Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
Stem cell therapies hold promise for enhancing infertility treatments through improved differentiation and cytokine secretion mechanisms, particularly autologous stem cells known for safety and compatibility. Collaboration and ongoing research are essential for clinical adoption. Additionally, cell-based treatments offer potential solutions for non-obstructive azoospermia NOA, a condition characterized by absent sperm in ejaculate. Traditional therapies have limitations, prompting exploration of spermatogonial stem cells SSCs and induced pluripotent stem cells iPSCs. Animal studies demonstrate successful fertility restoration via SSC transplantation, and progress has been made in characterizing human SSCs. However, challenges persist in expanding human SSCs and optimizing iPSC differentiation. Further research is necessary to overcome technical hurdles, ensure safety, and offer a novel NOA treatment option, ultimately restoring fertility.
Faculty of Science (Boys), Zoology Department, Al-Azhar University, Cairo, Egypt
Serag Eldin I. Elbehairi
Faculty of Science, Biology Department, King Khalid University, Abha, Saudi Arabia
Cell Culture Lab, Egyptian Organization for Biological Products and Vaccines (VACSERA Holding Company), Agouza, Giza, Egypt
Sayed Bakry
Faculty of Science (Boys), Zoology Department, Al-Azhar University, Cairo, Egypt
Ahmed B.M. Mehany
Faculty of Science (Boys), Zoology Department, Al-Azhar University, Cairo, Egypt
Mahmoud Ashry
Faculty of Science, Zoology Department, Al-Azhar University, Assuit, Egypt
Hussam Askar
Faculty of Science, Zoology Department, Al-Azhar University, Assuit, Egypt
Mohammad Y. Alfaifi
Faculty of Science, Biology Department, King Khalid University, Abha, Saudi Arabia
*Address all correspondence to: ahmed.atwa@azhar.edu.eg
1. Introduction
Spermatogenesis comprises three key stages. Initially, it involves the growth and maturation of spermatogonia. Next, meiosis (I and II) occurs, leading to the creation of haploid cells. Finally, spermiogenesis ensues, involving various biochemical and morphological changes in round spermatids. These alterations encompass chromatin compaction, acrosome formation, and flagellum assembly and elongation, ultimately yielding mature spermatozoa. Any anomalies in these specialized processes can hinder sperm cell production, resulting in NOA, which is known to exhibit significant genetic diversity. Studies have identified mutations in over 600 genes as contributors to reduced fertility in animal models [1]. Moreover, within the testis, 2274 genes are notably active, with 474 exclusively expressed in this organ. Presently, there are only two standard genetic tests for individuals affected by NOA. These tests involve karyotype analysis to detect sex chromosome abnormalities, particularly Klinefelter syndrome (47 XXY) and various translocations, as well as the investigation of microdeletions in the AZF region. However, these tests yield a diagnosis for only around 20% of the individuals studied, indicating that most affected individuals remain undiagnosed [2].
Most couples, regardless of diagnosis, desire natural conception. However, those with NOA face a unique challenge. Their only option for pregnancy is testicular sperm extraction (TESE) followed by in vitro fertilization (IVF) using intracytoplasmic sperm injection ICSI. Success rates are disappointingly low at 30–50%, making the invasive, time-consuming, emotionally distressing, and costly TESE-ICSI process worth avoiding if chances are slim. Physicians should recommend it only when benefits outweigh risks. Without a precise diagnosis, TESE-ICSI might be futile, overlooking quicker options like sperm donation or adoption. Infertile men, relatives, and potential offspring face increased health risks, notably cancer. Genetic diagnosis, providing prognostic value, is crucial for well-informed guidance during TESE and/or endocrine therapy [3, 4].
The World Health Organization (WHO) recognizes male infertility as a major global health issue affecting over 50 million couples. NOA is a condition where sperm are absent in the ejaculate, even after centrifugation and microscopic examination [5]. It affects about 1% of all males and 10% of males with infertility, with a wide range of genetic causes [6].
Azoospermia, the absence of spermatozoa in ejaculates, can be classified into obstructive and non-obstructive NOA forms. Distinguishing between these types is essential as obstructive azoospermia is more favorable, preserving spermatogenesis. Clinical examination, including evaluation of medical history, hormone levels, and physical examination, provides reliable means to differentiate the two types [7]. However, NOA, which accounts for approximately 10% of infertility cases, is characterized by the absence of spermatozoa due to spermatogenic deficiency. Often, azoospermia is associated with irreversible disorders of the testicles related to endocrine, genetic, and inflammatory diseases [8]. Idiopathic NOA, without a known cause, is also possible [9].
Non-obstructive azoospermia is typically characterized by small and flaccid testicles, as determined by palpation and measurement. In all azoospermia cases, measuring hormone levels such as follicle-stimulating hormone (FSH), luteinizing hormone (LH), prolactin, total testosterone, estradiol, and inhibin B is important [10]. In NOA, FSH levels are usually elevated, LH levels are increased or close to normal, and hypogonadism (low total testosterone) is prevalent, indicating a deficiency in Leydig cells obesity may lead to a reduction in testosterone levels and an elevation in serum estradiol levels, primarily as a result of androgen conversion occurring in peripheral tissues [11]. Nevertheless, in obese individuals, decreased testosterone levels could potentially be attributed to adjustments in sex hormone-binding globulin (SHBG) rather than a genuine deficiency in testosterone [7, 12].
Distinguishing obstructive azoospermia from NOA is crucial for treatment choices and the success of sperm extraction surgery. Men with azoospermia need a hormonal assessment, including FSH and total testosterone measurements [13]. NOA causes can be pretesticular (like endocrine issues) or testicular (acquired or congenital), often leading to elevated FSH levels. Common acquired causes include varicocele, orchitis, chemotherapy exposure, or trauma, detected through medical history and examination [14]. Congenital causes involve chromosomal abnormalities and Y chromosome microdeletions, detected via karyotyping and PCR [14, 15]. While primary testicular failure is usually irreversible, advancements like testicular sperm extraction offer a 50% sperm retrieval rate, enabling ICSI but requiring genetic screening [16].
Cellular transplantation addresses male infertility, especially NOA, in two ways: regenerative medicine enhances germ cell proliferation and differentiation while exploiting transplanted cells’ paracrine and anti-inflammatory effects to treat NOA caused by idiopathic and inflammatory factors.
2. The limitations of current treatment options and the need for the cell-based therapy
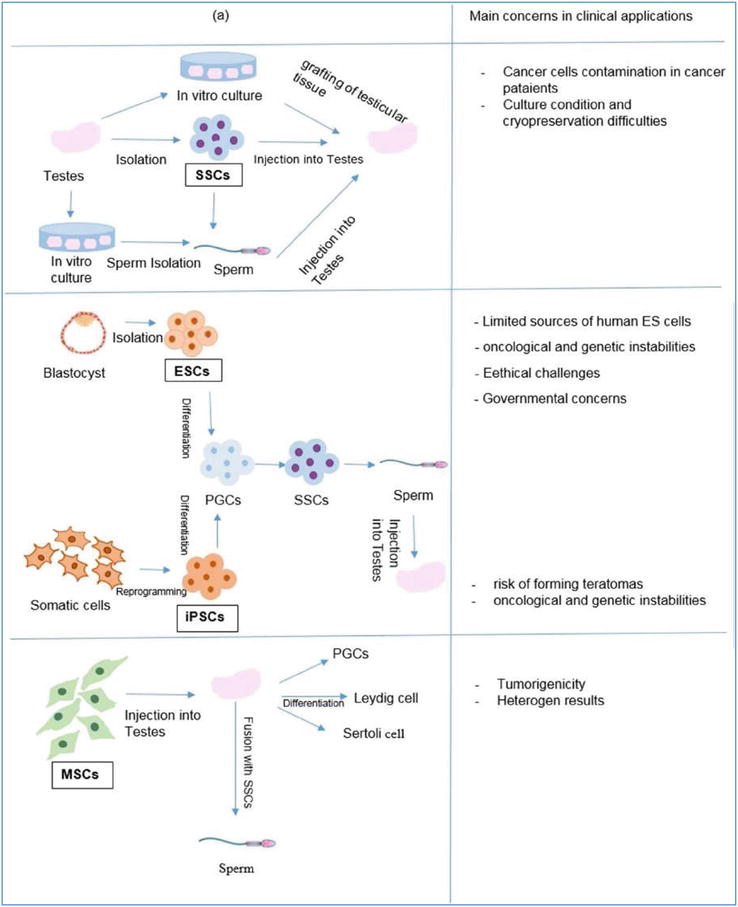
From the perspective of regenerative medicine, there are two experimental approaches aimed at restoring fertility in men with NOA. The first method, referred to as the in vivo approach, involves the transplantation of SSCs into the seminiferous tubules of infertile individuals. These SSCs serve as precursors to mature spermatids. On the other hand, the second method is based on in vitro studies and involves the cultivation and differentiation of various cell types into male germ cells. These cell types include embryonic stem cells [17], iPSCs [18], and MSCs [19, 20].
