Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
This chapter is devoted to elucidating the bioecological characteristics of the existence of five invasive leafminer moth species of the family Gracillariidae Steinton, 1854 (Parectopa robiniella (Clemens, 1863), Phyllonorycter issikii (Kumata, 1963), Phyllonorycter platani (Staudinger, 1870), Macrosaccus robiniella (Clemens, 1859) and Cameraria ohridella Deschka & Dimic, 1986) and extent of their impact on dendroflora in Ukraine. Features of their invasion into various ecosystems were revealed. The effects of Gracillariidae caterpillar feeding on host (food) plants were studied. The caterpillars of invasive leafminer moth species affected the functional state of host plants; it was confirmed by alterations in the content of soluble proteins, activity and isoenzyme composition of benzidine peroxidase during the growing season of the host plants. The study of dynamics of highly soluble proteins showed a decrease in their content at substantial leaf damage. The restructuring of both peroxidase activity and its isoenzyme profile was revealed to be indicators of the plant biochemical adaptation to the feeding of caterpillars of invasive leaf blotch miners. The results obtained show the activation of the enzymatic antioxidant protection system in host plants in response to the damaging effect of Gracillariidae caterpillars, which ensures the survival of the plant and completes its ontogenesis program in unfavorable conditions.
Keywords
invasive Gracillariidae
leafminer moths in the fauna of Ukraine
adaptation of invasive leafminers to a novel environment
effect of leafminers on the physiological state of host plant
biochemical adaptation of trees to caterpillar feeding
Research Institute of Biology, Oles Honchar Dnipro National University, Dnipro, Ukraine
Oksana Seliutina
Research Institute of Biology, Oles Honchar Dnipro National University, Dnipro, Ukraine
Larysa Shupranova
Research Institute of Biology, Oles Honchar Dnipro National University, Dnipro, Ukraine
Iryna Loza
Research Institute of Biology, Oles Honchar Dnipro National University, Dnipro, Ukraine
Svitlana Sytnyk
Research Laboratory of Forestry, Dnipro State Agrarian and Economic University, Dnipro, Ukraine
Viktoria Lovynska
Research Laboratory of Forestry, Dnipro State Agrarian and Economic University, Dnipro, Ukraine
Olexander Pakhomov
Faculty of Biology and Ecology, Department of Zoology and Ecology, Oles Honchar Dnipro National University, Dnipro, Ukraine
*Address all correspondence to: holoborodko.kk@gmail.com
1. Introduction
This chapter is devoted to elucidating the bioecological characteristics of the existence of invasive leafminer species of the family Gracillariidae (Lepidoptera) and the degree of their influence on dendroflora in Ukraine. The biological characteristics of invasive lepidopteran species were clarified. The scale and direction of invasion of five Gracillariidae species were determined in the territory of Ukraine. Features of settlement of various ecosystems were revealed.
This chapter found the effect of Gracillariidae caterpillar feeding on host (food) plants. The caterpillars of invasive insect species affect the functional state of host plants; it was confirmed by alterations of soluble protein content, activity and isoenzyme composition of benzidine peroxidase during the growing season of these plants. The study on the dynamics of highly soluble proteins showed their decrease in the case of high-level leaf damage.
The restructuring of both the activity and the isoenzyme profile of peroxidase turned out to be an indicator of plant biochemical adaptation to the feeding of caterpillars of invasive leaf blotch miners. Statistically significant differences were found for soluble peroxidase; its activity increased by an average of 2.1 times in the case of high-level damage of host plant leaves by the caterpillars. High-level leaf damage by the phytophagous insect was reflected in the alteration of the isoenzyme profile of benzidine peroxidase. The main pattern of negative effects of Gracillariidae caterpillars involves a significant increase in the activity of the most acidic molecular forms of cytoplasmic peroxidase in the leaves of host plants.
The results obtained demonstrated the activation of the enzymatic antioxidant defense system in host plants in response to the damaging effect of Gracillariidae caterpillars, which ensures the survival of the plant and completion successfully its ontogenesis program under the effect of unfavorable environmental conditions. The most significant factor in protecting the cells against caterpillar activity involves an increase in guaiacol peroxidase activity which indicates an enhancing of cell barrier properties.
It was established that the caterpillars of Gracillariidae invasive species can have a significant effect on photosynthetic apparatus of host plants. Such effect was clearly determined with the fluorescence technique which was carried out with a portable fluorometer, “Floratest.” Feeding of Cameraria ohridella caterpillars causes a decrease in the activity of photosynthetic apparatus in Aesculus hippocastanum regardless of the spatial arrangement of leaves in the tree crown. Under the influence of C. ohridella feeding on A. hippocastanum leaves, the amount of chlorophyll not involved in photosynthetic energy transfer in reaction centers increased. Conversely, during the development of only a single C. ohridella generation, a constant decrease in the quantum efficiency of PS II (inhibition of photosynthetic activity) was observed.
Already at the beginning of the moth mine development (first-aged C. ohridella caterpillars), a significant decrease in activity and an increase in the plateau coefficient were observed in the leaves in both illuminated and shaded part of the tree crown; it, in turn, indicates an inhibition of photosynthetic photophysical and photochemical processes and a reduction of electron acceptor pool in the electron transport chain. Such pathological alterations were caused by a decrease in the content of active chlorophyll (a component of PS II pigment-protein complexes) and its destruction. The values of the key parameters of chlorophyll fluorescence induction indicate a significant inhibition of photosynthesis processes and a violation of the coherence of reactions in the Calvin cycle. Being determined by local changes in chloroplast structure and functions, these metabolic transformations cause the damage of photosynthetic apparatus in plants resulted in a decrease in the content of photosynthetic pigments.
As a result of our research, no relationship was detected between the parameters of urbanocenoses in which the development of mines of invasive insects took place and the number of larvae infected by parasitoids. Only the relationship between the number of mines formed by invasive insect insects and the degree of their damage by parasitoids (Hymenoptera) has been established. The relative resistance of invasive Gracillariidae species to diseases in a novel environment was elucidated. The fact of damage to the preimaginal developmental stages by the entomopathogen was established only for Macrosaccus robiniella. The fact of infection of invasive caterpillars with the fungus Lecanicillium sp. (Verticillium) has been established for the first time in the territory of Ukraine.
2. Bioecological characteristics of Gracillariinae invasive species in the territory of Ukraine
Taxonomically, the leaf blotch miners (Gracillariidae Steinton, 1854) form a separate family. As a result of a molecular phylogenetic study performed in 2017 [1], eight following subfamilies were allocated: Acrocercopinae Kawahara & Ohshima, 2016; Gracillariinae Stainton, 1854; Lithocolletinae Stainton, 1854; Marmarinae Kawahara & Ohshima, 2016; Oecophyllembiinae Réal & Balachowsky, 1966; Ornixolinae Kuznetzov & Baryshnikova, 2001; Parornichinae Kawahara & Ohshima, 2016; Phyllocnistinae Herrich-Schäffer, 1857.
Ninety-eight genera and 1900 species of world fauna have been described. The Palearctic region possesses 42 genera with more than 500 species. In Ukraine, 94 species belonging to 18 families were described. Among Gracillariidae faunal complex, eight species have invasive status in Europe; among them, five following species were registered within Ukraine: Parectopa robiniella (Clemens, 1863), Phyllonorycter issikii (Kumata, 1963), Phyllonorycter platani (Staudinger, 1870), Macrosaccus robiniella (Clemens, 1859) and Cameraria ohridella Deschka & Dimic, 1986.
Almost all recent Gracillariidae species act as obligate leafminers throughout the development of all caterpillar ages. The vast majority of species are narrow oligophages or monophages, rarely polyphages, trophically associated with dicotyledons and partially with gymnosperms. Food chains of most Palearctic species were established with Fagaceae, Rosaceae, Fabaceae, Salicaceae, Aceraceae and others. They can damage fruit, ornamental and industrial plants, mainly introduced dendroflora. With mass reproduction, large-scale outbreaks of abundance can be observed. This chapter provides an assessment of the scale of modern invasion in the territory of Ukraine. The analysis of the experience of studying the bioecological features of invasive insect species in the novel range was carried out.
The studies have been conducted for 15 years (2006–2022) within the steppe zone of Ukraine in 10 stationary sample sites (the Dnipropetrovsk region: Dnipro city, Nikopol; the Dnipro-Oril Nature Reserve; Zaporizhia region: National Nature Park “Velykyi Luh”; the Mykolaiv region: Tylihulskiy Regional Landscape Park), reflecting the conditions of various types of artificial plantings of host plants that act as foodplants for invasive Gracillariidae. The ecosystems under study differed in geographical location, type and degree of anthropic load.
As an object of research, in the green areas of populated cities, various groups of model different-aged horse chestnut and black locust trees were identified with similar morphological and taxational features but with different degrees of leaf damage by invasive insect species. The degree of damage of leaf blades in horse chestnut by C. ohridella was evaluated visually using a modified scale proposed by Shupranova et al. [2].
All invasive species form mines on the leaves of host plants; such mines were recorded by visual examinations. Counting mines on the trees was carried out with the conventional “model branch” method [3]. A tree and a random branch with a length of at least 1 m were selected by randomization. After that, the number of leaves and mines formed by the invasive leafminer was counted. The photos of leaves damaged by miners were taken using a digital camera with a resolution of 5 megapixels. An object micrometer was applied to the leaves to calibrate the measured parameters. The measurements were made on digital photos using ToupView 3.7 software. The mine length (L) was measured along the central leaf vein; the maximum mine width (W) was measured perpendicularly to its length; also, the mine area (S) and the leaf blade area (Sl) were measured [3]. The mine length-to-mine width ratio (L/W) and the mine area-to-leaf blade area ratio (S/Sl) were calculated. Data processing was performed using Statistica 12.5 software package.
3.2 Analytical techniques for heavy metal content in plant tissues
The leaf blades were dehydrated in porcelain tiles using a drying cabinet at t° = 100°C. After being dehydrated, the dry leaves were weighed on electronic scales AXIS AD500 with an accuracy of 0.001 g (the weigh required for analysis was 0.5–1.0 g). Then, the weights were incinerated in a muffle furnace at t° = 450°C. The ash was poured into a conical flask, and 0.5 mL of concentrated nitric acid and 0.5 mL of double-distilled water were added. The resulting solution was adjusted to a volume of 10 mL with distillate water and filtered through an ash-free filter paper, then the tiles were washed with 10 mL of double-distilled water and the solution volume was adjusted to 25 mL. The content of Zn, Cu, Pb and Cd was analyzed in the samples. The heavy metal content in leaf blades was analyzed with atomic absorption spectrophotometry using an AAS-30 spectrophotometer according to the standard method proposed by Khavezov, 1983.
