Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
Identifying microalgae biodiversity is essential to unleashing the diverse potential applications for microalgae. The aim of the chapter is to define the various tools and methodologies possible for revealing microalgae diversity. Identifying microalgae from natural environments, hotspots, and extreme environments enables us to isolate naturally tolerant species that may thrive and exhibit unique characteristics. Identifying microalgae will enable researchers to uncover new applications for these versatile organisms, including biofuel production, wastewater treatment, and pharmaceutical development. Hence, it is crucial to define the different possible identification approaches, beginning with the sample collection protocols and extending to the identification methods employed to determine the diversity of microalgae in an ecosystem.
Biotechnology Graduate Program, School of Sciences and Engineering, The American University in Cairo, New Cairo, Egypt
Walid Fouad*
Department of Biology, School of Sciences and Engineering, The American University in Cairo, New Cairo, Egypt
*Address all correspondence to: wfouad@aucegypt.edu
1. Introduction
Algae are a highly diverse group of photosynthetic organisms that can thrive in various aquatic and soil environments. They are categorized based on their size, with macroalgae being visible to the naked eye and microalgae being microscopic. Macroalgae, also known as seaweed, height can vary from a few cm and can reach up to 53 m in height if present in ideal conditions [1], such as the Ochrophyta genus Sargassum and Macrocystis [2, 3]. Microscopic algae can range from 0.2–200 μm and are further classified into picoalgae (algae size between 2 and 5 μm), and microalgae [4, 5]. Algae can be prokaryotic or eukaryotic, with eukaryotic algae belonging to various phyla such as Rhodophyta (red algae), Chlorophyta (green algae), Glaucophyta, Euglenophyta, Dinoflagellates, Cryptophyta, Heterokonts, Ochrophyta (brown algae), Eustigmatophyta, Bacillariophyta (diatoms), and Haptophyta. Despite algae being tremendously rich with essential by-products that are useful for many industrial applications, only about 30,000 species of algae have been identified and studied due to limitations in research on microalgal biodiversity [6, 7]. The lack of taxonomists and researchers in this field has resulted in inadequate records of microalgae diversity and the loss of their potential uses [8].
Why is it important to study biodiversity? A remarkable feature of our planet Earth is its ability to sustain a great variety of life. However, one of the most prevalent occurring environmental changes is the loss of this biological variation. Scientists have determined that the leading cause of species extinction is due to the drastic anthropogenic changes we currently face [9, 10]. Research indicated that the rate at which different resources are deteriorating is increasing, and water quality is decreasing exponentially with the declining species diversity [11]. The decrease in diversity would eventually lead to the homogenization of an ecosystem and, as a result, would lead to a decrease in the functionality of this ecosystem [12, 13]. The absence of heterogeneity in an ecosystem would further result in the loss of energy fluxes and matter that are essential to all ecological processes, which also includes the loss of control over the abundance, biomass, and distribution of organisms in an ecosystem [10, 14, 15]. For instance, an example of a food chain would include microalgae and picoalgae as the primary producer in an aquatic ecosystem. The absence or abundance of algae can lead to the interruption of this food chain due to the low evenness, which means that the community has a few abundant species that are dominating the community and could consequently result in the loss of the following consumers in the food chain [16]. The functionality of an environment is considered preserved when evenness is high, and the community contains various species that equally exist, with no one species gaining the upper hand and dominating an ecosystem [17, 18, 19].
The United Nations (UN) has expressed concern about the loss of habitats and species, which threatens the functionality of our ecosystem and planet Earth. In response, the UN declared 2010 as the International Year of Biodiversity and developed the Strategic Plan for Biodiversity 2011–2020 (CBD, 2010), which aims to improve the science base and biodiversity-related technologies by 2020. To achieve this goal, biodiversity analysis and studies must be intensified to preserve and protect the remaining biodiversity [20]. This chapter will focus on microalgae and will discuss the different methods used to collect environmental samples and identify microalgae diversity present in their natural environments, which will ultimately facilitate the conservation of biodiversity and preserve the known and unknown potentials of microalgae and their by-products.
2. Microalgae cellular structure importance to microalgae identification
Microalgae are microorganisms that belong to the kingdom Protista. Microalgae have different morphological characteristics that, when observed under the microscope, aid in taxonomic-level identification and classification. However, this is not the case when it comes to picoalgae such as Synechococcus and Chrysochromulina, which are extremely small-sized and are difficult to be identified on a morphological level [21, 22]. Hence, they are mostly classified through ribosomal RNA (rRNA) or other identical tools that do not depend on morphological characteristics.
2.1 Microalgae basic cellular structure
Microalgae cellular organization that can be used to distinguish microalgae includes unicellular (non-motile and motile), colonial (coenobic and non-coenobic), and filamentous (simple, branched, and heterotrichous). A feature that can clearly help in the identification of microalgae is their ability to locomotion [23, 24]. The motility of microalgae is of two types: flagellar motility and gliding motility [25]. Euglena and Pedinomonas are examples of microalgae that have flagella. Microalgae that do not have flagella still have the ability to glide, such as Synechococcus and Oscillatoria [26, 27]. Chlorophyta is the only phylum of microalgae that grows in the form of coenobic colonies (such as Volvox) [28]. While non-coenobic microalgae retain a gelatinous matrix or are found to be attached by a gelatinous stalk [29, 30]. Eukaryotic microalgae cells have organelles that are membrane-bound and unlike bacteria [31]. The prokaryotic cyanobacteria, also known as blue-green algae, are considered photosynthetic bacteria. Their deoxyribonucleic acid (DNA) lies freely in the cytoplasm, yet they, like microalgae, can carry oxygenic photosynthesis (conversion of CO2 and water to sugars using the energy from light) even though they lack a true chloroplast. In cyanobacteria, photosynthetic electron transport occurs in the thylakoid membranes present in the cytoplasm [32, 33]. Through evolution, eukaryotic microalgae have gained their cellular complexity from cyanobacteria endosymbiosis.
2.2 Plastid evolution and microalgae pigments
The plastid in microalgae originated as a result of endosymbiosis with cyanobacterium (as the primary chloroplast ancestors) [34, 35]. Further evolution of the plastid in microalgae occurred through a series of endosymbiotic events in which the primary endosymbiosis gave rise to the plastid of red algae, green algae, and plants chloroplast [35, 36, 37]. The secondary symbiosis resulted in more diversification and complexity of the plastid of microalgae species. Euglenoids, chlorarachniophytes, and a small group of dinoflagellates have up-taken the green algae chloroplast through the secondary endosymbiosis resulting in the three-membrane plastid [37]. The secondary endosymbiosis of glaucophytes and red algae further resulted in the presence of four-membrane-bound plastids that can be identified in Chlorarachniophytes, Haptophytes, and Heterokonts.
Certain plastid genes have been noted to track the origins of the plastid to cyanobacteria. RuBisCO genes, responsible for the production of ribulose-1,5-bisphosphate carboxylase/oxygenase, a critical enzyme in carbon fixation, have created confusion about the endosymbiotic events and the existence of microalgae from cyanobacteria [38]. Interestingly, some algae groups contained similar non-cyanobacteria enzymes. However, this has been elucidated by confirming that several gene duplications and subsequent gene divergence resulted in the paralogous sequences of the gene that have been transferred through horizontal gene transfer, giving rise to the Rubisco phylogeny [39]. By comparing the genome of cyanobacteria to that of a eukaryotic microalgae plastid, it was found that the number of genes significantly decreased. This is because several plastid genes have been moved to the nucleus, and some genes have been lost due to their redundant function [34]. In addition, there are various heterotrophic microalgae that are non-photosynthesis, including certain lineages of Euglophytes (E. gracilis and E. longa), diatoms (Nitzschia sp. strain NIES-3581), and green algae Polytomella spp. and Helicosporidium sp. Although these microalgae lack genes related to photosynthesis in their plastids, the plastids still play a crucial role in their survival by participating in various biological pathways other than photosynthesis [40, 41, 42].
The type of pigmentation present within the plastids also reflects the plastid lineage and phylogeny of the microalgae. The three major pigment classes present in microalgae are chlorophylls, carotenoids, and phycobiliproteins. The pigments absorb light at different wavelengths making their productivity depend on the light regimen they are exposed to. Microalgae, such as Chlorophyta (green algae), contain chlorophyll a and b pigments that produce absorption peaks at wavelengths between 300 and 700 nm, which is the wavelength of the visible spectrum [43]. Chlorophyll a absorbs light in the red-violet region and is present in all the microalgae phyla, while chlorophyll b absorbs light in the red-blue region, and both reflect green light [44, 45, 46]. Hence, the green appearance of the green microalgae. Cyanobacteria, on the other hand, lack chlorophyll b and favor low light intensities. Even though cyanobacteria lack chlorophyll b, however, the capture of photons is compensated by the presence of Phycobilisomes (PBS) light-harvesting antennae that contain chromophores, also known as phycobilins that are capable of absorbing orange, green, and blue-green light at wavelength 620 nm, 545 nm, and 495 nm, respectively [47, 48]. Carotenoids are terpenoid pigments that play an important role in photosynthesis as they harvest light within 400 and 550 nm and transfer it to chlorophyll a [49]. They also play an important role in protecting the microalgae cell and its photosynthetic machinery from reactive oxygen species (ROS) [50]. Unicellular and multicellular microalgae can have one or more chloroplasts present in a cell. Chloroplast division in microalgae occurs once per cell cycle [51], while in a plant, each cell may have up to 20 chloroplasts in each mesophyll cell of a plant [52]. Microalgae cells under the algae order Chlorococcales and Volvocales follow a cell cycle known as the multiple fission cycle (Cn). Under optimum conditions (in terms of light and temperature), they can divide into 8–16 identical daughter cells in the cell division cycle of about 6–12 h [53, 54].
3. Acquisition of environmental samples for studying algal diversity
Microalgae are typically found in environmental sources such as rivers, lakes, oceans, and soil. To identify the diversity of microalgae in a particular ecosystem, selecting the appropriate sample site is crucial. When choosing a sampling site, certain conditions must be taken into account. The Environmental Protection Authority (EPA) recommends developing a monitoring plan that considers various factors to ensure consistency in sampling. These measures are necessary to maintain sample integrity, as any changes to the conditions may alter the sample and lead to misleading information. Mismanagement of water and soil resources can result from inaccurate analysis of such samples [55, 56].
3.1 Sample collection methods
3.1.1 Manual sampling (grab/hand samples)
Water or soil samples can be collected manually by hand in a collection bottle. Using this method limits sampling to surface sampling. This method is cost-effective and does not eliminate microalgae depending on their size; however, this method limits the collection to microalgae with high buoyancy due to the presence of gas vacuoles that are mainly present in cyanobacteria [57, 58, 59].
3.1.2 Plankton nets
Nets are made up of nylon or silk and have a cylinder or valve attached to the bottom for sample collection. The nets come in different mesh sizes that can range from 10 to 1000 microns [60]. Plankton nets are used to quantitatively and qualitatively identify microalgae present in water sources. The net can be used to collect samples horizontally from a water surface, by adding weights to the net a vertical sample can be collected from the depth of the water source. The mesh size also plays an important role in the identification of microalgae from environmental samples. The size of the mesh selected can lead to the elimination of certain microalgae from the analysis of the environmental sample. Furthermore, the identified microalgae only mirror the filtering capability of the mesh [55]. Small-sized mesh samples are easily clogged, while large-sized mesh leads to the loss of smaller-sized microalgae.