The upcoming discussion will focus on restoring fertility in men with NOA using the cell sources shown in Figure 1.
Figure 1.
Cell-based therapy for non-obstructive azoospermia. Regenerative medicine strategies are used for cell-based treatments of NOA. (b) Employing (MSCs and testicular somatic cells (DSCs) in NOA cell-based therapy aims to harness their secreted anti-inflammatory and paracrine factors [21].
Stem cells possess the unique ability to self-renew and differentiate into various human tissue cell types. Among these, MSCs have gained prominence in cellular therapy due to their potential. Derived from sources like bone marrow and adipose tissue, MSCs are prized by researchers and clinicians for their versatility, minimal immune reactivity, and active tissue repair capabilities. Compared to other stem cell types, MSCs offer advantages in clinical cell-based therapies, including easy sourcing, immune-suppressive qualities, suitability for autograft and allograft procedures, ethical acceptability, and limited replicative lifespan [22].
Cell and tissue-derived products’ origin is pivotal in regenerative medicine. To boost widespread adoption, we must produce ample cells with consistent quality and therapeutic effectiveness. This ensures steady therapeutic outcomes for patients [23].
This section examines various cell sources for cell therapy in different pathological conditions. We will also explore the transition from cell suspensions to complex tissue-engineered products. Diverse cell sources, including adult materials from living and deceased donors, fetal materials, and pluripotent stem cell lines, have been extensively researched in cell therapy and regenerative medicine [23, 24]. Humans possess a complex multicellular structure with specialized cell types, all originating from a single zygote. As development progresses, cells diversify and lose their ability to transform into other cell types. This ability is referred to as “cell potency” [25, 26] (Table 1).
Totipotency
During the blastocyst phase, a solitary cell possesses the capability to generate all cells until the stage of 16 cells.
Pluripotency
Embryonic stem cells can differentiate into cells that belong to all three germ layers.
Multipotency
The activation of genes imposes restrictions on the ability of these cells to differentiate into multiple cell types, albeit within a limited range. For instance, hematopoietic stem cells possess the capability to differentiate into diverse types of blood cells, including erythrocytes, lymphoid cells, neutrophils, and platelets.
Oligopotency
Certain stem cells possess the ability to differentiate into a restricted range of cell types. To illustrate, lymphoid stem cells can undergo development into either B cells or T cells.
Unipotency
Capability to specialize into a solitary cellular form, such as a precursor cell.
Table 1.
Cellular potency.
Adult cell material, obtained directly from patients, can be purified, or amplified in a lab, for example, mesenchymal stem cells or skin epithelial cells. This self-derived method reduces the risk of rejection but complicates manufacturing and supply logistics for large-scale production [27]. These challenges are more pronounced in diseases like age-related macular degeneration (AMD) or when many cells are needed, like extensive burn treatment. Alternatively, cells can come from deceased or, when possible, living donors (see Figure 2). Using adult cell sources introduce variability due to donor differences in characteristics like histocompatibility, age, and genotype. Additionally, adult cell sources have limited expansion potential, especially for terminally differentiated types (e.g., skeletal muscle cells) or organs with few endogenous stem cells [29].
Figure 2.
A simplified diagram showing a cross-section of a seminiferous tubule and the interstitial space around it is presented. The depicted cell-to-cell communication pathways that play a role in spermatogenesis are also indicated. It is important to note that the germ cells are not depicted as isolated entities; instead, they are connected through intercellular bridges, facilitating the exchange of cytoplasmic contents until later stages after meiosis, when these bridges no longer exist [28].
Potential sources of materials for cell therapy encompass aborted fetuses, which offer highly proliferative cells compared to adult cells. Fetal stem/progenitor cells possess limited differentiation potential but can yield cell types absent in adult tissues. The availability of fetal cells varies globally due to diverse abortion laws. Ethical concerns surround the use of human fetal tissue.
Obtaining fetuses at specific gestational stages poses challenges, as seen in the MIG-HD trial for Huntington’s disease. They attempted grafting fetal ganglionic eminences (GE), containing potential striatal cells, into HD patients’ brains. However, 163 surgeries were canceled out of 86 intended, due to inadequate or substandard fetal material, as fetal cells must be transplanted within 2 days post-abortion [30].
Human pluripotent stem cells hPSCs hold great promise for regenerative medicine due to their remarkable self-renewal and differentiation capabilities. They can be obtained from surplus in vitro fertilized embryos, known as human embryonic stem cells hESCs, or by reprogramming adult cells into a pluripotent state, termed human induced pluripotent stem cell hiPSC generation. Large-scale hiPSC production requires strict adherence to quality control standards and good manufacturing practices (GMP), akin to pharmaceuticals. While hESCs have been predominant in clinical trials, the field is increasingly favoring hiPSCs because they eliminate the need to destroy embryos, allowing wider use [31].
Cell therapy involves introducing healthy cells into diseased tissues, utilizing mature cells or undifferentiated stem cells that can adapt to specific conditions (as shown in Figure 2). Various treatment categories are available for male infertility, including optimizing sperm production, addressing obstructions, and employing surgical sperm retrieval techniques [32]. Men with NOA often require testicular sperm retrieval methods like testicular sperm extraction (TESA), conventional TESA (cTESE), or microsurgical TESA (micro-TESE). Among these techniques, micro-TESE has demonstrated superior success rates, reduced testicular tissue damage, increased sperm yield, and enhanced potential for sperm cryopreservation compared to alternative methods in NOA cases. Despite these advancements, the clinical pregnancy rate remains disappointingly low [33]. However, there is a glimmer of hope for NOA patients who have experienced unsuccessful pregnancies after undergoing micro-TESE surgery, as stem cell therapy emerges as a potential solution [34].
Mesenchymal stem cell (MSC) transplantation is a novel strategy to stimulate spermatogenesis and address male infertility. Sertoli cells (SCs) are crucial for cell survival, proliferation, migration, angiogenesis, and immune modulation. This makes MSCs an excellent choice for azoospermia treatment [35]. MSCs from bone marrow are a primary MSC source and have demonstrated the ability to differentiate into male germ cells in lab settings [36, 37]. Studies have observed spermatogenesis induction and MSC differentiation into germ cells when transplanted into NOA animal models. BM-MSC allotransplantation has successfully treated azoospermia in various animal models, including guinea pigs [38], hamsters [39], mice [40], and rats [41]. Stem cell therapy has significantly advanced NOA management, as seen in trials like NCT02025270, NCT02641769, and NCT02414295, evaluating bone marrow-derived mesenchymal stem cells (BM-MSCs) effects on hormonal profiles, testicular dimensions, and sexual function in NOA cases.
4. The process of spermatogenesis initiates during puberty, wherein spermatogonia under spermatogonial stem cells: a novel approach for treating impaired spermatogenesis
The process of spermatogenesis begins during puberty, when spermatogonia undergo mitotic and meiotic divisions, resulting in the production of haploid spermatids and spermatozoa. Since spermatogenesis relies heavily on stem cells [42], one potential treatment for male infertility caused by impaired spermatogenesis is the transplantation of stem cells (refer to Figure 3). SSCs can restore spermatogenesis in cases where spermatogonial cells have been damaged or depleted [44]. Consequently, stem cell transplantation holds promise as a technique to revive spermatogenesis in cancer patients and individuals experiencing impaired spermatogenesis [45].
Figure 3.
Fertility restoration routes. Following the freezing of a testicular tissue sample, various methods utilizing SSCs can be employed to restore fertility. These methods include SSC transplantation (SSCT), grafting of testicular tissue, and in vitro spermatogenesis, which can be carried out through either cell-based or tissue-based cultivation. With SSCT, there is potential for natural conception, while techniques involving grafting or in vitro spermatogenesis would require an ICSI procedure using elongated spermatids obtained from the respective method [43].
