Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
Phytohormones are chemicals released by plants for several mechanism which includes growth and development such as cell divisions, cell elongation and tissue differentiation, it also helps in stress tolerance and senescence. Major phytohormone groups include auxin, cytokinin, gibberellin, ethylene, abscisic acid, brassinosteroids and jasmonates. Phytohormones are naturally produced in low concentration. Certain naturally available soil microorganisms produce phytohormones, the current approach of plant growth regulators to crops improve yield by dual activity and genetic modifications is highly beneficial. The pilot study on metagenomic analysis on commercially important crops helped us to expand the study on identifying the nitrogen fixing bacteria also promoting phytohormone production. Expected outcome: Agrobacterium, Azospirillum, Bacillus, Enterobacter, Pseudomonas, Proteus, Klebsiella and Mycorrhizal are microorganisms that play dual activity. All these growth-promoting bacteria are proven to be involved in indole-3-acetic acid pathways which help in the biosynthesis of auxin and cytokinin. The dual benefit of the plant-growth promoting bacteria is that it can act as a diazotroph which helps in nitrogen fixation as well as the biosynthesis of phytohormones. Several microorganisms play crucial role in plants as nitrogen-fixing bacteria, phytohormone production, etc. they play multiple function in plant growth and development. These are essential microbes in application field of agriculture and biotechnology.
Department of Biotechnology, RV College of Engineering, Bangalore, Karnataka, India
Pooja Sureshkumar
Department of Biotechnology, RV College of Engineering, Bangalore, Karnataka, India
Likitha Shankara
Department of Biotechnology, RV College of Engineering, Bangalore, Karnataka, India
Gulab Khedkar
Department of Biotechnology, Biotechnology Industry Research Assistance Council (BIRAC), New Delhi, India
Jitendra Kumar*
Department of Zoology, Paul Hebert Centre for DNA Barcoding and Biodiversity Studies, Dr. Babasaheb Ambedkar Marathwada University, Aurangabad, Maharashtra, India
*Address all correspondence to: vidya.n@rvce.edu.in, director@bioinnovationcentre.com and jkumar110@yahoo.com
1. Introduction
Phytohormones are well known as Plant hormones. They play a vital role in regulating various physiological processes in plants. These chemical substances act as signaling molecules, coordinating the growth and development of plants [1]. Phytohormones are produced in one part of the plant and then transported to different sites where they elicit specific responses [2].
Phytohormones play a distinct role in plant growth and development. Auxins are responsible for cell elongation and root formation. They help plants bend towards light sources and facilitate the development of lateral roots [3, 4]. Cytokinins, on the other hand, promote cell division and delay senescence, allowing plants to maintain their youthful vigor. They are essential for proper shoot and leaf growth [5, 6].
Gibberellins are phytohormones that stimulate stem elongation and seed germination. They also regulate various developmental processes, such as flowering and fruit development [7]. Abscisic acid is another important hormone that regulates seed dormancy and plays a crucial role in plant responses to stress conditions, such as drought and high salinity [8].
Ethylene, often referred to as the “aging hormone,” influences fruit ripening and senescence. It is involved in the abscission (shedding) of leaves, flowers, and fruits [9]. Brassinosteroids, on the other hand, are involved in cell expansion and differentiation, contributing to overall plant growth and development [10].
In addition to these well-known phytohormones, there are other plant growth regulators worth mentioning. Polyamines (PAs) and karrikins (KARs) are small chemical molecules with less understood production and receptors. Although their precise roles are still being explored, they have been found to influence plant growth, development, and responses to environmental stimuli [2].
The long-term evolution of microorganisms in plant and its surrounding environment has acquired numerous beneficial strategies in enhancing the growth and development. The interaction of microbes with plants modulates in biotic and abiotic stresses. These microbes indirectly have a control over the phytohormone production [11].
Piriformospora indica is a root fungus that interferes with the synthesis and signaling of plant hormones that promote growth, the onset of flowering, differentiation, and systemic immunological responses. In reaction to the microbes, plants modify the hormone levels in their roots to regulate colonization and fungal growth [12].
Advance studies have proven the existence of microorganisms such as Pseudomonas, Enterobacter, Azospirillum and Klebsiella can stimulate the phytohormone production [13, 14].
Therefore, the chapter aims to explain beneficial microorganism and their mechanism in producing phytohormone as well as nitrogen fixation in plants, this will act as dual benefit to plants and reduce additional burden to farmers. The study of these microbes has extensive advantages in agricultural biotechnology.
The study of phytohormones has a rich and interesting historical background. The concept of plant hormones emerged in the late 19th and early 20th centuries when scientists began to observe and investigate the remarkable growth and developmental processes in plants. These scientists sought to understand the underlying factors that influenced plant growth and responses to various environmental stimuli.
The initial breakthrough in understanding phytohormones came with the discovery of auxins. In the late 19th century, Charles Darwin and his son Francis, observed that the tips of coleoptiles in grass seedlings showed bending towards a light source. This phenomenon, known as phototropism, led to the identification of a substance that was responsible for this growth response [15, 16]. In the 1920s, the Dutch scientist Frits Went isolated and characterized this substance, which he named auxin. This discovery marked the beginning of the era of plant hormone research [17, 18].
In 1930s, Japanese scientist Eiichi Kurosawa identified a group of compounds produced by fungi that caused excessive elongation of rice plants. These compounds were later named gibberellins, and their role in promoting stem elongation and flowering [19]. In the 1950s, another group of hormones, cytokinins, was discovered by Folke Skoog and colleagues, who found that these compounds stimulated cell division and played a crucial role in plant tissue culture [20, 21].
The identification of other plant hormones continued with the discovery of abscisic acid (ABA) in the 1960s, which was found to regulate plant responses to stress and dormancy. Ethylene, a gaseous hormone involved in fruit ripening and senescence, was discovered in the 20th century as well [22]. In more recent years, other classes of plant hormones, such as brassinosteroids, jasmonates, and strigolactones, have been identified and extensively studied [10, 23].
The understanding of plant hormones and their roles in plant growth and development has revolutionized the field of plant biology and agriculture. It has provided insights into how plants respond and adapt to their environment, and has led to the development of applications in crop production, horticulture, and plant biotechnology.
3. Classification of phytohormones and their mechanism
Phytohormones are extensively studied to understand their function and mechanism in plant physiology. Recent research has investigated on their biosynthesis, transport, perception and signal transduction pathways.