3.3 Morphometric methods
Photographs of the insects were taken through binocular MBS-10 equipped with 5 MP digital camera. To calibrate the measured parameters of insects, an object micrometer was applied. The measurements were made on digital photos using ToupView software. Primary data processing was performed in MS Excel 2019, and subsequent data processing was performed using Statistica 13 software package. Excess (Ex) and asymmetry (As) indicators were used to compare the detection of the morphometric characteristic deviations from the normal distribution. Coefficient of variation (CV) and standard deviation (SD) were used to analyze intrapopulation diversity. Univariate analysis of variance (ANOVA) was used to compare linear characteristics and indices to detect interpopulation polymorphism.
3.4 Methods for determining the effect of feeding of Gracillariidae caterpillars on biochemical processes in host plant tissues
To carry out biochemical analysis, the leaves were washed in water and immediately used for enzyme extraction. To isolate the enzyme preparation, tree leaves (0.3 g) were homogenized in 6 mL of 0.05 M tris-HCl buffer, pH 7.4, with 0.5% polyvinylpyrrolidone (PVP). The samples were extracted at +4°C for 1 hour and centrifuged for 15 min at 14,000 rpm. The supernatant was used to determine the activity and isoenzyme composition of benzidine peroxidase (BPOD) and guaiacol peroxidase (GPOD) activity. BPOD activity (BPOD, EC 1.11.1.7) was measured at 490 nm in a reaction mixture (0.8 mL of Na-acetic buffer, pH 5.4; 1 mL of benzidine solution and 0.2 mL of enzyme preparation) after adding 1% Н2О2. The activity was calculated within 1-min interval when the reaction rate reached a maximum [4]. The result was expressed in absorbance units/g of raw material min.
Izoenzyme composition of BPOD was determined by isoelectric focusing (IEF) in a 5% horizontal polyacrylamide gel (PAAG) using Ultraphor (LKB, Bromma, Sweden), pH range 3.5–6.5. A benzidine technique was used to detect enzymatic activity in PAAG. Blue-colored bands appeared, which turned brown after 10–15 min. These brown-colored bands represented the BPOD activity. The gels were scanned and analyzed using the 1D Phoretix software intended for determination of the relative content (%) of each isoform in the total peroxidase spectrum.
Guaiacol-depended peroxidase (GPOD, EC 1.11.1.7) activity was evaluated according to Ranieri et al. [5, 6] by determining guaiacol oxide at 470 nm in a reaction mixture containing an acetic buffer (pH 6.0), a 2 mM guaiacol solution, 0.2 mL of enzyme preparation and 0.15% Н2О2. The results were calculated considering the molar extinction coefficient (26.6 mM−1 cm−1) and expressed in mM guaiacol/g*raw material.
The protein content in the samples was determined by the Bradford [7] method with Coomassie brilliant blue G 250 dye relative to the bovine serum albumin standard (Serva, USA).
The results of studies on enzyme activity were presented as mean x ± SD (standard deviation). Data obtained were analyzed using Statistica (version 8, StatSoft, USA). Tukey’s Honestly Significant Difference test was used to determine the significant difference between the group means. The differences were found to be statistically significant at p < 0.05.
3.5 Methods for determining the effect of feeding of Gracillariidae caterpillar on the state of the photosynthetic apparatus in host plants
Diagnostics of photosynthetic disorders of native chlorophyll in fresh leaves were performed using a portable fluorometer “Floratest” developed by V. M. Glushkov Institute of Cybernetics of the National Academy of Sciences of Ukraine. The portable fluorometer “Floratest” comprises a base unit with a graphic liquid crystal display and remote optoelectronic sensor, connecting cable to the USB port of a personal computer, and a network adapter. LED, as a component of the remote optoelectronic sensor, has a maximum radiation intensity at λ = (470 + 20) nm. Irradiation parameters in the sensor were the following: irradiation wavelength 470 + 15 nm; irradiated spot area not less than 15 mm2; light intensity within the spot at least 2.4 W/m2. Signal reception indicators in an optoelectronic sensor: the spectral range of fluorescence intensity measurement was 670–800 nm; receiving window area 9 mm2; photodetector sensitivity at λ = 650 nm was 0.45 A/W.
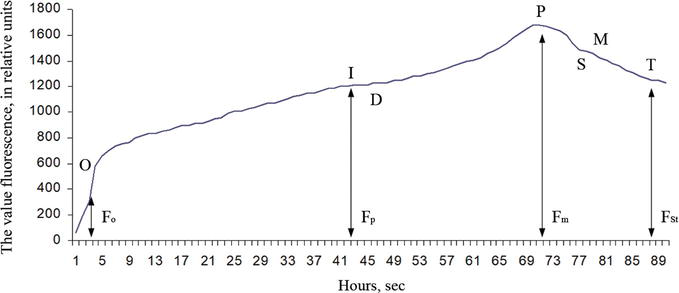
The observations were made on fresh A. hippocastanum leaves both damaged and undamaged by C. ohridella. After starting the light exposure, chlorophyll fluorescence intensity (induction of fluorescence or light-induced [caused] fluorescence) begins to change significantly over time. The time dependence of the chlorophyll fluorescence intensity (ChlF) has the characteristic form of a curve with one or more maximums, and it is called the ChlF induction curve, or the Kautsky curve (Figure 1).
Figure 1.
Typical chlorophyll fluorescence induction curve: F0 is an initial value of fluorescence induction after irradiation is turned on; Fp is a value of “plateau” fluorescence induction; Fm is a maximum value of fluorescence induction; FSt is a stationary value of fluorescence induction after light adaptation of a plant leaf.
The shape of this curve is quite sensitive to changes that occurred in the photosynthetic apparatus of plants during their adaptation to different environmental conditions, which has become the basis for the widespread use of the Kautsky effect in photosynthesis studies.
4. Mechanisms of invasion of different types of ecosystems by Gracillariidae species
4.1 Characteristics of invasion of field-protective forest belts
The invasion of field-protective forest belts occurs unevenly; we found the tree age to be the main factor in this process. We studied the characteristics of invasion of the black locust (Robinia pseudoacacia Linnaeus, 1753) plantings by the insect species because these trees occupy the most significant area among artificial field-protective plantings in Ukraine. It was found that R. pseudoacacia manifests itself relatively actively in almost all natural zones of Ukraine, mainly due to its spread by root growth. The forest belt surveys allowed us to distinguish three age-related variants of R. pseudoacacia trees (parent trees planted at the time of a forest belt creation (50–70 years), sprouting trees (15–25 years), and young trees (up to 15 years)). Pаrectopа robiniella (Clemens, 1863) and Macrosaccus robiniella (Clemens, 1859) were detected on black locust in all geographical areas of the country. Monitoring surveys have shown varying degrees of invasion of three age groups of trees by Gracillariidae. It was discovered that both invasive insect species mainly develop mines on the leaves of young trees (up to 15 years old).
By the results of calculating the average number of mines on a simple leaf, it turned out that the invasive insect species most often formed the mine on the lower (near the petiole) leaf segment in all three age groups of the trees. Determining the characteristics of the relative invasion of simple leaves related to the total number of damaged leaves showed that the invasion of leaf blades was the highest within the location of the middle simple leaves on the complex leaf of the black locust.
Such invasion patterns may be explained by the different growth rates of R. pseudoacacia leaf blades and the qualitative characteristics of leaf tissues. Thus, our survey confirmed the assumption that the topical specificity of invasion is derived from the peculiarities of divergent leaf development and acropetal development of the leaf of the host plant.
4.2 Features of invasion of urban green areas by invasive insect species
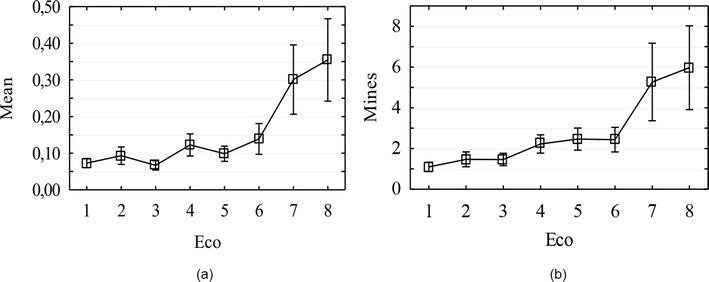
The study was conducted within Dnipro city, in seven largest green areas, and in Dnipro-Oril Nature Reserve. According to the results of statistical data processing, the largest number of mines was registered in park areas of Dnipro city (Figure 2), located at high terrain levels (higher than 100 m). The variability curve of the average number of mines on the leaf (a) is almost completely same to the variability curve of the absolute number of mines (b). Accordingly, the average number of mines does not depend on the number of leaves on the host plant. Based on the results of univariate variance analysis of the settlement of different green areas in Dnipro city by invasive leafminers (Table 1), statistically significant (p < 0.05) differences were found.
Figure 2.
Variability of settlement of the model branches of R. pseudoacacia L. by Parectopa robiniella: A—Variability of the average number of mines on leaves (Meаn), b—Variability of the absolute number of mines on leaves (mines); the x-axis: An ecosystem number (eco), the y-axis: A value of the characteristic.
Eco*
x ± SD
SS
MS
F
P
1
1.10 ± 0.32
1.0360
1.0360
1.2608
0.2682
2
1.47 ± 1.02
1
1.10 ± 0.32
1.0403
1.0403
1.4135
0.2404
3
1.46 ± 0.94
1
1.10 ± 0.32
9.8560
9.8560
6.8703
0.0120*
4
2.22 ± 1.33
1
1.10 ± 0.32
15.7926
15.7926
4.4560
0.0387*
5
2.46 ± 2.03
1
1.10 ± 0.32
14.2041
14.2041
5.0388
0.0295*
6
2.44 ± 1.86
1
1.10 ± 0.32
130.2083
130.2083
6.5556
0.0146*
7
5.27 ± 5.10
Table 1.
One-way analysis of variance of invasive species settlement in different green areas of Dnipro city (n = 242).