3.1.3 Van Dorn sample bottle
The Van Dorn samplers enable depth sampling of water sources. The sampler consists of a hollow tube that is made up of PVC (Polyvinyl chloride) or acrylic that is connected to a rope. The tube is fitted with a seal at both ends that seals to lock the water collected in. The sampler is deposited in the water source to the desired depth, and the seals are released to lock the water in. The sample volume collected is known and the sample is drained through a drain valve [61]. The Schindler-Patalas trap is a similar tool that combines the Van Dorn and a phytoplankton net. The trap box is fitted with a mesh net at the bottom of the wall of the trap through which the collected sample is filtered after collection.
3.1.4 Van Veen grab sampler
The Van Veen grab sampler can be used to collect samples from soil or sediments. The sampler consists of claws that can grab sediments. Soil samples need to be either soaked in autoclaved distilled water or an enrichment media to extract and enhance the soil microalgae into a liquid media to enable and ease the microalgae identification.
3.2 Sample processing
Sample preservation may be necessary to maintain the integrity of the morphological features of the microalgae collected. Common fixatives used in preserving and fixing microalgae and protists are formaldehyde and Lugol’s iodine solution. Fixative stains at a low concentration can aid in morphological identification. However, the addition of a higher concentration may lead to the loss of flagellum and sometimes cell loss [62] and may hinder the molecular screening process [63].
To isolate a single cell from liquid media, one can use a micropipette or a glass capillary tube. The latter can be made by manually stretching a Pasteur pipette under a flame to create a capillary tube. This technique is useful for isolating single microalgae cells from a small sample drop under an inverted microscope. However, it may require expertise in both the isolation technique and microalgae identification. Another method of microalgae isolation is the streak plating of environmental samples on solid agar, usually a concentration of agar between 0.8 and 2%. This can be directly from an environmental sample or following a serial dilution of the sample. The plating of environmental samples can limit the identification of non-motile microalgae. However, several other methodologies may be involved in the identification of microalgae. Serial dilution is one of the most common methods that includes the preparation of serial dilution of a collected water sample or soaked soil sample with autoclaved distilled water. The dilution factor is maintained at each step [64]. The dilution set created can be cultured on an enrichment liquid or solid media. This method enables isolating single microalgae cells from the environment [65, 66]. The enrichment of a dilution on a liquid enrichment media can enable the identification of motile and non-motile microalgae. Fluorescence-activated cell sorting (FACS) and flow cytometry (FCM) cell sorting are two high-throughput automated isolation and screening techniques. The technique allows isolation and identification of microorganisms based on particle volume, fluorescence, and the cell’s ability to scatter light [67, 68, 69].
3.3 Enrichment of sample by nutrient media
An easy and inexpensive approach that is commonly used for the identification of an environmental sample is the use of culture media to enrich species in an environmental sample, followed by using microscopy to identify and isolate the species observed in the sample or a more advanced identification tool. Even though sample enrichment is considered a “golden tool” to analyze the diversity of an unknown sample, it is similar to the direct identification method, which depends on the resolution of the microscope and the expertise of the workforce [70]. Currently, the enrichment of environmental samples is usually followed up with identification on a molecular level to confirm the phylogenetic classification of the microalgae isolated [66, 71, 72, 73, 74, 75, 76, 77]. Microalgae can either survive as free organisms or co-exist in the presence of other organisms, such as lichens, in which the microalgae are said to live in symbiosis with other fungi and bacteria [78]. Therefore, enriching an environmental sample can help identify species that are not easily observable but are present only in large populations. Hence, optimization of nutrient composition, pH, temperature, and light intensity to enrich microalgae species and mimic the naturalistic conditions is necessary to ensure the identification of maximum diversity enrichment. Commonly used media like Bold’s Basal Media (BBM) and Blue-green media (BG-11) can be used to prevent favoritism toward specific genera or phyla. However, selective enrichment media may also be required to nourish specific microalgae phyla that require extra elements, such as diatoms that need silica [79] found in F/2 nutrient media [80, 81] (Table 1). This section focuses on the different culturing requirements of microalgae since it is important to consider their effects on microalgae identification, growth, and by-product production. Some culturing parameters that may require optimization for improvement of microalgae identification and isolation include the following:
Synthetic media
Main phyla enriched
Illumination
Carbon source
Species enriched
Ref.
BBM
Chlorophyta
25 μmol photons m−2 day−1 (continuous) Fluorescent lamps-5 Days
Synthetic nutrient media commonly used for microalgae enrichment.
3.3.1 Light
Microalgae are photosynthetic microorganisms that absorb light energy and convert it to chemical energy in the photosynthetic apparatus, the photosystem. The photosystem complex contains pigments such as chlorophyll or carotenoid that absorb light energy. Light energy is measured in irradiance; photons of light that are used photosynthetically are known as photosynthetically active radiation (PAR). PAR is usually reported in μmol m−2 s−1 or in μE m−2 s−1. Light intensity has been vastly studied to increase algae production and growth rates [67, 68, 69]. Artificial light is widely used in photobioreactors, with light-emitting diodes (LEDs) being a common choice. Research has focused on the impact of light color on growth, carbohydrate, protein, and pigment content. For instance, red light has been found to increase biomass, while blue light increases the cell concentration of Chlorella kessleri [96]. In a study by Jin et al., monochromatic red light (660 nm) resulted in decreased biomass and cellular components, leading to reduced carbohydrate, carotenoid, and protein levels in Dunaliella salina. Conversely, monochromatic blue light (445 nm) did not significantly increase biomass production but led to the accumulation of lipids, proteins, and carbohydrates. Combining red and blue light resulted in higher dried biomass and cell content compared to white light [97, 98]. White light increased the cellular productivity and protein content of Isochrysis zhanjiangensis, while red light increased the specific growth rate. Greenlight improved the pigment content of Isochrysis sp. but reduced biomass, and blue light promoted carbohydrate content [99]. In another study, Chlamydomonas reinhardtii exposed to yellow light resulted in the highest biomass concentration, while blue and red light separately resulted in the lowest biomass [100]. The photosynthetic process and growth of microalgae are significantly affected by the duration, wavelength, and intensity of light exposure [101, 102]. Hence, the choice of light color is essential for both microalgae cultivation and the production of specific biochemical by-products, which at the end might favor one species over another, impacting population diversity.
3.3.2 Nutrients
The growth of microalgae and the synthesis of proteins, lipids, and carbohydrates require macronutrients, mainly nitrogen (N), phosphorus (P), and potassium (K). Nitrogen is considered a crucial element as it forms a significant portion of structural molecules in microalgae, including nucleic acid, amines, and is also needed for cell wall components and pigment production [103, 104, 105]. Microalgae can obtain various forms of nitrogen from their surroundings, inorganic nitrogen, such as nitrates, nitrites, ammonium, and the organic nitrogen in the form of urea. Ammonium nitrate is taken up more easily and at a higher rate by microalgae. Nitrates, on the other hand, are converted into ammonium inside the microalgae chloroplast, making ammonium a better option in terms of the energy needed for the nitrate-to-ammonium conversion [106, 107]. However, microalgae exposure to high ammonium concentration can suppress nitrate uptake, possibly due to a negative feedback loop, which may be attributed to the high production of glutamine in the cell upon exposure to ammonium [108]. Hence nitrates are currently being commonly used in synthetic media. Phosphorus is another essential macronutrient that is required by microalgae to produce nucleic acid, phospholipids, and ATP, as well as for cell division and growth [109]. Insufficient phosphorus can result in decreased lipid content [110] and a decline in chlorophyll a levels, as phosphorus is necessary for its production [111]. Even though they are required in minute quantities, vitamins significantly influence the cellular biochemistry of microalgae. Microalgae exist in a mutualistic symbiotic relationship with bacteria, where they obtain vitamin B (cobalamin, thiamine, and biotin) produced by bacteria in exchange for carbon fixation [112]. As microalgae are unable to produce these vitamins de novo, they must be supplemented in synthetic media [113, 114].
3.3.3 Carbon sources
Microalgae require carbon sources for cell growth and the production of necessary primary metabolites such as carbohydrates, lipids, and proteins. Microalgae also can utilize different carbon sources. They can be cultured under autotrophic conditions using inorganic carbon such as carbon dioxide (CO2) or bicarbonate and light to produce energy through the process of photosynthesis. Yeh et al. (2010) tested the impact of using sodium bicarbonate (NaHCO3) as a carbon source on Chlorella vulgaris, the higher the concentration of NaHCO3 (1 g/L), the higher the biomass production along with the optimum light intensity [115]. Microalgae can also be heterotrophs under lightless conditions using organic carbon sources such as glucose or glycerol [116, 117, 118], while microalgae in the presence of light in mixotrophic cultures can use either organic or inorganic carbon sources. Phaeodactylum tricornutum showed an increase in biomass and lipid production when grown in a media with glycerol as a carbon source and 24-h illumination regimen [119]. Similarly, Chlorella sorokiniana [120] and diatom Thalassiosira pseudonana [121] showed better biomass and lipid production when grown on glycerol compared to glucose. Glucose assimilation was considered higher in heterotrophic culture resulting in nine times higher biomass of Auxenochlorella protothecoides when compared to autotrophic mode indicating that the uptake of glucose by A. protothecoides may be inhibited by light [122]. This was also observed when tested on Chlorella and Scenedesmus cultures [123]. Yet, Pavlova lutheri cultured on sucrose showed better biomass and lipid production under mixotrophic conditions when compared to heterotrophic conditions and other carbon sources such as glucose and glycerol [124].
Although the use of an organic carbon source with a light source allows microalgae to function under mixotrophic conditions as an alternative to autotrophic and heterotrophic conditions, the presence of organic carbon reduces the microalgae CO2 capture efficiency for photosynthesis [125, 126]. Additionally, the presence of inorganic carbon sources or dissolved CO2 is known to decrease pH due to the dissociation of the inorganic carbon and the release of H+ ions, leading to media acidification and reduced microalgae activity [127]. Some algae can utilize both inorganic carbon sources and CO2 capture, while others only depend on the inorganic carbon sources if present, and this process is pH-dependent. Since the bicarbonate form of inorganic carbon is mainly predominant at pH 9, while at lower pH, both the aqueous form of CO2 and the bicarbonate form are equally present [128]. Therefore, managing the pH of microalgae culture is crucial as it plays a vital role in making essential elements available for microalgae growth and regulating microalgae biochemical pathways.
3.3.4 pH
pH is one of the important parameters that needs to be maintained to conserve the physiology of microalgae. However, in a natural habitat, pH is quite variable and not necessarily maintained. Therefore, microalgae collected from the environment may have developed tolerance to the pH of the extreme ends. Hence, the need to optimize pH conditions for the enrichment of a complete diversity or specific microalgae depending on the research aim. In a culture media, the presence of the inorganic carbon source bicarbonate is converted to CO2 by the enzyme carbonic anhydrase to release hydroxyl ions that result in an increase in pH. This increase in pH results in the inhibition of cell growth due to the reduction of nutrient accessibility; an increase in pH will eventually result in a decrease in cell growth [129]. Freshwater microalgae have been reported to tolerate high pH (>10) and extremes of pH in general, while marine algae are limited to growth within the pH range of 8.1–8.3 [129, 130]. An increase of pH (from pH 8.5 to pH >11) in the culture media also leads to the flocculation of algal cells, a desired feature in the industry, and is used to harvest algae, but is troublesome when studying diversity [131].