Cancer treatments like radiation and chemotherapy can lead to male infertility in cancer patients. Preserving fertility is a major concern for young boys with cancer. To address this, a procedure called testicular biopsy is performed to collect SSCs, which are frozen before cancer treatment. Later, a stem cell transplant is done in the testes. Researchers are also exploring other stem cell types like MSCs, embryonic stem cells (ESCs), very small embryonic-like stem cells (VSELs), and iPSCs to tackle azoospermia, which can be derived from regular somatic cells [46, 47].
SSCs possess remarkable self-renewal and pluripotent abilities, as highlighted by Seandel et al. [48]. These cells can differentiate into various stem cell types, playing a pivotal role in spermatogenesis and male fertility. However, SSCs are relatively scarce, comprising only 0.03% of germ cells in rodent testes. In contrast, differentiating cells like spermatogonia, spermatocytes, spermatids, and sperm, as observed by Phillips et al. [49], are more abundant. This balance ensures both stem cell preservation and the high sperm production demands of the testes. The emergence of SSCs presents significant potential for addressing human challenges through biotechnological advancements, as extensively explored in biomedical research, as discussed by Park et al. [50].
SSCs originate from gonocytes within the postnatal testis. These gonocytes trace their lineage back to primordial germ cells (PGCs) that emerge during early embryonic development. PGCs, initially identified as alkaline phosphatase-positive cells in the epiblast-stage embryo, appear around 7–7.25 days post-coitum (dpc) in rodents and around 24 dpc in humans, located near the allantois within the yolk sac wall (“post-coitum” is a widely accepted Latin term in biology and reproductive medicine for events after sexual intercourse) [51]. Human PGC presence relies on BMP4 and BMP8b expression within the extraembryonic ectoderm. PGCs passively migrate from the developing embryo, reaching the forming gonadal ridge between 8.5 and 12.5 dpc in mice and 29 and 33 dpc in humans, with approximately 3000 PGCs populating the genital ridges during this process [52]. Subsequently, PGCs differentiate into gonocytes within the male gonads around 13.5 dpc. These gonocytes are situated within testicular cords, composed of precursor Sertoli and peritubular myoid cells. Gonocytes can be categorized as mitotic (M)-prespermatogonia, T1-prospermatogonia, and T2-prospermatogonia. M prespermatogonia, located centrally within testicular cords, progress through various developmental stages during spermatogenesis [53]. Mitotic and meiotic divisions ultimately yield haploid spermatids and spermatozoa (see Figure 2). Given the reliance of spermatogenesis on stem cells, Kanatsu-Shinohara et al. explored stem cell transplantation as a potential solution for male infertility resulting from disrupted spermatogenesis [44], cancer patients undergoing radiation or chemotherapy often face male infertility as a side effect. Preserving fertility is crucial for prepubertal boys undergoing cancer treatment. In such cases, a testicular biopsy is performed to isolate and cryopreserve autologous SSCs before cancer treatment. Subsequently, stem cell transplantation is conducted within the testes, as described by Forbes et al. [46]. Additionally, various other stem cell types, such as MSCs, embryonic stem cells (ESCs), very small embryonic-like stem cells (VSELs), and iPSCs, derived from normal somatic cells, have been explored for treating azoospermia [47].
The presence of PGCs in humans depends on the expression of BMP4 and BMP8b in the extraembryonic ectoderm [44]. Afterward, PGCs are carried away from the developing embryo during allantois development until they migrate through the hindgut, eventually reaching the developing gonadal ridge. In mice, this migration typically occurs between 8.5 and 12.5 dpc, while in humans, it happens at 29–33 dpc. During this migration, approximately 3000 PGCs colonize the genital ridges. These PGCs eventually give rise to gonocytes, which become enclosed in testicular cords. These cords consist of Sertoli precursor cells and peritubular myoid cells and are present in male gonads at approximately 13.5 dpc. Gonocytes can be further classified into three categories: M-prespermatogonia, T1-prospermatogonia, and T2-prospermatogonia. M-prespermatogonia are located in the middle of the testicular cords, away from the basement membrane, and undergo various developmental stages throughout spermatogenesis [52].
6. In vitro proliferation and transplantation of SSCs
The potential restoration of fertility and natural conception in patients through the recolonization of seminiferous tubules and in vivo spermatogenesis can be achieved by auto-transplantation of SSCs collected from the patient before treatment (see Figure 3). The preferred method for reintroducing SSCs into the human testis is the ultrasonically guided injection of cells in the rete testis, as discussed comprehensively by Gul et al. [54].
Brinster and colleagues have successfully illustrated the establishment of colonization and spermatogenesis within the testes of recipient mice, resulting in offspring possessing the donor haplotype [55]. Additionally, Takashima and Shinohara [56] have reviewed studies demonstrating the accomplishment of SSC auto- or allo-transplantation in diverse mammalian species, encompassing rodents, non-rodents, and non-human primates [57, 58], which has subsequently led to functional spermatogenesis in the recipient organisms.
In human studies, clinical trials involving SSC transplantation have not yet been established. A single report on seven men receiving injections of cryopreserved testicular cells lacks a follow-up report on the outcome of the procedures [59]. However, studies in mice have shown that the number of transplanted SSC colonies gradually decreases during the homing process after transplantation [60], and the success of colonization and donor-derived spermatogenesis within the recipient testis is highly dependent on the concentration of transplanted SSCs [61, 62].
To address limited tissue samples from patients, promoting the expansion of SSCs (spermatogonial stem cells) becomes crucial. This expansion is necessary to generate enough for successful transplantation and re-establishment of seminiferous tubules in the recipient’s testicular tissue. Kanatsu-Shinohara and colleagues demonstrated sustained, long-term murine SSC expansion, leading to fertility restoration in sterile recipients [63]. Similarly, long-term propagation of testicular cells has been achieved from both adult sources [64] and pre-pubertal tissues [65]. Human SSCs have been confirmed in humans and validated through xenotransplantation into sterile mice, where they successfully migrated to the niche within seminiferous tubules housing SSCs.
Nevertheless, the task of pinpointing SSCs in testicular tissue or in vitro cell cultures remains a formidable challenge. In testicular cell cultures, a mixture of somatic cells and germ cells coexists, with the latter constituting a diverse population of spermatogonia at various stages of differentiation. At present, there is no universally accepted marker that definitively distinguishes human and/or non-human primate SSCs, whether within living organisms or in laboratory cultures. Many frequently employed markers, such as ITGA6, KIT, GPR125, and DAZL, are not exclusive to spermatogonia and are also present in somatic cells within the testicular environment [66, 67]. The expression of other markers, like GFRA1, THY1, and UCHL1, in spermatogonia is also controversial [68, 69]. Moreover, the presence of specific indicators is contingent upon the developmental phase of the testicular tissue [70].
Single-cell sequencing studies have identified different developmental states of germ cells, ranging from State 0 to State 4, each characterized by a unique set of markers, although there may be some overlap [71]. Examples of markers expressed in the earliest state include UTF1 and PIWIL4, with PIWIL4 being more specific [71, 72]. Another potential marker for undifferentiated spermatogonia with SSC characteristics is the LPPR3 protein [73].
Furthermore, it is uncertain whether various types of testicular cells maintain their transcriptomic and metabolic signatures when isolated from their natural niche [67]. The diversity found within testicular cell populations, coupled with the abundance of potential markers at our disposal, poses a challenge when it comes to comparing the outcomes and efficiency of culturing SSCs in different research studies. To advance our comprehension and practical use of SSCs, it is imperative to undertake further investigations geared toward pinpointing specific markers applicable to precisely defined spermatogonial subpopulations. Additionally, functional studies are necessary to assess the in vitro SSC potential of these identified markers. When seeking out these markers, it is advisable to prioritize surface markers over nuclear markers, as this approach is likely to be more advantageous for the isolation and enrichment of SSCs from cryopreserved biopsies or post-culture scenarios [43].
