 Open access peer-reviewed chapter
Open access peer-reviewed chapter
Abstract
Wild huckleberry (Vaccinium globare/membranaceum complex) is a keystone species in the Pacific Northwest of the United States. The fruits are a primary food source for grizzly bears and other wildlife, as well as an important traditional and contemporary human food. Huckleberry shrubs also provide cover and nesting habitat for many animal species, including small mammals and birds. The flowers provide nectar and pollen with crucial connections between bumble bees (Bombus species) and huckleberries. Native bee pollination is essential for successful berry development. Huckleberries flower early in the growing season and are some of the only floral resources available when bumble bee queens first emerge from hibernation and need to collect pollen and nectar for nesting. One of these species, the Western bumble bee (Bombus occidentalis), is in review for listing under the U.S. Endangered Species Act. Future climate change has the potential to influence huckleberry distribution, productivity, and phenology. These potential changes could have wide-ranging implications because of the economic, cultural, and ecological importance of huckleberry.
Keywords
- Vaccinium
- huckleberry
- pollination
- bumble bees
- wildlife
- keystone species
- ecological relationships
1. Introduction
The genus

Figure 1.
Huckleberry phenophases (life stages) showing flower buds (top left), flowers (top middle), developing green berries (top right), ripe berries (bottom left), ripe berries and red leaves (bottom middle), and harvested berries (bottom right) (SKC, USGS photos).
The shrubs of wild huckleberry are deciduous and grow up to 2 m tall, often in rhizomatous clumps [1, 4, 5]. Leaves are oval to elliptical, 11–50 mm, usually green, and finely serrated. Flower petals are fused, giving them a globe-like shape, and are light pink or sometimes white. They grow individually on the underside of stem nodes. The berries are red to deep purple when ripe with many small (1 mm) seeds. Roots may be as deep as 100 cm with rhizomes usually occurring at depths of 8–30 cm. Huckleberries reproduce through both seeds and rhizomes with patches often consisting of vegetative clones [1, 4, 5].
Wild huckleberries are usually in areas with low canopy cover, fine and well-drained loam soils, moderate slopes, and winter precipitation as snow [5, 6]. Periodic fire (every 20–50 years) is important to prevent competition from other plants and to maintain relatively open overhead canopy although timber harvesting, avalanche chutes, and even ski runs can also lead to suitable conditions [7, 8].
Wild huckleberry shrubs are commonly associated with beargrass (
Wild huckleberry is one of the most important food plants in the Pacific Northwest of North America [10, 11] and has strong cultural [10, 11, 12, 13], economical [10, 14, 15], and ecological value [2, 16]. The berries of huckleberries are highly valued by the people of the Pacific Northwest, past and present. They are a cultural keystone species for many Native American tribes that continue traditions and ceremonies surrounding collection, storage, and consumption of berries [10, 11, 12, 13], as well as caretaking of huckleberry patches by spreading berries and seeds, pruning berry bushes, and setting fires to clear dense vegetation and promote growth [2, 10, 11, 12, 13]. Fruits are often eaten fresh but also cooked or dried [4]. Some tribes use the berries in pemmican or sun cakes, and to make lavender dye [4]. Various parts of huckleberry plants are medicinal. Some people use the leaves for tea or mix them with other plants for smoking [10].
Early frontier people utilized huckleberries as subsistence food [15]. Today, harvesting huckleberries is an activity many people enjoy as a special treat. People mostly eat fresh berries or bake them in foods, as well as preserve them canned, frozen, and as jams and jellies. Huckleberries are difficult to cultivate, and so most commercial berry products are from wild harvested berries [3], with most companies paying pickers to harvest berries for their products [7, 8]. Some commercial pickers harvest huckleberries to supplement other income and others sell huckleberries to meet financial needs [15].
