 Open access peer-reviewed chapter
Open access peer-reviewed chapter


Abstract
It is estimated that approximately 185 million people worldwide are infected with hepatitis C virus (HCV). The global prevalence of HCV infection is known as 2–3%. Every year, 350,000 of these patients die from complications such as cirrhosis and HCC associated with chronic hepatitis C. Therefore, early diagnosis and treatment are of great importance. It is important to reach more patients because of the use of direct-acting antivirals that provide nearly 100% permanent viral response in the treatment of HCV. In line with the 2030 target of the World Health Organization for the elimination of hepatitis C, it is important to raise awareness that HCV is a treatable disease. This chapter aims to briefly review the structure and diagnostic methods of HCV.
Keywords
- hepatitis C
- chronic
- hepatitis C antibodies
- core protein p22
- hepatitis C diagnostic methods
1. Introduction
Hepatitis C virus (HCV) is a spherical, enveloped, positive-stranded, single-stranded RNA virus with a diameter of 40–80 nm. It belongs to the
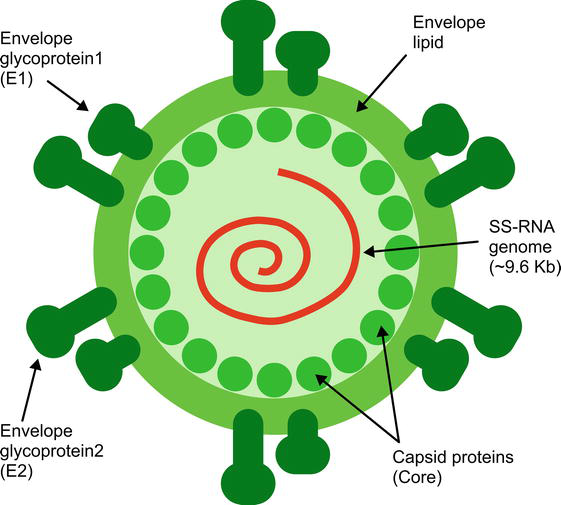
Figure 1.
HCV genome structure.
1.1 HCV genome
A single precursor protein containing 3020 amino acids is synthesized from the virus’s genome of approximately 9600 nucleotides [4]. The genome contains a long open reading frame (ORF) flanked by highly conserved 5’UTR (untranslated region) and 3’UTR regions. HCV genome has structural and nonstructural proteins (Figure 2).
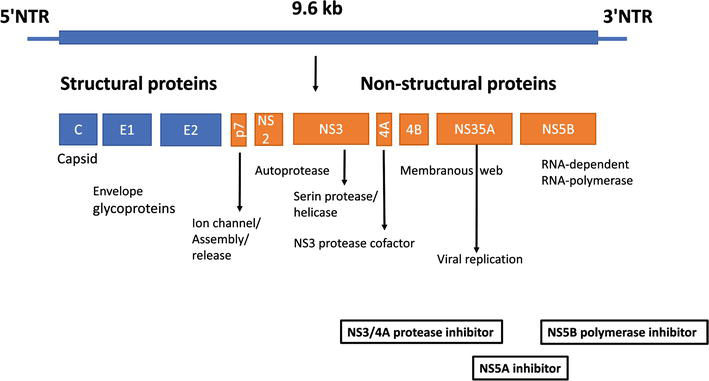
Figure 2.
HCV proteins.
1.2 HCV proteins
1.2.1 Structural proteins
1.2.1.1 Core protein
The HCV core protein is a highly basic RNA-binding protein that makes up the viral nucleocapsid and is released as a 191 amino acid, a precursor of 23-kDa (P23). Although proteins of various sizes (17 to 23 kDa) could be detected, the 21-kDa core protein (P21) appeared to be the predominant form [5]. It has three distinct regions: the N-terminal hydrophilic domain of 120 amino acids (D1), the C-terminal hydrophobic domain of about 50 amino acids (D2), and a single peptide domain of 20 amino acids (D3) that functions as a signal transducer [6, 7, 8]. D1 serves as three predictive nuclear localization signals (NLS) and is predicted to be involved in RNA binding and nuclear localization [9, 10]. D2 is responsible for the core protein relationship with endoplasmic reticulum (ER) membranes, outer mitochondrial membranes, and lipid droplets [10, 11]. It is thought to have a role in hepatosteatosis [12]. The core protein can also be translocated to the nucleus. It regulates the transcription of cellular genes (c-myc and c-fos), protooncogenes (ras) and has apoptotic and antiapoptotic functions [13]. It also has a role in suppressing hepatitis B virus (HBV) replication. Core protein may adversely affect the inhibition of natural killer cells (NK) by increasing the expression of major histocompatibility complex (MHC) class 1 and the immune response to be formed against it by interacting with the complement receptor [14]. In addition, miR-122 inhibits HCV RNA overproduction by reducing expression [15].