Cultural methods and media should support SSC proliferation while preventing their differentiation. In addition to basic nutrients, the culture medium may include specific cytokines, metabolites, hormones, and signaling molecules known to stimulate spermatogonial proliferation. Kanatsu-Shinohara et al. successfully developed a culture protocol in 2003 using StemPro-34 Serum-Free medium for long-term in vitro propagation of murine SSCs [63]. Although a similar medium can support long-term propagation of human SSCs [64, 65], the overgrowth of testicular somatic cells in the culture system remains a challenge, as it dilutes the SSC signature over time [63]. Feeder cells are beneficial for the survival of SSCs as they provide mechanical and metabolic support as well as paracrine signals [74, 75]. However, the utilization of external feeder layers is not conducive to clinical implementation. Consequently, somatic cells naturally present in the testicular suspension and facilitating SSC proliferation are employed for SSC cultivation. To prevent excessive proliferation of these somatic cells, the cultivation technique and medium must achieve an optimal equilibrium between the growth of somatic cells and SSCs. Alternatively, testicular somatic cells can be substituted with a synthetic matrix for cellular support, coupled with the supplementation of the medium with nutrients and growth factors to provide the metabolic sustenance typically offered by feeder cells for SSCs. Nonetheless, this process heavily relies on the ability to isolate and separate the somatic cell population from the germ cell population prior to cultivation, a task currently constrained by the absence of a specific SSC marker [76].
Hence, it is imperative to refine the in vitro cultivation technique for human SSCs before embarking on clinical applications. Investigating the SSC microenvironment in humans could prove pivotal in the discovery of essential factors essential for the efficient preservation and proliferation of SSCs [77].
Spermatogenesis encompasses a series of physiological, morphological, and biochemical changes that lead to the development of mature sperm cells. However, this process can be disrupted by various factors such as congenital or genetic abnormalities, as well as physical, chemical, and environmental factors. These disruptions can contribute to temporary or permanent infertility [78, 79, 80]. According to the latest report from the World Health Organization (WHO), a condition known as azoospermia, which refers to the absence of spermatozoa in the ejaculate, affects approximately 1% of the male population and 10–20% of infertile men [81, 82]. Besides genetic and congenital factors, chemotherapeutic drugs have the potential to substantially disrupt the typical processes of spermatogenesis and sperm characteristics. This disruption can result in the cessation of progenitor cell differentiation and a reduction in the germ cell reservoir [83].
8. Busulfan destroys DNA structure, prevents proliferation and differentiation of SSCs, and initiates apoptosis
Various animal models have been described to induce azoospermia, including busulfan injection [84], testicular heat stress [85], testicular torsion [86], radiation [87], and cryptorchidism induction [88]. Among these, busulfan injection and hyperthermia exposure are two commonly used methods to deplete germ cells from the testes [84, 85].
Busulfan, scientifically referred to as 1,4-butanediol dimethanesulfonate, is commonly utilized for the management of myeloproliferative syndromes, chronic myeloid leukemia (CML), lymphomas, and ovarian cancer [89]. This chemotherapeutic medication decreases the rate at which cells multiply by specifically intervening during the G1 phase of their growth cycle. It accomplishes this by forming connections between DNA proteins or DNA strands, effectively halting cell division during the mitosis/replication phase and instigating apoptosis [84]. Busulfan is also administered to leukemia patients prior to bone marrow transplantation, often in combination with cyclophosphamide and clofarabine, as a myelosuppressive/myeloablative drug [90, 91, 92]. However, it is important to note that both short-term and long-term side effects have been reported on various vital organs, including the urinary bladder, liver, skin, gonads, and nervous system [93, 94]. Impaired spermatogenesis has been observed in cancer patients receiving busulfan treatment [95]. The primary objective of this study is to examine the adverse impacts of busulfan on various rat organs, such as the liver, kidneys, testes, and bone marrow, utilizing histological, biochemical, and cytological assessments [41, 89].
The application of Busulfan treatment is intended to reduce germ cells for the purpose of transplantation research. Nevertheless, this method is linked to an inadequate elimination of germ cells, and the challenge of depleting endogenous spermatogenesis is greater in rats when compared to mice employed as recipients [96]. Various research groups have employed different doses (ranging from 10 to 50 mg/kg) of busulfan to eliminate testis function for their experiments, and different mouse strains exhibit varying sensitivities to busulfan doses. Recently, [97] in WBB6F1 WT mice, endogenous resumption was observed at 22 mg/kg busulfan dose, contrasting with the absence of resumption at 44 mg/kg. Our investigation in Swiss mice showed germ cell aplasia at or above 25 mg/kg, with doses exceeding 25 mg/kg causing premature mortality, hindering long-term safety assessment. Therefore, we consistently use a 25 mg/kg dose, achieving superior spermatogenesis restoration after niche cell transplantation compared to vehicle transplants post-chemotherapy. In humans, azoospermia in cancer survivors varies based on specific oncological drugs. A recent study using seven testicular biopsies from adult survivors of childhood cancers found a complete absence of germ cells, with only Sertoli cells and VSELs surviving [98].
9. Effects of heat stress on spermatogenesis and germ cells
Maintaining the temperature of the testicles within an optimal range is crucial for the process of spermatogenesis. Even slight increases in scrotal temperature, even within the normal physiological range, have a detrimental impact on sperm quality [99]. A rise of 1°C results in a 14% reduction in spermatogenesis (see Figure 4), leading to decreased sperm production [100]. The influence of temperature on male fertility is evident, as infertile men have higher mean scrotal temperatures compared to fertile men, and further increases in scrotal temperature result in a decline in sperm quality [101].
Figure 4.
The pathways involved in apoptosis: intrinsic and extrinsic. The intrinsic pathway centers on mitochondrial regulation, where Bax accumulates in response to heat stress, leading to cytochrome C release and caspase activation. The extrinsic pathway, initiated by Fas-FasL binding, activates caspases via the Fas/FADD complex. In hyperthermia, p53 relocates to the nucleus, inducing cell cycle arrest and apoptosis, while also influencing the intrinsic pathway by regulating Bax and Bcl-2. Both pathways converge at the executioner caspase cascade, resulting in germ cell death [85].
Spermatogenesis, specifically the process of spermatocyte and spermatid differentiation and maturation, is profoundly influenced by temperature. The ideal conditions for spermatogenesis require a temperature that is at least 2°C lower than the core temperature of the body [102]. However, elevated scrotal temperature leads to testicular germinal atrophy, spermatogenic arrest [103], and reduced levels of inhibin B (a biochemical marker of spermatogenesis) [104], resulting in lower sperm counts [105].
The susceptibility of germ cells to heat stress is higher due to their elevated mitotic activity [106]. Within the germ cell population, pachytene and diplotene spermatocytes, as well as early round spermatids, are particularly susceptible to heat stress in both humans [107] and rats [102, 108]. Several fundamental mechanisms contribute to germ cell damage, including germ cell apoptosis [108], autophagy [109], DNA damage caused by altered synapsis and strand breaks, and the generation of reactive oxygen species. The subsequent section, “Molecular response of male germ cells to heat stress,” will provide a more detailed explanation of the molecular reactions observed in germ cells under hyperthermic conditions [85].
10. Cell-based therapies for NOA via paracrine and anti-inflammatory pathways
Scientific evidence suggests that immune-related factors and inflammatory processes could be responsible for testicular damage and male infertility in approximately 30% of asymptomatic infertile patients [110]. Research indicates that immune cell infiltration has been observed in at least 20% of testicular biopsies taken from infertile patients with azoospermia, indicating a significant contribution of inflammatory infertility to male infertility [111].
Prior research has likewise illustrated the existence of immune cell infiltration and simultaneous inflammatory ailments in testicular biopsies from all dogs afflicted with NOA, encompassing M1 pro-inflammatory phenotype macrophages, pro-inflammatory monocytes, and cytokines [112].
In previous research, biopsies collected from males diagnosed with NOA revealed the presence of inflammatory lesions, characterized by the presence of lymphocytes and monocytes/macrophages, which were linked to compromised spermatogenesis. In contrast, samples taken from individuals with obstructive azoospermia (OA) exhibited uninhibited spermatogenesis without any signs of inflammation [113]. These investigations also revealed that inflammatory responses have the potential to inflict harm upon the testes and epididymis. Elevated concentrations of inflammatory mediators like tumor necrosis factor (TNF) and Activin A were detected in human testicular biopsies displaying compromised spermatogenesis, as observed in these studies [114]. Additionally, NOA has been observed in 10% of men with acute epididymitis. As mentioned earlier, (MSCs have been found to possess immunomodulatory, anti-inflammatory, anti-apoptotic, and proliferative effects through the secretion of cytokines and growth factors [115, 116].
Accumulative evidence suggests that the components of the spermatogonial stem cell niche play a significant role in the development of idiopathic male infertility. Infertility affects approximately 8–12% of couples worldwide, with male factors contributing to about 50% of cases. Idiopathic male infertility, accounting for 30–50% of male infertility cases, lacks identifiable causes [117]. Despite normal physical examinations and laboratory tests, semen analysis often reveals sperm abnormalities, either isolated or in combination [118].