3.3.5 Temperature
Another parameter that affects microalgae growth is temperature. The optimal temperature for microalgae to carry out cell functions and photosynthesis is generally between 15 and 30°C. Within the optimum temperature, ideally, an increase in temperature has a positive effect on the photosynthetic process and cell division. However, an unfavorable increase above the optimal temperature range can sharply decrease the growth rate of microalgae. This is because enzymatic functions are drastically affected by the high temperatures, hence affecting important enzymes of the electron transport chain, Calvin cycle, and membrane stability, eventually influencing the growth rate of the microalgae [132]. However, this is not always the case in cultures since microalgae are highly adaptive with the potential to adapt to extreme environments such as arid deserts [132, 133, 134, 135]. Furthermore, microalgae can be pre-adapted and acclimatized to certain temperatures depending on their habitat when cultured in the laboratory settings [136].
The direct identification of microalgae from environmental samples starts by collecting a sample either from water or soil as representative of the habitat. This is followed by direct observation through morphological characteristics of the different microalgae species under the microscope. There are several identification guidelines and keys available to aid in the identification of microalgae [31, 137, 138, 139, 140]. This method depends mainly on the presence of a research-grade microscope.
Scanning and transmission electron microscopes (SEM and TEM) are not options that are readily available in all research labs. However, they are used to study fine ultrastructure details on the species level of microalgae. Moreover, viewing microalgae under SEM and TEM requires specific cell preparation to preserve the ultrastructure of the microalgae. The preparations involve fixation, dehydration, and sputtering of the sample with gold particles [141, 142, 143]. Automated microalgae identification is a modern form of morphological level identification that is currently made available for the taxonomic identification of microalgae from an environmental sample. An automated system depends on machine learning and a training set that consists of a set of captured images of microalgae that a taxonomist has previously identified. This training set is used to train the software to classify the different microalgae genera [144, 145, 146, 147, 148, 149]. Yet, identifying microalgae based on their physical characteristics is challenging as their phenotypic traits are heavily influenced by the environment and their response to stress [150, 151]. To overcome this issue, molecular-based identification has emerged as an essential alternative approach that enables the identification of microalgae irrespective of any changes in their physical or morphological traits.
4.2 Molecular-level identification
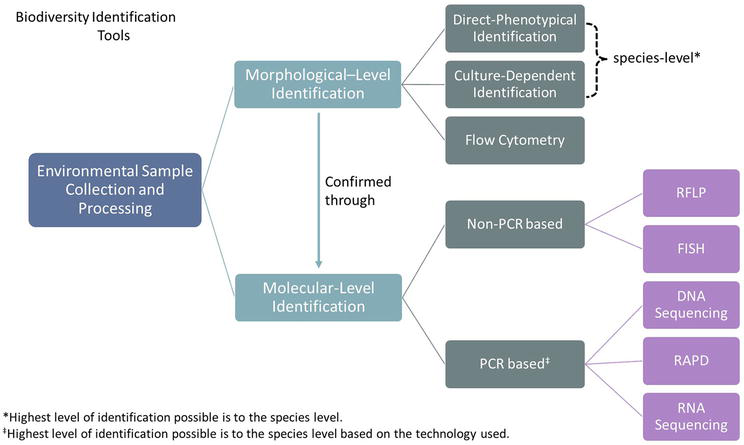
Molecular-level identification is the process of identifying microalgae on a genetic level. This process involves various steps and can be categorized into two main approaches: non-polymerase chain reaction (PCR)-based or PCR-based methods. These methods can be applied directly to environmental samples, enriched samples, or isolated species. Additionally, molecular level identification can serve as a confirmation tool for morphological level identification (Figure 1). This section will discuss the technologies utilized in both approaches for identifying microalgae diversity.
Figure 1.
Various methods that can be utilized to identify microalgae in environmental samples.
4.2.1 Non-PCR-based identification
4.2.1.1 Fluorescence in situ hybridization (FISH)
FISH is a technique that depends on the hybridization of a fluorescently labeled DNA sequence to its complementary sequence in a cell, followed by identification using epifluorescence microscopy. FISH has been used and optimized to identify harmful microalgae species mainly. This method has successfully identified dinoflagellates Gambierdiscus spp. [152], Biecheleria baltica [153] and the toxic Alexandrium sp. [154, 155, 156, 157], Chromophyta Heterosigma akashiwo [158].
4.2.1.2 Restriction fragment polymorphism (RFLP)
The identification of different organisms has been facilitated by RFLP, a technique that analyzes specific DNA sequence variations. This involves digesting DNA with restriction endonucleases and observing the digestion pattern through gel electrophoresis. Consequently, RFLP has been widely employed in gene mapping and DNA fingerprinting [159]. However, its use in microalgae identification has been limited [160]. PCR-RFLP or Terminal-RFLP (T-RFLP) are alternative methods to RFLP analysis, and they depend on the combination of PCR for DNA amplification of DNA barcodes and restriction fragment analysis, as discussed next.
4.2.2 PCR-based identification
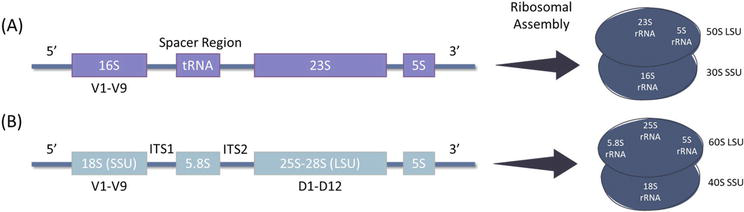
Polymerase chain reaction (PCR)-based molecular level identification depends on amplifying short conserved nucleic acid sequences known as DNA barcodes. Different DNA barcodes can also be used to identify different phyla of microalgae. For instance, the 16S ribosomal DNA (rDNA) sequence is mainly used for cyanobacteria identification region [161], while the 18S rDNA is used for eukaryotic microalgae by using primers designed to amplify the variable regions or the internal transcribed spacers (ITSs) in addition to the identification of eukaryotic microalgae through plastid genes [162] (Figure 2). The level of identification, whether to the genus or species level, depends on the reference database used for alignment. This method has gained popularity in taxonomical identification and phylogenetic analysis as it eliminates the need for culturing microalgae to confirm their presence, allowing the ease of direct identification from environmental samples. The identification and analysis of DNA barcodes have been facilitated by the development of various biotechnology tools (Table 2). The following are some tools that can be used to identify microalgae through DNA barcodes.
Figure 2.
Schematic diagram representing the ribosomal gene cluster (rDNA) of the (A) 16S and (B) 18S rDNA that are commonly used for the identification of cyanobacteria and eukaryotic microalgae, respectively. The hypervariable regions are denoted with V1-V9 on the 16S and 18S rDNA.
16S V3-V4 (for cyanobacteria) and 18S V4 rDNA (eukaryotic microalgae)
Colacium vesiculosum
Discoplastis spathirhyncha
Euglena sp.
Flexiglena variabilis
Lepocinclis sp.
Monomorphina pyrum
Phacus sp.
Trachelomonas sp.
Illumina MiSeq™ platform
Table 2.
Common DNA barcodes utilized for the identification of microalgae.
DNA barcode abbreviations: ribulose-1,5-bisphosphate carboxylase/oxygenase (rbcL), large subunit ribosomal DNA (LSU rDNA), small subunit ribosomal DNA (SSU rDNA), cytochrome c oxidase I (COI), encoding elongation factor (tufA gene), internal transcribed spacer region (ITS), cytochrome oxidase B (cob), calcium-modulated protein (calmodulin), damage checkpoint rad24 protein (rad24).
PCR-Restriction Fragment Polymorphism (PCR-RFLP) and Random Amplified Polymorphic DNA (RAPD) are two PCR-based techniques that have been utilized to identify different microalgae species. PCR-RFLP is a method that involves amplifying a DNA barcode before digesting the amplified region with restriction nucleases. This technique has been successfully used for the identification of microalgae such as Stihococcus sp., Nitschia sp., C. vulgaris, and Navicula accomoda through the PCR of 18S rDNA and its digestion [187]. PCR-RFLP has also been used to identify the dinoflagellate Gambierdiscus sp. isolated from the Caribbean Sea and Mexican coast was also identified through the amplification of the LSU rRNA D1-D2 sequences of the 18S rDNA [188]. Random Amplified Polymorphic DNA (RAPD) is another PCR-based technique that is useful for identifying genetic variation in different species [189]. RAPD has been used in the identification of different Chlorella strains [190], Microcystis species [191], Dunaliella species [192, 193], and diatom Chaetoceros species [194]. However, with the emergence of sequencing technology has provided a more reliable and accurate identification method over RAPD.
Sanger sequencing, also known as chain termination sequencing or first-generation sequencing, has been widely used as a confirmatory tool for identifying microalgae post-enrichment and isolation from environmental samples. The effectiveness of this method is dependent on the utilized DNA barcode, with a multiple primer approach often employed for identifying microalgae diversity across various phyla (Table 2). For instance, Ballesteros et al. used 16S rDNA and 16S–23S rDNA regions for cyanobacteria identification and 18S rDNA and ribulose-bisphosphate carboxylase (rbcL) gene for chlorophyta identification [163]. Other DNA barcodes used in identifying eukaryotic microalgae include the mitochondrial gene cytochrome c oxidase subunit 1 (cox1 or COI), ITS (ITS1–5.8S-ITS2), and elongation factor EF-Tu (tufA) gene [195].
Metagenomics involves the direct study of genomes in their natural habitat without isolation, and this is made possible due to the development of next-generation sequencing. Samples can be either directly used for DNA extraction, such as soil samples or water samples can be filtered with the appropriate filter paper followed by DNA extraction, and samples can be sequenced using Illumina MiSeq™ and HiSeq™ platform [186, 196, 197, 198]. Next-generation sequencing provides the best tool to identify the highest microalgal diversity from environmental samples; however, similar to any DNA-based tool, it depends on the sequences available in the database and is not usually combined with single species isolation.
4.3 Biochemical marker for microalgae identification
Several approaches have been employed to identify microalgae based on their pigment, also known as chemotaxonomic markers. Additionally, carbohydrates, lipids, and fatty acids may be used as chemotaxonomic markers. Microalgal photosynthetic pigments constitute a diverse range and are a highly valuable product with a vast array of commercial applications, including the food, cosmetics, and health industries [50, 199, 200, 201]. Despite the variety of chemotaxonomic markers, microalgae are known for their adaptability and plasticity as their cell composition and phenotypic characteristics are influenced by environmental changes such as temperature and nutrient composition [202, 203]. However, these changes can make it challenging to detect microalgae using such markers. Therefore, additional tests are necessary to account for the impact of various environmental conditions on microalgae cells, including limited nutrients, elevated CO2, and prolonged darkness, which affect cell composition [204, 205]. To address this issue, some research has focused on identifying specific macromolecules for different species [206, 207]. Serive et al. [208] determined that photosynthetic pigments cannot be used as the sole method of identification since the expression of these pigments depends upon various physiological and environmental factors, such as light and nutrients, and hence must be confirmed using other taxonomic identification tools [208].
