brain and peripheral 5-HT of rodents (in platelet-rich plasma, or PRP) subsequent to forced swimming. A subchronic treatment with fluoxetine elicited a significant increase in the level of 5-HT in PRP. On the other hand, the same treatment prior to forced swimming led to a lesser decrease in the level of 5-HT in PRP than that observed in the control group of rodents submitted to forced swimming without treatment [85]. Accordingly, the level of peripheral 5-HT is sensitive to forced swimming, fluoxetine treatment, and the combination of the two.
Platelets have been proposed as markers possibly capable of distinguishing between acute and chronic stress as well as between somatic symptoms and psychiatric disorders [86, 87]. The low level of 5-HT in the brain of patients in a depressive state has been regulated by the prescription of SRIs [41, 42]. However, no significant difference was detected in the platelet content of 5-HT or in 5-HT reuptake in patients with post-traumatic stress disorder compared to healthy individuals [88]. Hence, further investigation is needed on the effects of distinct kinds of stress on the dynamics of the platelet serotonergic system.
Interestingly, evidence exists of a greater quantity of plasma 5-HT and a higher density of 5-HT2A receptors on platelets in patients with a combination of major depressive disorder and cardiovascular disease [89, 90]. As can be appreciated, a coherent explanation of the participation of the peripheral serotonergic system in depression has not yet been established. A previous systematic review revealed three different outcomes for the peripheral levels of 5-HT after treatment with antidepressant drugs: an increase, a decrease, and no significant change [91]. As a consequence, the peripheral levels of 5-HT cannot be utilized as a biomarker for the diagnosis of depression or as a measurement of the efficacy of treatments.
Likewise, contradictory results have been found in research on the role of the peripheral serotonergic system among anxiety disorders. For instance, a higher level and activity of SERT have been described in the platelets of patients with anxiety disorders [92]. However, a study on patients with panic disorder discovered a diminished average platelet volume in comparison with the control group of healthy individuals [93].
Despite intense efforts in the last few years to explore the involvement of the platelet serotonergic system in the pathophysiology of neurodegenerative diseases (e.g., epilepsy) [94, 95], the findings have not yet provided a clear picture of the overall contribution of the corresponding elements. On the other hand, there are various reports on adverse effects in peripheral tissues produced by treatments aimed at the platelet serotonergic system in the brain. For example, cabergoline and valproic acid (agonists of 5-HT2A and 5-HT2B receptors used to treat Parkinson’s disease and epilepsy, respectively) have been linked to the development of valvular heart disease [96, 97].
5. The platelet serotonergic system in the pathophysiology of coagulopathies associated with viral infections
COVID-19 causes an broad spectrum of diseases due to an exaggerated proinflammatory response, which often progresses to hyperinflammation accompanied by coagulopathy and a procoagulant endothelial phenotype [98]. The close relation between the condition of hypercoagulability and infection by COVID-19 can explain various phenomena observed in clinical practice. For instance, thromboembolic events have been described, such as venous and arterial thrombosis, leading to a greater incidence of strokes, myocardial and cerebral infarction, acute coronary syndrome, and pulmonary embolism [99]. Moreover, the activation of platelet aggregation by endothelial damage and the ensuing interaction with other cells aggravates the potential of hyperinflammation, structural remodeling of the pulmonary vasculature, and cardiovascular disease [100, 101].
5-HT has been shown to decrease the harmful effects of COVID-19 infection [102]. It manifests its activity in macrophages and dendritic cells, suppressing the generation of proinflammatory cytokines and chemokines. Furthermore, 5-HT promotes enhanced cytotoxicity in natural killer cells and reduces the production of tumor necrosis factor alpha (TNF-α) [103], while stimulating the proliferation of endothelial cells by binding to their 5-HT2 receptors [104].
There is evidence that ketanserin (a 5-HT2 antagonist) diminishes pulmonary platelet trapping and inhibits platelet activation and aggregation, thus facilitating respiration and lowering the risk of pulmonary fibrosis and adverse consequences in the kidneys, the CNS, and the cardiovascular system. Additionally, this treatment is able to revert the pulmonary vasoconstriction mediated by 5-HT [105].
SRI antidepressants have been employed to treat COVID-19 patients as well. They prevent the release of inflammatory cytokines [106], present anticoagulant activity [34, 41], and make the immune response more effective through the inhibition of proinflammatory molecules such as TNF-α, IL- Υ, IL-1β, and IL-6 [107, 108]. A selective SRI, fluvoxamine, is among the most studied and used treatments in the outpatient care of COVID-19 patients with risk factors. It has been shown to be beneficial in treating hypercoagulability in patients with COVID-19, possibly due to the effect that this SRI has on the reduction in platelet 5-HT concentration [109].
Regarding infection with dengue virus, the specific mechanisms involved in the development of thrombocytopenia are unknown. It is believed that platelets and 5-HT play a crucial role in understanding the vascular pathophysiology of this disease [30]. In studies carried out on adult patients with severe dengue, the acute phase induced platelet activation, increased intraplatelet 5-TH concentration, and greater expression of the activated fibrinogen receptor, the lysosomal marker CD63, and P-selectin [110].