1.2.1.2 Envelope glycoproteins
E1 and E2 glycoproteins are major components of the virion envelope and are required for virus entry and fusion [16]. E1 and E2 are type I transmembrane glycoproteins and have a wide variety of functions, including membrane attachment, endoplasmic reticulum localization, and virus packaging [17, 18]. The segment consisting of 30 amino acid residues in the N-terminal region of the E2 envelope glycoprotein is called HVR-1. It is the most genetically diverse of the envelope proteins. Infected individuals often produce antibodies against the HVR-1 sequence. Different subtypes are thought to cause the production of different degrees of reactive HVR-1 antibodies. Available data indicate that HVR-1 has a few neutralizing epitopes, and these epitopes are sites where mutations responsible for immune escape occur during acute and chronic infection [19, 20, 21]. However, the fact that the virus can still infect chimpanzees with the deletion of this region shows that the virus is not critical in cell entry and release.
1.2.2 Non-structural proteins
1.2.2.1 Protein p7
The p7 protein, consisting of 63 amino acids, is an integral membrane protein that acts as a calcium ion channel necessary for the efficient recruitment and release of the proliferating virus [22]. p7 is essential because mutations or deletions in its cytoplasmic portion have suppressed the infectivity of intrahepatic transfection of HCV cDNA in chimpanzees [23]. It is a therapeutic target because it is inhibited by amantadine in vitro [24].
1.2.2.2 Protein NS2
NS2 is a nonstructural transmembrane protein weighing 21–23 kDa. It forms the NS2/3 protease with the N-terminal end of the NS3 protein. The NS2/3 protease is a zinc-dependent metalloprotease, the first of two virus-encoded proteases required for intramolecular cleavage of the HCV polyprotein [25]. It cuts the polyprotein from the NS2/NS3 junction and interacts with both structural and nonstructural proteins and plays a role in virion packaging (assembly) [26, 27].
1.2.2.3 Protein NS3
NS3 is a serine protease, RNA helicase, and nucleoside triphosphatase (NTPase) activity. NS4A is a cofactor for NS3 protease activity. NS3/4A carries additional features through its interaction with host cell pathways and proteins, which may be important in the life cycle and pathogenesis of infection; therefore it is one of the main targets of agents used for therapy [28]. It has recently been shown to antagonize the dsRNA-dependent interferon regulatory factor 3 (IRF-3) pathway, which is an important mediator of interferon induction in response to viral infection [29]. Thus, it is possible for the virus to escape from the natural cellular antiviral defense mechanisms.
1.2.2.4 Protein NS4A
The NS4A protein is a short polypeptide of 54 amino acids and the cofactor of the NS3 serine protease. It places NS3 protease on intracellular membranes through the N-terminal transmembrane segment in its structure, contributes to its correct folding by joining the N-terminal protease region, stabilizes protease against proteolytic degradation, and activates protease activity [30].
1.2.2.5 Protein NS4B
NS4B is an integral membrane protein weighing 27 kDa, which is predicted to contain at least four transmembrane domains. It is in contact with the ER membranes. NS4B has the ability to induce the formation of specialized membrane folds, also called membranous webs, that serve as scaffolds for the HCV replication complex [31].
1.2.2.6 Protein NS5A
The NS5A protein is a membrane-located phosphoprotein containing 458 amino acids. A region of NS5A consisting of 40 amino acids is called the interferon sensitivity determining region (ISDR), and mutations in this region are associated with interferon resistance [32, 33]. It plays a role in interferon resistance by inhibiting double-stranded RNA-activated protein kinase R (PKR), induced by interferon, which is an important component of the cellular antiviral and antiproliferative immune response [34]. NS5A also inhibits IRES-dependent replication through PKR inactivation [35]. NS5A is an RNA-binding protein. Positive and negative-stranded HCV RNA has the capacity to bind to its 3′ ends [36]. It also takes part in viral replication, packaging, and release of HCV particles.
1.2.2.7 Protein NS5B
NS5B has RNA-dependent RNA polymerase activity. HCV replication begins with the synthesis of complementary negative-stranded RNA from HCV’s positive-stranded RNA. Then, positive-stranded RNA is synthesized from this negative-stranded RNA. The main enzyme responsible for both of these steps is NS5B RNA-dependent RNA polymerase (RdRp) [3].