Huckleberry shrubs provide cover, nesting habitat, and food for many animal species, including small mammals and birds [17]. The flowers of
Meteorological conditions often determine annual huckleberry phenology and distribution [28]. Huckleberry productivity in the Cabinet-Yaak Region was positively correlated with cooler, wetter springs, and 70% of the inter-annual variability was linked to April–June growing degree days and July temperature range based on a twenty-one year dataset [16]. Livestock grazing, mining, and the introduction of invasive weed species are often detrimental [12]. Fire suppression creates unfavorable conditions for huckleberry plants due to denser forest canopy and more competition from understory plants. Fire was an important tool for the Salish and Pend d’Oreille people who burned low-intensity fires to remove unhealthy plants and simulated higher berry productivity [13]. Much of the United States and Canada began preventing fire setting by indigenous people as early as the late 1800s [10]. Although there is growing recognition by mainstream forest managers that prescribed fire is important for the health of montane ecosystems, fuel buildup from past fire suppression has led to implementation challenges [29].
Furthermore, the Pacific Northwest is experiencing warming temperatures (0.7 degrees C warmer on average) and less precipitation [30], leading to decreases in snowpack and soil moisture. These factors are shifting the timing of berry ripening in some areas; thus, timing for harvesting plants has become less predictable [28, 31]. Models are being used to help determine how climate change might influence huckleberry distribution and phenological stages (flowering, berry ripening) in the Pacific Northwest region [28]. A study investigating how climate change could affect the wild huckleberry concluded that the suitable range of huckleberry across the Pacific Northwest United States will likely shrink by 5–40% by the end of the twenty-first century [28]. The study further suggests that the phenology of huckleberry will change, with flowering advancing 23–50 days and fruiting advancing 24–52 days, by the end of the twenty-first century [28]. Models are also being developed to predict sites where huckleberries are most likely to be found for human harvesting and bears [6, 32, 33, 34].
Climate fluctuations leading to changes in snow depth and timing of snowmelt may lead to phenological mismatches, thus interrupting species interactions and reducing populations of some species. Earlier snowmelt and shifts in the timing of flowering can increase exposure of flowers to frost and reduce berry production, resulting in years with poor to no berries [35, 36, 37, 38]. Furthermore, frost damage to flowers can impact pollinators. For example, fecundity of the alpine butterfly (
There are also concerns that climate change could increase population size and spread of pest species and pathogens. Warmer temperatures are expected to make conditions more suitable for invasive plant species, resulting in increases at higher elevations and latitudes [41]. Invasive plants tend to have rapid growth rates, wide dispersal, and the ability to outcompete native plants [41]. Many of the pest insect species in the Order Hemiptera (true bugs) have piercing/sucking mouthparts and can cause economic damage [42]. Three hemipteran families collected from the leaves and stems of wild huckleberry plants in Montana, Cicadellidae (leafhoppers), Aphidadae (aphids), and Miridae (plant bugs) include species known to cause damage to fruit crops and two species in particular, the leafhopper (
Species-specific responses to climate change can also disrupt plant-pollinator interactions [45, 46]. Environmental cues such as air temperature and day length influence the phenological responses of plants, insects, and other animal species [35, 40, 47, 48]. Snowpack can play a major role in the emergence of plants and insects [49, 50, 51]. In Japan, early snowmelt also increased the risk of phenological mismatch between a spring ephemeral plant and its bumble bee pollinators, likely because snow melt happened sooner than increasing soil temperatures [52]. The indirect effects on flowers can even influence bee abundances [53].
Pollinator declines and shifts in assemblages are already impacting wild plant species in some places [54]. Most commercially grown
Greater than 85% of fruit set and yield of plants in the genus
Potential changes in the habitat, health, productivity, abundance, and distribution of huckleberry plants and the species they interact with have wide-ranging implications because of the economic, cultural, and ecological importance of huckleberry. Climate change has a high potential to impact huckleberry ecology and the species they interact with such as pollinators. To better understand the potential impact of climate change on wild huckleberry, we conducted an array of studies on huckleberry phenology and plant visitors (particularly pollinators) during the years of 2015 through 2021.