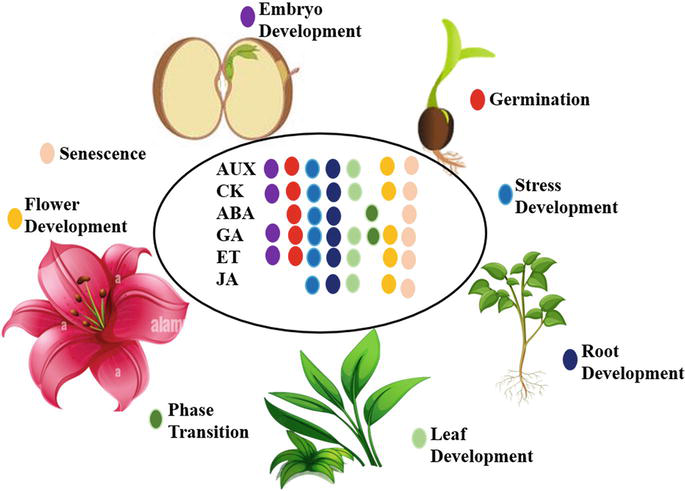
Additionally, there are different pathways in which the phytohormones play a key role which includes: Cell Division and Growth, Apical Dominance, Root Development, Tropisms, Seed Germination and Dormancy, Flowering, Fruit Ripening and Senescence and Stress responses. (Figure 1 and Table 1) [24].
Figure 1.
Representation of phytohormones and their function. The six classes of phytohormones and the plant development processes.
Title
Phytohormone and their mechanism
Cell division and growth
Auxin and Gibberellins promote stem elongation and cell division [25]. Cytokinins stimulate the cell division and also promote shoot and root growth [5].
Apical dominance
Auxin stimulate the dominance of apical meristem, over lateral buds, controlling the growth and branching pattern [18]
Auxin regulate phototropism and gravitropism, Ethylene influences the triple response in plants, causing change in growth and bending during gravitropism and phototropism [26]
Seed germination and dormancy
Gibberellin promotes seed germination and break seed dormancy [7]
Flowering
Gibberellins are involved in regulation of flowering, particularly in transition from vegetative to reproductive growth [27]
Fruit ripening and senescence
Ethylene is key regulator of fruit ripening and senescence processes, while Abscisic acid influences the timing of seed dormancy and germination and promote leaf and fruit senescence [28]
Stress responses
Abscisic acid is involved in response to drought stress, regulating stomatal closure and promoting water conservation [8] Jasmonates are involved in plant defense responses against insects, pathogens and other biotic stresses [29]
4. Exploiting microbial potential in production of phytohormones
The advanced research shows that plant-microbes plays a vital role in inducing plant growth promoters i.e., phytohormones (Figure 2). In addition to the production of hormones by plants these microorganisms also produce, stimulate or involve in the mechanism of converting compounds that are similar to phytohormones. These microorganisms are naturally present in the fertile soil which usually involved in supplying nutrition to the plants. Due to the high nutritional content of root exudates, plant roots are extensively populated with microorganisms. They also produce antifungal substances, enzymes, and suitable solutes. These microbial metabolites are essential for plant development, growth, and nourishment. In response to drought, salt, high heat, and heavy metal toxicity, microbial phytohormones play a critical role in altering root shape. They also have an impact on the metabolism of endogenous growth regulators in plant tissue [30, 31].
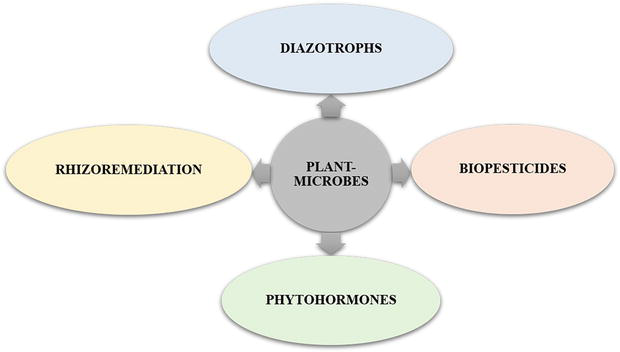
Figure 2.
Classification of plant-growth promoting microbes. The plant microbes play multiple role that includes diazotrophs, biopesticides, phytohormones and rhizoremediation which are highlighted.
The pilot study on various commercially important crops such as wheat, rice, maize, soyabean, peanut etc. has helped us to conclude that certain microorganisms can do multi-function in enhancing the plant as it can improve the plant health, provide nutrient efficiency and resistance to biotic and abiotic stresses [32, 33, 34]. These can also facilitate the symbiotic relationship between the nitrogen fixation by rhizobia or mycorrhizal associations which also mediate the production of phytohormones.
Benefit of using microorganism to produce phytohormones:
Cost-effective: Bacteria and fungi are used as efficient and low-cost production systems for phytohormones. They can be grown in large-scale fermentation processes, which are often more economically viable than the extraction or synthesis of natural or synthetic hormones [35, 36, 37].
Sustainable and eco-friendly: Phytohormones produced by microbes are sustainable alternative to the hormones produced by the plant or the artificial-hormones supplemented to plants. It reduces the need for plant material collection, minimizing the impact on natural resources. Moreover, microbial production can be carried out in controlled environments, reducing the use of chemical solvents and minimizing the release of pollutants [38].
Increased availability and scalability: Microbial production allows for consistent and scalable production of phytohormones. With appropriate cultivation and optimization techniques, large quantities of phytohormones can be generated, ensuring a steady and reliable supply for agricultural applications [39].
Manipulation and customization: Microorganisms can be genetically engineered or modified to produce specific phytohormones or variants with enhanced properties are the advanced technique to optimized characteristics, such as increased stability, activity, or specificity.
Reduced dependence on scarce resources: Some phytohormones, especially rare or complex natural compounds, may be challenging to obtain from plant sources due to their low abundance or difficulty in extraction. Microbial production provides an alternative source for these compounds, reducing the dependence on scarce plant resources.
Quality control and purity: Microbial production allows for precise control over the production process, ensuring consistent quality and purity of the phytohormones. This is particularly important for agricultural applications, where the effectiveness and reliability of the hormones play a crucial role in crop growth and development.
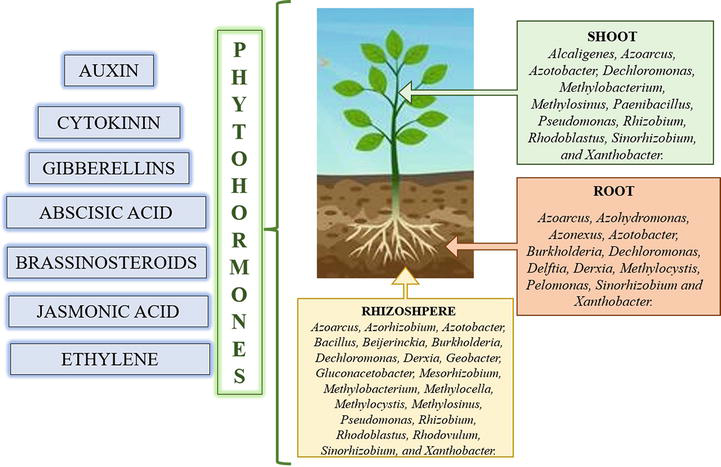
Understanding the plant-microbes and their mechanism in producing the phytohormones which offers promising avenues for sustainable agriculture, biocontrol strategies and improving plant performance in various ecosystem (Figure 3).