* Urbocenoses: 1—Manuylovsky Park; 2—Lazaria Hloby Park; 3—Druzhby Narodiv Forest Park; 4—Pridneprovsky Park; 5—Park 40th anniversary of the liberation of Dnipropetrovsk; 6—T. G. Shevchenko Park; 7—Oles Honchar Dnipro National University (DNU) Botanical Garden.
Morphological characteristics of mines can be considered the main indicators of successful feeding of leafminer caterpillars [3]. Assessment of the mine area determines the feeding activity of caterpillars within this mine [8]. To test these hypotheses, we conducted morphometric studies of mines formed by Parectopa robiniella and Macrosaccus robiniella. As a result of the research conducted, it turned out that both insect species studied show relatively good adaptability to the choice of suitable environmental conditions. This was evidenced by the statistical data obtained by us in morphological studies of mines formed by both insect species (Table 2).
Characteristics
Parectopa robiniella
Macrosaccus robiniella
CV
SD
X
CV
SD
x
L
0.40
5.50
13.70
0.30
4.20
15.80
W
0.60
4.90
8.40
0.20
1.30
7.40
S
0.80
3178.10
3787.60
0.30
1131.90
3628.20
Sl
0.40
11038.60
27137.70
0.30
10281.20
31877.40
L/W
0.30
0.50
1.90
0.10
0.30
2.10
S/Sl
0.80
0.10
0.10
0.40
0.01
0.10
Table 2.
Morphometric variability of mines in populations of Parectopa robiniella (n = 184) and Macrosaccus robiniella (n = 180) on the territory of DNU botanical garden.
According to the results of the study, both coefficient of variation and standard deviation indicated that the morphometric plasticity of mines formed by Parectopa robiniella showed a more variable polymorphism than of mines formed by Macrosaccus robiniella. Average values of length of mines formed by Parectopa robiniella were less than of mines formed by Macrosaccus robiniella. At the same time, average width (W) and area (S) of mines formed by Parectopa robiniella were larger, and the leaf surface area (Sl) of leaves with mines formed by Parectopa robiniella was less than the surface area of leaves with mines formed by Macrosaccus robiniella. The mine length-to-mine width ratio (L/W) was greater in the mines formed by Macrosaccus robiniella, which indicates a higher elongation of the mines formed by this insect.
4.3 Effect of heavy metals on distribution of invasive insects in urban agglomerations
The content of pollutants of various origins in the atmospheric air is considered to be one of the leading factors of pollution in urban environments. Woody vegetation in urboecosystems is constantly exposed to such pollution, which is reflected, among other, by an increase in heavy metal content in leaf tissues. Being primary consumers, phytophages are affected by the content of heavy metals, and their certain concentrations can even influence the vital processes of larval stages of insect development.
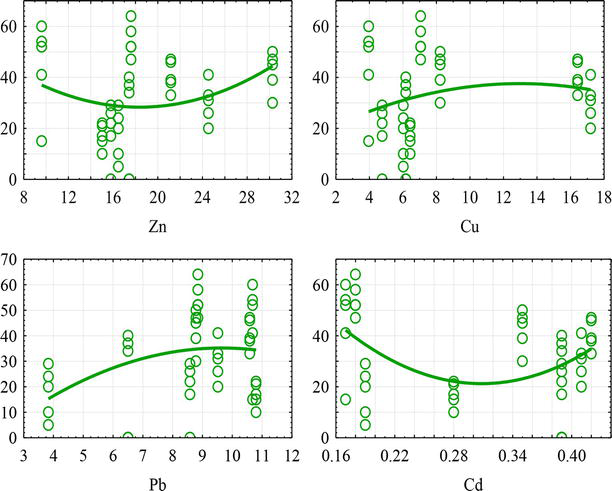
Research of the effects of heavy metals (Zn, Cu, Pb, Cd) on urban populations of Parectopa robiniella showed significant differences in the types of urboecosystems (Figure 3). It turned out that the higher the heavy metal content (especially zinc, copper and lead) in the leaf tissues of a host plant, the greater the amount of insect mines. Results of a general linear model of the effect of heavy metal content and terrain height on the number of mines formed by Parectopa robiniella in the conditions of Dnipro city (Table 3) also demonstrated this regularity.
Figure 3.
Dependence of the number of mines formed by Parectopa robiniella (y-axis) on heavy metal content (x-axis, in mcg/g).
Effect
Sum of squares
Degrees of freedom
Mean sum of squares
F-ratio
p-level
Beta regression coefficients
Constant
2170.4
1
2170.4
14.75
<0.001
—
H
537.7
1
537.7
3.65
0.06
0.25 ± 0.13
Year
2170.7
1
2170.7
14.75
<0.001
0.42 ± 0.11
Zn
1288.0
1
1288.0
8.75
0.01
0.43 ± 0.15
Cu
0.2
1
0.2
0.00
0.97
−0.01 ± 0.17
Pb
1146.0
1
1146.0
7.79
0.01
0.34 ± 0.12
Cd
1606.1
1
1606.1
10.92
<0.001
−0.50 ± 0.15
Error
5591.4
38
147.1
—
—
—
Table 3.
Results of a general linear model of the effect of heavy metal content and terrain height on the number of mines formed by Parectopa robiniella (R2adj = 0.48, F = 7.8, p < 0.001).
5. Adaptive features of invasive Gracillariidae species to the novel environmental conditions
5.1 Characteristics of morphometric variability within the population group of single urbocenosis
According to our study, the distribution of Macrosaccus robiniella pupae collected in Pridneprovsky Park (Table 4) shows among linear characteristics a significant asymmetry in height of body (Hb, As = 1.33) and length of elytra (Le, As = 0.76), as well as among indices the length of body-to-height of body ratio (Lb/Hb, As = −1.09), and the length of elytra-to-height of body ratio (Le/Hb, As = −0.97). The excess values were reliable for height of body (Hb, Ex = 0.82) and the length of body-to-length of elytra ratio (Lb/Le, Ex = −1.11). Length of body (Lb) and length of elytra (Le) showed no significant asymmetry or excess. A significant coefficient of variation (CV) was also observed in height of body (Hb), the length of body-to-height of body ratio (Lb/Hb) and the length of elytra-to-height of body ratio (Le/Hb).
Urbo
n
Characteristics
x ± SD
CV
As
Ex
Pridneprovsky Park
16
Lb, mm
3.48 ± 0.17
0.05
−0.20
0.12
Hb, mm
0.79 ± 0.16
0.20
1.33
0.82
Le, mm
2.12 ± 0.12
0.06
0.76
0.31
Lb/Le
1.64 ± 0.11
0.07
0.18
−1.11
Lb/Hb
4.55 ± 0.86
0.19
−1.09
0.05
Le/Hb
2.76 ± 0.45
0.16
−0.97
0.02
Oles Honchar Dnipro National University Botanical Garden
34
Lb, mm
3.34 ± 0.28
0.08
−0.31
−1.15
Hb, mm
0.82 ± 0.08
0.10
0.66
0.52
Le, mm
1.88 ± 0.20
0.10
0.01
−1.03
Lb/Le
1.79 ± 0.17
0.09
0.18
−0.43
Lb/Hb
4.11 ± 0.42
0.10
0.08
−0.46
Le/Hb
2.30 ± 0.20
0.09
−0.73
0.74
Druzhby Narodiv Forest Park
18
Lb, mm
3.30 ± 0.19
0.06
−0.91
−0.32
Hb, mm
0.81 ± 0.09
0.11
0.79
0.73
Le, mm
1.73 ± 0.15
0.09
0.25
−0.17
Lb/Le
1.92 ± 0.13
0.07
−1.49
1.35
Lb/Hb
4.10 ± 0.37
0.09
−1.37
1.75
Le/Hb
2.14 ± 0.18
0.08
0.65
−1.08
Park 40th anniversary of the liberation of Dnipropetrovsk
22
Lb, mm
3.46 ± 0.16
0.05
−0.23
−0.80
Hb, mm
0.76 ± 0.09
0.12
0.28
−0.17
Le, mm
1.96 ± 0.12
0.06
0.62
0.07
Lb/Le
1.76 ± 0.08
0.05
0.27
−0.76
Lb/Hb
4.60 ± 0.51
0.11
0.38
−1.14
Le/Hb
2.61 ± 0.28
0.11
0.47
−0.58
T. G. Shevchenko Park
22
Lb, mm
3.44 ± 0.20
0.06
−1.17
0.37
Hb, mm
0.78 ± 0.05
0.07
0.65
−0.37
Le, mm
1.87 ± 0.15
0.08
−0.57
−0.83
Lb/Le
1.85 ± 0.10
0.05
0.53
0.45
Lb/Hb
4.40 ± 0.33
0.07
0.51
−0.22
Le/Hb
2.38 ± 0.17
0.07
0.11
−1.21
The Dnipro-Oril Nature Reserve
28
Lb, mm
2.79 ± 0.09
0.03
0.37
−0.95
Hb, mm
0.67 ± 0.06
0.09
0.03
−0.88
Le, mm
1.68 ± 0.16
0.10
2.19
4.82
Lb/Le
1.67 ± 0.10
0.06
−2.32
5.39
Lb/Hb
4.23 ± 0.36
0.08
0.40
−0.44
Le/Hb
2.54 ± 0.27
0.11
0.51
−0.80
Table 4.
Morphometric variability of Macrosaccus robiniella within population groups (n = 140).
Note: Urbo—name of urbocenosis; n—number of pupae measured; characteristics—parameters and indices studied; x ± SD—mean ± standard deviation; CV—coefficient of variation; As—asymmetry coefficient; Ex—excess coefficient.
Among the pupae collected in Oles Honchar Dnipro National University Botanical Garden, a reliably significant asymmetry was observed only in height of body (Hb, As = 0.66), in the length of elytra-to-height of body ratio (Le/Hb, As = −0.73), while a significant excess was observed in the length of body (Lb, Ex = −1.15), the length of elytra (Le, Ex = −1.03), and the length of elytra-to-height of body ratio (Le/Hb, Ex = 0.74). The body-length-to-body-height ratio (Lb/Hb) and the body-length-to-wing-length ratio (Lb/Le) showed no significant asymmetry or excess. None of the characteristics studied showed a significant CV.