Despite the fact that chemotaxonomic markers are not reliable in identifying microalgae, quantifying their biochemical composition after identification from their environment is an essential next step. To achieve this, Fourier-transformed infra-red (FTIR) spectroscopy is a powerful tool for the rapid identification of microalgal lipids, carbohydrates, and proteins through biomass characterization via the identification of functional groups [209, 210] (Table 3). However, FTIR is limited for use on environmental samples containing unidentified species, as it can only identify species whose chemical composition is already present in its library [204]. For the identification of fatty acid from microalgae, gas chromatography-flame ionization detection (GC-FID), gas chromatography-mass spectroscopy (GC–MS), and nuclear magnetic resonance (NMR) have been commonly used (Table 3). NMR has also been utilized to identify carbohydrates and pigments from microalgae. Other tools, such as high-performance liquid chromatography (HPLC) and ultraviolet-visible (UV–vis) spectrophotometry, have been employed for pigment analysis, ultimately allowing for the classification of microalgae up to the phyla level based on their pigments [233].
Identification tool
Microalgae biochemical product
Method of identification
Refs.
FTIR
Lipids, Carbohydrates, Proteins
The vibrational energy of functional groups such as CH2, OH, and C=O induced by the absorption of infrared radiation.
GC connected to flame ionization detectors is based on the burning of esterified fatty acids using a hydrogen flame after separation using gas chromatography.
Abbreviations: Fourier-transform infrared spectroscopy (FTIR); Gas chromatography-Flame Ionization Detection (GC-FID); Gas chromatography-mass spectroscopy (GC-MS); High-performance liquid chromatography (HPLC); Nuclear magnetic resonance (NMR); HPLC-diode-array detector (HPLC-DAD); Ultraviolet-visible (UV–vis) spectrophotometry.
4.4 Conclusion
Identification of microalgae is important for determining their value. Microalgae are rich in nutrients such as proteins, vitamins, minerals, essential fatty acids, and different pigments, making them a valuable source for human food, animal feed, and various industrial applications. In addition, microalgae are fast-growing and can be easily cultivated up to large scales, which make them a cost-effective solution for environmental clean-up. Different species of microalgae are known to be effective in bioremediation. Hence, researchers are continuously investigating different targeted strategies using such species for cleaning up polluted environments. This can lead to a more sustainable and eco-friendly approach to environmental remediation. By identifying the specific species of microalgae, it is possible to determine the potential health and industrial benefits. Therefore, accurate identification of microalgae is crucial for assessing their nutritional value and potential applications in different industries; additionally, biochemical characterization of microalgae can help in the discovery of new bioactive compounds with potential in various applications.
Identification and isolation of microalgae from their natural habitats are crucial for their utilization and extraction of diverse by-products. Various factors come into play when isolating and maintaining microalgae, such as sample collection, post-collection processing, and identification methods. Direct morphological identification, which involves identifying different phenotypical features of microalgae under the microscope, is a golden method that requires highly qualified taxonomists. To further ease the identification of microalgae from their environmental samples, enrichment of microalgae using different nutrients and optimizing their growth enables the ease of identifying low-density microalgae through enriching them. Molecular level identification is another means of identification that can be used to confirm the morphological level identification of microalgae or can be used to directly identify microalgae without culturing or isolating it. Utilizing several identification methods provides a holistic approach to identifying microalgae from environmental samples. Thus, the need for continuous optimization of the different identification methods and to aid the continuous monitoring of the microalgae diversity in our environment.
1.Srednick GS, Steele MA. Macroalgal physical structure predicts variation in some attributes of temperate fish assemblages better than macroalgal species composition. Marine Biology. 2022;169(11):1-15
2.Miller RJ, Lafferty KD, Lamy T, Kui L, Rassweiler A, Reed DC. Giant kelp, Macrocystis pyrifera, increases faunal diversity through physical engineering. Proceedings of the Biological Sciences 2018;285(1874):1-8
3.Marks LM, Reed DC, Holbrook SJ. Life history traits of the invasive seaweed Sargassum horneri at Santa Catalina Island, California. Aquatic Invasions. 2018;13(3):339-350
4.Belevich TA, Ilyash LV, Milyutina IA, Logacheva MD, Goryunov DV, Troitsky AV. Metagenomic analyses of white sea picoalgae: First data. Biochemistry (Moscow). 2015;80(11):1514-1521
5.Massana R. Eukaryotic picoplankton in surface oceans. Annual Review of Microbiology. 2011;65:91-110
6.May RM. The future of biological diversity in a crowded world. Current Science. 2002;82(11):1325-1331
7.Guiry MD. How many species of algae are there? Journal of Phycology. 2012;48(5):1057-1063
8.Scheffers BR, Joppa LN, Pimm SL, Laurance WF. What we know and don’t know about Earth’s missing biodiversity. Trends in Ecology & Evolution. 2012;27(9):501-510
9.Naeem S, Thompson LJ, Lawler SP, Lawton JH, Woodfin RM. Declining biodiversity can alter the performance of ecosystems. Nature. 1994;368:734-737
10.Cardinale BJ, Srivastava DS, Duffy JE, Wright JP, Downing AL, Sankaran M, et al. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature. 2006;443(7114):989
11.Worm B, Barbier EB, Beaumont N, Duffy JE, Folke C, Halpern BS, et al. Impacts of biodiversity loss on ocean ecosystem services. Science. 1979;314(5800):787-790
12.Adebayo O. Loss of biodiversity: The burgeoning threat to humans. Annals of Ibadan Postgraduate Medicine. 2019;17(1):1
13.Lefcheck JS, Byrnes JEK, Isbell F, Gamfeldt L, Griffin JN, Eisenhauer N, et al. Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats. Natural Communication. 2015;6:6936
14.Alsterberg C, Roger F, Sundbäck K, Juhanson J, Hulth S, Hallin S, et al. Habitat diversity and ecosystem multifunctionality—The importance of direct and indirect effects. Science Advances. 2017;3(2):e1601475
15.Barnes AD, Jochum M, Lefcheck JS, Eisenhauer N, Scherber C, O’Connor MI, et al. Energy flux: The link between multitrophic biodiversity and ecosystem functioning. Trends in Ecology & Evolution. 2018;33(3):186
16.Jochum M, Barnes AD, Brose U, Gauzens B, Sünnemann M, Amyntas A, et al. For flux’s sake: General considerations for energy-flux calculations in ecological communities. Ecology and Evolution. 2021;11(19):12948-12969
17.Pauli W, Jax K, Berger S. Protozoa in wastewater treatment: Function and importance. In: Beek B, editor. Biodegradation and Persistence. Berlin, Heidelberg: Springer Berlin Heidelberg; 2001. pp. 203-252. (The Handbook of Environmental Chemistry)
18.Medipally SR, Yusoff FM, Banerjee S, Shariff M. Microalgae as sustainable renewable energy feedstock for biofuel production. BioMed Research International. 2015:519513
19.Mohamed Ramli N, Yusoff FM, Giatsis C, Tan GYA, Verreth JAJ, Verdegem MCJ. Effects of Stigeoclonium nanum, a freshwater periphytic microalga on water quality in a small-scale recirculating aquaculture system. Aquaculture Research. 2018;49(11):3529-3540
20.Magurran AE, Dornelas M. Biological diversity in a changing world. Philosophical Transactions of the Royal Society B: Biological Sciences. 2010;365(1558):3593-3597
21.Agegian CR, editor. Collection M to ML. In: Biogeochemical Cycling and Fluxes Between the Deep Euphotic Zone and Other Oceanic Realms. U.S. Department of Commerce, National Oceanic and Atmospheric Administration, Oceanic and Atmospheric Research, Office of Undersea Research; 1988. p. 176pp. (National Undersea Research Program research report; vol. 88 (1))
22.Dussart BH. Les différentes catégories de plancton. Hydrobiologia. 1965;26(1-2):72-74
23.Cooper MB, Smith AG. Exploring mutualistic interactions between microalgae and bacteria in the omics age. Current Opinion in Plant Biology. 2015;26:147-153
24.Sime I. In: John DM, Whitton BA, Brook AJ, editors. The Freshwater Algal Flora of the British Isles: An Identification Guide to Freshwater and Terrestrial Aalgae. Cambridge: Cambridge University Press; 2002. p. 702. Aquat Conserv. 2004;14(1):105-105
25.Häder DP, Hoiczyk E. Gliding motility. In: Melkonian M, editor. Algal Cell Motility. Chapman and Hall, New York, London: Springer Science & Business Media; 1991. pp. 1-38
26.Halfen LN, Castenholz RW. Gliding motility in the blue-green alga Oscillatoria Princeps. Journal of Phycology. 1971;7(2):133-145
27.Heimann K, Benting J, Timmermann S, Melkonian M. The flagellar developmental cycle in algae – Two types of flagellar development in uniflagellated algae. Protoplasma. 1989;153(1-2):14-23
28.Starr RC. Colony formation in algae. In: Linskens HF, Heslop-Harrison J, editors. Cellular Interactions. Berlin, Heidelberg: Springer Berlin Heidelberg; 1984. pp. 261-290
29.Leliaert F, Smith DR, Moreau H, Herron MD, Verbruggen H, Delwiche CF, et al. Phylogeny and molecular evolution of the Green Algae. Critical Reviews in Plant Sciences. 2012;31(1):1-46
30.Lortou U, Gkelis S. Polyphasic taxonomy of green algae strains isolated from Mediterranean freshwaters. Journal of Biological Research (Greece). 2019;26(1):1-12
31.Kim SK. Handbook of Marine Microalgae: Biotechnology Advances. USA: Elsevier Science; 2015. pp. 1-585
32.Vermaas WF. Photosynthesis and respiration in Cyanobacteria. In: John Wiley & Sons, Ltd, editor. Encyclopedia of Life Sciences. Chichester, UK: John Wiley & Sons, Ltd; 2001
33.Shimakawa G, Dietz KJ. Electron transport in cyanobacterial thylakoid membranes: Are cyanobacteria simple models for photosynthetic organisms? Journal of Experimental Botany. 2023;74(12):3476-3487