Figure 3.
Illustration of microbes and phytohormones. Right side of the image shows microbes which is present in shoot, root and rhizosphere. And left side of the image shows phytohormones produced by the microbes.
Biosynthesis of phytohormones refers to the natural process by which plants produce these vital chemical compounds. The biosynthesis of phytohormones occurs within specific cells or tissues of plants, where enzymes and biochemical pathways catalyze the production of these hormones’ molecules [40, 41, 42]. The microbial biosynthesis of phytohormones holds great promise for various applications in agriculture and biotechnology [43]. Using microorganisms are sustainable and eco-friendly approaches to enhance the crops productivity, improve stress tolerance and optimize plant growth.
5.1 Auxin (AUX)
Auxins are one of the most essential plant hormones it plays curial role in growth and development of the plants. Auxin has an impact on a variety of leaf development processes, including initiation, growth ax specification, morphogenesis, and marginal patterning [4, 18]. Although it is evident that some of the processes by which auxin exerts significantly various across plant species.– Modification in auxin distribution has an impact on leaf and leaflet initiation, adaxial-abaxial polarity, and marginal patterning in many plant species. The placement of serrations and the initiation of leaflets and lobes from the border of leaf primordia are determined by auxin, which also coordinates the phyllotaxis of leaf initiation from the shoot apical meristem (SAM). Auxin and cytokinin, which is crucial for SAM maintenance, are both involved in leaf initiation [6].
Auxin usually obtained as natural and synthetic form. The natural form of auxins is Indole-3-acetic acid (IAA), Indole-3-butyric acid (IBA) and 4-chloroindole-3-acetic acid (4-Cl-IAA). The synthetic form of auxins is Naphthaleneacetic acid (NAA), 2,4-Dichlorophenoxyacetic acid (2,4-D), 1-Naphthaleneacetamide (NAD) and 2-Naphthoxyacetic acid (NOA) [18].
Recent studies have shown certain genes and biochemical reactions interfere in the auxin biosynthesis. Auxin are essential for the following processes; gametogenesis, embryogenesis, seedling growth, vascular patterning and flower development. Auxin biosynthesis has the most complexity as it contributes to several pathways to de novo auxin production. Indole-3-acetic acid (IAA) can also be released from IAA conjugates by hydrolytic cleavage of IAA-amino acids, IAA-sugars and IAA methyl ester. The evolutionarily conserved core mechanisms for auxin biosynthesis may differ from plant species which can have unique strategies and modification to optimize their IAA biosynthesis [44]. To overcome the cost burden and to promote dual efficiency certain bacteria are also involved in producing phytohormones such as Agrobacterium and Pseudomonas are known as Tryptophan-2-monooxygenase (iaaM) these convert the tryptophan to indole-3-acetamide (IAM) which is subsequently hydrolyzed into IAA by the hydrolase iaaH. IAA can also be produced by try through indole-3-pyruvate pathways (IPA).
The IPA decarboxylase has been cloned from Enterobacter cloacae and Azospirillum brasilense, but the genes that catalyze the conversion of tryptophan to IPA and the enzymes that catalyze the conversion of indole-3-acetaldehyde to IAA have not been definitively identified in microorganisms. The function of IPA decarboxylase produced by microbes in plants are yet to be investigation [3, 18]. Thus, plant growth promotor microbes are highly essential for the production of phytohormones to reduce the chemicals or artificial hormones which are supplemented.
5.2 Cytokinins (CK)
Cytokinin is another crucial plant-hormones which regulate the development of cell division and cytokinesis in roots and shoots. Cytokinin might be thought of as a “juvenility” element in leaf development, increasing morphogenesis and postponing differentiation and senescence. The plant hormone cytokinin (CK) participates in orchestrating a variety of growth-related activities in the plant, including the constructive control of SAM size and activity. Major cell specification processes are regulated by auxin and cytokinin during embryogenesis [6]. A brief antagonistic interaction between auxin and cytokinin is indicated as the role of cytokinin in early development.
Cytokinin are divided into two groups adenine-type and phenylurea-type cytokinins. The adenine cytokinin groups includes natural and synthetic compounds such as zeatin, isopentenyl adenine, kinetin, and 6-benzylaminopurine, while phenylurea-type includes synthetic compounds such as diphenylurea and thidiazuron all these molecules exhibit biological activity on plant tissues [45]. Several physiological and biological processes, such as the development of roots, shoots, leaves, controlling apical dominance in the shoot, chloroplast development, and organ formation in a variety of tissues are all impacted by the stimulatory or repressive actions of cytokinins. Senescence is also delayed by an increase in chlorophyll [21].
Different rhizobacterial strains that produce cytokinins as phytohormones include Halomonas desiderata, Proteus mirabilis, P. vulgaris, Klebsiella pneumoniae, Bacillus megaterium, B. cereus, B. subtilis, and Escherichia coli [45].
The loss of cytokinins during a drought may actually harm drought-stressed plants. Inoculating well-watered plants with Bacillus IB-22 efficiently increases shoot zeatin and its riboside and strengthens dependability, shorter root length and increased root mass in the inoculated cabbage revealed that the root axes had swollen, most likely as a result of a post-transcriptional acceleration of ethylene production. It was also indicated that the majority of cytokinin accumulation occurred in the shoot rather than the roots [46].
The cytokinin route and auxin function are closely related, and both hormones are known to regulate the beginning of lateral organs from stem cell niches, a behavior that is well-described during root development and apical dominance in plants. To balance cell division and differentiation, auxin and cytokinin work in opposition to one another. In contrast to cytokinin, which encourages cell differentiation towards the elongation zone by suppressing polar auxin transport, auxin maintains stem cell division in the root apical meristem.
5.3 Gibberellins (GA)
Biologically active gibberellins (bioactive GAs) regulate many different aspects of plant growth and development, such as seed germination, stem lengthening, leaf enlargement, and the production of flowers and seeds [27]. The four main bioactive GAs: GA1, GA3, GA4, and GA7.
In plants, terpene synthases (TPSs), cytochrome P450 monooxygenases (P450s), and 2-oxoglutarate-dependent dioxygenases (2ODDs) are necessary for the production of bioactive Gibberellins from trans-geranylgeranyl diphosphate (GGDP). There are multiple possible ways that Gibberellins are metabolically degraded. The 2-hydroxylation reaction, which is catalyzed by a family of 2ODDs called Gibberellins 2-oxidases (GA2oxs), is the best understood deactivation process. Gibberellins are created in the embryo and subsequently transferred to the aleurone cells during the post germinative development of cereal grains. In these cells, −amylase gene expression is stimulated to facilitate the hydrolysis of endosperm starch [7].
Rhizobium spp enhance nitrogen fixation and also influence in plant hormonal balance which includes gibberellin metabolism [47]. Mycorrhizal fungi enhance the nutritional uptake and also plays a role in gibberellin biosynthesis [48]. Bacillus and Pseudomonas which are plant growth promoting rhizobacteria stimulate the plant growth and development and also enhance the hormonal signaling pathways which can indirectly affect gibberellins production and metabolism in plants [49].