6. The platelet serotonergic system in the pathophysiology of acute myocardial infarction
Acute myocardial infarction is the main cause of mortality around the world [111]. The prognosis of patients has been improved by opportune treatment, including pharmacological thrombolysis and percutaneous coronary intervention. However, the restauration of coronary blood flow provokes tissue damage, denominated myocardial reperfusion injury [32].
Platelets play a critical role in response to myocardial injury. Firstly, platelet glycoproteins IIb, IIIa, and VI have an essential function in the adhesion and aggregation of platelets. Secondly, activated platelets recruit proinflammatory leukocytes to the site of ischemic tissue, providing reperfusion during the first few days following a lesion. Thirdly, platelets release exosomes, which increase inflammation within the ischemic myocardium. They also release microvesicles and apoptotic bodies involved in myocardial tissue regeneration. Finally, 5-HT has been found to aggravate inflammation in the area of an infarct [111], suggesting that the respective receptors could be targeted to afford protective effects in post-infarct patients [112]. Some recently developed drugs have been administered to patients with acute myocardial infarction in the early phase of recovery. For instance, ondansetron shows anti-inflammatory effects by modulating the immune system and diminishing platelet activation [112].
7. The platelet serotonergic system in the pathophysiology of preeclampsia
Preeclampsia is a severe hypertensive complication of pregnancy that constitutes a significant risk for the mother and fetus [113, 114]. In women with preeclampsia, the following conditions have been detected: (1) enhanced availability of tryptophan, (2) a decrease in the activity of the kynurenine pathway and thus in the synthesis of indoleamine 2,3 dioxygenase, and (3) a reduction in the degradation of 5-HT by monoamine oxidase type A, resulting in a higher concentration of free 5-HT in maternal circulation [115, 116]. During pregnancy, 5-HT is synthesized in the syncytiotrophoblasts of chorionic villi. With preeclampsia, the generation of 5-HT is significantly lower in the syncytiotrophoblasts, but extracellular 5-HT probably causes placental hyperserotonemia (as found in normal maternal circulation) [117]. Various studies carried out on women with preeclampsia and gestational hypertension demonstrate an increase in 5-HT in the placenta, correlating positively with blood pressure and the severity of the disease [116, 117, 118].
Moreover, a release of 5-HT from mastocytes, basophils, and platelets is triggered by lesion or inflammation and can initiate cascades and other immune processes, among which are chemotaxis mediated by 5-HT1A receptors and cellular phagocytosis [119, 120]. Hence, 5-HT is necessary for the normal production of cytokines involved in the activation of inflammatory processes, including IFN-γ, IL-1B, IL-8, IL-12, TNFα, IL-17, and IL-6 [121, 122].
According to multiple reports on women with preeclampsia, the existence of hyperserotonemia during preeclampsia exacerbates several pathological processes, such as endothelial cell damage, platelet aggregation, the development of turbulence in blood flow, and microvascular damage in the placenta, all leading to more abundant inflammation [24, 123]. Damage to endothelial cells promotes platelet aggregation and as a consequence the release of 5-HT, which induces vasoconstriction mediated by 5-HT2 receptors in smooth muscle tissue and the uterine artery [25, 124].
8. Conclusions
The platelet serotonergic system is complex, affecting factors in the CNS and peripheral tissues. SRIs and other serotonergic drugs (agonists and antagonists of 5-HTRs) not only interact with the immune and central nervous system to treat certain diseases, but also may participate in coagulation, vascular permeability, and peripheral vascular resistance. It is crucial to consider both the central and peripheral locations and functions of the platelet serotonergic system before prescribing a medication that targets SERT or a serotonergic receptor (5-HT1A, 5-HT2A, 5-HT3, or 5-HT4) in order to avoid or reduce adverse effects (Figure 2). These targets represent an opportunity to treat the pathophysiology of infectious diseases (e.g., dengue and COVID-19), neuropsychological disorders (e.g., stress, depression, anxiety, and epilepsy), and cardiovascular diseases. They also represent potential biomarkers of the same disorders. Further research is required to provide greater insights into the mechanisms at play in the platelet serotonergic system. The challenge in drug development is to discover selective ligands capable of giving rise to the desired activity in the CNS or peripheral tissues (without provoking adverse effects) by increasing or decreasing the activation of platelets.
References
- 1.
Thon JN, Italiano JE. Platelets: Production, morphology and ultrastructure. Handbook of Experimental Pharmacology. 2012; 210 :3-22 - 2.
Ren DW, Liu WJ. Regulation of the megakaryocyte development and the platelet production—Review. Zhongguo Shi Yan Xue Ye Xue Za Zhi. 2018; 26 (6):1876-1880 - 3.
Hayashi T, Tanaka S, Hori Y, Hirayama F, Sato EF, Inoue M. Role of mitochondria in the maintenance of platelet function during in vitro storage. Transfusion Medicine. 2011;21 (3):166-174 - 4.