A significant asymmetry was observed among the pupae sampled in Druzhby Narodiv Forest Park: length of body (Lb, As = −0.91), height of body (Hb, As = 0.79), the length of body-to-elytra ratio (Lb/Le, As = −1.49), the length of body-to-height of body ratio (Lb/Hb, As = −1.37) and the length of elytra-to-height of body ratio (Le/Hb, As = 0.65).
A reliably positive excess was observed in height of body (Hb, Ex = 0.73), the length of body-to-length of elytra ratio (Lb/Le, Ex = 1.35) and the length of body-to-height of body ratio (Lb/Hb, Ex = 1.75); a negative excess was observed in the length of body-to-height of body ratio (Lb/Hb, Ex = −1.08). There was no significant asymmetry or excess in length of elytra (Le). The studied characteristics showed no significant indicators in CV.
In the pupae collected in Park 40th anniversary of the liberation of Dnipro, none of the studied characteristics showed a reliable asymmetry, except for length of elytra (Le, As = 0.62). However, a significant negative excess was observed for length of body (Lb, Ex = −0.80), the length of body-to-length of elytra ratio (Lb/Le, Ex = −0.76) and the body-length-to-height ratio (Lb/Hb, Ex = −1.14). Significant asymmetry and excess were not shown in height of body (Hb) and in the length of elytra-to-height of body ratio (Le/Hb). The coefficient of variation was significant in terms of height of body (Hb), the length of body-to-length of elytra ratio (Lb/Le) and the length of body-to-height of body ratio (Lb/Hb). Significant asymmetry among pupae collected in T. G. Shevchenko Park was found in the length of body (Lb, As = −1.17) and height of body (Hb, As = 0.65). Significant negative excess was observed in terms of the length of elytra (Le, Ex = −0.83) and in the length of elytra-to-height of body ratio (Le/Hb, Ex = −1.21). Significant asymmetry and excess were not shown in the length of body-to-height of body ratio (Lb/Hb).
Pupae collected in Dnipro-Oril Nature Reserve showed a significant positive asymmetry in length of elytra (Le, As = 2.19), as well as in the length of body-to-length of elytra ratio (Lb/Le, As = −2.32). The significant excess was seen in all the characteristics studied, except for the length of body-to-height of body ratio (Lb/Hb): by length of body (Lb, Ex = −0.95), height of body (Hb, Ex = −0.88) and length of elytra (Le, Ex = 4.82). Significant asymmetry and excess were not shown in the length of body-to-height of body ratio (Lb/Hb).
5.2 Characteristics of morphometric variability of population groups in different urbocenoses
Based on the results of a one-way analysis of variance based on linear characteristics, a significant difference (P < 1*10−6) between datasets obtained in different urbocenoses. By length of body (Lb), intergroup sum of squares was greater than intragroup sum of squares. By other linear characteristics, intragroup sum of squares was greater than intergroup ones. Based on a one-way analysis of variance ANOVA of interpopulation variability of index characteristics, Macrosaccus robiniella (as in the case of linear characteristics) showed a significant (P < 0.001) difference between urbocenoses (Table 5) for all the indices studied. Intragroup sum of squares was greater than that of all the indices studied.
Characteristics
Urbo
n
x ± SD
SSi
SSb
F
P
Lb/Le
1
16
1.64 ± 0.11
2.064
1.0733
13.9359
<1*10−6
2
34
1.79 ± 0.17
3
18
1.92 ± 0.13
4
22
1.76 ± 0.08
5
22
1.85 ± 0.10
6
28
1.67 ± 0.10
Lb/Hb
1
16
4.55 ± 0.86
30.7168
5.3343
4.6541
0.0006
2
34
4.11 ± 0.42
3
18
4.10 ± 0.37
4
22
4.60 ± 0.51
5
22
4.40 ± 0.33
6
28
4.23 ± 0.36
Le/Hb
1
16
2.76 ± 0.45
9.14
4.9003
14.3685
<1*10−6
2
34
2.30 ± 0.20
3
18
2.14 ± 0.18
4
22
2.61 ± 0.28
5
22
2.38 ± 0.17
6
28
2.54 ± 0.27
Table 5.
One-way analysis of variance ANOVA of morphological features variability of Macrosaccus robiniella index characteristics (n = 140).
Note: Urbo—number of urbocenosis; x ± SD, SSi—sum of squares within the populations studied; SSb—sum of squares between the populations studied; F—Fisher value; P—degree of confidence.
Morphological variability is one of the manifestations of adaptations that develop the accommodation of living organisms to environmental changes. Effects of factor influence can be accumulated in biological objects over a certain period of time. Morphological features of living organisms depend largely on their habitat. This is conditioned by the peculiarities of the feeding, the breeding season, adaptation to a particular environment, etc. Morphological variability is characterized by changes in weight and linear indicators; it is the result of the influence of environmental factors.
Morphological variability of the population is an instance of general genetic polymorphism and an indicator of potential population stability under conditions of high anthropogenic load on natural ecosystems. The study of morphological variability of invertebrates allows us to assess the capacity of a population to maintain its constancy, the possibility of changes within the same species and deviations from the average size, as well as to make an assessment of environmental quality.
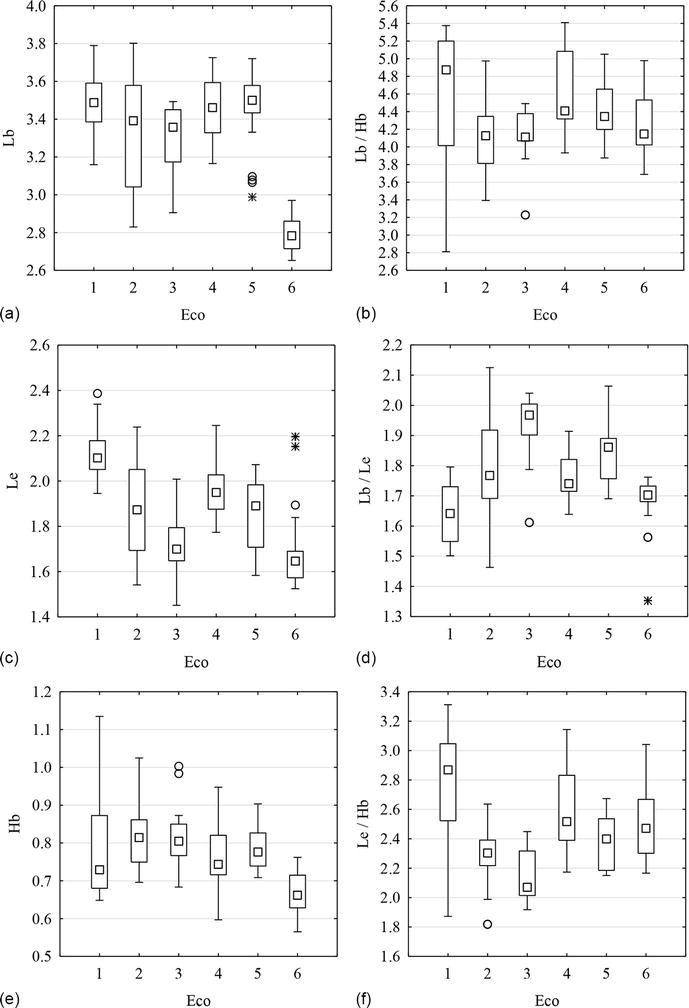
According to the results of our study, significant differences in all linear characteristics and indices between groups of Macrosaccus robiniella pupae were found. Each group corresponded to the urbocenosis in which the leaves damaged by the leafminer were collected. However, on the constructed box graphs shown in Figure 4, it can be seen that the distribution and values of both linear characteristics and indices in the initial five urbocenoses were very different from the sixth urbocenose. The sixth urbocenose, Dnipro-Oril Nature Reserve, being located at a considerable distance from the city center and industrial areas, can act as a reference site aimed to comparison of its characteristics with urbocenoses located within the city. These differences may be caused by significantly different environmental conditions (Figure 4a). Pupae collected in the Dnipro-Oril Nature Reserve show less polymorphism compared to ones collected within the city in terms of length of body, height of body, length of elytra and the length of body-to-length of elytra ratio (Figure 4a,с–e).
Figure 4.
Box plots of Macrosaccus robiniella variability: A—length of body, b—the length of body-to-height of body ratio, c—length of elytra, d—the length of body-to-length of elytra ratio, e—height of body, f—the length of elytra-to-height of body ratio.
Polymorphism within the individual population group in terms of two linear characteristics and three indices was greater than that between different population groups. From this, we can conclude that the diversity of pupae within a single population group or urbocenose was greater than the diversity of pupae collected in different urbocenoses. That is, environmental conditions within the city have little effect on the leafminer pupae diversity. However, morphometric characteristics can vary quite a lot in the pupae collected within the city compared to the pupae collected outside the city.
Studies of morphometric characteristics of Macrosaccus robiniella pupae have shown that both linear characteristics and indices of pupae in the population were stable relative to the averages, since a significant coefficient of variation was observed only in pupae collected in Pridneprovsky Park. This may be conditioned by an insufficient dataset size, so a similar analysis of a larger dataset should be conducted in further studies. However, asymmetry and excess coefficients indicate that most of the studied population groups of Macrosaccus robiniella showed a tendency to deviate from the normal distribution. Moreover, in this case, the dataset obtained in the Dnipro-Orel Nature Reserve also differed greatly: the higher coefficient of excess for length of elytra and for the length of body-to-length of elytra ratio was found compared to the coefficients in other urbocenoses. The population group closest to this parameter was the group collected in Druzhby Narodiv Forest Park which also underwent less anthropogenic pressure compared to the city center. A high excess coefficient indicates a tendency to reduce the number of specimens with characteristics that differ significantly from the averages.
However, in other population groups, on the contrary, an increase in the number of individuals potentially differ greatly from the averages was observed. This may be due to the fact that Macrosaccus robiniella within the city limits was affected by more rigorous environmental conditions, so the specimen unable to be adapted was removed from the population. The asymmetry coefficient also indicates similar processes in populations: specimen living in the urban environment showed a tendency to a deviation of various indicators, while in the Dnipro-Orel Nature Reserve, the tendency to a deviation was manifested only in the length of elytra, but the other indicators studied were relatively stable. Such processes in populations indicated that Macrosaccus robiniella pupae were influenced by stressful factors to a certain extent, in particular, by anthropogenic ones.