5.4 Abscisic acid (ABA)
Abscisic acid (ABA) is a phytohormone which is essential for adapting to stress conditions like drought and excessive salt. Under water stress, it builds up in plant cells, encourages stomatal closure in guard cells, and controls the production of several genes whose by-products may shield vegetative tissues from dehydration or excessive osmotic pressure. Furthermore, abscisic acid is crucial for numerous developmental phases, including seed maturation and dormancy [8].
Abscisic acid has the ability to modify the biosynthesis of ethylene and auxin, it also influences in related gene expression which altogether regulates fruit ripening in climacteric species [28].
The hormone may slow (late effect in immature fruit) or hasten (early effect in more mature fruit) ripening depending on fruit developmental stage and, consequently, on flesh texture and physiological status, as shown by repression or enhancement of ethylene-, cell wall-, and auxin-related ripening induced gene expression, respectively. By influencing ethylene biosynthesis at a certain developmental stage, which affects metabolic and signaling cross-talk, abscisic acid may either weaken or augment ethylene/auxin action [50].
5.5 Brassinosteroids (BRs)
Brassinosteroids (BRs) are plant-specific steroidal hormones that play a variety of roles in monitoring a wide range of plant growth and developmental processes, including seed germination, cell elongation, cell division, senescence, vascular differentiation, reproduction, root development, sex differentiation, seedling gravitropism, fruit ripening and photomorphogenesis and also responding to various biotics and abiotic stresses [51]. They are distinguished by their polyhydroxylated sterol structure. Catharanthus roseus cell lines in culture were originally used to unravel the biosynthesis routes of Brassinosteroids. Bioactive brassinosteroids, such as brassinolide, bind to an island domain of the BRI1 receptor kinase, which is located on the plasma membrane, to initiate BR signaling. The synthetic auxin 2, 4-dichlorophenoxyacetic acid (2,4-D) greatly enhanced DWF4 expression, notably in the root tips, according to physiological tests utilizing a DWF4pro: GUS transgenic line. Auxin induces DWF4 expression, which in turn raises the number of brassinosteroids via the auxin signaling pathway. DWF4 is a crucial flux-determining enzyme that regulates the endogenous level of Brassinosteroids [52].
Studies showed that the BR biosynthesis route is a triterpenoid pathway after extensive research. The triterpenoid process uses evalonic acid as a precursor. It is compressed and changed into 2,3-oxidosqualene, which then goes through further modifications to create important plant sterols including sitosterols and campesterols. In order to create brassinolide (BL), the most active form of BR, campesterol is transformed to campestenal [53, 54].
The main receptor, BRI1, has a widespread expression pattern, whereas the expression of its two homologs only affects the vasculature and has a more restricted impact on growth. High BR activity promotes SAM cell differentiation and proliferation, which can be induced outside the border zone [55].
A complicated interaction between BRs and GA has been suggested as the cause of BRs’ opposing influence on cell elongation in the leaf sheath in rice. By increasing GA production, physiological levels of BRs cause cell elongation in the leaf sheath. High BR concentrations, on the other hand, prevent cell elongation by inducing GA catabolism when administered exogenously to the leaf sheath [56].
5.6 Jasmonic acid (JA) and ethylene (ET)
Plant defense responses against a variety of diseases, pests, and abiotic stressors such wounding and exposure to ozone are known to be significantly regulated by three phytohormones, SA, JA, and ET. When plants are exposed to pathogens, their salicylic acid levels rise, and exogenous applications lead to the activation of genes associated with pathogenesis (PR) and improved resistance to a wide variety of infections. The defensive signaling network that the plant activates and uses depends on the kind of pathogen and its mechanism of pathogenicity. JA and ET are often involved with defense against necrotrophic diseases and herbivorous insects. JAs are also engaged in a variety of processes, including fruit ripening, leaf senescence, stomatal opening, tendril coiling, root development, tuber formation, and tendril coiling. Coronatine insensitive 1 (COI1), jasmonate resistant 1 (JAR1), and Jasmonate insensitive 1/MYC2 (JIN1/MYC2) are the three primary JA-signaling components [29].
6. Microbes associated in production of phytohormones
Microorganisms in plants do multi-function and these are well known as plant growth-promoting rhizobacteria (PGPR), which is a synonym for PGRs, to describe helpful, free-living bacteria that colonize roots. The term “PGPR” refers to any bacteria that live in plant roots and impact plant growth favorably through a variety of methods, including direct influences like enhanced nutrient absorption and indirect influences like disease suppression (biocontrol). Bacteria from the genera Azotobacter, Azospirillum, Klebsiella, Bacillus, Pseudomonas, Arthrobacter, Enterobacter, and Serratia are included in the well-known PGPR (Table 2) [67].
Present in shoot region of the plant and help in distributing the nutrient and absorbing the essential components [57]
Rheinheimera sp. J3-AN42
It is isolated from younger lateral roots. VFR5–3 (vegetative state of soil fertilized with N roots) has the ability to fix nitrogen and produce IAA and moderately produce cellulase and pectinase [58].
Burkholderia vietnamiensis
They show increased N-fixing as its marked with GUS gene and higher IAA production [59]
Corynebacterium sp
Produced the highest IAA in presences of tryptophan, among the isolates from seven types of rice soils [60]
Azospirillum amazonense
increased dry matter accumulation of rice grain, the number of panicles and nitrogen accumulation at grain maturation [61]
Azospirillum brasilense
Increase yield of rice and also promote cell elongation, root development and tropic response [62]
They showed highest acetylene reduction (ARA) activity and significant biocontrol activity with complete inhibition of sclerotium germination compared to control and other test isolates. In the quantitative estimation of GA endophytic isolate L1 showed the highest value [64].
Detected with highest protease activity and highest IAA production [65]
A.tumefaciens, Klebsiella sp., Ochrobactrum anthropic, P. stutzeri, Pseudomonas sp.
Bacteria’s has ability to exhibit catalase activity and produce ammonia, synthesize IAA Phosphate solubilizing activity, nitogenase activity, ACC deminase activity, significant increase in growth and salt tolerant capacity compared to positive control [66]
Table 2.
Function/mechanism of the microbes.
Diazotrophic bacteria are also members of the PGPR due to their capacity to transform nitrogen into usable ammonia for plants [68]. Therefore, apart from nitrogen fixation they also contribute in synthesis of phytohormone. There have been reports of diazotrophs in a number of bacterial genera, including Acetobacter, Azoarcus, Azospirillum, Azotobacter, Beijerinckia, Burkholderia, Enterobacter, Herbaspirillum, Klebsiella, Paenibacillus, and Pseudomonas, which may be loosely or more closely (endophytes) associated with plants [69].