34.Delwiche CF. Tracing the thread of plastid diversity through the tapestry of life. The American Naturalist. 1999;154(S4):S164-S177
35.Waller RF, Kořený L. Plastid complexity in dinoflagellates: A picture of gains, losses, replacements and revisions. In: Advances in Botanical Research. USA: Elsevier; 2017. pp. 105-143
36.Bhattacharya D, Medlin and L. Algal phylogeny and the origin of land plants. Plant Physiology. 1998;116(1):9-15
37.Keeling PJ. The endosymbiotic origin, diversification and fate of plastids. Philosophical Transactions of the Royal Society B: Biological Sciences. 2010;365(1541):729-748
38.McFadden GI. Primary and secondary endosymbiosis and the origin of plastids. Journal of Phycology. 2001;37:951-959
39.Nowicka B, Kruk J. Powered by light: Phototrophy and photosynthesis in prokaryotes and its evolution. Microbiological Research. 2016;186-187:99-118
40.Novák Vanclová AMG, Zoltner M, Kelly S, Soukal P, Záhonová K, Füssy Z, et al. Metabolic quirks and the colourful history of the Euglena gracilis secondary plastid. New Phytologist. 2020;225(4):1578-1592
41.Richtová J, Sheiner L, Gruber A, Yang SM, Kořený L, Striepen B, et al. Using diatom and apicomplexan models to study the heme pathway of chromera velia. International Journal of Molecular Sciences. 2021;22(12):1-23
42.Kayama M, Chen JF, Nakada T, Nishimura Y, Shikanai T, Azuma T, et al. A non-photosynthetic green alga illuminates the reductive evolution of plastid electron transport systems. BMC Biology. 2020;18(1):1-15
43.Brown JS. Absorption and fluorescence of chlorophyll A in particle fractions from different plants. Biophysical Journal. 1969;9:1542-1552
44.Eroglu E, Eggers PK, Winslade M, Smith SM, Raston CL. Enhanced accumulation of microalgal pigments using metal nanoparticle solutions as light filtering devices. Green Chemistry. 2013;15(11):3155-3159
45.Kommareddy A, Anderson G. Study of light requirements of a Photobioreactor. Time. 2004;300:1-8
46.Munawaroh HSH, Fathur RM, Gumilar G, Aisyah S, Yuliani G, Mudzakir A, et al. Characterization and physicochemical properties of chlorophyll extract from Spirulina sp. Journal of Physics Conference Series. 2019;1280(2):22013
47.Luimstra VM, Schuurmans JM, Verschoor AM, Hellingwerf KJ, Huisman J, Matthijs HCP. Blue light reduces photosynthetic efficiency of cyanobacteria through an imbalance between photosystems I and II. Photosynthesis Research. 2018;138(2):177-189
48.Pagels F, Guedes AC, Amaro HM, Kijjoa A, Vasconcelos V. Phycobiliproteins from cyanobacteria: Chemistry and biotechnological applications. Biotechnology Advances. 2019;37(3):422-443
49.Pagels F, Salvaterra D, Amaro HM, Guedes AC. Pigments from microalgae. In: Handbook of Microalgae-Based Processes and Products. USA: Elsevier; 2020. pp. 465-492
50.Chini Zittelli G, Lauceri R, Faraloni C, Silva Benavides AM, Torzillo G. Valuable pigments from microalgae: Phycobiliproteins, primary carotenoids, and fucoxanthin. Photochemical & Photobiological Sciences. 2023;2023:1-57
51.Okazaki K, Kabeya Y, Miyagishima S, ya. The evolution of the regulatory mechanism of chloroplast division. Plant Signaling & Behavior. 2010;5(2):164-167
52.Kubínová Z, Janáček J, Lhotáková Z, Kubínová L, Albrechtová J. Unbiased estimation of chloroplast number in mesophyll cells: Advantage of a genuine three-dimensional approach. Journal of Experimental Botany. 2014;65(2):609-620
53.Sayre R. Microalgae: The potential for carbon capture. BioScience. 2010;60(9):722-727
54.Zachleder V, Bišová K, Vítová M. The cell cycle of microalgae. In: Borowitzka MA, Beardall J, Raven JA, editors. The Physiology of Microalgae. Cham: Springer International Publishing; 2016. pp. 3-46
55.Ingebrigtsen RA, Hansen E, Andersen JH, Eilertsen HC. Field sampling marine plankton for biodiscovery. Scientific Reports. 2017;7(1):1-12
56.Creach V, Tett P, Bresnan E, Best M. CEMP Guidelines: Phytoplankton monitoring (OSPAR Agreement 2016-06); 2016
57.Reynolds CS, Oliver RL, Walsby AE. Cyanobacterial dominance: The role of buoyancy regulation in dynamic lake environments. New Zealand Journal of Marine Freshwater Research. 1987;21(3):379-390
58.Walsby AE. Structure and function of gas vacuoles. Bacteriological Reviews. 1972;1972:36
59.Walsby AE. The identification of gas vacuoles and their abundance in the hypolimnetic bacteria of Arco Lake, Minnesota. Microbial Ecology. 1974;1:51-61
60.Johnson WS, Allen DM, Fylling M. Zooplankton of the Atlantic and Gulf Coasts: Guide to their identification and ecology. In: Zooplankton of the Atlantic and Gulf Coasts: Guide to their Identification and Ecology: Second Edition. USA: Johns Hopkins University Press; 2012. pp. 1-453. (Zooplankton of the Atlantic and Gulf Coasts)
61.Usda Forest Service. Field guide for surface water sample and data collection. Water. 2001;2.3.4:1-3
62.Nollet LML, De Gelder LSP. Handbook of Water Analysis. Vol. 51. Boca Raton: CRC Press; 2014. pp. 51-4444-51-4444
63.Auinger BM, Pfandl K, Boenigk J. Improved methodology for identification of protists and microalgae from plankton samples preserved in Lugol’s iodine solution: Combining microscopic analysis with single-cell PCR. Applied and Environmental Microbiology, American Society for Microbiology. 2008;74(8):2505-2510
64.Cullen JJ, HL MI. On the use of the serial dilution culture method to enumerate viable phytoplankton in natural communities of plankton subjected to ballast water treatment. Journal of Applied Phycology. 2016;28:279-298
65.Bacha S. Techniques for isolation and purification of marine microalgae from the coast of Qatar. Qatar Foundation Annual Research Forum Proceedings. 2013;2013:EEP-033
66.Lee K, Eisterhold ML, Rindi F, Palanisami S, Nam PK. Isolation and screening of microalgae from natural habitats in the midwestern United States of America for biomass and biodiesel sources. Journal of Natural Science and Biological Medicine. 2014;5(2):333-339
67.Cellamare M, Rolland A, Jacquet S. Flow cytometry sorting of freshwater phytoplankton. Journal of Applied Phycology. 2010;22(1):87-100
68.Pereira H, Barreira L, Custódio L, Alrokayan S, Mouffouk F, Varela J, et al. Isolation and fatty acid profile of selected microalgae strains from the red sea for biofuel production. Energies (Basel). 2013;6(6):2773-2783
69.Sensen CW, Heimann K, Melkonian M. The production of clonal and axenic cultures of microalgae using fluorescence-activated cell sorting. European Journal of Phycology. 1993;28(2):93-97
70.Ebenezer V, Medlin LK, Ki JS. Molecular detection, quantification, and diversity evaluation of microalgae. Marine Biotechnology. 2012;14(2):129-142
71.Ali E, Mirza SS. A new method to isolate algal species from mix algal culture. bioRxiv. 2017;2017:1-16
72.Chik MN, Yahya L, Kamarudin KF, Tao DG, Takriff MS. Isolation, purification, and identification of microalgae from coal-fired power plant environment. Malaysian Journal of Analytical Sciences. 2017;21(2):460-469
73.Gumbi ST, Majeke BM, Olaniran AO, Mutanda T. Isolation, identification and high-throughput screening of neutral lipid producing indigenous microalgae from South African aquatic habitats. Applied Biochemical Biotechnology. 2017;182(1):382-399
74.Hutagalung RA, Sukoco AE, Soedharma D, Goreti LM, Andrean I, Elshaddai B, et al. Isolation, identification and growth optimization of microalgae derived from soft coral Dendronephthya sp. APCBEE Procedia. 2014;10:305-310
75.Liu P, Wei J, Yang K, Massey IY, Guo J, Zhang C, et al. Isolation, molecular identification, and characterization of a unique toxic cyanobacterium \textit{{Microcystis}} sp. found in {Hunan} {Province}, {China}. Journal of Toxicology and Environmental Health. Part A. 2018;81(21):1142-1149
76.Minhas AK, Hodgson P, Barrow CJ, Sashidhar B, Adholeya A. The isolation and identification of new microalgal strains producing oil and carotenoid simultaneously with biofuel potential. Bioresource Technology. 2016;211:556-565
77.Thangavel K, Radha Krishnan P, Nagaiah S, Kuppusamy S, Chinnasamy S, Rajadorai JS, et al. Growth and metabolic characteristics of oleaginous microalgal isolates from Nilgiri biosphere Reserve of India. BMC Microbiology. 2018;18:1-17
78.Mittermeier RA, Rylands AB. Biodiversity hotspots. In: Rescigno V, Maletta S, editors. Encyclopedia of the Anthropocene. New York: Nova Science Publishers; 2017. pp. 67-75 (Environmental science, engineering and technology series; vols. 1-5)
79.Theriot E, Herbarium D, Round FE, Crawford RM, Mann DG. The diatoms. Biology and morphology of the genera. Systematic Biology. 1992;41:125
80.Guillard RRL, J.H. R. Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt, and Detonula confervacea (cleve) gran. Canadian Journal of Microbiology. 1962;8:229-239
81.Guillard RRL. Culture of phytoplankton for feeding marine invertebrates. In: Culture of Marine Invertebrate Animals. USA: Springer US; 1975. pp. 29-60
82.Wu LF, Chen PC, Huang AP, Lee CM. The feasibility of biodiesel production by microalgae using industrial wastewater. Bioresource Technology. 2012;113:14-18
83.Sorokina KN, Samoylova YV, Parmon VN. Comparative analysis of microalgae metabolism on BBM and municipal wastewater during salt induced lipid accumulation. Bioresources Technology Reports. 2020;11:100548
84.Kumar V, Nanda M, Verma M. Application of agar liquid-gel transition in cultivation and harvesting of microalgae for biodiesel production. Bioresources Technology. 2017;243:163-168
85.Zittelli GC, Silva Benavides AM, Silovic T, Ranglová K, Masojídek J, Cicchi B, et al. Productivity and nutrient removal by the microalga Chlamydopodium fusiforme grown outdoors in BG-11 and piggery wastewater. Frontiers in Marine Science. 2022;9:2406
86.Norena-caro DA, Malone TM, Benton MG. Nitrogen sources and iron availability affect pigment biosynthesis and nutrient consumption in Anabaena sp. UTEX 2576. Microorganisms. 2021;9(2):431
87.Endar V, Sarjito S, Hutabarat J, Prayitno B. Effect of using Guillard and Walne technical culture media on growth and fatty acid profiles of microalgae Skeletonema SP. in mass culture. Journal of Coastal Zone Management. 2012;16(1):50-56