Epigenetic elements, specifically alterations in DNA methylation at a global or gene-specific level, could potentially play a role in the onset of idiopathic male infertility. However, the involvement of histone modifications and chromatin protamination in spermatogenesis remains somewhat uncertain [119]. Specific configurations of differential DNA methylation regions may function as prognostic indicators for the efficacy of particular treatments in addressing idiopathic male infertility [120]. Nonetheless, there is a need for more comprehensive research to elucidate the mechanisms responsible for changes in DNA methylation patterns and their significance in the context of male infertility. Likewise, although extensive investigations have been carried out on other epigenetic elements like miRNAs, additional data are essential to establish a definitive link between abnormal miRNA expression and unexplained male infertility [121].
Abnormal mRNA expression could also be correlated with unexplained male infertility. Research has indicated that numerous genes associated with unexplained male infertility are connected to reactive oxygen species (ROS). This might elucidate the observed disproportion in ROS genes and their protein products in seminal plasma and impaired spermatozoa [122]. The expression levels of glutathione transferase genes were discovered to be elevated in samples obtained from individuals afflicted with NOA and oligospermia, which may contribute to the detoxification of reactive oxygen species (ROS) [123]. Aberrant expression is not limited to cells in the spermatogenic epithelium but also extends to somatic cells [124]. Transcriptome analysis of single Sertoli cells obtained from patients with idiopathic male infertility revealed the presence of immature Sertoli cell fractions. These cells exhibited characteristics resembling infantile and pubertal Sertoli cells, displayed increased in vitro proliferation, and demonstrated energy metabolism patterns typical of immature Sertoli cells. Although their ability to support germ cell colonies was slightly impaired compared to normal adult Sertoli cells, this functional immaturity could be reversed through inhibition of the Wnt pathway, suggesting potential therapeutic modulation of SSC niche properties [125]. Empirical investigations have revealed noteworthy indications of compromised Leydig cell performance in males with a record of unexplained male infertility. This could be attributed to potential disturbances in the paracrine communication between the seminiferous epithelium and Leydig cells, or congenital dysfunction affecting both of these essential components [126]. Furthermore, within a mouse model, it was observed that the generation of glial cell line-derived neurotrophic factor GDNF by peritubular myoid cells plays a crucial role in spermatogonial growth. This underscores the significance of the interaction among key components of the spermatogonial stem cell niche, including Leydig cells and peritubular myoid cells, given that testosterone stimulates the expression of GDNF [118].
11. Challenges and future directions
11.1 The importance of optimizing cell isolation, transplantation techniques, and immunosuppression protocols
NOA, a condition characterized by low or absent sperm production, poses a significant challenge in male infertility. However, advancements in gene editing and combination therapies hold promise for its future treatment. Possible directions include:
Gene editing to correct genetic defects: CRISPR-Cas9, a gene editing tool, could correct specific genetic defects linked to NOA, potentially restoring normal sperm production [127].
Induced pluripotent stem cell therapy: iPSCs, reprogrammed adult cells, can transform into different cell types, including sperm cells. Scientists are investigating iPSCs for generating functional sperm in the lab, potentially offering a sperm source for individuals with NOA [128].
Testicular sperm extraction (TESE) combined with assisted reproductive techniques: TESE, which surgically retrieves sperm from the testicles, can be coupled with assisted reproductive methods like ICSI to facilitate egg fertilization and embryo transfer. This approach exhibits potential for successful pregnancies in certain instances of NOA [129].
Hormonal therapies: Hormonal treatments, including FSH and hCG, can boost sperm production in those with NOA, though their effectiveness varies by case [130].
Combination therapies: Combining various treatments can improve outcomes for NOA by addressing underlying causes and boosting sperm production. This may involve gene editing, hormonal therapies, and assisted reproductive techniques [131].
11.2 Ethical considerations and regulatory framework
Respect for human life: A central ethical concern pertains to the moral status of embryos, particularly in the context of embryonic stem cell research. Extracting these cells often entails destroying human embryos, prompting debates on whether this constitutes the loss of potential human life. This raises ethical questions about the treatment and utilization of human embryos in scientific or therapeutic endeavors [132].
Informed consent: Ethical concerns in cell-based therapies encompass securing informed consent, especially for embryonic stem cell use and genetic modifications. These techniques bear significant implications for individuals and their offspring. Clear communication of potential risks and benefits empowers informed decision-making by participants [133].
Equity and access: The ethical concerns surrounding cell-based therapies encompass equity and accessibility. These treatments are intricate and expensive, posing barriers for some. Ensuring fair access is vital to prevent worsening health disparities and promote healthcare justice.
Genetic modifications and enhancement: Genetic modifications raise ethical questions regarding the differentiation between therapeutic interventions and enhancements. They can address genetic disorders or enhance traits, leading to ongoing ethical debates and the need to define ethical boundaries [134].
Long-term consequences and unintended effects: Long-term implications and unintended outcomes of cell-based therapies, including genetic modifications, require thorough evaluation. Ethical responsibility necessitates rigorous research, safety assessments, and continuous monitoring to mitigate potential harm and unforeseen consequences for future generations.
Regulatory oversight: To address the complexity and risks of cell-based therapies, establishing effective regulatory frameworks is vital. Ethical concerns require the creation of strong regulations and oversight systems to ensure the ethical and responsible application of these technologies. Regulatory authorities must aim for a balance between innovation, patient safety, and public confidence to prevent unethical conduct and safeguard individuals’ well-being [135].
11.3 The existing regulatory framework and guidelines for the clinical application of cell-based therapies in reproductive medicine
As of September 2021, I can discuss the regulatory framework and guidelines for using cell-based therapies in reproductive medicine. Keep in mind that these rules may change, so consult current sources and regulatory bodies for updates.
Many countries have government agencies that oversee cell-based therapies in reproductive medicine to ensure safety, efficacy, and ethical use. Here are key points about these regulations and guidelines:
Regulatory authorities: Various countries have specific regulatory agencies overseeing cell-based therapies. In the United States, the FDA regulates these therapies, and in the European Union, the EMA is responsible for oversight [136].
Classification of cell therapies: Regulatory bodies classify cell-based therapies based on intended use, risk, and manufacturing methods. This classification dictates oversight and regulatory routes. Categories, like advanced therapy medicinal products (ATMPs) or biological products, vary by country but remain consistent in the general framework [137].
Preclinical testing: Cell-based therapies undergo preclinical evaluation to assess safety and efficacy before clinical trials. This involves lab research and animal testing to gather evidence on benefits and risks.
Clinical trials: Clinical trials assess cell-based therapy safety and efficacy in humans. Regulatory bodies mandate researchers to secure approval and adhere to precise trial guidelines, encompassing participant selection, informed consent, treatment administration, and data gathering [138].
Good manufacturing practices (GMP): Cell-based therapies used in reproductive medicine are often subject to stringent manufacturing requirements to ensure their quality, consistency, and safety. GMP guidelines regulate various aspects of manufacturing, including facilities, personnel, equipment, documentation, and quality control.
Ethical considerations: Cell-based therapies in reproductive medicine may raise ethical concerns related to the source of cells, their manipulation, and the implications of their use. Regulatory frameworks often include guidelines or principles to address these ethical considerations, such as informed consent, privacy protection, and appropriate handling of human tissues [139].
12. Future perspectives
Infertility, a multifaceted condition encompassing genetic, environmental, physical, and psychological factors alongside issues in germ cell production and transmission, persists as a challenge. Assisted reproductive technology ART, notably IVF, has facilitated numerous births but raises concerns about patient and offspring well-being. This review delves into refractory infertility, its traits, and treatment approaches, with an emphasis on stem cell potential in treatment.
Stem cell therapies, due to their pluripotent capabilities, emerge as ART alternatives. Research explores their utility in enhancing infertility treatment. The review covers translational studies investigating stem cell-based treatments’ mechanisms: direct differentiation and cytokine secretion, customized for specific conditions. For age-related or irreversible fertility problems, germ-line stem cells have induced active gamete production, and mesenchymal stem cells contribute to immunoregulation and organ restoration.
Notably, stem cell therapies are presently in preclinical stages, prompting ethical concerns and diverse opinions. To transition into clinical practice, meticulous, long-term planning, thorough assessments, and vigilant oversight are essential to ensure precision, excellence, and safety. Autologous stem cells, sourced from the patient, show promise for future clinical use due to their safety and immunogenicity benefits.