1.2.2.8 Protein ARFP/F/Core+1
The HCV F/ARFP/Core+1 protein, where ARFP indicates alternative reading frame protein, F indicates frameshift, and Core+1 indicates its position, suppresses type I and III interferon induction, presumably mediated through the retinoic acid-inducible gene I (RIG-I) signaling pathway, together with other viral factors. It plays a role in the modulation of host immunity [37]. It is thought to play a role in the pathogenesis of hepatocellular carcinoma (HCC) [38].
1.3 HCV replication
The entry of HCV into the cell occurs in a pH and clathrin-dependent manner. HCV primarily binds to glycosaminoglycans (GAG) and low-density lipoprotein receptor (LDL-R). Following its binding to LDL-R and GAGs, it binds to CD81 and scavenger receptor class B type I (SR-BI) via the HVR1 region of the E2 envelope glycoprotein [39, 40]. SR-BI belongs to the CD36 family and is a high-density lipoprotein (HDL) receptor [41]. Following the interaction with HCV, CD81, and SR-BI, it is directed to tight junctions and binds to tight junction proteins claudin-1 and occludin [42, 43, 44]. The virus then undergoes clathrin-dependent endocytosis. The virus, which enters the cell with the endosome, reaches the cytoplasm as a result of the conformational change and fusion in the envelope proteins, as the pH in the endosome decreases. The nonenveloped capsid that enters the cytoplasm opens and viral RNA is released into the cytoplasm. The translation is initiated by IRES on the smooth ER membrane and is cleaved translationally and posttranslationally by the HCV polyprotein, the NS2/3 and NS3/4A proteases of HCV, and the cellular proteases of the host. Local pH changes also convert envelope proteins into their three-dimensional shape. HCV replication is catalyzed by the NS5B RNA-dependent RNA polymerase. Replication ends with the fusion of the virion with the endosomal membrane (Figure 3). After replication of the RNA, the viral particles are packaged, the virion matures and is released from the host cell [45, 46]. In addition to the liver, HCV replicates in peripheral blood mononuclear cells, lymphoid follicles, and bone marrow [45, 46]. HCV has mostly (more than 85%) the same secretion pathways in serum as lipoproteins, and these structures are termed “lipoviral particles” (LVP) [47].
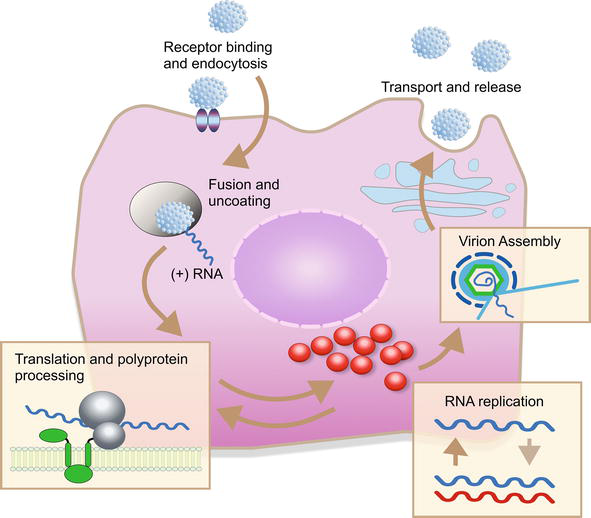
Figure 3.
HCV replication.
1.4 Epidemiology of hepatitis C virus
Approximately 185 million people worldwide are infected with HCV. The global prevalence of infection is known as 2–3% [30, 48]. The most affected regions are Eastern Mediterranean and European regions, with a prevalence of 2.3 and 1.5%, respectively. Egypt has a reported seroprevalence of about 14.7% which is the highest in the world. Substantial regional differences exist in the distribution of HCV genotypes in the world [49]. Every year, 350,000 of these patients die from complications, such as cirrhosis and HCC, associated with chronic hepatitis C (CHC) [50].
1.5 Methods of diagnosis of hepatitis C
Tests used for diagnosis of hepatitis C disease can be divided into serological tests that measure antibodies to hepatitis C and molecular analyzes that measure or detect HCV RNA. Genotype test, biochemical parameters showing fibrosis in serum and liver biopsy are other tests that should be done because they show the response to treatment and prognosis. European Association for the Study of the Liver (EASL) and Centers for Disease Control and Prevention (CDC) currently recommend HCV RNA testing together with the detection of anti-HCV antibodies in the diagnosis of HCV infection [51, 52]. The groups that should be screened for HCV are listed below [53].