Mohammad-Zadeh LF, Moses L, Gwaltney-Brant SM. Serotonin: A review. Journal of Veterinary Pharmacology and Therapeutics. 2008; 31 (3):187-199 - 5.
Yubero-Lahoz S, Robledo P, Farré M, de laTorre R. Platelet SERT as a peripheral biomarker of serotonergic neurotransmission in the central nervous system. Current Medicinal Chemistry. 2013; 20 (11):1382-1396 - 6.
Jolas T, Aghajanian GK. Neurotensin and the serotonergic system. Progress in Neurobiology. 1997; 52 (6):455-468 - 7.
Erspamer V, Asero B. Identification of enteramine, the specific hormone of the enterochromaffin cell system, as 5-hydroxytryptamine. Nature. 1952; 169 (4306):800-801 - 8.
Lam DD, Heisler LK. Serotonin and energy balance: Molecular mechanisms and implications for type 2 diabetes. Expert Reviews in Molecular Medicine. 2007; 9 (5):1-24 - 9.
Wiersma DA, Roth RA. Clearance of 5-hydroxytryptamine by rat lung and liver: The importance of relative perfusion and intrinsic clearance. The Journal of Pharmacology and Experimental Therapeutics. 1980; 212 (1):97-102 - 10.
Jones LA, Sun EW, Martin AM, Keating DJ. The ever-changing roles of serotonin. The International Journal of Biochemistry & Cell Biology. 2020; 125 :105776 - 11.
Rendu F, Brohard-Bohn B. The platelet release reaction: Granules’ constituents, secretion and functions. Platelets. 2001; 12 (5):261-273 - 12.
del Río J, Lasheras B. Serotoninergic receptors: Physiological and therapeutic implications. Revista de Medicina de la Universidad de Navarra. 1993; 38 (3):29-38 - 13.
Bayer H, Müller T, Myrtek D, Sorichter S, Ziegenhagen M, Norgauer J, et al. Serotoninergic receptors on human airway epithelial cells. American Journal of Respiratory Cell and Molecular Biology. 2007; 36 (1):85-93 - 14.
Zhang ZJ, Wang D, Man SC, Ng R, McAlonan GM, Wong HK, et al. Platelet 5-HT(1A) receptor correlates with major depressive disorder in drug-free patients. Progress in Neuro-Psychopharmacology & Biological Psychiatry. 2014; 53 :74-79 - 15.
Liu YS, Yang M. The effect of 5-hydroxtryptamine on the regulation of megakaryocytopoiesis. Hematology. 2006; 11 (1):53-56 - 16.
Spigset O, Wilhelmsson C, Mjörndal T, Eriksson S. Serotonin 5-HT2A receptor binding in platelets from patients with Alzheimer’s disease or vascular dementia. International Psychogeriatrics. 2000; 12 (4):537-545 - 17.
Stratz C, Trenk D, Bhatia HS, Valina C, Neumann FJ, Fiebich BL. Identification of 5-HT3 receptors on human platelets: Increased surface immunoreactivity after activation with adenosine diphosphate (ADP) and thrombin receptor-activating peptide (TRAP). Thrombosis and Haemostasis. 2008; 99 (4):784-786 - 18.
Serebruany VL, El Mouelhi M, Pfannkuche HJ, Rose K, Marro M, Angiolillo DJ. Investigations on 5-HT₄ receptor expression and effects of tegaserod on human platelet aggregation in vitro. American Journal of Therapeutics. 2010; 17 (6):543-552 - 19.
Dib PRB, Quirino-Teixeira AC, Merij LB, Pinheiro MBM, Rozini SV, Andrade FB, et al. Innate immune receptors in platelets and platelet-leukocyte interactions. Journal of Leukocyte Biology. 2020; 108 (4):1157-1182 - 20.
Harifi G, Sibilia J. Pathogenic role of platelets in rheumatoid arthritis and systemic autoimmune diseases. Perspectives and therapeutic aspects. Saudi Medical Journal. 2016; 37 (4):354-360 - 21.
Habets KLL, Huizinga TWJ, Toes REM. Platelets and autoimmunity. European Journal of Clinical Investigation. 2013; 43 (7):746-757 - 22.
Hamzeh-Cognasse H, Berthelot P, Tardy B, Pozzetto B, Bourlet T, Laradi S, et al. Platelet toll-like receptors are crucial sensors of infectious danger moieties. Platelets. 2018; 29 (6):533-540 - 23.
Stasi R, Willis F, Shannon MS, Gordon-Smith EC. Infectious causes of chronic immune thrombocytopenia. Hematology/Oncology Clinics of North America. 2009; 23 (6):1275-1297 - 24.
Gumusoglu S, Scroggins S, Vignato J, Santillan D, Santillan M. The serotonin-immune Axis in preeclampsia. Current Hypertension Reports. 2021; 23 (7):37 - 25.
Stott DJ, Saniabadi AR, Hosie J, Lowe GD, Ball SG. The effects of the 5 HT2 antagonist ritanserin on blood pressure and serotonin-induced platelet aggregation in patients with untreated essential hypertension. European Journal of Clinical Pharmacology. 1988; 35 (2):123-129 - 26.