Endophytic diazotrophs can be discovered in the roots, stems, leaves, and even seeds of plants. Through processes like chemotaxis, biofilm formation, and the secretion of compounds that stimulate plant growth, they are able to colonize these tissues. Tropical plants including sugarcane, maize, and rice have been shown to contain exceptionally high concentrations of endophytic diazotrophs. They are crucial in these plants for encouraging plant development and improving soil nitrogen fixation.
Di-nitrogenase and Di-nitrogen reductase, two proteins that work together to catalyze the conversion of atmospheric nitrogen to ammonia, make up the complex enzyme system known as nitrogenase. A wide range of diazotrophs are capable of converting atmospheric nitrogen into a form that may be used by plants and other living things. They are essential components of the global nitrogen cycle, supplying the biosphere with nitrogen and preserving life as we know it [70]. Piriformospora indica is an endophytic root fungus which stimulate the production of phytohormones. The mechanism of drought and salt tolerance mediated by fungi in plants involves in the production of cytokinin and abscisic acid. This fungus also stimulate lateral root growth and increase root surface area by producing auxin and gibberellin, this helps the plant to take up more water and minerals during stresses, thereby enhancing the yield [12].
Azospirillum, Rhizobium, and Klebsiella species are some of the diazotrophs that are most frequently utilized as bio-fertilizers. The process of transforming atmospheric nitrogen gas into a form that plants can utilize is known as nitrogen fixation, and it is promoted by these bacteria’s ability to colonize the roots of plants and develop symbiotic partnerships. Diazotrophs can boost plant growth and production in a number of crops, including rice, maize, and wheat, according to studies on their usage as bio-fertilizers. They have also been demonstrated to increase the sustainability of agricultural practises, increase soil fertility, and decrease environmental pollution from synthetic fertilizers [38, 71, 72]. These bacteria are carefully studied to understand their mechanism as diazotrophs and phytohormone synthesizer. Azospirillum, Rhizobium and Klebsiella are essential in root promotion and increase IAA production in plants, therefore they contribute in dual benefits.
The most prevalent diazotrophs identified in rice plants are individuals from the genera Azospirillum, Herbaspirillum, and Burkholderia. It is known that these organisms colonize the roots of rice plants and encourage nitrogen fixation by activating nitrogenase enzymes [71, 73].
The concentration of the most significant plant hormones generated by Azospirillum species, such as auxins (IAA), cytokinins (Z), and gibberellins (GA3), rises during the exponential and/or stationary growth phase. By changing the homeostasis of the phytohormones, co-inoculation with rhizobacteria that produce them might affect the result of the symbiotic relationship. Researchers have focused on this genus to create particular strains as inoculants for use in agriculture since Azospirillum sp. has beneficial impacts on the development of various plant species [74, 75, 76, 77].
An experiment was conducted on the availability of tryptophan as precursors in which IAA concentration produced by Azospirillum was monitored. The amount of IAA generated is significantly influenced by the pH of the culture medium. After the carbon source in the medium used for batch culture has been depleted, the stationary phase of the bacterium cells controls the release of huge quantities of IAA by Azospirillum spp. cultures [78]. Several PGPR have the ability to enhance the plant development, heavy metal stress such as Arsenic (As) adversely affects certain cascade pathways and phytohormonal signaling stimulations [79]. An experiment was conducted on wheat and sunflower plant to examine the arsenic toxicity level. The results conclude, plants produced more biomass when Bradyrhizobium japonicum CB1809, was present in the medium which was also quite tolerant of various arsenic concentration [80].
1-aminocyclopropane-1-carboxylate (ACC) deaminase-producing rhizobacteria that promote plant growth help plants grow and develop by lowering ethylene levels, increasing salt tolerance, and lessening drought stress. A large number of rhizobacteria consume the ethylene precursor ACC and transform it into 2-oxobutanoate and NH3. Plant roots elongate, shoot development is encouraged, rhizobia nodulation and N, P, and K intake are improved, and mycorrhizal colonization occurs in a variety of crops as a result of seed/root inoculation with ACC deaminase-producing rhizobacteria [81, 82, 83].
Thus, bacteria are the most crucial component which play multiple function in plant’s growth and development. Azospirillum is the most widely studied organism concerning nitrogen fixation and phytohormone production such as auxin. There are several such organism which play multi-function in plant which are described in Table-2. Microorganisms are equally important in plant, research has proven they functions as biofertilizers, biopesticides, natural decomposers, nitrogen fixation bacteria, plant growth promotors and many more [84]. Genetic engineering is an operation tool to enhance the mechanism of these microbes to produce the desirable products [85]. Therefore, their great potential for the application field of agriculture and biotechnology.
Plant Growth Regulation: Phytohormones are used as plant growth regulators (PGRs) to manipulate plant growth and development. They can control plant height, enhance branching, promote flowering, regulate fruit development, and improve overall plant architecture [86].
Seed Treatment and Germination: Phytohormones are applied to seeds to enhance germination rates, overcome dormancy, and promote uniform seedling emergence. This is particularly useful for improving the establishment of crops with low germination rates or challenging environmental conditions.
Fruit Ripening and Shelf-Life Extension: Phytohormones, such as ethylene, are involved in fruit ripening and senescence. By regulating ethylene levels using inhibitors or ethylene-releasing compounds, the post-harvest shelf life of fruits can be extended, reducing spoilage and maintaining product quality.
Flowering Induction: Phytohormones can be used to induce flowering in certain crops. By applying specific hormones at the appropriate stage, flowering can be promoted, leading to synchronized and increased flower production.
Crop Yield Improvement: Phytohormones play a crucial role in optimizing crop yields. By understanding the hormonal regulation of plant growth and development, farmers can apply phytohormones strategically to maximize productivity and improve crop quality [39].
Stress Tolerance: Phytohormones, such as abscisic acid (ABA), play a role in plant responses to various stresses, including drought, salinity, and temperature extremes. By applying ABA or other stress-related hormones, plants can exhibit improved tolerance and resilience in challenging environmental conditions [37].
Plant Tissue Culture: Phytohormones are essential in plant tissue culture techniques for in vitro propagation and regeneration of plants. They stimulate cell division, callus formation, and differentiation, allowing for the production of large numbers of uniform plantlets [87].
Weed and Pest Control: Phytohormones can be used as part of integrated pest management strategies to control weeds and pests. By manipulating hormone levels, it is possible to disrupt their growth and reproductive processes, offering an alternative approach to chemical pesticides [39].
Root Development and Nutrient Uptake: Phytohormones, particularly auxins, play a crucial role in root development and nutrient uptake. By applying auxins, root growth can be stimulated, leading to enhanced nutrient absorption and overall plant vigor [39].
Environmental Stimulation: Phytohormones can be applied to induce specific responses in plants, such as stimulating bud break, promoting lateral shoot development, or enhancing rooting. These applications are commonly used in horticulture and landscaping practices [86].