Those born between 1945 and 1965
People who use intravenous drugs
Those with HIV infection
Patients with hemophilia who received factor concentrate before 1987
Patients on long-term hemodialysis treatment
Transfusion or transplant recipients
Blood or organ recipients from donors with HCV infection
Children born to mothers with HCV infection
HCV-contaminated needle stick or mucosal exposure
Those who have a sexual partner with HCV infection
Those with extrahepatic manifestations
Those with abnormal liver function tests
The diagnosis of HCV infection is made by detecting anti-HCV antibodies by ELISA and then demonstrating HCV RNA by molecular methods. Quantitative measurement of HCV RNA is used to confirm the diagnosis and the detection limit should be 25 IU/ml or less. In resource-limited locations or where HCV RNA testing is not available, HCV core antigen testing is a viable alternative. Despite the high specificity (>99%) of the anti-HCV test, positivity can be seen in previous acute or chronic hepatitis and false positive results are not uncommon. Especially pregnant women are patient groups that can be false positives in immunological or hematological disease states. At the same time, false negative results can be detected in dialysis patients, transplant patients, HIV-positive patients, and patients with severe immunodeficiency [54, 55]. In such cases, the result should be confirmed by HCV RNA testing. In addition, it should be considered that anti-HCV becomes positive 2–6 months after exposure in patients with acute hepatitis C, and HCV RNA should be requested for diagnosis.
Reactive antibody and negative HCV RNA: A negative HCV RNA test confirms the absence of chronic HCV. False-negative results of RNA are unlikely when sensitive qualitative or quantitative assays that can measure <50 IU/ml are used. As a result, this occurs either after false positive antibody tests for technical reasons or after successfully treating chronic hepatitis C infection. Other rare causes are the transmission of antibodies from a mother with HCV to her baby resulting from antibodies passively passed through blood transfusion.
1.5.1 Detection of anti- HCV antibodies
Antibodies to HCV can be detected by a variety of business models, including standardized laboratory immunoassays, rapid point-of-care immunoassays, and self-administered home testing.
1.5.1.1 Standard immunoassay test
To detect anti-HCV antibodies in serum and plasma, most clinical laboratories use tests that give a positive signal as a result of an enzymatic reaction (EIA or ELISA) or light emission (chemoluminescent assay). These tests provide many advantages, including ease of use, low variability, ease of automation, and relatively low cost. There are several immunoassay tests that target different viral antigens and detect antibodies with varying accuracy. Third-generation EIAs (EIA-3) currently in routine use usually detect antibodies against antigens originating from the core, NS3, NS4, and NS5 proteins. These tests have very high sensitivity and high specificity [56, 57]. dAnti-HCV EIA tests become positive as early as 8 weeks after exposure, and most patients are seroconverted two to 6 months after exposure [58, 59].
1.5.1.2 POCT (point of care test), rapid immunoassay tests
Several rapid tests have been developed for HCV antibodies with performance comparable to standard laboratory-based immunoassays. These tests can be performed on venous blood, fingertip blood, serum, plasma, and oral fluid, and results are usually available in less than 30 minutes. The tests are designed as point-of-care testing to provide more opportunities for HCV testing outside of traditional clinical settings [60].
In the United States, a rapid test (OraQuick HCV Rapid Antibody Test) has been approved by the US Food and Drug Administration (FDA). The test gives results quickly using venous blood, fingertip blood, and saliva. The data suggest that the sensitivity and specificity of the test are equivalent to the EIA test [61].
1.5.1.3 Self-assembled tests
An over-the-counter antibody testing kit (hepatitis C check) has been approved by the FDA. A sample is sent to the lab and results are reported within 4–10 business days.
1.5.1.4 Recombinant immunoblot analysis (RIBA)
Recombinant immunoblot analysis (RIBA) is a test that detects HCV antibodies with similar sensitivity but higher specificity as the second-generation EIA. Where RIBA is available, it can help differentiate between a previous infection (RIBA positive) and a false positive antibody test (RIBA negative) in people with reactive antibody and negative HCV RNA testing.
1.5.2 HCV RNA detection
HCV RNA detection and measurement are essential tools in the diagnosis and management of individuals with chronic HCV infection. HCV RNA measurements are used to confirm the presence or absence of infection and to quantify the amount of HCV RNA present and can be used to guide decisions regarding the duration of treatment with certain regimens. Various methods can be used to detect and quantify HCV RNA and have varying levels of sensitivity. These include polymerase chain reaction (PCR)-based methods, transcription-mediated amplification (TMA), and branched DNA testing.