Yamamoto M, Jin JJ, Wu Z, Abe M, Tabara Y, Nagai T, et al. Interaction between serotonin 2A receptor and endothelin-1 variants in association with hypertension in Japanese. Hypertension Research. 2006; 29 (4):227-232 - 27.
Mammadova-Bach E, Mauler M, Braun A, Duerschmied D. Autocrine and paracrine regulatory functions of platelet serotonin. Platelets. 2018; 29 (6):541-548 - 28.
Ponomarev ED. Fresh evidence for platelets as neuronal and innate immune cells: Their role in the activation, differentiation, and deactivation of Th1, Th17, and Tregs during tissue inflammation. Frontiers in Immunology. 2018; 9 :406 - 29.
Canobbio I. Blood platelets: Circulating mirrors of neurons? Research and Practice in Thrombosis and Haemostasis. 2019; 3 (4):564-565 - 30.
Corzo-Gómez J, Picazo O, Castellanos-Pérez M, Briones-Aranda A. Systematic review of the serotonergic system in the pathophysiology of severe dengue: The theory of thrombocytopenia and vascular extravasation. Mini Reviews in Medicinal Chemistry. 2023; 23 (2):230-243 - 31.
Santos AP, Couto CF, Pereira SS, Monteiro MP. Is serotonin the missing link between COVID-19 course of severity in patients with diabetes and obesity? Neuroendocrinology. 2022; 112 (11):1039-1045 - 32.
Ziegler M, Wang X, Peter K. Platelets in cardiac ischaemia/reperfusion injury: A promising therapeutic target. Cardiovascular Research. 2019; 115 (7):1178-1188 - 33.
Kopeikina E, Ponomarev ED. The role of platelets in the stimulation of neuronal synaptic plasticity, electric activity, and oxidative phosphorylation: Possibilities for new therapy of neurodegenerative diseases. Frontiers in Cellular Neuroscience. 2021; 15 :680126 - 34.
Halperin D, Reber G. Influence of antidepressants on hemostasis. Dialogues in Clinical Neuroscience. 2007; 9 (1):47-59 - 35.
Zhu MM, Li HL, Shi LH, Chen XP, Luo J, Zhang ZL. The pharmacogenomics of valproic acid. Journal of Human Genetics. 2017; 62 (12):1009-1014 - 36.
Johnston JP, Nerenberg SF. Valproic acid-induced thrombocytopenia-related spontaneous systemic bleeding. American Journal of Case Reports. 2020; 21 :e927830 - 37.
Ni W, Watts SW. 5-hydroxytryptamine in the cardiovascular system: Focus on the serotonin transporter (SERT). Clinical and Experimental Pharmacology & Physiology. 2006; 33 (7):575-583 - 38.
Linder AE, Ni W, Szasz T, Burnett R, Diaz J, Geddes TJ, et al. A serotonergic system in veins: Serotonin transporter-independent uptake. Journal of Pharmacology and Experimental Therapeutics. 2008; 325 (3):714-722 - 39.
Brindley RL, Bauer MB, Blakely RD, Currie KPM. An interplay between the serotonin transporter (SERT) and 5-HT receptors controls stimulus-secretion coupling in sympathoadrenal chromaffin cells. Neuropharmacology. 2016; 110 :438-448 - 40.
Mercado CP, Kilic F. Molecular mechanisms of SERT in platelets: Regulation of plasma serotonin levels. Molecular Interventions. 2010; 10 (4):231-241 - 41.
Bismuth-Evenzal Y, Gonopolsky Y, Gurwitz D, Iancu I, Weizman A, Rehavi M. Decreased serotonin content and reduced agonist-induced aggregation in platelets of patients chronically medicated with SSRI drugs. Journal of Affective Disorders. 2012; 136 (1-2):99-103 - 42.
de Abajo FJ, Montero D, Garcia Rodriguez LA, Madurga M. Antidepressants and risk of upper gastrointestinal bleeding. Basic & Clinical Pharmacology & Toxicology. 2006; 98 (3):304-310 - 43.
Holck A, Wolkowitz OM, Mellon SH, Reus VI, Nelson JC, Westrin Å, et al. Plasma serotonin levels are associated with antidepressant response to SSRIs. Journal of Affective Disorders. 2019; 250 :65-70 - 44.
Paczkowski FA, Bryan-Lluka LJ, Pörzgen P, Brüss M, Bönisch H. Comparison of the pharmacological properties of cloned rat, human, and bovine norepinephrine transporters. The Journal of Pharmacology and Experimental Therapeutics. 1999; 290 (2):761-767 - 45.
Berger M, Gray JA, Roth BL. The expanded biology of serotonin. Annual Review of Medicine. 2009; 60 :355-356 - 46.
De Meyer SF, Vanhoorelbeke K, Broos K, Salles II, Deckmyn H. Antiplatelet drugs. British Journal of Haematology. 2008; 142 (4):515-528 - 47.
Machida T, Iizuka K, Hirafuji M. 5-Hydroxytryptamine and its receptors in systemic vascular walls. Biological & Pharmaceutical Bulletin. 2013; 36 (9):1416-1419 - 48.