Phytohormones regulate the physiological processes in plants. Their ability to influence plant growth, development and responses to environment stimuli. The use of phytohormones which is synthesized by microbes holds higher application in research field. The regulatory power of phytohormones can optimize the plant growth, improve yields, overcome challenges and ensure sustainable and efficient crop production. Auxin, Cytokinin and Gibberellin are used as the plant growth regulator as it helps in enhancing the plant height, widening of branch, promote flowering and also regulate fruits development. It has also been found that it promotes seed treatment and germination which is most challenging environmental conditions. Phytohormones are naturally produced by plants but in low concentration. To enhance the plant growth regulator certain artificially synthesized plant-hormones are supplemented which in-turn affects the plant in long-term. The pilot study of nitrogen fixing bacteria in several commercially important crops has helped us to expand the study on phytohormone producing bacteria. The plant-growth promoting bacteria plays dual benefit for the plants which not only help in nitrogen fixation but also helps in producing phytohormones.
Agrobacterium, Pseudomonas, Enterobacter cloacae and Azospirillum brasilense stimulate in auxin production. Halomonas desiderata, Proteus mirabilis, P. vulgaris, Klebsiella pneumoniae, Bacillus megaterium, B. cereus, B. subtilis, and Escherichia coli influence in cytokinin production while Bacillus, Pseudomonas and Mycorrhizal fungi enhance the nutritional uptake and also plays a role in gibberellin biosynthesis. Nevertheless, Azospirillum is well studied in the production of auxin i.e., IAA productions. There are similar microorganisms which play multiple function in plant growth and development whose mechanism is yet to be discovered.
Genetic Engineering in modification of the microorganism to enhance the yield and productivity has contributed with respect to application in agricultural and biotechnology field. Therefore, these advanced approaches improve resources efficiency, minimizes environmental impacts and maximizes crop yield and quality. This benefits the farmers and society to increase the economic production.
The authors would like to thank Akshay Uttarkar for his crucial insights in improving the presentation. Our heartfelt thanks to the staff and administration at R V College of Engineering for the support.
The authors declare there is no conflict of interest.
Funding
The funding was acquired from the Bangalore Bioinnovation Centre; the Karnataka Innovation and Technology Society; the Department of Electronics, IT, BT and S&T; and the Government of Karnataka, India and was put towards paying the publication cost.
References
1.Altmann M, Altmann S, Rodriguez PA, Weller B, Elorduy Vergara L, Palme J, et al. Extensive signal integration by the phytohormone protein network. Nature. 2020;583(7815):271-276
2.Zhao B, Liu Q , Wang B, Yuan F. Roles of Phytohormones and their Signaling pathways in leaf development and stress responses. Journal of Agricultural and Food Chemistry. 2021;69(12):3566-3584
3.Korasick DA, Enders TA, Strader LC. Auxin biosynthesis and storage forms. Journal of Experimental Botany. 2013;64(9):2541-2555
4.Michniewicz M, Brewer PB, Friml JÍ. Polar auxin transport and asymmetric auxin distribution. Arabidopsis Book. 2007;5:e0108
5.Cortleven A, Leuendorf JE, Frank M, Pezzetta D, Bolt S, Schmülling T. Cytokinin action in response to abiotic and biotic stresses in plants: The role of cytokinin in stress responses. Plant, Cell & Environment. 2019;42(3):998-1018
6.Shwartz I, Levy M, Ori N, Bar M. Hormones in tomato leaf development. Developmental Biology. 2016;419(1):132-142
7.Yamaguchi S. Gibberellin metabolism and its regulation. Annual Review of Plant Biology. 2008;59(1):225-251
8.Umezawa T, Nakashima K, Miyakawa T, Kuromori T, Tanokura M, Shinozaki K, et al. Molecular basis of the Core regulatory network in ABA responses: Sensing, Signaling and transport. Plant and Cell Physiology. 2010;51(11):1821-1839
9.Schaller GE. Ethylene and the regulation of plant development. BMC Biology. 2012;10(1):9
10.Oh MH, Honey SH, Tax FE. The control of cell expansion, cell division, and vascular development by Brassinosteroids: A historical perspective. IJMS. 2020;21(5):1743
11.Yan L, Zhu J, Zhao X, Shi J, Jiang C, Shao D. Beneficial effects of endophytic fungi colonization on plants. Applied Microbiology and Biotechnology. 2019;103(8):3327-3340. DOI: 10.1007/s00253-019-09713-2
12.Xu L, Wu C, Oelmüller R, Zhang W. Role of Phytohormones in Piriformospora indica-induced growth promotion and stress tolerance in plants: More questions than answers. Frontiers in Microbiology. 2018;9:1646
13.Laird TS, Flores N, Leveau JHJ. Bacterial catabolism of indole-3-acetic acid. Applied Microbiology and Biotechnology. 2020;104(22):9535-9550
14.Fukami J, Cerezini P, Hungria M. Azospirillum: Benefits that go far beyond biological nitrogen fixation. AMB Express. 2018;8(1):73
15.Kutschera U, Niklas KJ. Evolutionary plant physiology: Charles Darwin’s forgotten synthesis. Die Naturwissenschaften. 2009;96(11):1339-1354
16.Hurný A, Benková E. Methodological advances in auxin and Cytokinin biology. In: Dandekar T, Naseem M, editors. Auxins and Cytokinins in Plant Biology [Internet]. New York, NY: Springer New York; 2017 [cited June 17, 2023]. p. 1, 29 (Methods in Molecular Biology; vol. 1569). Available from: http://link.springer.com/10.1007/978-1-4939-6831-2_1
17.Pennazio S. The discovery of the chemical nature of the plant hormone auxin. Rivista di Biologia. 2002;95(2):289-308
18.Enders TA, Strader LC. Auxin activity: Past, present, and future. American Journal of Botany. 2015;102(2):180-196
19.Gray RA. Alteration of LEAF size and shape and other changes caused by gibberellins IN plants. American Journal of Botany. 1957;44(8):674-682
20.Eckardt NA. A new classic of Cytokinin research: Cytokinin-deficient Arabidopsis plants provide new insights into Cytokinin biology. The Plant Cell. 2003;15(11):2489-2492
21.Kamínek M. Tracking the story of Cytokinin research. Journal of Plant Growth Regulation. 2015;34(4):723-739
22.Bakshi A, Shemansky JM, Chang C, Binder BM. History of research on the plant hormone ethylene. Journal of Plant Growth Regulation. 2015;34(4):809-827
23.Tamogami S, Agrawal GK, Rakwal R. Jasmonates to Jasmolites in plants. In: Advances in Botanical Research [Internet]. Netherlands: Elsevier; 2011 [cited May 24, 2023]. pp. 309-348. Available from: https://linkinghub.elsevier.com/retrieve/pii/B9780123858511000068
24.Curaba J, Singh MB, Bhalla PL. miRNAs in the crosstalk between phytohormone signalling pathways. Journal of Experimental Botany. 2014;65(6):1425-1438
25.Takatsuka H, Umeda M. Hormonal control of cell division and elongation along differentiation trajectories in roots. Journal of Experimental Botany. 2014;65(10):2633-2643
26.Nziengui H, Lasok H, Kochersperger P, Ruperti B, Rébeillé F, Palme K, et al. Root gravitropism is regulated by a crosstalk between Para -Aminobenzoic acid, ethylene, and auxin. Plant Physiology. 2018;178(3):1370-1389
27.Martins AO, Nunes-Nesi A, Araújo WL, Fernie AR. To bring flowers or do a runner: Gibberellins make the decision. Molecular Plant. 2018;11(1):4-6
28.Zhang M, Yuan B, Leng P. The role of ABA in triggering ethylene biosynthesis and ripening of tomato fruit. Journal of Experimental Botany. 2009;60(6):1579-1588
29.Bari R, Jones JDG. Role of plant hormones in plant defence responses. Plant Molecular Biology. 2009;69(4):473-488
30.Egamberdieva D, Wirth SJ, Alqarawi AA, EF Abd_Allah, Hashem A. Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Frontiers in Microbiology. 2017;8:2104
31.Fahad S, Hussain S, Matloob A, Khan FA, Khaliq A, Saud S, et al. Phytohormones and plant responses to salinity stress: A review. Plant Growth Regulation. 2015;75(2):391-404
32.Niranjan V, Rao A, Janaki B, Uttarkar A, Setlur AS, K C, et al. Molecular docking and interaction studies of identified abscisic acid receptors in Oryza sativa: An In-silico perspective on comprehending stress tolerance mechanisms. Current Genomics 2021;22(8):607-619.