13. Conclusion
Cell-based therapy shows promise in addressing NOA, a condition marked by the absence of sperm in ejaculation due to testicular dysfunction. Traditional treatments have limitations, prompting exploration of cell-based approaches using SSCs or iPSCs to restore spermatogenesis. Animal studies demonstrate successful fertility restoration via SSC transplantation, while human research advances in isolating and characterizing human SSCs. Challenges persist in scaling human SSCs and optimizing iPSC differentiation. Further research is needed to overcome technical hurdles and ensure safety and sustained functionality of transplanted cells, potentially providing a novel NOA treatment option and fertility restoration.
References
1.Matzuk MM, Lamb DJ. The biology of infertility: Research advances and clinical challenges. Nature Medicine. 2008;14(11):1197-1213
2.Uhlén M et al. Tissue-based map of the human proteome. Science. 2015;347(6220):1260419
3.Zhang X et al. Embryo development and live birth resulted from artificial oocyte activation after microdissection testicular sperm extraction with ICSI in patients with non-obstructive azoospermia. Frontiers in Endocrinology. 2023;14:1123541
4.Logan S, Anazodo A. The psychological importance of fertility preservation counseling and support for cancer patients. Acta Obstetricia et Gynecologica Scandinavica. 2019;98(5):583-597
5.Kherraf Z-E et al. Whole-exome sequencing improves the diagnosis and care of men with non-obstructive azoospermia. The American Journal of Human Genetics. 2022;109(3):508-517
6.Tournaye H, Krausz C, Oates RD. Novel concepts in the aetiology of male reproductive impairment. The Lancet Diabetes & Endocrinology. 2017;5(7):544-553
7.Zhankina R et al. Mesenchymal stromal/stem cells and their exosomes for restoration of spermatogenesis in non-obstructive azoospermia: A systemic review. Stem Cell Research & Therapy. 2021;12(1):229
8.Wosnitzer M, Goldstein M, Hardy MP. Review of Azoospermia. Spermatogenesis. 2014;4(1):e28218
9.Oud MS et al. Validation and application of a novel integrated genetic screening method to a cohort of 1,112 men with idiopathic azoospermia or severe oligozoospermia. Human Mutation. 2017;38(11):1592-1605
10.Gordetsky J, van Wijngaarden E, O'Brien J. Redefining abnormal follicle-stimulating hormone in the male infertility population. BJU International. 2012;110(4):568-572
11.Fernandez CJ, Chacko EC, Pappachan JM. Male obesity-related secondary hypogonadism—Pathophysiology, clinical implications and management. European Endocrinology. 2019;15(2):83-90
12.Hammond GL. Sex hormone-binding globulin and the metabolic syndrome. In: Male Hypogonadism: Basic, Clinical and Therapeutic Principles. Cham: Springer International Publishing, Humana Press; 2017. pp. 305-324
13.Hwang K et al. Evaluation of the azoospermic male: A committee opinion. Fertility and Sterility. 2018;109(5):777-782
14.Peña VN, Kohn TP, Herati AS. Genetic mutations contributing to non-obstructive azoospermia. Best Practice & Research Clinical Endocrinology & Metabolism. 2020;34(6):101479
15.Sharlip ID et al. Best practice policies for male infertility. Fertility and Sterility. 2002;77(5):873-882
16.Corona G et al. Sperm recovery and ICSI outcomes in men with non-obstructive azoospermia: A systematic review and meta-analysis. Human Reproduction Update. 2019;25(6):733-757
17.Bucay N et al. A novel approach for the derivation of putative primordial germ cells and sertoli cells from human embryonic stem cells. Stem Cells. 2009;27(1):68-77
18.Yang W et al. iPSC reprogramming from human peripheral blood using Sendai virus mediated gene transfer. International Journal of Biomedical and Health Sciences. 2021;10(4)
19.Nayernia K et al. In vitro-differentiated embryonic stem cells give rise to male gametes that can generate offspring mice. Developmental Cell. 2006;11(1):125-132
20.El-Sayed ME et al. Mesenchymal stem cell transplantation in burn wound healing: Uncovering the mechanisms of local regeneration and tissue repair. Histochemistry and Cell Biology. 2023
21.Roshandel E et al. Cell-based therapy approaches in treatment of non-obstructive azoospermia. Reproductive Sciences. 2023;30(5):1482-1494
22.Kim HJ, Park JS. Usage of human mesenchymal stem cells in cell-based therapy: Advantages and disadvantages. Development & Reproduction. 2017;21(1):1-10
23.Jarrige M et al. The future of regenerative medicine: Cell therapy using pluripotent stem cells and acellular therapies based on extracellular vesicles. Cells. 2021;10. DOI: 10.3390/cells10020240
24.Atwa A et al. Biodegradable materials from natural origin for tissue engineering and stem cells technologies. In: Ali GAM, Makhlouf ASH, editors. Handbook of Biodegradable Materials. Cham: Springer International Publishing; 2022. pp. 1-40
25.Sampogna G, Guraya SY, Forgione A. Regenerative medicine: Historical roots and potential strategies in modern medicine. Journal of Microscopy and Ultrastructure. 2015;3(3):101-107
26.Ashry M et al. Hormonal and inflammatory modulatory effects of hesperidin in hyperthyroidism-modeled rats. Frontiers in Immunology. 2023;14:1087397. DOI: 10.3389/fimmu.2023.1087397
27.Ratcliffe E, Thomas RJ, Williams D. Current understanding and challenges in bioprocessing of stem cell-based therapies for regenerative medicine. British Medical Bulletin. 2011;100(1):137
28.Robinson M et al. Human in vitro spermatogenesis as a regenerative therapy — Where do we stand? Nature Reviews Urology. 2023
29.Chowdhury S et al. Stem cells an overview. Stem Cells: Biology and Therapeutics. 2021:1-21
30.Kawasaki H et al. Current status and legal/ethical problems in the research use of the tissues of aborted human fetuses in Japan. Congenital Anomalies. 2020;60(6):166-174
31.Ozawa H, Matsumoto T, Nakagawa M. Culturing human pluripotent stem cells for regenerative medicine. Expert Opinion on Biological Therapy. 2023:1-11
32.Cullen I, Muneer A. Surgical sperm retrieval and MicroTESE. In: Allahbadia GN et al., editors. Textbook of Assisted Reproduction. Singapore: Springer Singapore; 2020. pp. 193-202
33.Kang C, Punjani N, Schlegel PN. Reproductive chances of men with azoospermia due to spermatogenic dysfunction. Journal of Clinical Medicine. 2021;10:1400. DOI: 10.3390/jcm10071400
34.Deng C-C, Liu G-H. Stem cell therapy for non-obstructive azoospermia. Zhonghua nan ke xue = National Journal of Andrology. 2020;26(4):351-356
35.Tamadon A et al. Mesenchymal stem cell therapy of male infertility. In: Male Reproductive Health. 2019. p. 105
36.Yan G et al. Ectopic expression of DAZL gene in goat bone marrow-derived mesenchymal stem cells enhances the trans-differentiation to putative germ cells compared to the exogenous treatment of retinoic acid or bone morphogenetic protein 4 signalling molecules. Cell Biology International. 2015;39(1):74-83
37.Salem M et al. Germ cell differentiation of bone marrow mesenchymal stem cells. Andrologia. 2019;51(4):e13229
38.Hajihoseini M et al. Induction of spermatogenesis after stem cell therapy of azoospermic Guinea pigs. Veterinary Archives. 2017;87(3):333-350
39.Tamadon A et al. Induction of spermatogenesis by bone marrow-derived mesenchymal stem cells in busulfan-induced azoospermia in hamster. International Journal of Stem Cells. 2015;8(2):134-145
40.Kadam P et al. Co-transplantation of mesenchymal stem cells improves spermatogonial stem cell transplantation efficiency in mice. Stem Cell Research & Therapy. 2018;9(1):317
41.Atwa A, Mansour AM, Bakry S. Mesenchymal stem cells restore fertility in induced azoospermic rats following chemotherapy administration. Journal of Reproduction and Infertility. 2014;5(2):52-60
42.Schuppe HC et al. Chronic orchitis: A neglected cause of male infertility? Andrologia. 2008;40(2):84-91
43.Sanou I et al. Spermatogonial stem cell-based therapies: Taking preclinical research to the next level. Frontiers in Endocrinology. 2022;13
44.Kanatsu-Shinohara M et al. Functional assessment of self-renewal activity of male germline stem cells following cytotoxic damage and serial transplantation1. Biology of Reproduction. 2003;68(5):1801-1807