Gocht A, Distler JHW, Spriewald B, Ramsperger-Gleixner M, Weyand M, Ensminger SM, et al. Effects of different serotonin receptor subtype antagonists on the development of cardiac allograft vasculopathy in murine aortic allografts. Transplant Immunology. 2018; 49 :43-53 - 49.
Hoyer D, Hannon JP, Martin GR. Molecular, pharmacological and functional diversity of 5-HT receptors. Pharmacology, Biochemistry, and Behavior. 2002; 71 (4):533-554 - 50.
Ellis ES, Byrne C, Murphy OE, Tilford NS, Baxter GS. Mediation by 5-hydroxytryptamine 2B receptors of endothelium-dependent relaxation in rat jugular vein. British Journal of Pharmacology. 1995; 114 (2):400-404 - 51.
Borman RA, Tilford NS, Harmer DW, Day N, Ellis ES, Sheldrick RLG, et al. 5-HT(2B) receptors play a key role in mediating the excitatory effects of 5-HT in human colon in vitro . British Journal of Pharmacology. 2002;135 (5):1144-1151 - 52.
Ramage A, Villalon C. 5-Hydroxytryptamine and cardiovascular regulation. Trends in Pharmacological Sciences. 2008; 29 (9):472-481 - 53.
Ramage AG. Central cardiovascular regulation and 5-hydroxytryptamine receptors. Brain Research Bulletin. 2001; 56 (5):425-439 - 54.
Bootle DJ, Adcock JJ, Ramage AG. The role of central 5-HT receptors in the bronchoconstriction evoked by inhaled capsaicin in anaesthetised Guinea-pigs. Neuropharmacology. 1998; 37 (2):243-250 - 55.
Ramage AG. The role of central 5-hydroxytryptamine (5-HT, serotonin) receptors in the control of micturition. British Journal of Pharmacology. 2006; 147 (S2):S120-S131 - 56.
Gradin K, Pettersson A, Hedner T, Persson B. Acute administration of 8-hydroxy-2-(di-n-propylamino) tetralin (8-OH-DPAT), a selective 5-HT-receptor agonist, causes a biphasic blood pressure response and a bradycardia in the normotensive Sprague-Dawley rat and in the spontaneously hypertensive rat. Journal of Neural Transmission. 1985; 62 (3-4):305-319 - 57.
Walstab J, Rappold G, Niesler B. 5-HT3 receptors: Role in disease and target of drugs. Pharmacology & Therapeutics. 2010; 128 (1):146-169 - 58.
Thompson AJ, Lummis SCR. The 5-HT3 receptor as a therapeutic target. Expert Opinion on Therapeutic Targets. 2007; 11 (4):527-540 - 59.
Reeves DC, Goren EN, Akabas MH, Lummis SCR. Structural and electrostatic properties of the 5-HT 3 receptor pore revealed by substituted cysteine accessibility mutagenesis*. Journal of Biological Chemistry [Internet]. 2001; 276 (45):42035-42042. Available from:http://www.jbc.org - 60.
Derkach V, Surprenant A, North RA. 5-HT3 receptors are membrane ion channels. Nature. 1989; 339 (6227):706-709 - 61.
Maricq AV, Peterson AS, Brake AJ, Myers RM, Julius D. Primary structure and functional expression of the 5HT3 receptor, a serotonin-gated ion channel. Science. 1979; 254 (5030):432-437 - 62.
Parker RMC, Barnes JM, Ge J, Barber PC, Barnes NM. Autoradiographic distribution of [3 H]-(S)-zacopride-labelled 5-HT 3 receptors in human brain. Journal of the Neurological Sciences. 1996; 144 (1-2):119-127 - 63.
Reeves DC, Lummis SCR. The molecular basis of the structure and function of the 5-HT 3 receptor: A model ligand-gated ion channel (review). Molecular Membrane Biology. 2002; 19 (1):11-26 - 64.
Fatima SK. Effect of D-serine on the serotonin receptors of human platelets. Experimental Brain Research. 2006; 173 (2):353-356 - 65.
Nichols DE, Nichols CD. Serotonin receptors. Chemical Reviews. 2008; 108 (5):1614-1641 - 66.
Kemp A, Manahan-Vaughan D. The 5-hydroxytryptamine 4 receptor exhibits frequency-dependent properties in synaptic plasticity and behavioural metaplasticity in the hippocampal CA1 region in vivo . Cerebral Cortex. 2005;15 (7):1037-1043 - 67.
Kadowaki M, Wang XO, Shimatani H, Yoneda S, Takaki M. 5-HT4 receptor enhances the propulsive power of the peristaltic reflex in the rat distal colon. Autonomic Neuroscience. 2002; 99 (1):62-65 - 68.
Gershon MD. Review article: Serotonin receptors and transporters—Roles in normal and abnormal gastrointestinal motility. Alimentary Pharmacology & Therapeutics. 2004; 20 (Suppl. 7):3-14 - 69.