33.Sujitha D, Kumar H, Thapliayal G, Pal G, Vanitha A, Uttarkar A, et al. Transcription factors controlling the expression of oxidative stress associated genes in rice (Oryza sativa L.). Plant Biotechnology Reports. 2023:1-17. DOI: 10.1007/s11816-023-00819-8
34.Pal G, Bakade R, Deshpande S, Sureshkumar V, Patil SS, Dawane A, et al. Transcriptomic responses under combined bacterial blight and drought stress in rice reveal potential genes to improve multi-stress tolerance. BMC Plant Biology. 2022;22(1):349
35.Shi TQ , Peng H, Zeng SY, Ji RY, Shi K, Huang H, et al. Microbial production of plant hormones: Opportunities and challenges. Bioengineered. 2017;8(2):124-128
36.Chiaranunt P, White JF. Plant beneficial Bacteria and their potential applications in vertical farming systems. Plants. 2023;12(2):400
37.Ali S, Xie L. Plant growth promoting and stress mitigating abilities of soil born microorganisms. FNA. 2020;11(2):96-104
38.Bhattacharyya PN, Jha DK. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World Journal of Microbiology and Biotechnology. 2012;28(4):1327-1350
39.Bishnoi U. PGPR interaction. In: Advances in Botanical Research [Internet]. Netherlands: Elsevier; 2015 [cited May 24, 2023]. pp. 81-113. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0065229615000671
40.Costacurta A, Vanderleyden J. Synthesis of Phytohormones by plant-associated Bacteria. Critical Reviews in Microbiology. 1995;21(1):1-18
41.Novák O, Napier R, Ljung K. Zooming In on plant hormone analysis: Tissue- and cell-specific approaches. Annual Review of Plant Biology. 2017;68(1):323-348
42.Kende H, Jad Z. The five “classical” plant hormones. The Plant Cell. 1997;9:1197-1210
43.Stirk WA, van Staden J. Potential of phytohormones as a strategy to improve microalgae productivity for biotechnological applications. Biotechnology Advances. 2020;44:107612
44.Woodward AW. Auxin: Regulation, action, and interaction. Annals of Botany. 2005;95(5):707-735
45.Gupta S, Seth R, Sharma A, Seth R, Sharma A. Plant growth-promoting Rhizobacteria play a role as Phytostimulators for sustainable agriculture. In: Choudhary DK, Varma A, Tuteja N, editors. Plant-Microbe Interaction: An Approach to Sustainable Agriculture [Internet]. Singapore: Springer Singapore; 2016 [cited May 7, 2023]. pp. 475-493. Available from: http://link.springer.com/10.1007/978-981-10-2854-0_22
46.Arkhipova TN, Prinsen E, Veselov SU, Martinenko EV, Melentiev AI, Kudoyarova GR. Cytokinin producing bacteria enhance plant growth in drying soil. Plant and Soil. 2007;292(1-2):305-315
48.Bottini R, Fulchieri M, Pearce D, Pharis RP. Identification of gibberellins a 1 , a 3 , and Iso-a 3 in cultures of Azospirillum lipoferum. Plant Physiology. 1989;90(1):45-47
49.Ryu CM, Hu CH, Locy RD, Kloepper JW. Study of mechanisms for plant growth promotion elicited by rhizobacteria in Arabidopsis thaliana. Plant and Soil. 2005;268(1):285-292
50.Soto A, Ruiz KB, Ravaglia D, Costa G, Torrigiani P. ABA may promote or delay peach fruit ripening through modulation of ripening- and hormone-related gene expression depending on the developmental stage. Plant Physiology and Biochemistry. 2013;64:11-24
51.Gruszka D. The Brassinosteroid Signaling pathway—New key players and interconnections with other Signaling networks crucial for plant development and stress tolerance. IJMS. 2013;14(5):8740-8774
52.Saini S, Sharma I, Pati PK. Versatile roles of brassinosteroid in plants in the context of its homoeostasis, signaling and crosstalks. Frontiers in Plant Science. 2015;6:950. DOI: 10.3389/fpls.2015.00950
53.Chung Y, Choe S. The regulation of Brassinosteroid biosynthesis in Arabidopsis. Critical Reviews in Plant Sciences. 2013;32(6):396-410
54.Fujioka S, Yokota T. Biosynthesis and metabolism of Brassinosteroids. Annual Review of Plant Biology. 2003;54(1):137-164
55.Singh AP, Savaldi-Goldstein S. Growth control: Brassinosteroid activity gets context. Journal of Experimental Botany. 2015;66(4):1123-1132
56.Tong H, Xiao Y, Liu D, Gao S, Liu L, Yin Y, et al. Brassinosteroid regulates cell elongation by modulating gibberellin metabolism in Rice. The Plant Cell. 2014;26(11):4376-4393
57.Roesch LFW, Camargo FAO, Bento FM, Triplett EW. Biodiversity of diazotrophic bacteria within the soil, root and stem of field-grown maize. Plant and Soil. 2008;302(1-2):91-104
58.Prakamhang J, Minamisawa K, Teamtaisong K, Boonkerd N, Teaumroong N. The communities of endophytic diazotrophic bacteria in cultivated rice (Oryza sativa L.). Applied Soil Ecology. 2009;42(2):141-149
59.Govindarajan M, Balandreau J, Kwon SW, Weon HY, Lakshminarasimhan C. Effects of the inoculation of Burkholderia vietnamensis and related endophytic Diazotrophic Bacteria on grain yield of Rice. Microbial Ecology. 2008;55(1):21-37
60.Naher UA, Radziah O, Shamsuddin ZH, Halimi MS, Razi IM. Isolation of Diazotrophs from different soils of Tanjong Karang Rice growing area in Malaysia. International Journal of Agriculture and Biology. 2009;11(5):1560-8530
61.Rodrigues EP, Rodrigues LS, De Oliveira ALM, Baldani VLD, Teixeira KRDS, Urquiaga S, et al. Azospirillum amazonense inoculation: Effects on growth, yield and N2 fixation of rice (Oryza sativa L.). Plant and Soil. 2008;302(1-2):249-261