45.Valli H, Gassei K, Orwig KE. Stem cell therapies for male infertility: Where are we now and where are we going? Biennial Review of Infertility. 2015;4:17-39
46.Forbes CM, Flannigan R, Schlegel PN. Spermatogonial stem cell transplantation and male infertility: Current status and future directions. Arab Journal of Urology. 2018;16(1):171-180
47.Pourmoghadam Z et al. Current approaches for the treatment of male infertility with stem cell therapy. Journal of Cellular Physiology. 2018;233(10):6455-6469
48.Seandel M et al. Generation of functional multipotent adult stem cells from GPR125+ germline progenitors. Nature Medicine. 2007;449(7160):346-350
49.Phillips BT, Gassei K, Orwig KE. Spermatogonial stem cell regulation and spermatogenesis. Philosophical Transactions of the Royal Society B: Biological Sciences. 2010;365(1546):1663-1678
50.Park JE et al. Development of a three-dimensional hydrogel system for the maintenance of porcine spermatogonial stem cell self-renewal. Jet. 2017;32(4):343-351
51.Freeman B. The active migration of germ cells in the embryos of mice and men is a myth. Reproduction-Cambridge. 2003;125(5):635-643
52.Møllgård K et al. Human primordial germ cells migrate along nerve fibers and Schwann cells from the dorsal hind gut mesentery to the gonadal ridge. Molecular Human Reproduction. 2010;16(9):621-631
53.Cremades N et al. In-vitro maturation of round spermatids using co-culture on Vero cells. Human Reproduction. 1999;14(5):1287-1293
54.Gul M et al. Review of injection techniques for spermatogonial stem cell transplantation. Human Reproduction Update. 2020;26(3):368-391
55.Brinster RL, Zimmermann JW. Spermatogenesis following male germ-cell transplantation. Proceedings of the National Academy of Sciences. 1994;91(24):11298-11302
56.Takashima S, Shinohara T. Culture and transplantation of spermatogonial stem cells. Stem Cell Research. 2018;29:46-55
57.Hermann BP et al. Spermatogonial stem cell transplantation into rhesus testes regenerates spermatogenesis producing functional sperm. Cell Stem Cell. 2012;11(5):715-726
58.Shetty G et al. Postpubertal spermatogonial stem cell transplantation restores functional sperm production in rhesus monkeys irradiated before and after puberty. Andrology. 2021;9(5):1603-1616
59.Radford JA. Is prevention of sterility possible in men? Annals of Oncology. 2000;11(Suppl 3):173-174
60.Nagano MC. Homing efficiency and proliferation kinetics of male germ line stem cells following transplantation in mice. Biology of Reproduction. 2003;69(2):701-707
61.Mirzapour T et al. Xenotransplantation assessment: Morphometric study of human spermatogonial stem cells in recipient mouse testes. Andrologia. 2015;47(6):626-633
62.Dobrinski I et al. Computer assisted image analysis to assess colonization of recipient seminiferous tubules by spermatogonial stem cells from transgenic donor mice. Molecular Reproduction and Development. 1999;53(2):142-148
63.Kanatsu-Shinohara M et al. Long-term proliferation in culture and germline transmission of mouse male germline stem cells. Biology of Reproduction. 2003;69(2):612-616
64.Sadri-Ardekani H et al. Propagation of human spermatogonial stem cells In vitro. JAMA. 2009;302(19):2127-2134
65.Sadri-Ardekani H et al. In vitro propagation of human prepubertal spermatogonial stem cells. JAMA. 2011;305(23):2416-2418
66.Kossack N et al. A combined approach facilitates the reliable detection of human spermatogonia in vitro. Human Reproduction. 2013;28(11):3012-3025
67.Struijk RB et al. ITGA6+ human testicular cell populations acquire a mesenchymal rather than germ cell transcriptional signature during long-term culture. International Journal of Molecular Sciences. 2020:21. DOI: 10.3390/ijms21218269
68.Baert Y et al. Orthotopic grafting of cryopreserved prepubertal testicular tissue: In search of a simple yet effective cryopreservation protocol. Fertility and Sterility. 2012;97(5):1152-1157
69.McCoy DE et al. Structural absorption by barbule microstructures of super black bird of paradise feathers. Nature Communications. 2018;9(1):1
70.Guo F et al. The transcriptome and DNA methylome landscapes of human primordial germ cells. Cell. 2015;161(6):1437-1452
71.Sara Di P et al. EGR4-dependent decrease of UTF1 is associated with failure to reserve spermatogonial stem cells in infertile men. bioRxiv. 2021:2021.02.02.429371
72.Di Persio S et al. Spermatogonial kinetics in humans. Development. 2017;144(19):3430-3439
73.Sohni A et al. The neonatal and adult human testis defined at the single-cell level. Cell Reports. 2019;26(6):1501-1517.e4
74.Jetten AM, Kang HS, Takeda Y. Retinoic acid-related orphan receptors α and γ: Key regulators of lipid/glucose metabolism, inflammation, and insulin sensitivity. Frontiers in Endocrinology. 2013;4:1
75.Voigt AL et al. Metabolic requirements for spermatogonial stem cell establishment and maintenance In vivo and In vitro. International Journal of Molecular Sciences. 2021;22. DOI: 10.3390/ijms22041998
76.Murdock MH et al. Human testis extracellular matrix enhances human spermatogonial stem cell survival In vitro. Tissue Engineering Part A. 2018;25(7-8):663-676
77.Kurek M et al. Spermatogonia loss correlates with LAMA 1 expression in human prepubertal testes stored for fertility preservation. Cells. 2021;10(2)
78.de Michele F et al. Preserved seminiferous tubule integrity with spermatogonial survival and induction of Sertoli and Leydig cell maturation after long-term organotypic culture of prepubertal human testicular tissue. Human Reproduction. 2017;32(1):32-45
79.Nayernia K et al. Derivation of male germ cells from bone marrow stem cells. Laboratory Investigation. 2006;86(7):654-663
80.Aboul Fotouh GI et al. Histological study on the protective effect of endogenous stem cell mobilization in busulfan-induced testicular injury in albino rats. Journal of Microscopy and Ultrastructure. 2018;6(4):197-204
81.Alfano M et al. Anti-Mullerian hormone-to-testosterone ratio is predictive of positive sperm retrieval in men with idiopathic non-obstructive azoospermia. Scientific Reports. 2017;7(1):17638
82.Menkveld R. Clinical significance of the low normal sperm morphology value as proposed in the fifth edition of the WHO Laboratory manual for the examination and processing of human semen. Asian Journal of Andrology. 2010;12(1):47-58
83.Gandini L et al. Effect of chemo- or radiotherapy on sperm parameters of testicular cancer patients. Human Reproduction. 2006;21(11):2882-2889
84.Chen X, Liang M, Wang D. Progress on the study of the mechanism of busulfan cytotoxicity. Cytotechnology. 2018;70:497-502
85.Durairajanayagam D, Agarwal A, Ong C. Causes, effects and molecular mechanisms of testicular heat stress. Reproductive Biomedicine Online. 2015;30(1):14-27
86.Azizollahi S et al. Recruiting testicular torsion introduces an azoospermic mouse model for spermatogonial stem cell transplantation. Urology Journal. 2014;11(3):1648-1655
87.Abuelhija M et al. Rat models of post-irradiation recovery of spermatogenesis: Interstrain differences. Andrology. 2013;1(2):206-215
88.Absalan F, Movahedin M, Mowla SJ. Evaluation of apoptotic genes expression and its protein after treatment of cryptorchid mice. Iranian Biomedical Journal. 2012;16(2):77-83
89.Mobarak H et al. Intratesticular versus intraperitoneal injection of busulfan for the induction of azoospermia in a rat model. BMC Pharmacology and Toxicology. 2022;23(1):50
90.Andersson BS et al. Clofarabine ± fludarabine with once daily i.v. busulfan as pretransplant conditioning therapy for advanced myeloid leukemia and MDS. Biology of Blood and Marrow Transplantation. 2011;17(6):893-900
91.Gooptu M et al. A comparison of the myeloablative conditioning regimen fludarabine/busulfan with cyclophosphamide/total body irradiation, for allogeneic stem cell transplantation in the modern era: A cohort analysis. Biology of Blood and Marrow Transplantation. 2018;24(8):1733-1740
92.Gayoso J et al. Busulfan-based myeloablative conditioning regimens for haploidentical transplantation in high-risk acute leukemias and myelodysplastic syndromes. European Journal of Haematology. 2018;101(3):332-339