88.Harini AB, Sarangi NV, Nisha N, Rajkumar R. Cultivation of a marine diatom, Amphora sp., in municipal wastewater for enhancing lipid production toward sustainable biofuel production. South African Journal of Botany. 2023;155(1):288-297
89.Duong VT, Thomas-Hall SR, Schenk PM. Growth and lipid accumulation of microalgae from fluctuating brackish and sea water locations in South East Queensland—Australia. Frontiers in Plant Science. 2015;6:1-8
90.Kang KH, Ryu BM, Kim SK, Qian ZJ. Characterization of growth and protein contents from microalgae Navicula incerta with the investigation of antioxidant activity of enzymatic hydrolysates. Food Science and Biotechnology. 2011;20(1):183-191
91.Song Z, Lye GJ, Parker BM. Morphological and biochemical changes in Phaeodactylum tricornutum triggered by culture media: Implications for industrial exploitation. Algal Research. 2020;47:101822
92.Rani V, Deepika S, Abarna K, Uma A. Isolation, identification, and optimization of growth conditions for the marine microalgae isolated from the Gulf of Mannar, South-east coast of India. Regulation Studies in Marine Science. 2022;51:102189
93.Kinnunen V, Rintala J. The effect of low-temperature pretreatment on the solubilization and biomethane potential of microalgae biomass grown in synthetic and wastewater media. Bioresource Technology. 2016;221:78-84
94.Duangjan K, Kumsiri B, Pumas C. Lipid production by microalga Scenedesmus sp. AARL G022 in the cultivation with effluent from chicken manure biogas plant. Desalination Water Treatment. 2016;57(56):27191-27198
95.Choi HJ, Yu SW. Influence of crude glycerol on the biomass and lipid content of microalgae. Biotechnology & Biotechnological Equipment. 2015;29(3):506-513
96.Koc C, Anderson GA, Kommareddy A. Use of red and blue light-emitting diodes (LED) and fluorescent lamps to grow microalgae in a photobioreactor. Israeli Journal of Aquaculture - Bamidgeh. 2013;65(1):1-8
97.Jin C, Yu B, Qian S, Liu Q , Zhou X. Impact of combined monochromatic light on the biocomponent productivity of Dunaliella salina. Journal of Renewable and Sustainable Energy. 2021;13(2):023101 1-8
98.Pereira S, Otero A. Effect of light quality on carotenogenic and non-carotenogenic species of the genus Dunaliella under nitrogen deficiency. Algal Research. 2019;44:101725
99.Lv B, Liu Z, Chen Y, Lan S, Mao J, Gu Z, et al. Effect of different Colored LED lighting on the growth and pigment content of Isochrysis zhanjiangensis under laboratory conditions. Journal of Marine Science and Engineering. 2022;10(11):1752
100.de Mooij T, de Vries G, Latsos C, Wijffels RH, Janssen M. Impact of light color on photobioreactor productivity. Algal Research. 2016;15:32-42
101.Maltsev Y, Maltseva K, Kulikovskiy M, Maltseva S. Influence of light conditions on microalgae growth and content of lipids, carotenoids, and fatty acid composition. Biology (Basel). 2021;10(10):1-24
102.Kwan PP, Banerjee S, Shariff M, Yusoff F. Influence of light on biomass and lipid production in microalgae cultivation. Aquaculture Research. 2021;52(4):1337-1347
103.Becker EW. Micro-algae as a source of protein. Biotechnology Advances. 2007;25(2):207-210
104.Yaakob MA, Mohamed RMSR, Al-Gheethi A, Ravishankar GA, Ambati RR. Influence of nitrogen and phosphorus on microalgal growth, biomass, lipid, and fatty acid production: An overview. Cell. 2021;10(2):1-19
105.Zarrinmehr MJ, Farhadian O, Heyrati FP, Keramat J, Koutra E, Kornaros M, et al. Effect of nitrogen concentration on the growth rate and biochemical composition of the microalga, Isochrysis galbana. The Egyptian Journal of Aquatic Research. 2020;46(2):153-158
106.Salbitani G, Carfagna S. Ammonium utilization in microalgae: A sustainable method for wastewater treatment. Sustainability. 2021;13(2):956
107.Lourenço SO, Barbarino E, Mancini-Filho J, Schinke KP, Aidar E. Effects of different nitrogen sources on the growth and biochemical profile of 10 marine microalgae in batch culture: An evaluation for aquaculture. Phycologia. 2002;41(2):158-168
108.Lachmann SC, Mettler-Altmann T, Wacker A, Spijkerman E. Nitrate or ammonium: Influences of nitrogen source on the physiology of a green alga. Ecology and Evolution. 2019;9(3):1070
109.Fu L, Li Q , Yan G, Zhou D, Crittenden JC. Hormesis effects of phosphorus on the viability of Chlorella regularis cells under nitrogen limitation. Biotechnology for Biofuels. 2019;12(1):1-9
110.Chen M, Tang H, Ma H, Holland TC, Ng KYS, Salley SO. Effect of nutrients on growth and lipid accumulation in the green algae Dunaliella tertiolecta. Bioresource Technology. 2011;102(2):1649-1655
111.Xing Y, Guo L, Wang Y, Zhao Y, Jin C, Gao M, et al. An insight into the phosphorus distribution in extracellular and intracellular cell of Chlorella vulgaris under mixotrophic cultivation. Algal Research. 2021;60:102482
112.Tandon P, Jin Q , Huang L. A promising approach to enhance microalgae productivity by exogenous supply of vitamins. Microbial Cell Factories. 2017;16(1):1-13
113.Badr AA, Fouad WM. Identification of culturable microalgae diversity in the River Nile in Egypt using enrichment media. African Journal of Biological Sciences. 2021;3(2):50
114.Croft MT, Lawrence AD, Raux-Deery E, Warren MJ, Smith AG. Algae acquire vitamin B12 through a symbiotic relationship with bacteria. Nature. 2005;438(7064):90-93
115.Yeh KL, Chang JS, Chen WM. Effect of light supply and carbon source on cell growth and cellular composition of a newly isolated microalga Chlorella vulgaris ESP-31. Engineering in Life Sciences. 2010;10(3):201-208
116.Barros A, Pereira H, Campos J, Marques A, Varela J, Silva J. Heterotrophy as a tool to overcome the long and costly autotrophic scale-up process for large scale production of microalgae. Scientific Reports. 2019;9(1):1-7
117.Liang MH, Xue LL, Jiang JG. Two-stage cultivation of Dunaliella tertiolecta with glycerol and triethylamine for lipid accumulation: A viable way to alleviate the inhibitory effect of triethylamine on biomass. Applied Environmental Microbiology. 2019;85(4):1-10
118.Patel A, Matsakas L, Rova U, Christakopoulos P. Heterotrophic cultivation of Auxenochlorella protothecoides using forest biomass as a feedstock for sustainable biodiesel production. Biotechnology Biofuels. 2018;11(1):169
119.Morais KCC, Conceição D, Vargas JVC, Mitchell DA, Mariano AB, Ordonez JC, et al. Enhanced microalgae biomass and lipid output for increased biodiesel productivity. Renewable Energy. 2021;163:138-145
120.Cheah WY, Show PL, Juan JC, Chang JS, Ling TC. Enhancing biomass and lipid productions of microalgae in palm oil mill effluent using carbon and nutrient supplementation. Energy Conversion and Management. 2018;164:188-197
121.Baldisserotto C, Sabia A, Guerrini A, Demaria S, Maglie M, Ferroni L, et al. Mixotrophic cultivation of Thalassiosira pseudonana with pure and crude glycerol: Impact on lipid profile. Algal Research. 2021;54:102194
122.Xiao Y, Guo J, Zhu H, Muhammad A, Deng H, Hu Z, et al. Inhibition of glucose assimilation in Auxenochlorella protothecoides by light. Biotechnology for Biofuels. 2020;13(1):1-13
123.Bardone E, Bravi M, Keshavarz T, Vidotti ADS, Coelho RS, Franco LM, et al. Miniaturized culture for heterotrophic microalgae using low cost carbon sources as a tool to isolate fast and economical strains. Chemical Engineering Transactions. 2014;38:325-330
124.Bashir KMI, Mansoor S, Kim NR, Grohmann FR, Shah AA, Cho MG. Effect of organic carbon sources and environmental factors on cell growth and lipid content of Pavlova lutheri. Annales de Microbiologie. 2019;69(4):353-368
125.Ogbonna IO, Ogbonna JC, Ogbonna IO, Ogbonna JC. Effects of carbon source on growth characteristics and lipid accumulation by microalga Dictyosphaerium sp. with potential for biodiesel production. Energy and Power Engineering. 2018;10(2):29-42
126.Chavoshi ZZ, Shariati M. Lipid production in Dunaliella bardawil under autotrophic, heterotrophic and mixotrophic conditions. Brazilian Journal of Oceanography. 2019;67:e19249
127.Meier L, Vilchez C, Cuaresma M, Torres-Aravena Á, Jeison D. Effect of pH change on the microalgae-based biogas upgrading process. Applied Sciences. 2022;12(23):12194
128.Qiu R, Gao S, Lopez PA, Ogden KL. Effects of pH on cell growth, lipid production and CO2 addition of microalgae Chlorella sorokiniana. Algal Research. 2017;28:192-199
129.Goldman JC, Azov Y, Riley CB, Dennett MR. The effect of pH in intensive microalgal cultures. I. Biomass regulation. Journal of Experimental Marine Biology and Ecology. 1982;57(1):1-13
130.Taraldsvik M, Myklestad S. The effect of pH on growth rate, biochemical composition and extracellular carbohydrate production of the marine diatom skeletonema costatum. European Journal of Phycology. 2000;35(2):189-194
131.Fan J, Zheng L, Bai Y, Saroussi S, Grossman AR. Flocculation of Chlamydomonas reinhardtii with different phenotypic traits by metal cations and high pH. Frontiers in Plant Science. 2017;8:1997
132.Ras M, Steyer JP, Bernard O. Temperature effect on microalgae: A crucial factor for outdoor production. Reviews in Environmental Science and Biotechnology. 2013;12(2):153-164
133.Azua-Bustos A, González-Silva C, Arenas-Fajardo C, Vicuña R. Extreme environments as potential drivers of convergent evolution by exaptation: The Atacama Desert Coastal Range case. Frontiers in Microbiology. 2021;10(10):1-24
134.Cameron RE, Blank GB, Aeronautics U, States N, Administration S. Desert Algae: Soil Crusts and Diaphanous Substrata as Algal Habitats. Jet Propulsion Laboratory, USA: California Institute of Technology; 1966 (JPL Technical Report, 32-971)
135.Perera I, Subashchandrabose SR, Venkateswarlu K, Naidu R, Megharaj M. Consortia of cyanobacteria/microalgae and bacteria in desert soils: an underexplored microbiota. Applied Microbiology and Biotechnology. 2018;102:7351-7363
136.Bleeke F, Rwehumbiza VM, Winckelmann D, Klöck G. Isolation and characterization of new temperature tolerant microalgal strains for biomass production. Energies (Basel). 2014;7(12):7847-7856
137.Serediak N, Huynh M. Algae Identification Field Guide: An Illustrative Field Guide on Identifying Common Algae Found in the Canadian Prairies. Ottawa: Agriculture and Agri-Food Canada; 2011
138.Evans RI, Prescott GW. How to Know the Fresh-Water Algae. Bulletin of the Torrey Botanical Club. Dubuque, Iowa: W.C. Brown Company; 1956. p. 311. (Pictured key nature series; vol. 83)