Irving HR, Tan YY, Tochon-Danguy N, Liu H, Chetty N, Desmond PV, et al. Comparison of 5-HT4 and 5-HT7 receptor expression and function in the circular muscle of the human colon. Life Sciences. 2007; 80 (13):1198-1205 - 70.
Bockaert J, Claeysen S, Compan V, Dumuis A. 5-HT4 receptors. Current Drug Target-CNS & Neurological Disorders. 2004; 3 (1):39-51 - 71.
Liu N, Sun S, Wang P, Sun Y, Hu Q , Wang X. The mechanism of secretion and metabolism of gut-derived 5-hydroxytryptamine. International Journal of Molecular Sciences. 2021; 22 (15):7931 - 72.
Bertrand PP, Bertrand RL. Serotonin release and uptake in the gastrointestinal tract. Autonomic Neuroscience [Internet]. 2010; 153 (1-2):47-57. Available from:https://pubmed.ncbi.nlm.nih.gov/19729349/ - 73.
Terry N, Margolis KG. Serotonergic mechanisms regulating the GI tract: Experimental evidence and therapeutic relevance. Handbook of Experimental Pharmacology [Internet]. 2017; 239 :319-342. Available from:https://pubmed.ncbi.nlm.nih.gov/28035530/ - 74.
Dizeyi N, Hedlund P, Bjartell A, Tinzl M, Austild-Taskén K, Abrahamsson PA. Serotonin activates MAP kinase and PI3K/Akt signaling pathways in prostate cancer cell lines. Urologic Oncology: Seminars and Original Investigations. 2011; 29 (4):436-445 - 75.
Boczek T, Mackiewicz J, Sobolczyk M, Wawrzyniak J, Lisek M, Ferenc B, et al. The role of G protein-coupled receptors (GPCRs) and calcium signaling in schizophrenia. Focus on GPCRs activated by neurotransmitters and chemokines. Cell. 2021; 10 (5):1228 - 76.
Guillet-Deniau I, Burnol AF, Girard J. Identification and localization of a skeletal muscle secrotonin 5-HT2A receptor coupled to the Jak/STAT pathway. The Journal of Biological Chemistry. 1997; 272 (23):14825-14829 - 77.
Baldys A, Raymond JR. Role of c-Cbl carboxyl terminus in serotonin 5-HT2A receptor recycling and resensitization. Journal of Biological Chemistry. 2011; 286 (28):24656-24665 - 78.
Marcinkowska M, Kubacka M, Zagorska A, Jaromin A, Fajkis-Zajaczkowska N, Kolaczkowski M. Exploring the antiplatelet activity of serotonin 5-HT2A receptor antagonists bearing 6-fluorobenzo [d]isoxazol-3-ylpropyl motif–as potential therapeutic agents in the prevention of cardiovascular diseases. Biomedicine & Pharmacotherapy. 2022; 145 :112424 - 79.
Czopek A, Kubacka M, Bucki A, Siwek A, Filipek B, Pawłowski M, et al. Novel serotonin 5-HT2A receptor antagonists derived from 4-phenylcyclohexane-5-spiro-and 5-methyl-5-phenyl-hydantoin, for use as potential antiplatelet agents. Pharmacological Reports [Internet]. 2021; 73 (5):1361-1372. Available from:https://pubmed.ncbi.nlm.nih.gov/34115343/ - 80.
Green T, Stauffer KA, Lummis SCR. Expression of recombinant homo-oligomeric 5-hydroxytryptamine 3 receptors provides new insights into their maturation and structure. Journal of Biological Chemistry. 1995; 270 (11):6056-6061 - 81.
Boess FG, Beroukhim R, Martin IL. Ultrastructure of the 5-Hydroxytryptamine 3 receptor. Journal of Neurochemistry. 1995; 64 (3):1401-1405 - 82.
Barrera NP, Herbert P, Henderson RM, Martin IL, Edwardson JM. Atomic force microscopy reveals the stoichiometry and subunit arrangement of 5-HT3 receptors. National Academy of Sciences of the United States of America. 2005; 102 (35):12595-12600 - 83.
Marumo M, Ekawa K, Wakabayashi I. Resveratrol inhibits Ca2+ signals and aggregation of platelets. Environmental Health and Preventive Medicine. 2020; 25 (1):70 - 84.
Barthet G, Framery B, Gaven F, Pellissier L, Reiter E, Claeysen S, et al. 5-hydroxytryptamine 4 receptor activation of the extracellular signal-regulated kinase pathway depends on Src activation but not on G protein or beta-arrestin signaling. Molecular Biology of the Cell [Internet]. 2007; 18 (6):1979-1991. Available from:https://pubmed.ncbi.nlm.nih.gov/17377064/ - 85.
Bianchi M, Moser C, Lazzarini C, Vecchiato E, Crespi F. Forced swimming test and fluoxetine treatment: in vivo evidence that peripheral 5-HT in rat platelet-rich plasma mirrors cerebral extracellular 5-HT levels, whilst 5-HT in isolated platelets mirrors neuronal 5-HT changes. Experimental Brain Research. 2002;143 (2):191-197 - 86.