6. Influence of feeding of invasive Gracillariidae caterpillars on biochemical processes in the leaves of host (food) plants
Studying of relationships between host plants and phytophage at the biochemical level is important for understanding of the adaptational pattern of black locust plants to the action of the leafminers Pаrectopа robiniellа and Macrosaccus robiniella. During the staying in the host’s body, complex physiological processes occur between the parasite and the plant resulted in the phytophage survival or death. When the pest insect survives, it affects the host body causing changes in metabolism that result in changes in the plant’s functional state [9]. In response to plant damage by phytopathogens and pest insects, such protective reactions are initiated as increased barrier properties (lignification), gene expression, synthesis of secondary metabolites, and protective proteins [10]. The universal stress-protective systems of the plant body include an enzymatic antioxidant complex that provides neutralization of reactive oxygen species (ROS), the level of which increases under stressful effects [11, 12, 13, 14, 15, 16]. Peroxidases are known to be very labile enzymes that respond to cellular homeostasis disruption under the action of stressors of various origins [17, 18, 19, 20].
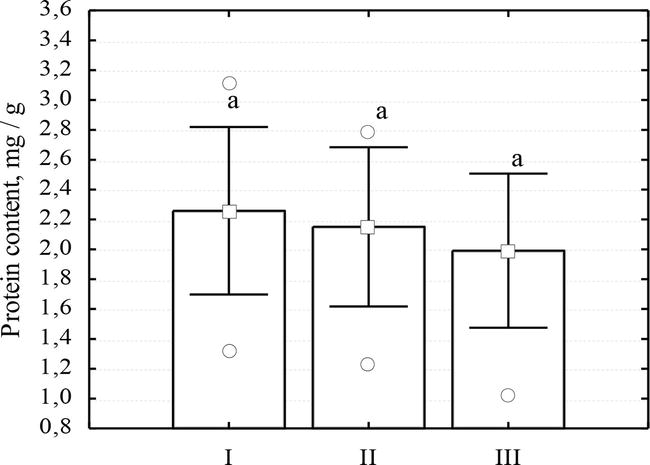
In order to find out the consequences of the impact of the leafminers P. robiniella and M. robiniella on the functional state of R. pseudoacacia trees, the activity and composition of the following antioxidant enzymes were determined: benzidine peroxidase (BPOD), guaiacol peroxidase (GPOD), as well as the content of highly soluble proteins. Highly soluble proteins are among the biochemical indicators that respond actively to cell damage by insects [21, 22, 23, 24]. Our data show a slight fluctuation in the level of soluble proteins of black locust leaves between experimental samples; their values varied in the range of 1.99–2.26 mg/g (Figure 5).
Figure 5.
Soluble protein content in leaves of R. pseudoacacia under the action of Pаrectopа robiniellа and Macrosaccus robiniella: I—Leaves without mines (mine-free leaves); II—Leaves with mines formed by P. robiniella; leaves with mines formed by M. robiniella.
In the leaves with mines formed both by P. robiniella and M robiniella, there was a tendency to reduce the protein content under the influence of M. robiniella to a greater extent (the protein level decreased by 11.9%) compared to the action of P. robiniella. The difference between the reference and the experiment samples under the action of P robiniellа was 4.9%. The average range of variability of this parameter was determined to be 21.5–27.1%.
Reducing the soluble protein content in the phase of active outflow of assimilates is a normal physiological process, but this process in a leaf damaged by a pest insect can be accelerated compared to reference.
The protective enzymatic antioxidant system in the cell comprises a series of enzyme systems including peroxidases (BPODs) actively involved in the mechanisms of plant adaptation to both abiotic and biotic influences [9, 17, 25]. Being regulated a number of processes directly or indirectly related to plant resistance to insect pests, peroxidases can be induced in a number of plants in response to insect attack, and they act as an important component of the direct plant response to damage [26, 27]. Our research has shown that the peroxidase system intended for detoxification of the effects of oxidative stress caused by the action of P. robiniella and M. robiniella is activated in the leaves of R. pseudoacacia trees (Table 6).
Variants of invasion of leaves
n
BPOD, U/g FW min (middle-aged trees)
СV, %
n
BPOD, U/g FW min (5-year-old tree)
СV, %
n
GPOD, mm TG guaiacol/g FW min
СV, %
Mine-free leaves
24
592.13 ± 140.77a
23.8
4
728.90 ± 3.57a
0.49
30
6.09 ± 2.87a
47.1
P. robiniella
34
737.59 ± 408.25a
55.3
4
2784.81 ± 20.68b
0.74
26
9.94 ± 5.10b
51.3
М. robiniella
24
948.42 ± 320.27b
33.8
—
—
—
12
15.97 ± 5.92c
37.1
Table 6.
Total activity of antioxidant enzymes (BPOD, GPOD) in Robinia pseudoacacia leaves under the action of Parectopa robiniella and Macrosaccus robiniella (mean ± SD).
Notes: values in column marked with different letters (a, b, с) were significantly different according to Tukey’s t-test P < 0.05. BPOD—benzidine peroxidase; GPOD—guaiacol peroxidase; TG—tetraguaiacol; U—units; FW—fresh weight.
In 35–40-year-old trees with the presence of leaf blotch miners P. robiniella, the leaves showed a tendency to increase BPOD activity by 24.6% compared to reference (F = 2.80; p = 0.10). A significant variability (55.3%) of this indicator was found. A significant increase in benzidine peroxidase activity was noted under the action of leaf blotch miner M. robiniella at the level of 60.2% (F = 24.89; P = 9.11.10−6) compared to reference and at the level of 28.6% compared to the action of P. robiniella (F = 4.46; p = 0.04).
Five-year-old trees showed the highest response to the damages caused by P. robiniella: in leaves damaged by the pest, activity of the enzyme increased by 3.8 times compared to reference (F = 3,8.10−5; Р = 1.19.10−12). A significantly increased GPOD response to leaf damage by the invasive insects was shown in the presence of both P. robiniella and M. robiniella. In the presence of P. robiniella, the activity value increased by 63.2% (F = 9.51; P = 3.4.10−3), and under the action of M. robiniella it increased by 2.6 times (F = 48.12; p = 3.44.10−3). A significant difference in enzyme activity was also recorded between Parectopa robiniella and Macrosaccus robiniella. GPOD activity of the latter species was higher by 60.7% (F = 8.77; P = 5.7.10−3).
Peroxidases are often present in multiple isoforms, which is consistent with different physiological functions of the peroxidase family [28, 29]. Acidic and basic isoperoxidases are associated with the mechanisms of stress in plants. Most of the scientific works were devoted to study the effect of phytopathogens on activity and composition of the peroxidase system [30]. In the scientific paper was shown that activation of the basic peroxidases occurs at the beginning of infection, and alterations associated with auxin and ethylene metabolism induce intensified synthesis of acidic isoenzymes as a later stage of plant response or defense [26].
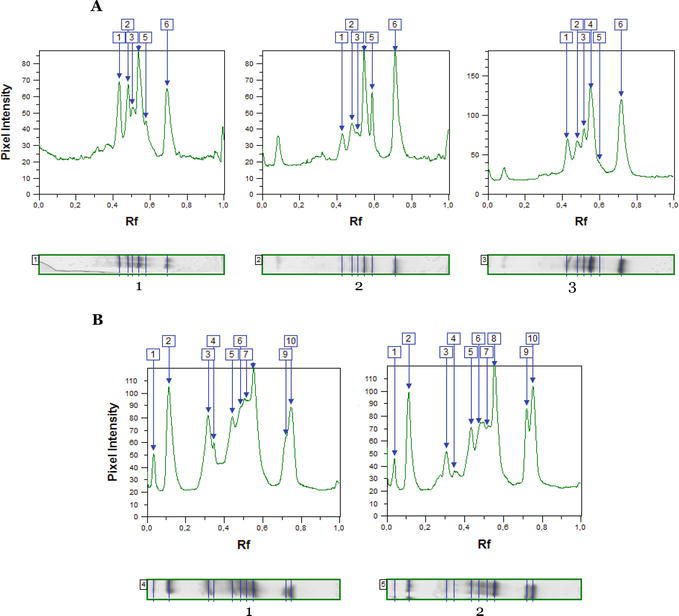
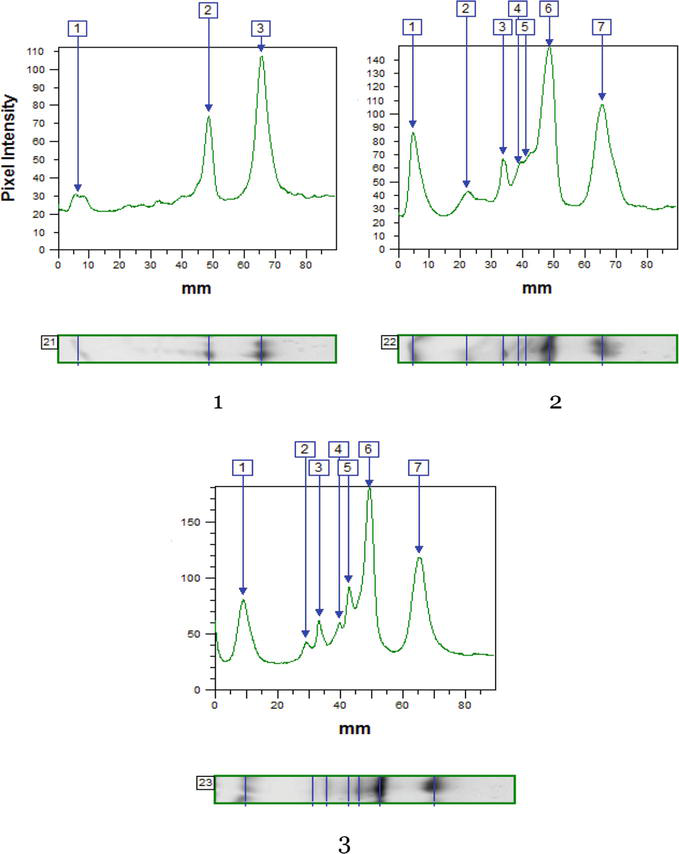
It was found that active rearrangements of the peroxidase system in the leaves occur when black locusts are affected by the leaf-mining insects. The stressful situation caused by pest attacks leads to changes in the expression of isoperoxidases in black locust leaves in the ecologically favorable area of Oles Honchar Dnipro National University Botanical Garden, where six isoforms were clearly visualized (Figure 6; Tables 7 and 8).