1.5.3 HCV core antigen test (HCV cAg)
Various immunoassays have been developed to detect the HCV core (HCV cAg) protein, which is a component of the viral particle [62, 63]. In resource-limited populations where nuclear acid testing (NAT) is not available, World Health Organization (WHO) guidelines recommend using an HCV cAg test to confirm viremia [64]. It is a test based on the detection of core antigens of HCV using specific monoclonal antibodies and works with the ELISA or chemoluminescent immune assay (CLIA) method [65]. The lower detection limit varies depending on the HCV genotype but is 500–3000 IU/ml [66, 67]. Although HCV RNA tests have a low detection level of 5–15 IU/mL, approximately 90% of HCV RNA positive samples are above 10,000 IU/mL, which is within the sensitivity range of the HCV core antigen test. Therefore, the next step following the anti-HCV positive screening test may be HCV core antigen detection. In a systematic review of studies evaluating the accuracy of these tests, the most studied analyses (Abbott ARCHITECT HCV Ag test and Ortho HCV Ag ELISA) detected HCV viremia in approximately 93 and 99%, respectively [62].
1.6 Additional examinations
Determining the extent of liver damage by liver biopsy is important for a newly diagnosed chronic HCV patient. It determines the duration of treatment and the treatment regimen. In addition, knowing the additional diseases of the patient is important in terms of predicting the response to treatment, urgency of treatment, and treatment complications. Basal laboratory tests, such as serum aminotransferases, bilirubin, prothrombin time, albumin, complete blood count, glucose, renal functions, urine analysis, 25 hydroxy vitamin D, and pregnancy test, should be requested. In addition, other causes of chronic liver disease, such as autoimmune hepatitis and hemochromatosis, should be investigated in patients with elevated liver function tests. Hepatitis A, B, and HIV serology should be screened and vaccination should be offered if necessary.
Since kidney diseases, such as mixed cryoglobulinemia and membranoproliferative glomerulonephritis, can be seen in HCV, patients should be examined for proteinuria, hematuria, hypertension, and kidney function tests. Additional tests for cryoglobulinemia, complement levels, and rheumatoid factor should be requested, and it should be kept in mind that renal biopsy may be required for diagnosis when significant proteinuria or unrecovered renal functions are observed.
Since HCV genotype is important in determining which treatment regimen will be given for how long, at what dose, and in determining the response to treatment, it is another routine test.
2. Conclusion
In conclusion, since HCV is usually asymptomatic, it is important to know and disseminate diagnostic methods in order to reach more people to be treated. Recently, with the use of DAAs for treating HCV, the success rate has exceeded 90%. The implementation of these strong therapies has reduced the role of monitoring therapy with HCV-RNA tests. Since HCV cAg detects viremia, it can be used especially in populations with limited resources where nuclear acid test is not available.
References
- 1.
Bartenschlager R, Frese M, Pietschmann T. Novel insights into hepatitis C virus replication and persistence. Advances in Virus Research. 2004; 63 :71-180 - 2.
Penin F, Dubuisson J, Rey FA, Moradpour D, Pawlotsky JM. Structural biology of hepatitis C virus. Hepatology. 2004; 39 (1):5-19 - 3.
Moradpour D, Penin F, Rice CM. Replication of hepatitis C virus. Nature Reviews. Microbiology. 2007; 5 :453-463 - 4.
Brass V, Moradpour D, Blum HE. Molecular virology of hepatitis C virus(HCV):2006 update. International Journal of Medical Sciences. 2006; 3 :29-34 - 5.
Yasui K, Wakita T, Tsukiyama-Kohara K, et al. The native form and maturation process of hepatitis C virus core protein. Journal of Virology. 1998; 72 :6048-6055 - 6.
Grakoui A, Wychowski C, Lin C, Feinstone SM, Rice CM. Expression and identification of hepatitis C virus polyprotein cleavage products. Journal of Virology. 1993; 67 :1385-1395 - 7.
Harada S, Watanabe Y, Takeuchi K, et al. Expression of processed core protein of hepatitis C virus in mammalian cells. Journal of Virology. 1991; 65 :3015-3021 - 8.
Santolini E, Migliaccio G, La Monica N. Biosynthesis and biochemical properties of the hepatitis C virus core protein. Journal of Virology. 1994; 68 :3631-3641 - 9.