62.Thakuria D, Talukdar NC, Goswami C, Hazarika S, Boro RC, Khan MR. Characterization and screening of bacteria from rhizosphere of rice grown in acidic soils of Assam. Current Science. 2004;86(7):978-985
63.Rangjaroen C, Rerkasem B, Teaumroong N, Noisangiam R, Lumyong S. Promoting plant growth in a commercial rice cultivar by endophytic diazotrophic bacteria isolated from rice landraces. Annales de Microbiologie. 2015;65(1):253-266
64.Shabanamol S, Divya K, George TK, Rishad KS, Sreekumar TS, Jisha MS. Characterization and in planta nitrogen fixation of plant growth promoting endophytic diazotrophic Lysinibacillus sphaericus isolated from rice (Oryza sativa). Physiological and Molecular Plant Pathology. 2018;102:46-54
65.Patel JK, Archana G. Diverse culturable diazotrophic endophytic bacteria from Poaceae plants show cross-colonization and plant growth promotion in wheat. Plant and Soil. 2017;417(1-2):99-116
66.Sharma S, Kulkarni J, Jha B. Halotolerant Rhizobacteria promote growth and enhance salinity tolerance in Peanut. Frontiers in Microbiology. 2016;7:1600. DOI: 10.3389/fmicb.2016.01600/full
67.Frankenberger WT, Arshad M. Phytohormones in Soils: Microbial Production and Function. New York: M. Dekker; 1995. 503 p (Books in soils, plants, and the environment)
68.Hu J, Richwine JD, Keyser PD, Li L, Yao F, Jagadamma S, et al. Nitrogen fertilization and native C4 grass species Alter abundance, activity, and diversity of soil Diazotrophic communities. Frontiers in Microbiology. 2021;12:675693
69.Dobbelaere S, Vanderleyden J, Okon Y. Plant growth-promoting effects of Diazotrophs in the rhizosphere. Critical Reviews in Plant Sciences. 2003;22(2):107-149
70.González PJ, Correia C, Moura I, Brondino CD, Moura JJG. Bacterial nitrate reductases: Molecular and biological aspects of nitrate reduction. Journal of Inorganic Biochemistry. 2006;100(5-6):1015-1023
71.Döbereiner J, Baldani VLD, Reis VM. Endophytic occurrence of Diazotrophic Bacteria in non-leguminous crops. In: Fendrik I, del Gallo M, Vanderleyden J, de Zamaroczy M, editors. Azospirillum VI and Related Microorganisms [Internet]. Berlin, Heidelberg: Springer Berlin Heidelberg; 1995 [cited June 16, 2023]. p. 3-14. 10.1007/978-3-642-79906-8_1
72.Ahemad M, Kibret M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. Journal of King Saud University - Science. 2014;26(1):1-20
73.Bally R, Thomas-Bauzon D, Heulin T, Balandreau J, Richard C, Ley JD. Determination of the most frequent N2 -fixing bacteria in a rice rhizosphere. Canadian Journal of Microbiology. 1983;29(8):881-887
74.Cassán F, Diaz-Zorita M. Azospirillum sp. in current agriculture: From the laboratory to the field. Soil Biology and Biochemistry. 2016;103:117-130
75.Strzelczyk E, Kampert M, Li CY. Cytokinin-like substances and ethylene production by Azospirillum in media with different carbon sources. Microbiological Research. 1994;149(1):55-60
76.Cohen AC, Bottini R, Piccoli PN. Azospirillum brasilense Sp 245 produces ABA in chemically-defined culture medium and increases ABA content in arabidopsis plants. Plant Growth Regulation. 2008;54(2):97-103
77.Cassán F, Maiale S, Masciarelli O, Vidal A, Luna V, Ruiz O. Cadaverine production by Azospirillum brasilense and its possible role in plant growth promotion and osmotic stress mitigation. European Journal of Soil Biology. 2009;45(1):12-19
78.Costacurta A, Keijers V, Vanderleyden J. Molecular cloning and sequence analysis of an Azospirilium brasilense indole-3-pyruvate decarboxylase gene. Molecular Genetics and Genomics. 1994;243(4):463-472
79.Aymen U, Khan M, Singh R, Parihar P, Pandey N. Role of Phytohormones in regulating arsenic‐induced toxicity in plants. In: Srivastava PK, Singh R, Parihar P, Prasad SM, editors. Arsenic in Plants [Internet]. 1st ed. Canada: Wiley; 2022 [cited June 17, 2023]. pp. 198-222. DOI: 10.1002/9781119791461.ch11
80.Reichman SM. Probing the plant growth-promoting and heavy metal tolerance characteristics of Bradyrhizobium japonicum CB1809. European Journal of Soil Biology. 2014;63:7-13
81.Nadeem SM, Zahir ZA, Naveed M, Arshad M. Rhizobacteria containing ACC-deaminase confer salt tolerance in maize grown on salt-affected fields. Canadian Journal of Microbiology. 2009;55(11):1302-1309
82.Nadeem SM, Zahir ZA, Naveed M, Arshad M. Preliminary investigations on inducing salt tolerance in maize through inoculation with rhizobacteria containing ACC deaminase activity. Canadian Journal of Microbiology. 2007;53(10):1141-1149
83.Arshad M, Saleem M, Hussain S. Perspectives of bacterial ACC deaminase in phytoremediation. Trends in Biotechnology. 2007;25(8):356-362
84.Prakash O, Sharma R, Rahi P, Karthikeyan N. Role of microorganisms in plant nutrition and health. In: Rakshit A, Singh HB, Sen A, editors. Nutrient Use Efficiency: From Basics to Advances [Internet]. New Delhi: Springer India; 2015 [cited June 17, 2023]. pp. 125-161. DOI: 10.1007/978-81-322-2169-2_9
85.Liu Y, Feng J, Pan H, Zhang X, Zhang Y. Genetically engineered bacterium: Principles, practices, and prospects. Frontiers in Microbiology. 2022;13:997587
86.Basra AS, editor. Plant Growth Regulators in Agriculture and Horticulture: Their Role and Commercial Uses. New York: Food Products Press; 2000. p. 264
 Open access peer-reviewed chapter
Open access peer-reviewed chapter