Figure 6.
IEF benzidine peroxidase profiles of leaves of 30- to 40-year-old (A) and 5-year-old (B) trees of black locust sampled in the botanical garden under the influence of invasive insects; 1—Mine-free leaves; 2—Leaves with mines formed by P. robiniella; 3—Leaves with mines formed by M. robiniella; Rf—Electrophoretic mobility.
Variants of invasion of leaves
Rf value of isoperoxidase
0.08
0.43
0.48
0.51
0.54
0.71
Mine-free leaves
17.89 ± 0.39a
16.10 ± 0.46a
10.39 ± 0.48a
24.24 ± 0.66a
9.74 ± 1.99a
21.64 ± 0.99a
P. robiniella
12.19 ± 0.57b
13.63 ± 1.03b
5.32 ± 0.25b
25.70 ± 0.24b
14.35 ± 0.56b
28.81 ± 2.33b
М. robiniella
13.49 ± 0.04c
12.58 ± 1.15b
10.82 ± 0.06a
27.74 ± 0.16c
6.22 ± 0.64c
29.17 ± 1.14b
Table 7.
The relative content of BPOD isoforms in the leaves of middle-aged Robinia pseudoacacia trees under the action of Pаrectopа robiniellа and Macrosaccus robiniella (n = 3; ± SD).
Variants of invasion of leaves
Rf value of isoperoxidase
0.03
0.08
0.30
0.34
0.43
0.48
0.51
0.54
0.71
0.75
Mine-free leaves
4.13 ± 0.11a
15.23 ± 0.36a
12.64 ± 0.32a
3.59 ± 0.09a
13.03 ± 0.80a
8.25 ± 0.21a
9.68 ± 0.15a
15.66 ± 0.52a
6.01 ± 0.25a
11.77 ± 0.30a
P. robiniella
4.18 ± 0.08a
13.43 ± 0.26a
9.06 ± 0.18b
3.21 ± 0.25a
11.28 ± 0.72a
10.40 ± 0.20b
7.56 ± 0.77a
18.24 ± 0.74b
8.25 ± 0.08b
14.39 ± 0.38b
Table 8.
The relative content of BPOD isoforms in the leaves of 5-year-old Robinia pseudoacacia trees which grow in Oles Honchar Dnipro National University Botanical Garden, under the action of P. robiniella and M. robiniella (n = 3; ± SD).
Benzidine peroxidase isolated from the leaf tissue of medium-aged black locust trees was divided into six fractions with Rf values from 0.08 to 0.76 (Figure 5a and Table 7). The relative content of all six isoperoxidases in leaves damaged by the pests differs from the reference samples. In trees of black locust, the proportion of 0.51 and 0.71 isoforms in the samples studied was significantly higher compared to the reference trees by 6.0 and 13.1% (Pаrectopа robiniella) and 14.4 and 34.8% (Macrosaccus robiniella), respectively. In the leaves of young R. pseudoacacia trees sampled in the Botanical Garden (in contrast to 30–40-year-olds), 10 peroxidase isoforms were isolated (Figure 5), and the relative content of components with Rf 0.48, 0.54, 0.71 and 0.75 was increased by 26.1, 16.5, 37.3 and 22.3%, respectively (Table 8).
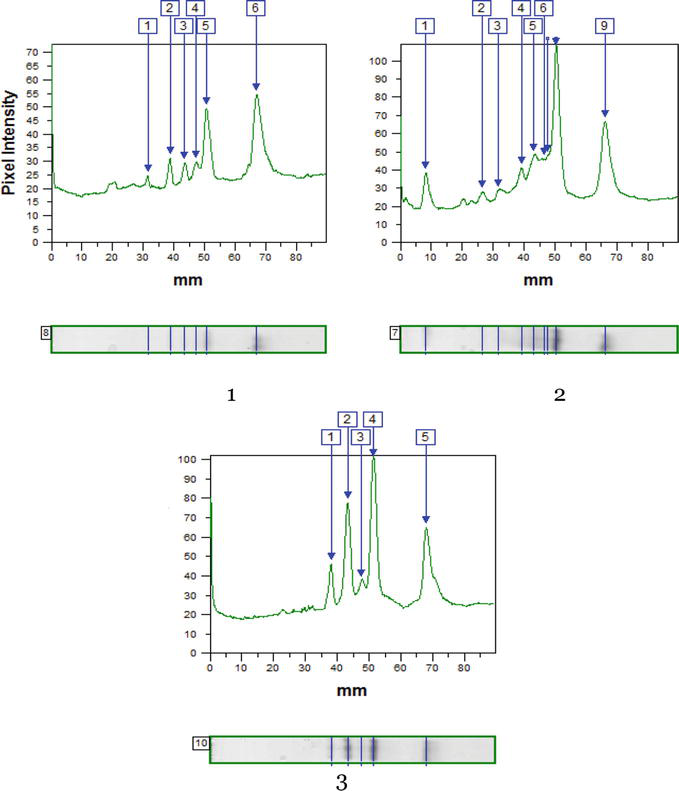
In the leaves of R. pseudoacacia trees sampled in T. G. Shevchenko Park, more significant changes in the spectrum of BPOD isoforms were observed (Figure 7). Under the action of P. robiniella, two isoforms of peroxidase were isolated with Rf 0.09 and 0.29; these isoforms were absent in the reference samples and in the samples affected by M. robiniella (Table 9).
Figure 7.
Alterations of IEF peroxidase profiles in leaves of R. pseudoacacia trees sampled in T. G. Shevchenko Park under the action of P. robiniella and M. robiniella.
Variants of invasion of leaves
Rf value of isoperoxidase
0.08
0.30
0.37
0.43
0.48
0.51
0.54
0.56
0.75
Mine-free leaves
—
—
3.83 ± 2.52a
10.08 ± 0.72a
8.87 ± 3.81a
—
8.36 ± 3.53a
23.40 ± 10.8a
34.64 ± 15.1a
P. robiniella
9.14 ± 0.55
4.90 ± 0.86
6.41 ± 0.84b
8.61 ± 0.35b
10.78 ± 0.71b
5.90 ± 0.81
4.23 ± 0.27b
26.57 ± 1.05a
25.15 ± 1.88b
M. robiniella
—
—
—
10.62 ± 0.11a
17.31 ± 11.2c
—
6.69 ± 4.11с
22.74 ± 14.8a
20.83 ± 13.4c
Table 9.
The relative content of BPOD isoforms in leaves of different-aged Robinia pseudoacacia trees sampled in T. G. Shevchenko Park under the action of P. robiniella and M. robiniella (n = 3; ± SD).
Under the action of M. robiniella, the number of components decreased by one isoform (with Rf 0.51) compared to mine-free leaves. Common areas with reference have values of Rf 0.43, 0.48, 0.54, 0.56 and 0.75. The latter isoform reduces activity under the action of the pest insects: by 27.4% (P. robiniella) and by 39.8% (M. robiniella). The activity of the component with Rf 0.48 significantly increases (by 21.5 and 92.5% according to the variants affected by P. robiniella and M. robiniella). A decrease in the relative proportions under the action of the insects was noted for isoperoxidase with Rf 0.54 (by 49.4 and 20.0%, respectively, according to the variants affected by P. robiniella and M. robiniella). Study of peroxidase in leaves of black locust trees undamaged by pests sampled in Pridneprovsky Park showed only three isoforms in their composition. The main activity occurred in the component with Rf 0.75 (Figure 8 and Table 10).
Figure 8.
Alterations of IEF peroxidase profiles in leaves of R. pseudoacacia trees sampled in Pridneprovsky Park under the action of P. robiniella and M. robiniella (n = 3; ± SD).
Variants of invasion of leaves
Rf value of isoperoxidase
0.08
0.25
0.34
0.37
0.43
0.48
0.54
0.75
Mine-free leaves
17.25 ± 0.41a
—
—
—
—
—
28.46 ± 0.63a
54.29 ± 1.05a
P. robiniella
14.54 ± 0.50b
7.33 ± 0.59
—
8.81 ± 0.31
3.98 ± 0.61
8.94 ± 0.74
29.47 ± 0.55a
26.92 ± 0.94b
M. robiniella
16.35 ± 0.64а
—
4.72 ± 0.18
6.49 ± 0.26
5.36 ± 0.84
9.53 ± 0.77
29.13 ± 1.12a
28.40 ± 0.52b
Table 10.
The relative content of BPOD isoforms in leaves of different-aged Robinia pseudoacacia trees sampled in Pridneprovsky Park under the action of P. robiniella and M. robiniella (n = 3; ± SD).
The leaves damaged by P. robiniella and M. robiniella showed seven active zones. Molecular forms with Rf 0.08, 0.54, and 0.75 were found to be isoperoxidases common with the reference. Compared to mine-free leaves, leaves damaged by P. robiniella and M. robiniella components with Rf 0.08 and 0.75 showed a decrease in enzyme activity by 15.9 and 5.2%, and with Rf 0.75—by 50.2 and 47.7%, respectively.
Therefore, the increase in BPOD and GPOD activity established by us under the action of leaf blotch miners on the leaves of black locusts indicates the involvement of peroxidases in mechanisms of plant defense against the attacks of phytophagous insect, which was also noted in a number of other studies [2, 22, 31, 32, 33]. The role of antioxidants consists both in neutralizing reactive oxygen species with the involvement of hydrogen peroxide (benzidine peroxidase) and in protecting cell walls by their lignification to prevent pest invasion (guaiacol peroxidase). Summarizing the data on the electrophoretic profiles of benzidine peroxidase in black locust leaves, it should be noted that peroxidase in the leaves of the trees studied has a fairly wide range of isoforms (a total of 13 molecular forms were recorded) in the acidic pH range (3.5–6.5), and changes in the level of their expressiveness and the number of components in the system may indicate their involvement in the processes of cellular defense against the complex action of stressors of both biotic and abiotic origins. We have shown that molecular forms of peroxidase located in the most acidic pH range (Rf values from 0.51 to 0.75) were subjected to the main load in the peroxidase system under the influence of stress caused by mechanical damage of leaf tissue. Thus, in middle-aged trees, the main BPOD activity was inherent in isoforms 0.51 and 0.71; their proportion in the total spectrum was 45.9% (reference leaves), 51.5% (leaves with mines formed by P. robiniella) and 56.9% (leaves with mines formed by M. robiniella). In young trees, the relative content of the components was distributed more evenly, indicating a fairly high level of complexity of the peroxidase system in young trees. In the leaves of trees sampled in T. G. Shevchenko Park, high activity was focused in areas with Rf 0.56 and 0.75. The highest activity was registered in the reference leaves. This amounted to 58.0% and was slightly lower in the damaged leaves: 51.7 (in the leaves with mines formed by P. robiniella) and 43.6% (in the leaves with mines formed by M. robiniella).