Sandrini L, Ieraci A, Amadio P, Zarà M, Barbieri SS. Impact of acute and chronic stress on thrombosis in healthy individuals and cardiovascular disease patients. International Journal of Molecular Sciences. 2020; 21 (21):7818 - 87.
Koudouovoh-Tripp P, Hüfner K, Egeter J, Kandler C, Giesinger JM, Sopper S, et al. Stress enhances proinflammatory platelet activity: The impact of acute and chronic mental stress. Journal of Neuroimmune Pharmacology. 2021; 16 (2):500-512 - 88.
Cicin-Sain L, Mimica N, Hranilovic D, Balija M, Ljubin T, Makarić G, et al. Posttraumatic stress disorder and platelet serotonin measures. Journal of Psychiatric Research. 2000; 34 (2):155-161 - 89.
Schins A, Honig A, Crijns H, Baur L, Hamulyák K. Increased coronary events in depressed cardiovascular patients: 5-HT2A receptor as missing link? Psychosomatic Medicine. 2003; 65 (5):729-737 - 90.
Williams MS, Ziegelstein RC, McCann UD, Gould NF, Ashvetiya T, Vaidya D. Platelet serotonin signaling in patients with cardiovascular disease and comorbid depression. Psychosomatic Medicine. 2019; 81 (4):352-362 - 91.
Wiaderna D, Gralewicz S, Tomas T. Behavioural changes following a four-week inhalation exposure to hemimellitene (1,2,3-trimethylbenzene) in rats. International Journal of Occupational Medicine and Environmental Health. 1998; 11 (4):319-334 - 92.
Zhuang X, Xu H, Fang Z, Xu C, Xue C, Hong X. Platelet serotonin and serotonin transporter as peripheral surrogates in depression and anxiety patients. European Journal of Pharmacology. 2018; 834 :213-220 - 93.
Göğçegöz Gül I, Eryılmaz G, Ozten E, Hızlı SG. Decreased mean platelet volume in panic disorder. Neuropsychiatric Disease and Treatment. 2014; 10 :1665-1669 - 94.
Somogyi GT, de Groat WC. Evidence for inhibitory nicotinic and facilitatory muscarinic receptors in cholinergic nerve terminals of the rat urinary bladder. Journal of the Autonomic Nervous System. 1992; 37 (2):89-97 - 95.
Cupello A, Audenino D, Scarrone S, Fornaro M, Gatta E, Fornaro P, et al. Epileptic seizures but not pseudoseizures are associated with decreased density of the serotonin transporter in blood platelet membranes. Neurochemical Research. 2008; 33 (11):2263-2268 - 96.
Pullan PT. Dopamine agonists, Dopplers and doubt: Cabergoline-associated cardiac valvulopathy. Internal Medicine Journal. 2009; 39 (4):213-215 - 97.
Solela G, Fedlu M. Rare recurrence of Sydenham chorea in an adult: A case report. International Medical Case Reports Journal. 2023; 16 :265-268 - 98.
Wool GD, Miller JL. The impact of COVID-19 disease on platelets and coagulation. Pathobiology. 2021; 88 (1):15-27 - 99.
Monteil V, Kwon H, Prado P, Hagelkrüys A, Wimmer RA, Stahl M, et al. Inhibition of SARS-CoV-2 infections in engineered human tissues using clinical-grade soluble human ACE2. Cell. 2020; 181 (4):905-913.e7 - 100.
Malerba M, Clini E, Malagola M, Avanzi GC. Platelet activation as a novel mechanism of atherothrombotic risk in chronic obstructive pulmonary disease. Expert Review of Hematology. 2013; 6 (4):475-483 - 101.
Malerba M, Nardin M, Radaeli A, Montuschi P, Carpagnano GE, Clini E. The potential role of endothelial dysfunction and platelet activation in the development of thrombotic risk in COPD patients. Expert Review of Hematology. 2017; 10 (9):821-832 - 102.
Szabo A, Gogolak P, Koncz G, Foldvari Z, Pazmandi K, Miltner N, et al. Immunomodulatory capacity of the serotonin receptor 5-HT2B in a subset of human dendritic cells. Scientific Reports. 2018; 8 (1):1765 - 103.
Lu B, Kwan K, Levine YA, Olofsson PS, Yang H, Li J, et al. α7 nicotinic acetylcholine receptor signaling inhibits inflammasome activation by preventing mitochondrial DNA release. Molecular Medicine. 2014; 20 (1):350-358 - 104.
Warner-Schmidt JL, Duman RS. VEGF is an essential mediator of the neurogenic and behavioral actions of antidepressants. Proceedings of the National Academy of Sciences of the United States of America. 2007; 104 (11):4647-4652 - 105.
Kuindersma M, Spronk PE. Ketanserin as potential additive drug to improve V/Q mismatch in COVID-19? Critical Care. 2020; 24 (1):526 - 106.
Roumestan C, Michel A, Bichon F, Portet K, Detoc M, Henriquet C, et al. Anti-inflammatory properties of desipramine and fluoxetine. Respiratory Research. 2007; 8 (1):35 - 107.