Chang SC, Yen JH, Kang HY, Jang MH, Chang MF. Nuclear localization signals in the core protein of hepatitis C virus. Biochemical and Biophysical Research Communications. 1994; 205 :1284-1290 - 10.
Suzuki R, Sakamoto S, Tsutsumi T, et al. Molecular determinants for subcellular localization of hepatitis C virus core protein. Journal of Virology. 2005; 79 :1271-1281 - 11.
Schwer B, Ren S, Pietschmann T et al. Targeting of hepatitis C virus core protein to mitochondria through a novel C-terminal localization motif. Journal of Virology. 2004; 78 :7958-7968 - 12.
Asselah T, Rubbia-Brandt L, Marcellin P, Negro F. Steatosis in chronic hepatitis C: Why does it really matter? Gut. 2006; 55 :123-130 - 13.
Chou AH, Tsai HF, Wu YY, et al. Hepatitis C virus core protein modulates TRAIL-mediated apoptosis by enhancing bid cleavage and activation of mitochondria apoptosis signaling pathway. Journal of Immunology. 2005; 174 :2160-2166 - 14.
Akhan S. Hepatit C Virusu. Willke Topçu A, Söyletir G, Doğanay M (Editörler). Enfeksiyon Hastalıkları ve Klinik Mikrobiyolojisi. 3. baskı. İstanbul: Nobel Tıp Kitabevleri; 2008. pp. 1911-1929 - 15.
Kim G-W, Lee S-H, Cho H, et al. Hepatitis C virus Core protein promotes miR-122 destabilization by inhibiting GLD-2. PLoS Pathogens. 2016; 12 (7):e1005714 - 16.
Bartosch B, Dubuisson J, Cosset FL. Infectious hepatitis C virus pseudo-particles containing functional E1–E2 envelope protein complexes. The Journal of Experimental Medicine. 2003; 197 :633-642 - 17.
Cocquerel L, Meunier JC, Pillez A, Wychowski C, Dubuisson J. A retention signal necessary and sufficient for endoplasmic reticulum localization maps to the transmembrane domain of hepatitis C virus glycoprotein E2. Journal of Virology. 1998; 72 :2183-2191.48 - 18.
Cocquerel L, Wychowski C, Minner F, Penin F, Dubuisson J. Charged residues in the transmembrane domains of hepatitis C virus glycoproteins play a major role in the processing, subcellular localization, and assembly of these envelope proteins. Journal of Virology. 2000; 74 :3623-3633 - 19.
Farci P, Shimoda A, Wong D, et al. Prevention of hepatitis C virus infection in chimpanzees by hyperimmune serum against the hypervariable region 1 of the envelope 2 protein. Medical Science. 1996; 93 (26):15394-15399 - 20.
Kato N, Sekiya H, Ootsuyama Y. Humoral immune response to hypervariable region 1 of the putative envelope glycoprotein (gp70) of hepatitis C virus. Journal of Virology. 1993; 67 (7):3923-3930 - 21.
Zvon Hahn T, Yoon JC, Alter H, et al. Hepatitis C virus continuously escapes from neutralizing antibody and T-cell responses during chronic infection In vivo. Gastroenterology. 2007; 132 (2):667-678 - 22.
Carrere-Kremer S, Montpellier-Pala C, Cocquerel L, Wychowski C, Penin F, Dubuisson J. Subcellular localization and topology of the p7 polypeptide of hepatitis C virus. Journal of Virology. 2002; 76 :3720-3730 - 23.
Sakai A, Claire MS, Faulk K, et al. The p7 polypeptide of hepatitis C virus is critical for infectivity and contains functionally important genotype-specific sequences. Proceedings of the National Academy of Sciences USA. 2003; 100 :11646-11651 - 24.
Pavlović D, Neville DCA, Argaud O, et al. The p7 protein of hepatitis C virus forms an ion channel that is blocked by the antiviral drug, amantadine. FEBS Letters. 2003; 535 (10):34-38 - 25.
Welbourn S, Pause A. The hepatitis C virus NS2/3 protease. Current Issues in Molecular Biology. 2007; 9 (1):63-69 - 26.
Boson B, Granio O, Bartenschlager R, Cosset FL. A concerted action of hepatitis C virus p7 and nonstructural protein 2 regulates core localization at the endoplasmic reticulum and virus assembly. PLoS Pathogens. 2011; 7 :e1002144 - 27.