In Pridneprovsky Park, the main activity was also observed in two isoperoxidases with Rf 0.54 and 0.75, which accounted for more than 50% of all molecular forms of peroxidase. A number of studies have noted that overexpression of anionic peroxidase contributes to plant resistance to the action of lepidopteran and coleopteran insect pests [34, 35]; the peroxidase exhibits the capacity to detoxify ROS and is involved in lignin and suberin synthesis; cationic peroxidases can participate in the production of hydrogen peroxide and have NADPH oxidase activity.
The data obtained allow us to conclude that R. pseudoacacia is a tree species highly sensitive to the invasion by the larvae of leaf blotch miners that feed on its leaves. It is also necessary to consider the anthropogenic pressure exerted by the environment on both Robinia pseudoacacia and leafminers. So T. G. Shevchenko Park and Pridneprovsky Park are located in the zone of intensive sedimentation of harmful impurities emitted by a thermal power plant, a pipe rolling plant and motor vehicles. Therefore, we observed more significant changes in IEF peroxidase profiles of black locust trees growing in these areas: Depletion of enzyme spectra or almost complete decrease in the intensity of certain components of the system was recorded in addition to alterations in the expressiveness of ВРОD isoforms.
7. Effect of feeding of invasive Gracillariidae caterpillars on parameters of chlorophyll fluorescence induction curves in host plants
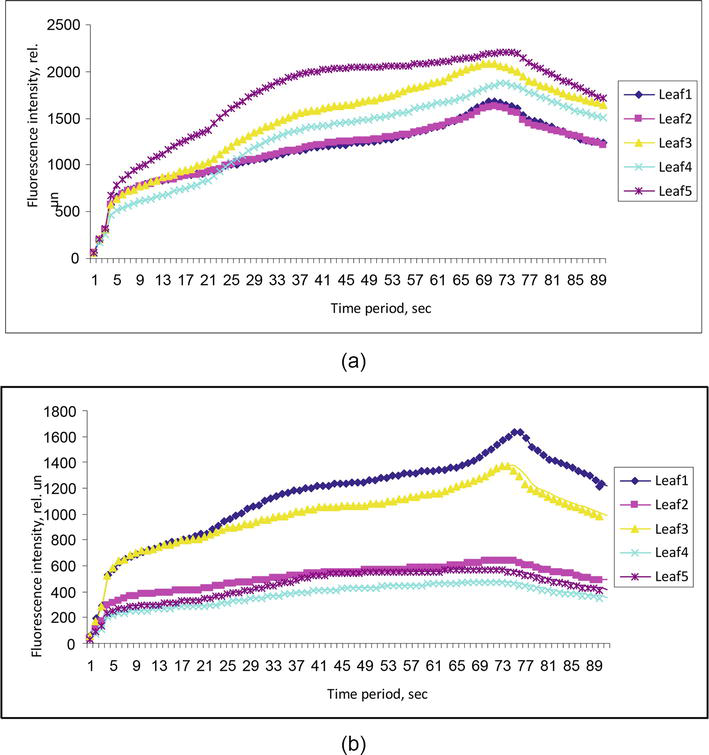
Using a portable fluorometer “Floratest” on fresh A. hippocastanum leaves both damaged and undamaged by Cameraria ohridella, and we obtained data that clearly demonstrate the effect of the leafminer’s caterpillars feeding on chlorophyll fluorescence intensity (the Kautsky curve) (Figure 9).
Figure 9.
Chlorophyll fluorescence induction curves (the Kautsky curves): A—Leaves undamaged by the insect (n = 5); b—Leaves damaged by the insect (n = 5).
Under normal conditions, the fluorescence level was insignificant (1–2% of the total absorbed light), which indicates the active use of absorbed light energy by cells. After lightening the leaves of plants adapted to the dark, a sharp increase in the intensity of chlorophyll fluorescence as a fast phase was observed in the first few seconds. Then, a gradual decrease occurred within a few minutes through certain stages, from the slow phase to the stationary level of FSt [36].
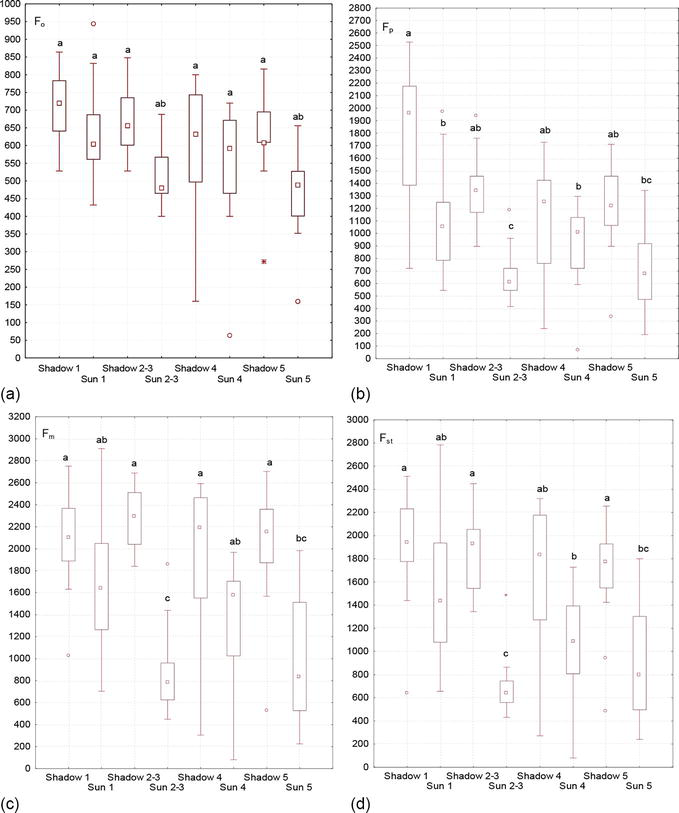
At the initial time point, all channels of photosynthetic electron transfer are open, and the maximum energy of electron excitation goes into the photosynthetic process. During this period, chlorophyll fluorescence is minimal, and its intensity on the Kautsky curve is denoted by the letter F with the “0” index, i.e., F0. Background fluorescence index (F0) characterizes the amount of inactive chlorophyll that has no functional connections with the reaction centers; that is, it acts as the initial level of ChlF. It depends on the loss of excitation energy during migration along with the pigment matrix. F0–Fр transition is associated with a decrease in electron transport. It characterizes the thermal adaptation period of the leaf. The transition is steeper when the adaptation period is short, but the transition is slower when the adaptation period is large [5]. Fm parameter indicates the highest level of fluorescence, which is recorded as the maximum on the induction curve. The entire F0–Fm area is called the fast phase of fluorescence. The slow phase of chlorophyll fluorescence induction represents all transitions of the induction after reaching the maximum value (peak). The steady-state fluorescence level (FSt) is characterized by a dynamic equilibrium between the processes that cause an increase in fluorescence and the processes that lead to its decrease. During the caterpillar development (5 generations of Cameraria ohridella caterpillars), a gradual decrease in the values of all the main indicators (F0, Fm, Fp and FSt) occur in the chlorophyll fluorescence induction curve (the Kautsky curve) (Figure 10) of damaged A. hippocastanum leaves.
Figure 10.
Variability of indicators of the chlorophyll fluorescence induction curve (the Kautsky curve) in Aesculus hippocastanum leaves damaged by Cameraria ohridella: F0—The initial value of fluorescence induction after switching on the light; Fp—A “plateau” value of fluorescence induction; Fm—A maximum value of fluorescence induction; FSt—A steady-state value of fluorescence induction after light adaptation of the plant leaf.
A significant difference in the influence of different-aged caterpillars and leaf position in tree crowns was established for all the main indicators of the chlorophyll fluorescence induction curve. The only exception is the initial value of fluorescence induction (F0) for which no such dependence has been established.
Long-term studies of invasive Gracillariidae as components of artificial and natural ecosystems in Ukraine allowed us to find out the scale of their invasion, mechanisms of invasion into various ecosystems, adaptive capabilities in a novel environment, features of trophic relationships and influence on the physiological and biochemical state of host plants which can be confirmed by the following conclusions:
According to the statistical data processing, the largest number of mines formed by invasive Gracillariidae on the leaves of host plants was recorded in ecosystems environmentally beneficial for these host plants, where the plant age and the position of tree plantation above sea level acted as key factors; for urban plantings, key factor was the content of heavy metals (Zn, Cu, Pb and Cd) in leaf tissues.
Statistical data obtained during the study of mines showed a high variability (40.1–100%) of morphometric indicators in most cases. According to the results of the study, both coefficient of variation and standard deviation indicated that the morphometric plasticity of mines formed by Parectopa robiniella showed a more variable polymorphism than that of mines formed by Macrosaccus robiniella.
Polymorphism of Parectopa robiniella pupae collected in green areas of the city was larger compared to the pupae collected in the Nature Reserve (reference) ecosystem. For all linear characteristics and indices, there were significant intergroup differences of Parectopa robiniella pupae collected in different urbocenoses by the level of anthropogenic load.
Polymorphism of two linear characteristics and three indexes of six studied characteristics within a single population group were greater than polymorphism established for different population groups. From this, it can be concluded that the morphological diversity of pupae within a single group or urbocenose was greater than that in different urbocenoses. That is, environmental conditions within the city have a slight effect on the variety of morphometric parameters in the pupae. However, morphometric characteristics can vary quite a lot in the pupae collected within the city compared to the pupae collected outside the city.
In the leaves of the host plants, an increase in the activity of both benzidine and guaiacol peroxidase was found under the conditions of invasive insect caterpillars feeding on the host plant. The enzyme activity level depended on the age of the trees and the environmental conditions in which the trees grow. In 30–40-year-old trees, BPOD activity increased by 24.6% under the influence of caterpillars on the leaves compared to the reference. A significant increase in benzidine peroxidase activity was detected under the action of Macrosaccus robiniella (by 60.2%) compared to the reference, and by 28.6% compared to the action of Parectopa robiniella. A 5-year-old tree showed the highest response to the action of Parectopa robiniella: the enzyme activity increased by 3.8 times in leaves damaged by the pest insect compared to the reference.