Zhang J, Zhang N, Lei J, Jing B, Li M, Tian H, et al. Fluoxetine shows neuroprotective effects against LPS-induced neuroinflammation via the notch signaling pathway. International Immunopharmacology. 2022; 113 (Pt A):109417 - 108.
Zheng W, Sun HL, Cai H, Zhang Q , Ng CH, Xiang YT. Antidepressants for COVID-19: A systematic review. Journal of Affective Disorders. 2022; 307 :108-114 - 109.
Reis G, dos Santos Moreira-Silva EA, Silva DCM, Thabane L, Milagres AC, Ferreira TS, et al. Effect of early treatment with fluvoxamine on risk of emergency care and hospitalisation among patients with COVID-19: The together randomised, platform clinical trial. The Lancet Global Health. 2022; 10 (1):e42-e51 - 110.
Michels M, Alisjahbana B, de Groot PG, Indrati AR, Fijnheer R, Puspita M, et al. Platelet function alterations in dengue are associated with plasma leakage. Thrombosis and Haemostasis. 2014; 112 (08):352-362 - 111.
Mauler M, Herr N, Schoenichen C, Witsch T, Marchini T, Härdtner C, et al. Platelet serotonin aggravates myocardial ischemia/reperfusion injury via neutrophil degranulation. Circulation. 2019; 139 (7):918-931 - 112.
Boshen Y, Yuankang Z, Taixi L, Kaifan N, Zhixiang W, Liang L, et al. Effects of ondansetron treatment on outcomes of critically ill patients with myocardial infarction partly through its anti-inflammatory activity. International Journal of Medical Sciences. 2023; 20 (6):709-716 - 113.
Bolte AC, van Geijn HP, Dekker GA. Pathophysiology of preeclampsia and the role of serotonin. European Journal of Obstetrics, Gynecology, and Reproductive Biology. 2001; 95 (1):12-21 - 114.
Gumusoglu SB, Chilukuri ASS, Santillan DA, Santillan MK, Stevens HE. Neurodevelopmental outcomes of prenatal preeclampsia exposure. Trends in Neurosciences. 2020; 43 (4):253-268 - 115.
Santillan MK, Pelham CJ, Ketsawatsomkron P, Santillan DA, Davis DR, Devor EJ, et al. Pregnant mice lacking indoleamine 2,3-dioxygenase exhibit preeclampsia phenotypes. Physiological Reports. 2015; 3 (1):e12257 - 116.
Ren D, Huang W, Yang M. Immunohistochemical locating and quantitative studies of 5-hydroxytryptamine and 5-hydroxytryptamine receptor in placental villi of patients with pregnancy induced hypertension. Zhonghua Fu Chan Ke Za Zhi. 1997; 32 (8):480-482 - 117.
Middelkoop CM, Dekker GA, Kraayenbrink AA, Popp-Snijders C. Platelet-poor plasma serotonin in normal and preeclamptic pregnancy. Clinical Chemistry. 1993; 39 (8):1675-1678 - 118.
Gujrati VR, Shanker K, Vrat S, Chandravati PSS. Novel appearance of placental nuclear monoamine oxidase: Biochemical and histochemical evidence for hyperserotonomic state in preeclampsia-eclampsia. American Journal of Obstetrics and Gynecology. 1996; 175 (6):1543-1550 - 119.
Kushnir-Sukhov NM, Gilfillan AM, Coleman JW, Brown JM, Bruening S, Toth M, et al. 5-hydroxytryptamine induces mast cell adhesion and migration. The Journal of Immunology. 2006; 177 (9):6422-6432 - 120.
Ieni JR, Meyerson LR. The 5-HT1A receptor probe [3H]8-OH-DPAT labels the 5-HT transporter in human platelets. Life Sciences. 1988; 42 (3):311-320 - 121.
Lichtblau N, Schmidt FM, Schumann R, Kirkby KC, Himmerich H. Cytokines as biomarkers in depressive disorder: Current standing and prospects. International Review of Psychiatry. 2013; 25 (5):592-603 - 122.
Cloëz-Tayarani I, Petit-Bertron AF, Venters HD, Cavaillon JM. Differential effect of serotonin on cytokine production in lipopolysaccharide-stimulated human peripheral blood mononuclear cells: Involvement of 5-hydroxytryptamine 2A receptors. International Immunology. 2003; 15 (2):233-240 - 123.
Gonzalez C, Cruz MA, Gallardo V, Albornoz J, Bravo I. Serotonin-induced vasoconstriction in human placental chorionic veins: Interaction with prostaglandin F2 alpha. Gynecologic and Obstetric Investigation. 1993; 35 (2):86-90 - 124.
Tanaka N, Nakamura E, Ohkura M, Kuwabara M, Yamashita A, Onitsuka T, et al. Both 5-hydroxytryptamine 5-HT2A and 5-HT1B receptors are involved in the vasoconstrictor response to 5-HT in the human isolated internal thoracic artery. Clinical and Experimental Pharmacology & Physiology. 2008; 35 (7):836-840