139.John DM, Whitton BA, Brook AJ. The Freshwater algal flora of the British Isles: an identification guide to freshwater and terrestrial algae. Choice Reviews Online. 2012;49:49-6880-49-6880
140.Tomas CR, Hasle GR. Identifying Marine Phytoplankton. San Diego: Academic Press; 1997
141.Gärtner G, Uzunov B, Ingolic E, Kofler W, Gacheva G, Pilarski P, et al. Microscopic investigations (LM, TEM and SEM) and identification of Chlorella isolate R-06/2 from extreme habitat in Bulgaria with a strong biological activity and resistance to environmental stress factors. Biotechnology and Biotechnological Equipment. 2015;29(3):536-540
142.Jeon SM, Kim JH, Kim T, Park A, Ko AR, Ju SJ, et al. Morphological, molecular, and biochemical characterization of monounsaturated fatty acids-rich chlamydomonas sp. Kiost-1 isolated from Korea. Journal of Microbiology and Biotechnology. 2015;25(5):723-731
143.Murtey M, Ramasamy P. Sample preparations for scanning Electron Microscopy – Life Sciences. In: Modern Electron Microscopy in Physical and Life Sciences. London, UK: InTech; 2016
144.Deglint JL, Jin C, Chao A, Wong A. The feasibility of automated identification of six algae types using feed-forward neural networks and fluorescence-based spectral-morphological features. IEEE Access. 2019;7:7041-7053
145.Pardeshi R, Deshmukh PD. Classification of microscopic algae: An observational study with AlexNet. In: Advances in Intelligent Systems and Computing. MRCET, Hyderabad: Springer; 2020. pp. 309-316
147.Reimann R, Zeng B, Jakopec M, Burdukiewicz M, Petrick I, Schierack P, et al. Classification of dead and living microalgae Chlorella vulgaris by bioimage informatics and machine learning. Algal Research. 2020;48:101908
148.Santhi N, Pradeepa C, Subashini P, Kalaiselvi S. Automatic identification of algal community from microscopic images. Bioinform Biol Insights. 2013;7:327-334
149.Schulze K, Tillich UM, Dandekar T, Frohme M. PlanktoVision – an automated analysis system for the identification of phytoplankton. BMC Bioinformatics. 2013;14(1):115
150.Leung C, Grulois D, Chevin LM. Plasticity across levels: Relating epigenomic, transcriptomic, and phenotypic responses to osmotic stress in a halotolerant microalga. Molecular Ecology. 2022;31(18):4672
151.Abinandan S, Venkateswarlu K, Megharaj M. Phenotypic changes in microalgae at acidic pH mediate their tolerance to higher concentrations of transition metals. Current Research Microbiological Science. 2021;2:100081
152.Pitz KJ, Richlen ML, Fachon E, Smith TB, Parsons ML, Anderson DM. Development of fluorescence in situ hybridization (FISH) probes to detect and enumerate Gambierdiscus species. Harmful Algae. 2021;101:101914
153.Sundström AM, Kremp A, Tammilehto A, Tuimala J, Larsson U. Detection of the bloom-forming cold-water dinoflagellate Biecheleria baltica in the Baltic Sea using LSU rRNA probes. Aquatic Microbial Ecology. 2010;61(2):129-140
154.Sako Y, Hosoi-Tanabe S, Uchida A. Fluorescence in-situ hybridization using rRNA-targeted probes for simple and rapid identification of the toxic dinoflagellates Alexandrium tamarense and A Catenella. Journal of Phycology. 2004;40(3):598-605
155.Hosoi-Tanabe S, Sako Y. Rapid detection of natural cells of Alexandrium tamarense and A. catenella (Dinophyceae) by fluorescence in situ hybridization. Harmful Algac. 2005;4(2):319-328
156.Hosoi-Tanabe S, Sako Y. Development and application of fluorescence in situ hybridization (FISH) method for simple and rapid identification of the toxic dinoflagellates Alexandrium tamarense and Alexandrium catenella in cultured and natural seawater. Fisheries Science. 2006;72(1):77-82
157.Tang X, Yu R, Zhou M, Yu Z. Application of rRNA probes and fluorescence in situ hybridization for rapid detection of the toxic dinoflagellate Alexandrium minutum. Chinese Journal of Oceanology and Limnology. 2012;30(2):256-263
158.Chen GF, Wang GC, Zhang CY, Zhang BY, Wang XK, Zhou BC. Development of rRNA and rDNA-targeted probes for fluorescence in situ hybridization to detect Heterosigma akashiwo (Raphidophyceae). Journal of Experimental Marine Biology and Ecology. 2008;355(1):66-75
159.Mittal B, Chaturvedi P, Tulsyan S. Restriction Fragment Length Polymorphism: Brenner’s Encyclopedia of Genetics: Second Edition. 2013:190-193
160.Beetul K, Sadally SB, Taleb-Hossenkhan N, Bhagooli R, Puchooa D. An investigation of biodiesel production from microalgae found in Mauritian waters. Biofuel Research Journal. 2014;1(2):58-64
161.Nübel U, Garcia-Pichel F, Muyzer G. PCR primers to amplify 16S rRNA genes from cyanobacteria. Applied and Environmental Microbiology. 1997;1997
162.Burja AM, Tamagnini P, Bustard MT, Wright PC. Identification of the green alga, Chlorella vulgaris (SDC1) using cyanobacteria derived 16S rDNA primers: Targeting the chloroplast. FEMS Microbiology Letters. 2001;202(2):195-203
163.Ballesteros I, Terán P, Guamán-Burneo C, González N, Cruz A, Castillejo P. DNA barcoding approach to characterize microalgae isolated from freshwater systems in Ecuador. Neotrop Biodivers. 2021;7(1):170-183
164.Yanuhar U, Caesar NR, Musa M. Identification of local isolate of microalgae Chlorella vulgaris using Ribulose-1,5-Bisphosphate Carboxylase/Oxygenase Large Subunit (rbcL) Gene. IOP Conference Series Material Science Engineering. 2019;546(2):1-6
165.Fitriyah F, Faramitha Y, Sari DA, Kresnawaty I, Panji T, Santoso D. Molecular identification and phylogenetic analysis of Chlorella isolates from Indonesia using rbcL gene. E-Journal Menara Perkebunan. 2021;89(1):17-25
166.Wang C, Zou S, Fei C, Wang C, Gao Z, Bao Y, et al. How DNA barcoding can be more effective in microalgae identification: a case of cryptic diversity revelation in Scenedesmus (Chlorophyceae). Scientific Reports. 2016;6(1):1-13
167.Strunecký O, Kopejtka K, Goecke F, Tomasch J, Lukavský J, Neori A, et al. High diversity of thermophilic cyanobacteria in Rupite hot spring identified by microscopy, cultivation, single-cell PCR and amplicon sequencing. Extremophiles. 2019;23(1):35-48
168.Kurobe T, Baxa DV, Mioni CE, Kudela RM, Smythe TR, Waller S, et al. Identification of harmful cyanobacteria in the Sacramento-San Joaquin Delta and Clear Lake, California by DNA barcoding. Springerplus. 2013;2(1):1-12
169.Johansen JR, Mareš J, Pietrasiak N, Bohunická M, Zima J, Štenclová L, et al. Highly divergent 16S rRNA sequences in ribosomal operons of Scytonema hyalinum (Cyanobacteria). PLoS One; 2017;12(10):1-16
170.Webb VL, Maas EW. Sequence analysis of 16S rRNA gene of cyanobacteria associated with the marine sponge Mycale (Carmia) hentscheli. FEMS Microbiology Letters. 2002;207(1):43-47
171.Svenning MM, Eriksson T, Rasmussen U. Phylogeny of symbiotic cyanobacteria within the genus Nostoc based on 16S rDNA sequence analyses. Archives of Microbiology. 2005;183(1):19-26
172.Casamatta DA, Vis ML, Sheath RG. Cryptic species in cyanobacterial systematics: A case study of Phormidium retzii (Oscillatoriales) using RAPD molecular markers and 16S rDNA sequence data. Aquatic Botany. 2003;77(4):295-309
173.Zou S, Bao Y, Wu X, Wang C. DNA barcoding diatoms from China with multiple genes. Frontier in Marine Science. 2021;8:1423
174.Hamsher SE, Evans KM, Mann DG, Poulíčková A, Saunders GW. Barcoding diatoms: Exploring alternatives to COI-5P. Protist. 2011;162(3):405-422
175.Piredda R, Claverie JM, Decelle J, de Vargas C, Dunthorn M, Edvardsen B, et al. Diatom diversity through HTS-metabarcoding in coastal European seas. Scientific Reports. 2018;8(1):1-12
176.Urbánková P, Veselá J. DNA-barcoding: A case study in the diatom genus Frustulia (Bacillariophyceae). Nova Hedwigia. 2013
177.Urbánková P, Scharfen V, Kulichová J. Molecular and automated identification of the diatom genus Frustulia in northern Europe. Diatom Research. 2016;31(3):217-229
178.Stern RF, Horak A, Andrew RL, Coffroth MA, Andersen RA, Küpper FC, et al. Environmental barcoding reveals massive Dinoflagellate diversity in marine environments. PLoS One. 2010;5(11)
179.Hwang J, Kang HW, Moon SJ, Hyung JH, Lee ES, Park J. Metagenomic analysis of the species composition and seasonal distribution of marine Dinoflagellate communities in Four Korean Coastal Regions. Microorganisms. 2022;10(7):1-14
180.Pochon X, Putnam HM, Burki F, Gates RD. Identifying and characterizing alternative molecular markers for the symbiotic and free-living Dinoflagellate genus Symbiodinium. PLoS One. 2012;7(1):e29816
181.Lin S, Zhang H, Hou Y, Zhuang Y, Miranda L. High-level diversity of dinoflagellates in the natural environment, revealed by assessment of mitochondrial cox1 and cob genes for dinoflagellate DNA barcoding. Applied and Environmental Microbiology. 2009;75(5):1279-1290
182.Endo H, Ogata H, Suzuki K. Contrasting biogeography and diversity patterns between diatoms and haptophytes in the central Pacific Ocean. Scientific Reports. 2018;8(1):1-13
183.Liu H, Probert I, Uitz J, Claustre H, Aris-Brosou S, Frada M, et al. Extreme diversity in noncalcifying haptophytes explains a major pigment paradox in open oceans. Proceedings of the National Academy Science USA. 2009;106(31):12803-12808
184.Egge E, Bittner L, Andersen T, Audic S, de Vargas C, Edvardsen B. 454 pyrosequencing to describe microbial eukaryotic community composition, diversity and relative abundance: A test for marine haptophytes. PLoS One. 2013;8(9):e74371
185.Łukomska-Kowalczyk M, Karnkowska A, Krupska M, Milanowski R, Zakryś B. DNA barcoding in autotrophic euglenids: Evaluation of COI and 18s rDNA. Journal of Phycology. 2016;52(6):951-960
186.Salmaso N, Vasselon V, Rimet F, Vautier M, Elersek T, Boscaini A, et al. DNA sequence and taxonomic gap analyses to quantify the coverage of aquatic cyanobacteria and eukaryotic microalgae in reference databases: Results of a survey in the Alpine region. Science of The Total Environment. 2022;834:155175
187.Bérard A, Dorigo U, Humbert JF, Martin-Laurent F. Microalgae community structure analysis based on 18S rDNA amplification from DNA extracted directly from soil as a potential soil bioindicator. Agronomie. 2005;25(2):285-291
188.Lozano-Duque Y, Richlen ML, Smith TB, Anderson DM, Erdner DL. Development and validation of PCR-RFLP assay for identification of Gambierdiscus species in the greater Caribbean region. Journal of Applied Phycology. 2018;30(6):3529