Active rearrangements of the peroxidase system in the leaves of host plants indicate the involvement of benzidine peroxidase and guaiacol peroxidase in plant defense mechanisms against the action of Gracillariidae caterpillars. Benzidine peroxidase is probably involved in the neutralization of reactive oxygen species with the participation of hydrogen peroxide, and guaiacol peroxidase is involved in protecting of cell walls by lignification and suberization to prevent pest invasion.
Caterpillar feeding affects the functional state of the host plant which was confirmed by changes in the soluble protein content. The studies have shown a decrease in the content of highly soluble proteins in leaves damaged by invasive insects. The protein content was significantly reduced in leaves affected by the invasive insects, as the plant reduced the rate of synthesis of highly soluble protein under biotic stress, and the entire translation mechanism was shifted to the production of proteins associated with the plant defense response.
It was established that the caterpillars of Gracillariidae invasive species affect significantly the photosynthetic apparatus of host plants. This effect was clearly determined with fluorescence analysis. Analysis of the Kautsky curve characteristics of both damaged and undamaged leaves showed that caterpillar feeding affected significantly four critical parameters.
Feeding of Cameraria ohridella caterpillars causes a decrease in the activity of the photosynthetic apparatus of A. hippocastanum regardless of the spatial arrangement of leaves in the tree crown. In damaged leaves, the amount of chlorophyll not involved in photosynthetic energy transfer to reaction centers increases. Conversely, a constant decrease in the quantum efficiency of PS II (inhibition of photosynthetic activity) was observed during the development of only a single Cameraria ohridella generation.
References
1.Kawahara AY, Plotkin D, Ohshima I, Lopez-Vaamonde C, Houlihan PR, Breinholt JW, et al. A molecular phylogeny and revised higher-level classification for the leaf-mining moth family Gracillariidae and its implications for larval host-use evolution. Systematic Entomology. 2017;42:60-81
2.Shupranova LV, Holoborodko KK, Seliutina OV, Pakhomov OY. The influence of Cameraria ohridella (Lepidoptera, Gracillariidae) on the activity of the enzymatic antioxidant system of protection of the assimilating organs of Aesculus hippocastanum in an urbogenic environment. Biosystems Diversity. 2019;27(3):238-243. DOI: 10.15421/011933
3.Holoborodko KK, Rusynov VI, Seliutina OV. Addition to analysis of morphological parameters of mines on two invasive leaf-mining Lepidoptera species (Parectopa robiniella (Clemens, 1863) and Phyllonorycter robiniella (Clemens, 1859)) on black locust. Problems of Bioindications and Ecology. 2018;23(2):134-141. DOI: 10.26661/2312-2056/2018-23/2-09
4.Gregory RPF. A rapid assay for peroxidase activity. The Biochemical Journal. 1966;101(3):582-583
5.Ranieri A, Castagna A, Baldam B, Soldatini GF. Iron deficiency differently affects peroxidase isoforms in sunflower. Journal of Experimental Botany. 2001;52(354):25-35
6.Rühle T, Reiter В, Leister D. Chlorophyll fluorescence video imaging: A versatile tool for identifying factors related to photosynthesis. Frontiers Plant Science. 2018;9(55):1-13
7.Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry. 1976;72:248-254. DOI: 10.1016/0003-2697(76)90527-3
8.Fodor E, Hâruţa O. Niche partition of two invasive insect species, Parectopa robiniella (Lepidoptera, Gracillariidae) and Phyllonorycter robiniella (Clem.) (Lepidoptera, Gracillariidae). Research Journal of Agricultural Science. 2009;41(2):261-269
9.Mithöfer A, Boland W. Plant defense against herbivores: Chemical aspects. Annual Review Plant Biology. 2012;63:431-450. DOI: 10.1146/annurev-arplant-042110-103854
10.Bagnoli F, Capuana M, Racchi ML. Developmental changes of catalase and superoxide dismutase isoenzyme, in zygotic and somatic embryos of horse chestnut. Australian Journal of Plant Physiology. 1998;25(8):909-913. DOI: 10.1071/PP98068
11.Dowd PF, Hermis DA, Berhow MA, Lagrimini LM. Mechanism of insect resistance in transgenic plants (over) expressing a tobacco anionic peroxidase. Plant Peroxidase Newsletter. 2000;14:93-101
12.Hirayama T, Shinozaki K. Research on plant abiotic stress responses in the post-genome era: Past, present and future. The Plant Journal. 2010;61:1041-1052. DOI: 10.1111/j.1365-313X.2010.04124.x
13.Huseynova IM, Aliyeva DR, Mammadov AC, Aliyev JA. Hydrogen peroxide generation and antioxidant enzyme activities in the leaves and roots of wheat cultivars subjected to long-term soil drought stress. Photosynthesis Research. 2015;125:279-289. DOI: 10.1007/s11120-015-0160-7
14.Jia X, Zhao YH, Liu T, He YH. Leaf defense system of Robinia pseudoacacia L. seedlings exposed to 3years of elevated atmospheric CO2 and Cd-contaminated soils. Science of the Total Environment. 2017;605-606:48-57. DOI: 10.1016/j.scitotenv.2017.06.172
15.Koch KG, Chapman K, Louis J, Heng-Moss T, Sarath G. Plant tolerance: A unique approach to control hemipteran pests. Frontiers in Plant Science. 2016;7:1363. DOI: 10.3389/fpls.2016.01363
16.Lushchak VI. Free radicals, reactive oxygen species, oxidative stresses and their classifications. Ukrainian Biochemical Journal. 2015;87(6):11-18. DOI: 10.15407/ubj87.06.011
17.Batra S. Toxicity mediated oxidative stress and its mitigation strategies in crop plants. Journal of Environmental Engineering and Landscape Management. 2021;29(4):499-508. DOI: 10.3846/jeelm.2021.14382
20.Tognolli M, Penel C, Greppin H, Simon P, et al. Analysis and expression of the class III peroxidase large gene family in Arabidopsis thaliana. Gene. 2002;288(1-2):129-138. DOI: 10.1016/S0378-1119(02)00465-1
21.Ashry NA, Mohamed HI. Impact of secondary metabolites and related enzymes in flax resistance and or susceptibility to Powdery Mildew. World Journal of Agricultural Sciences. 2011;7:78-85
22.Deans CA, Sword GA, Vogel H, Behmer ST. Quantity versus quality: Effects of diet protein-carbohydrate ratios and amounts on insect herbivore gene expression. Insect Biochemistry and Molecular Biology. 2022;145:103773. DOI: 10.1016/j.ibmb.2022.103773
23.Esteban-Carrasco A, López-Serrano M, Zapata JM, Sabater B, Martín M. Oxidation of phenolic compounds from Aloe barbadensis by peroxidase activity: Possible involvement in defense reactions. Plant Physiology and Biochemistry. 2001;39:521-527. DOI: 10.1016/S0981-9428(01)01270-0
24.Michalak A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Polish Journal of Environmental Studies. 2006;15:523-530
25.Staszak AM, Ratajczak E, Leśniewska J, Piotrowska-Niczyporuk A, Kostro-Ambroziak A. A broad spectrum of host plant responses to the actions of the gall midge: Case study of Robinia pseudoacacia L. and Obolodiplosis robiniae (Haldeman). BMC Plant Biology. 2023;23:19. DOI: 10.1186/s12870-022-03914-0
26.Takashima T, Suzuki M, Ishiguri F, Iizuka K, Yoshizawa N, Yokota S. Cationic peroxidase related to basal resistance of Betula platyphylla var. japonica plantlet No. 8 against canker-rot fungus Inonotus obliquus strain IO-U1. Plant Biotechnology. 2013;30(2):199-205. DOI: 10.5511/plantbiotechnology.13.0312b
27.War AR, Paulraj MG, Ahmad T, Buhroo AA, Hussain B, Ignacimuthu S, et al. Mechanisms of plant defense against insect herbivores. Plant Signaling & Behavior. 2013;7(10):1306-1320. DOI: 10.4161/psb.21663
28.Duroux L, Welinder KG. The peroxidase gene family in plants: A phylogenetic overview. Journal of Molecular Evolution. 2003;57:397-407. DOI: 10.1007/s00239-003-2489-3
30.Chamani M, Naseri B, Rafiee-Dastjerdi H, Emaratpardaz J, Ebadollahi A, Palla F. Some physiological effects of nanofertilizers on wheat-aphid interactions. Plants. 2023;12(14):2602. DOI: 10.3390/plants12142602
31.Breda C, Buffard D, Vanhuystee RB, Esnault R. Differential expression of 2 peanut peroxidase cDNA clones in peanut plants and cells in suspension-culture in response to stress. Plant Cell Reports. 1993;12:268-272. DOI: 10.1007/BF00237133
32.Grene R. Oxidative stress and acclimation mechanism in plants. Arabidopsis Book. 2002;1:e0036. DOI: 10.1199/tab.0036.1
33.Seliutina OV, Shupranova LV, Holoborodko KK, Shulman MV, Bobylev YP. Effect of Cameraria ohridella on accumulation of proteins, peroxidase activity and composition in Aesculus hippocastanum leaves. Regulatory Mechanisms in Biosystems. 2020;11(2):299-304. DOI: 10.15421/022045
34.Dowd PF, Lagrimini LM. The role of peroxidase in host insect defenses. In: Carozzi N, Koziel M, editors. Advances in Insect Control: The Role of Transgenic Plants. London: Taylor and Francis; 1997. pp. 195-223
35.Kukavica BM, Jovanovic SD, Menckhoff L, Lüthje S. Cell wall-bound cationic and anionic class III isoperoxidases of pea root: Biochemical characterization and function in root growth. Journal of Experimental Botany. 2012;63(12):4631-4645. DOI: 10.1093/jxb/ers139
36.Martinazzo EG, Ramm А, Bacarin MA. The chlorophyll a fluorescence as an indicator of the temperature stress in the leaves of Prunus persica. Brazilian Journal of Plant Physiology. 2012;24(4):237-246
 Open access peer-reviewed chapter
Open access peer-reviewed chapter