93.Chi H et al. Effect of genistein administration on the recovery of spermatogenesis in the busulfan-treated rat testis. Clinical and Experimental Reproductive Medicine. 2013;40(2):60-66
94.Suttorp M, Millot F. Treatment of pediatric chronic myeloid leukemia in the year 2010: Use of tyrosine kinase inhibitors and stem-cell transplantation. Hematology. 2010;2010(1):368-376
95.Ghasemi FM et al. Effect of melatonin on proliferative activity and apoptosis in spermatogenic cells in mouse under chemotherapy. Journal of Reproduction and Contraception. 2010;21(2):79-94
96.Ogawa T, Dobrinski I, Brinster R. Recipient preparation is critical for spermatogonial transplantation in the rat. Tissue and Cell Biology International. 1999;31(5):461-472
97.Kanatsu-Shinohara M et al. Germline niche transplantation restores fertility in infertile mice. Human Reproduction. 2005;20(9):2376-2382
98.Kurkure P et al. Very small embryonic-like stem cells (VSELs) detected in azoospermic testicular biopsies of adult survivors of childhood cancer. Reproductive Biology and Endocrinology. 2015;13:122
99.Hjollund NH et al. Diurnal scrotal skin temperature and semen quality. The Danish First Pregnancy Planner Study Team. International Journal of Andrology. 2000;23(5):309-318
100.Wang C et al. Effect of increased scrotal temperature on sperm production in normal men. Fertility and Sterility. 1997;68(2):334-339
101.Mieusset R, Bengoudifa B, Bujan L. Effect of posture and clothing on scrotal temperature in fertile men. Journal of Andrology. 2007;28(1):170-175
102.Chowdhury AK, Steinberger E. Early changes in the germinal epithelium of rat testes following exposure to heat. Journal of Reproduction and Fertility. 1970;22(2):205-212
103.Munkelwitz R, Gilbert BR. Are boxer shorts really better? A critical analysis of the role of underwear type in male subfertility. The Journal of Urology. 1998;160(4):1329-1333
104.Jensen TK et al. Inhibin B as a serum marker of spermatogenesis: Correlation to differences in sperm concentration and follicle-stimulating hormone levels. A study of 349 Danish men. The Journal of Clinical Endocrinology and Metabolism. 1997;82(12):4059-4063
105.Hjollund NH et al. The relation between daily activities and scrotal temperature. Reproductive Toxicology. 2002;16(3):209-214
106.Shiraishi K, Matsuyama H, Takihara H. Pathophysiology of varicocele in male infertility in the era of assisted reproductive technology. International Journal of Urology. 2012;19(6):538-550
107.Carlsen E et al. History of febrile illness and variation in semen quality. Human Reproduction. 2003;18(10):2089-2092
108.Lue YH et al. Single exposure to heat induces stage-specific germ cell apoptosis in rats: Role of intratesticular testosterone on stage specificity. Endocrinology. 1999;140(4):1709-1717
109.Zhang M et al. Autophagy and apoptosis act as partners to induce germ cell death after heat stress in mice. PLoS One. 2012;7(7):e41412
110.Fijak M et al. Infectious, inflammatory and 'autoimmune' male factor infertility: How do rodent models inform clinical practice? Human Reproduction Update. 2018;24(4):416-441
111.Hasan H et al. Mechanism of inflammatory associated impairment of sperm function, spermatogenesis and steroidogenesis. Frontiers in Endocrinology. 2022;13:897029
112.Pröbstl C et al. Immune cell characterization in spontaneous autoimmune orchitis in dogs. Theriogenology. 2022;187:219-226
113.Nieschlag E et al. Disorders at the testicular level. Andrology: Male Reproductive Health Dysfunction. 2010:193-238
114.Nicolas N et al. Testicular activin and follistatin levels are elevated during the course of experimental autoimmune epididymo-orchitis in mice. Scientific Reports. 2017;7:42391
115.Rusz A et al. Influence of urogenital infections and inflammation on semen quality and male fertility. World Journal of Urology. 2012;30(1):23-30
116.Abdallah AE et al. Immunomodulatory quinazoline-based thalidomide analogs: Design, synthesis, apoptosis and anticancer evaluations. Journal of Molecular Structure. 2023;1281:135164
117.Agarwal A et al. Male infertility. The Lancet Diabetes & Endocrinology. 2021;397(10271):319-333
118.Sagaradze G et al. Regenerative medicine for male infertility: A focus on stem cell niche injury models. Biomedical Journal. 2022;45(4):607-614
119.Gunes S et al. The role of epigenetics in idiopathic male infertility. Journal of Assisted Reproduction. 2016;33:553-569
120.Luján S et al. Sperm DNA methylation epimutation biomarkers for male infertility and FSH therapeutic responsiveness. Scientific Reports. 2019;9(1):16786
121.Salas-Huetos A et al. The role of miRNAs in male human reproduction: A systematic review. Andrology. 2020;8(1):7-26. Epub 2019/10/04. DOI: 10.1111/andr.12714
122.Kothandaraman N et al. Pathogenic landscape of idiopathic male infertility: New insight towards its regulatory networks. NPJ Genomic Medicine. 2016;1(1):1-9
123.Razavi SM et al. Comprehensive functional enrichment analysis of male infertility. Scientific Reports. 2017;7(1)
124.Bansal SK et al. Differential genes expression between fertile and infertile spermatozoa revealed by transcriptome analysis. PLoS One. 2015;10(5):e0127007
125.Zhao LY et al. Single-cell analysis of developing and azoospermia human testicles reveals central role of Sertoli cells. Nature. Communications. 2020;11(1)
126.Andersson AM et al. Impaired Leydig cell function in infertile men: A study of 357 idiopathic infertile men and 318 proven fertile controls. Journal of Clinical Endocrinology and Metabolism. 2004;89(7):3161-3167
127.Cinà DP, Phillips D, Flannigan R. CRISPR/Cas9 in male factor infertility. Current Tissue Microenvironment Reports. 2020;1(3):89-97
128.Liu J et al. Generation and characterization of reprogrammed sheep induced pluripotent stem cells. Theriogenology. 2012;77(2):338-346 e1
129.Aydos K, Aydos OS. Sperm selection procedures for optimizing the outcome of ICSI in patients with NOA. Journal of Clinical Medicine. 2021;10(12):2687
130.Kim D et al. Long-acting recombinant human follicle-stimulating hormone (SAFA-FSH) enhances spermatogenesis. Frontiers in Endocrinology. 2023;14:1132172
131.Assidi M. Infertility in men: Advances towards a comprehensive and integrative strategy for precision theranostics. Cell. 2022;11(10):1711
132.Sivaraman MAF. Ethical guiding principles of “do no harm” and the “intention to save lives” in relation to human embryonic stem cell research: Finding common ground between religious views and principles of medical ethics. Asian Bioethics Review. 2019;11(4):409-435
133.de Jongh D et al. Early-phase clinical trials of bio-artificial organ technology: A systematic review of ethical issues. Transplant International. 2022;35:10751
134.Macpherson I, Roqué MV, Segarra I. Ethical challenges of germline genetic enhancement. Frontiers in Genetics. 2019;10:767
135.Manghwar H et al. CRISPR/Cas systems in genome editing: Methodologies and tools for sgRNA design, off-target evaluation, and strategies to mitigate off-target effects. Advanced Science. 2020;7(6):1902312
136.Von Tigerstrom B. Revising the regulation of stem cell-based therapies: Critical assessment of potential models. Food and Drug. 2015;70:315
137.Laurencin CT, McClinton A. Regenerative cell-based therapies: Cutting edge, bleeding edge, and off the edge. Regenerative Engineering and Translational Medicine. 2020;6:78-89
138.Rayment EA, Williams DJ. Concise review: Mind the gap: Challenges in characterizing and quantifying cell-and tissue-based therapies for clinical translation. Stem Cells. 2010;28(5):996-1004
139.Najar M et al. Mesenchymal stromal cell-based therapy: New perspectives and challenges. Journal of Clinical Medicine. 2019;8(5):626
Written By
Ahmed Atwa, Serag Eldin I. Elbehairi, Sayed Bakry, Ahmed B.M. Mehany, Mahmoud Ashry, Hussam Askar and Mohammad Y. Alfaifi
Submitted: 01 September 2023Reviewed: 22 September 2023Published: 15 November 2023
 Open access peer-reviewed chapter
Open access peer-reviewed chapter