189.Panigrahi S, Velraj P, Subba RT. Functional microbial diversity in contaminated environment and application in bioremediation. Microbial Diversity in the Genomic Era. 2019:359-385
190.Cho JJ, Kim YT, Hur SB, Kim Y. Random amplified polymorphic DNA (RAPD) identification of genetic variation in Chlorella species. Korean Journal of Fisheries and Aquatic Sciences. 1996;29(6):761-769
191.Nishihara H, Miwa H, Watanabe M, Nagashima M, Yagi O, Takamura Y. Random amplified polymorphic DNA (RAPD) analyses for discriminating genotypes of Microcystis cyanobacteria. Bioscience, Biotechnology, and Biochemistry. 1997;61(7):1067-1072
192.Gómez PI, González MA. Genetic variation among seven strains of Dunaliella salina (Chlorophyta) with industrial potential, based on RAPD banding patterns and on nuclear ITS rDNA sequences. Aquaculture. 2004;233(1-4):149-162
193.Tsai M. Application of the random amplified polymorphic DNA (RAPD) technique to the identification and classification of Dunaliella species, Department of Biology, University of Wollongong, 1994. Available from: https://ro.uow.edu.au/theses/2754
194.Kutako M, Hiransuchalert R, Watchasit S, Kaewduang M, Hanchana O, Setthamongkol P, et al. Morphological and molecular comparison as a useful tool for identification of the three centric marine diatoms (Bacillariophyceae: Chaetoceros). Archives of Microbiology. 2023;205(5):173
195.Suarez-Montes D, Borrell YJ, Gonzalez JM, Rico JM. Isolation and identification of microalgal strains with potential as carotenoids producers from a municipal solid waste landfill. Science of The Total Environment. 2022;802:149755
196.Choi J, Park JS. Comparative analyses of the V4 and V9 regions of 18S rDNA for the extant eukaryotic community using the Illumina platform. Scientific Reports. 2020;10(1):1-11
197.Yun HS, Kim YS, Yoon HS. Illumina MiSeq analysis and comparison of freshwater microalgal communities on Ulleungdo and Dokdo Islands. Polish Journal of Microbiology. 2019;68(4):527
198.Kumar V, Al Momin S, Kumar VV, Ahmed J, Al-Musallam L, Shajan AB, et al. Distribution and diversity of eukaryotic microalgae in Kuwait waters assessed using 18S rRNA gene sequencing. PLoS One. 2021;16(4):e0250645
199.Begum H, Yusoff FMD, Banerjee S, Khatoon H, Shariff M. Availability and utilization of pigments from microalgae. Critical Reviews in Food Science and Nutrition. 2015;56(13):2209-2222
200.Sun H, Wang Y, He Y, Liu B, Mou H, Chen F, et al. Microalgae-derived pigments for the food industry. Marine Drugs. 2023;21(2):82
201.Patel AK, Albarico FPJB, Perumal PK, Vadrale AP, Ntan CT, Chau HTB, et al. Algae as an emerging source of bioactive pigments. Bioresource Technology. 2022;351:126910
202.Sassenhagen I, Wilken S, Godhe A, Rengefors K. Phenotypic plasticity and differentiation in an invasive freshwater microalga. Harmful Algae. 2015;41:38-45
203.Thrane J, Hessen DO, Andersen T. Plasticity in algal stoichiometry: {Experimental} evidence of a temperature-induced shift in optimal supply {N}:{P} ratio. Limnology and Oceanography. 2017;62(4):1346-1354
204.Domenighini A, Giordano M. Fourier transform infrared spectroscopy of microalgae As a novel tool for biodiversity studies, species identification, and the assessment of water quality. Journal of Phycology. 2009;45(2):522-531
205.Giordano M, Ratti S, Domenighini A, Vogt F. Spectroscopic classification of 14 different microalga species: First steps towards spectroscopic measurement of phytoplankton biodiversity. Plant Ecological Diversity. 2009;2(2):155-164
206.Duygu DY. Identification and characterization of some species of Cyanobacteria, Chlorophyta and Bacillariophyta using Fourier-Transform Infrared (FTIR). Spectroscopy. 2019;2019:20-27
207.Duygu D, Udoh AU, Ozer T, Akbulut A, Erkaya I, Yildiz K, et al. Fourier transform infrared (FTIR) spectroscopy for identification of Chlorella vulgaris Beijerinck 1890 and Scenedesmus obliquus (Turpin) Kützing 1833. African Journal of Biotechnology. 2014;11(16):3817-3824
208.Serive B, Nicolau E, Bérard JB, Kaas R, Pasquet V, Picot L, et al. Community analysis of pigment patterns from 37 microalgae strains reveals new carotenoids and porphyrins characteristic of distinct strains and taxonomic groups. PLoS One. 2017;1(2):2
209.Dean AP, Sigee DC, Estrada B, Pittman JK. Using FTIR spectroscopy for rapid determination of lipid accumulation in response to nitrogen limitation in freshwater microalgae. Bioresource Technology. 2010;101(12):4499-4507
210.Jebsen C, Norici A, Wagner H, Palmucci M, Giordano M, Wilhelm C. FTIR spectra of algal species can be used as physiological fingerprints to assess their actual growth potential. Physiologia Plantarum. 2012;146(4):427-438
211.Singh P, Venkata Mohan S, Mohanty K. Dairy wastewater treatment using Monoraphidium sp. KMC4 and its potential as hydrothermal liquefaction feedstock. Bioresource Technology. 2023;376:128877
212.Ferreira AF, Dias APS, Silva CM, Costa M. Effect of low frequency ultrasound on microalgae solvent extraction: Analysis of products, energy consumption and emissions. Algal Research. 2016;14:9-16
213.Wagner H, Liu Z, Langner U, Stehfest K, Wilhelm C. The use of FTIR spectroscopy to assess quantitative changes in the biochemical composition of microalgae. Journal of Biophotonics. 2010;3(8-9):557-566
214.Marcilla A, Gómez-Siurana A, Gomis C, Chápuli E, Catalá MC, Valdés FJ. Characterization of microalgal species through TGA/FTIR analysis: Application to nannochloropsis sp. Thermochimica Acta. 2009;484(1-2):41-47
215.Arif M, Li Y, El-Dalatony MM, Zhang C, Li X, Salama ES. A complete characterization of microalgal biomass through FTIR/TGA/CHNS analysis: An approach for biofuel generation and nutrients removal. Renewable Energy. 2021;163:1973-1982
216.Almutairi AW, El-Sayed AEKB, Reda MM. Evaluation of high salinity adaptation for lipid bio-accumulation in the green microalga Chlorella vulgaris. Saudi Journal of Biological Science. 2021;28(7):3981-3988
217.Kurnia D, Yuliantini A, Cendana IS, Nurachman Z. Fatty acid analysis of marine microalgae Chlorella vulgaris in modified medium used GC-FID. Journal of Physics Conference Series. 2019;1338(1):012007
218.Shaima AF, Mohd Yasin NH, Ibrahim N, Takriff MS, Gunasekaran D, MYY I. Unveiling antimicrobial activity of microalgae Chlorella sorokiniana (UKM2), Chlorella sp. (UKM8) and Scenedesmus sp. (UKM9). Saudi Journal of Biological Science. 2022;29(2):1043
219.Doomun SNE, Loke S, O’Callaghan S, Callahan DL. A simple method for measuring Carbon-13 fatty acid enrichment in the major lipid classes of microalgae using GC-MS. Metabolites. 2016;6(4)
220.Tarazona Delgado R, Guarieiro M, Antunes PW, Cassini ST, Terreros HM, Fernandes V. Effect of nitrogen limitation on growth, biochemical composition, and cell ultrastructure of the microalga Picocystis salinarum. Journal of Applied Phycology. 2021;33(4):2083-2092
221.Akase S, Yoshikawa T, Sakata T. Pigment analysis of marine microalgae by TLC and HPLC methods. Japanese Society for Aquaculture Science. 1999;47(4):511-518
222.Schlüter L, Lauridsen TL, Krogh G, Jørgensen T. Identification and quantification of phytoplankton groups in lakes using new pigment ratios – A comparison between pigment analysis by HPLC and microscopy. Freshwater Biology. 2006;51(8):1474-1485
223.van Leeuwe MA, Villerius LA, Roggeveld J, Visser RJW, Stefels J. An optimized method for automated analysis of algal pigments by HPLC. Marine Chemistry. 2006;102(3-4):267-275
224.Irigoien X, Meyer B, Harris R, Harbour D. Using HPLC pigment analysis to investigate phytoplankton taxonomy: The importance of knowing your species. Helgoland Marine Research. 2004;58(2):77-82
225.Lourenço-Lopes C, Fraga-Corral M, Garcia-Perez P, Carreira-Casais A, Silva A, Simal-Gandara J, et al. A HPLC-DAD method for identifying and estimating the content of fucoxanthin, β-carotene and chlorophyll a in brown algal extracts. Food Chemistry Advances. 2022;1:100095
226.Giovannetti R, Alibabaei L, Zannotti M, Ferraro S, Petetta L. HPLC-DAD-ESI/MS identification of light harvesting and light screening pigments in the lake sediments at Edmonson point. The Scientific World Journal. 2013;2013:1-9
227.Bisht B, Kumar V, Gururani P, Tomar MS, Nanda M, Vlaskin MS, et al. The potential of nuclear magnetic resonance (NMR) in metabolomics and lipidomics of microalgae- a review. Archives of Biochemistry and Biophysics. 2021;710:108987
228.Sarpal AS, Teixeira CMLL, Silva PRM, da Costa Monteiro TV, da Silva JI, da Cunha VS, et al. NMR techniques for determination of lipid content in microalgal biomass and their use in monitoring the cultivation with biodiesel potential. Applied Microbiology and Biotechnology. 2016;100(5):2471-2485
229.Sarpal AS, Teixeira CMLL, Silva PRM, Lima GM, Silva SR, Monteiro TV, et al. Determination of lipid content of oleaginous microalgal biomass by NMR spectroscopic and GC-MS techniques. Analytical and Bioanalytical Chemistry. 2015;407(13):3799-3816
230.Aguilera-Sáez LM, Abreu AC, Camacho-Rodríguez J, González-López CV, Del Carmen C-GM, Fernández I. NMR metabolomics as an effective tool to unravel the effect of light intensity and temperature on the composition of the marine microalgae Isochrysis galbana. Journal of Agricultural and Food Chemistry. 2019;67(14):3879-3889
231.Rinawati M, Sari LA, Pursetyo KT. Chlorophyll and carotenoids analysis spectrophotometer using method on microalgae. IOP Conference Series Earth Environmental Science. 2020;441(1):1-21
232.Picazo A, Vicente C, Miracle E, Rosa M, Antonio C. Spectrophotometric methods for the determination of photosynthetic pigments in stratified lakes: a critical analysis based on comparisons with HPLC determinations in a model lake. Limnetica. 2013;15(32):139-158
233.Schlüter L, Møhlenberg F, Kaas H. Temporal and spatial variability of phytoplankton monitored by a combination of monitoring buoys, pigment analysis and fast screening microscopy in the {Fehmarn} {Belt} {Estuary}. Environmental Monitoring and Assessment. 2014;186(8):5167-5184
Written By
Amal Badr and Walid Fouad
Submitted: 30 June 2023Reviewed: 07 July 2023Published: 25 August 2023