Suzuki R, Matsuda M, Watashi K, et al. Signal peptidase complex subunit 1 participates in the assembly of hepatitis C virus through an interaction with E2 and NS2. PLoS Pathogens. 2013; 9 :e1003589 - 28.
Pawlotsky JM, McHutchison JG. Development of new drugs and clinical trials: Promises and pitfalls. Summary of an AASLD hepatitis single topic conference, Chicago, IL, February 27–march 1, 2003. Hepatology. 2004; 39 :554-567 - 29.
Foy E, Li K, Wang C, et al. Regulation of interferon regulatory factor-3 by the hepatitis C virus serine protease. Science. 2003; 300 :1145-1148 - 30.
Butkiewicz NJ, Yao N, Wright-Minogue J, et al. Hepatitis C NS3 protease: Restoration of NS4A cofactor activity by N-biotinylation of mutated NS4A using synthetic peptides. Biochemical and Biophysical Research Communications. 2000; 267 (1):278-282. DOI: 10.1006/bbrc.1999.1898 - 31.
Lundin M, Monne M, Widell A, Von Heijne G, Persson MA. Topology of the membrane-associated hepatitis C virus protein NS4B. Journal of Virology. 2003; 77 (9):5428-5438 - 32.
Enomoto N, Sakuma I, Asahina Y, et al. Comparison of full-length sequences of interferon sensitive and resistant hepatitis C virus 1b. Sensitivity to interferon is conferred by amino acid substitutions in the NS5A region. Journal of Clinical Investigation. 1995; 96 :224-230 - 33.
Enomoto N, Sakuma I, Asahina Y, et al. Mutations in the nonstructural protein 5A gene and response to interferon in patients with chronic hepatitis C virus 1b infection. The New England Journal of Medicine. 1996; 334 :77-81 - 34.
Gale MJ Jr, Korth MJ, Tang NM, et al. Evidence that hepatitis C virus resistance to interferon is mediated through repression of the PKR protein kinase by the nonstructural 5A protein. Virology. 1997; 230 :217-227 - 35.
Karamichali E, Foka P, Tsitoura E, et al. HCV NS5A co-operates with PKR in modulating HCV IRES- dependent translation. Infection, Genetics and Evolution. 2014; 26C :113-122 - 36.
Huang L, Hwang J, Sharma SD, et al. Hepatitis C virus nonstructural protein 5A (NS5A) is an RNA-binding protein. The Journal of Biological Chemistry. 2005; 280 :36417-36428 - 37.
Park SB, Seronello S, Mayer W, Ojcius DM. Hepatitis C virus frameshift/alternate Reading frame protein suppresses interferon responses mediated by pattern recognition receptor retinoic-acid-inducible gene-I. PLoS One. 2016; 11 (7):e0158419 - 38.
Dalagiorgou G, Vassilaki N, Foka P, et al. High levels of HCV core+1 antibodies in HCV patients with hepatocellular carcinoma. The Journal of General Virology. 2011; 92 (6):1343-1351 - 39.
Molina S, Castet V, Pichard-Garcia L, et al. Serum-derived hepatitis C virus infection of primary human hepatocytes is tetraspanin CD81 dependent. Journal of Virology. 2008; 82 :569-574 - 40.
Scarselli E, Ansuini H, Cerino R, et al. The human scavenger receptor class B type I is a novel candidate receptor for the hepatitis C virus. The EMBO Journal. 2002; 21 :5017-5025 - 41.
Krieger M. Scavenger receptor class B type I is a multiligand HDL receptor that influences diverse physiologic systems. The Journal of Clinical Investigation. 2001; 108 (6):793-797 - 42.
Evans MJ, von Hahn T, Tscherne DM, et al. Claudin-1 is a hepatitis C virus co-receptor required for a late step in entry. Nature. 2007; 446 :801-805 - 43.
Liu S, Yang W, Shen L, et al. Tight junction proteins claudin-1 and occludin control hepatitis C virus entry and are downregulated during infection to prevent superinfection. Journal of Virology. 2009; 83 :2011-2014 - 44.
Ploss A, Evans MJ, Gaysinskaya VA, et al. Human occludin is a hepatitis C virus entry factor required for infection of mouse cells. Nature. 2009; 457 :882-886 - 45.
Petersen SV, Thiel S, Jensenius JC. The manan-binding lectin pathway of complement activation: Biology and disease association. Mol. Immun. 2001; 38 :133-149 - 46.
Eisen DP, Minchinton RB. Impact of mannose-binding on susceptibility to infectious diseases. CID. 2003; 37 :1496-1505 - 47.
André P, Komurian-Pradel F, Deforges S, et al. Characterization of low- and very-low-density hepatitis C virus RNA-containing particles. Journal of Virology. 2002; 76 :6919-6928 - 48.
Lavanchy D. The global burden of hepatitis C. Liver International. 2009; 1 :74-81 - 49.
Mohamoud YA, Mumtaz GR, Riome S, Miller D, Abu-Raddad LJ. The epidemiology of hepatitis C virus in Egypt: A systematic review and data synthesis. BMC Infectious Diseases. 2013; 13 :288 - 50.
Aygen B, Demirtürk N, Türker N, et al. Management of chronic hepatitis C virüs infection: A consensus report of the study group for viral hepatitis of the Turkish Society of Clinical Microbiology and Infectious Diseases-2017. Update. 2017; 30 :2-36 - 51.
European Association for the Study of the Liver. EASL clinical practice guidelines: Management of hepatitis C virus infection. Journal of Hepatology. 2011; 55 :245-264 - 52.
Centers for Disease Control and Prevention (CDC). Testing for HCV infection: An update of guidance for clinicians and laboratorians. MMWR. Morbidity and Mortality Weekly Report. 2013; 62 :362-365 - 53.
Up to date- HCV. Available from: https://www.uptodate.com/contents/diagnosis-and-evaluation-of-chronic-hepatitis-c-virusinfection?search=hepatitis%20c%20whom%20to%20test&source=search_result&selectedTitle=2~150&usage_type=default&display_rank=2 . [Accessed: December 13, 2022] - 54.
Pereira BJ, Levey AS. Hepatitis C virus infection in dialysis and renal transplantation. Kidney International. 1997; 51 :981 - 55.
Lau JY, Davis GL, Brunson ME, et al. Hepatitis C virus infection in kidney transplant recipients. Hepatology. 1993; 18 :1027 - 56.
Gretch DR. Diagnostic tests for hepatitis C. Hepatology. 1997; 3 (1):43S-47S - 57.
Uyttendaele S, Claeys H, Mertens W, et al. Evaluation of third-generation screening and confirmatory assays for HCV antibodies. Vox Sanguinis. 1994; 66 :122 - 58.
Maheshwari A, Thuluvath PJ. Management of acute hepatitis C. Clinics in Liver Disease. 2010; 14 :169 - 59.
Kamili S, Drobeniuc J, Araujo AC, Hayden TM. Laboratory diagnostics for hepatitis C virus infection. Clinical Infectious Diseases. 2012; 55 (1):43 - 60.
Stockman LJ, Guilfoye SM, Benoit AL, et al. Rapid hepatitis C testing among persons at increased risk for infection--Wisconsin, 2012-2013. MMWR. Morbidity and Mortality Weekly Report. 2014; 63 :309 - 61.
Lee SR, Yearwood GD, Guillon GB, et al. Evaluation of a rapid, point-of-care test device for the diagnosis of hepatitis C infection. Journal of Clinical Virology. 2010; 48 :15 - 62.
Freiman JM, Tran TM, Schumacher SG, et al. Hepatitis C Core antigen testing for diagnosis of hepatitis C virus infection: A systematic review and meta-analysis. Annals of Internal Medicine. 2016; 165 :345 - 63.
Khan H, Hill A, Main J, et al. Can hepatitis C virus antigen testing replace ribonucleic acid polymerase chain reaction analysis for detecting hepatitis C virus? A systematic review. Open forum. Infectious Diseases. 2017; 4 (2):ofw252 - 64.
World Health Organization. WHO Guidelines on Hepatitis B and C Testing. 2017. Available from: http://apps.who.int/iris/bitstream/10665/254621/1/9789241549981-eng.pdf?ua=1. [Accessed: December 13, 2022] - 65.
Lorenzo J, Castro A, Aguilera A, et al. Total HCV core antigen assay: A new marker of HCV viremia and its application during treatment of chronic hepatitis C. Journal of Virological Methods. 2004; 120 (2):173-177 - 66.
Chevaliez S, Feld J, Cheng K, et al. Clinical utility of HCV core antigen detection and quantification in the diagnosis and management of patients with chronic hepatitis C receiving an all-oral, interferon-free regimen. Antiviral Therapy. 2018; 23 (3):211-217 - 67.
Heidrich B, Pischke S, Helfritz FA, et al. Hepatitis C virus core antigen testing in liver and kidney transplant recipients. Journal of Viral Hepatitis. 2014; 21 :769-779