2. Research on the Flathead Indian reservation, Montana
2.1 Study area
The research described in this section took place on the Flathead Indian Reservation of the Confederated Salish and Kootenai Tribes in northwest Montana, USA (Figure 2). Ten study sites were selected based on aspect, elevation (1200 to 1900 m), slope, plant community, geographical location, and accessibility. Dominant tree species recorded at these sites were Douglas fir, ponderosa pine, western larch, lodgepole, subalpine fir (
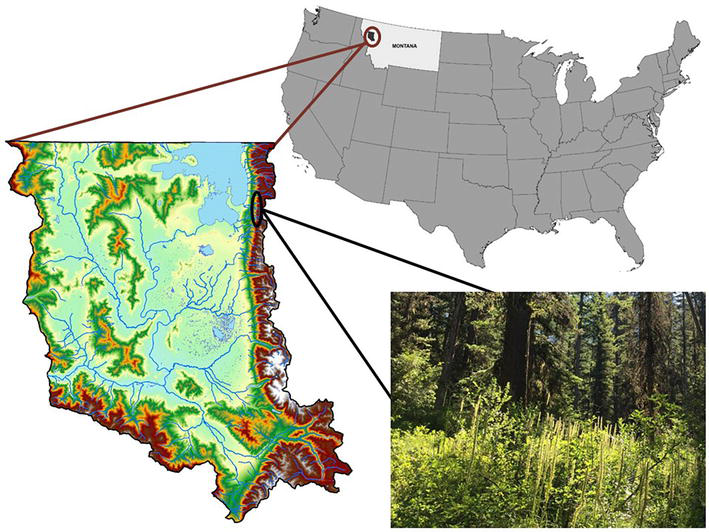
Figure 2.
Location of the Flathead reservation in northwestern Montana, USA, showing topography and landmarks within the reservation boundary, and an example of one of the study sites with huckleberry and beargrass in the foreground (CSKT maps, SKC photo).
The average annual temperature in northwest Montana is 4.8°C, ranging from −4.6°C in winter to 14.7°C in summer [67]. Temperatures are cooler and more variable at higher elevations. The state growing season averaged 206 days (1981–2010). The average annual precipitation in northwest Montana is 82.2 cm with a range of an average winter minimum of 23.9 cm to an average summer maximum of 15.5 cm. Higher mountain elevations can receive as much as 150.8 cm annually. Most winter precipitation in Montana is snow that accumulates as snowpack [67].
Based on data from the nearest USDA NRCS SNOTEL station within the elevation range at our sites (Bisson Creek, 1500 m), snow was present during the months of December through April for the winters of 2014–2015 and 2015–2016 [68]. During the winters of 2016 through 2021 snow was similarly present in December but also continued into the month of May. During this study, the highest snow water equivalent (SWE) recorded was 348 mm at the start of April 2018 and the lowest SWE recorded in April was 124 mm in the year 2015. Mean spring precipitation (March–June) was the highest in 2017 (666 mm) followed by 2018 (590.25 mm) and 2015 (548.5 mm). Spring precipitation was the lowest in 2021 (419.75 mm) followed by 2020 (421.25 mm), 2016 (472.5 mm), and then 2019 (485.75 mm) [68].
2.2 Huckleberry and vertebrate visitors
A variety of methods were utilized to record huckleberry visitors including standardized bird point counts, standardized insect observations, incidental sightings, and remote cameras. During 12 hours of point count surveys at 4 sites (each visited 3 times), we documented a total of 18 species and 388 individual birds. Of these 18 species, those known to eat or that were observed eating ripe huckleberries were ruffed grouse (
Bird species photographed in the huckleberry plants while berries were ripe included ruffed grouse, American Robin, Canada jay, Western tanager, and chipping sparrow (

Figure 3.
Some of the bird species photographed near huckleberry plants at study sites located on the Flathead reservation. Calliope hummingbird (top left), calliope hummingbird before daylight (top right), ruffed grouse (bottom left), and dark-eyed junco (bottom right) (SKC, USGS camera photos).
Mammals photographed feeding on huckleberries were chipmunk (

Figure 4.
Some of the mammal species photographed near huckleberry plants at study sites located on the Flathead reservation. Snowshoe hare (top left), northern flying squirrel (middle left), white-tailed deer fawn (bottom left), black bear (top right), and deer mouse (bottom right) (SKC, USGS camera photos).
2.3 Huckleberry flowers and insect visitors
Flower-visiting insects were more specifically targeted with time-constrained surveys, net captures, photographs, and incidental sightings. The majority of visitors observed were in the Order Hymenoptera (bees, wasps, hornets, ants). Some Lepidoptera (butterflies and moths) and Diptera (flies) were observed including several bee mimics; clearwing moths (
Standardized, time-constrained (15 min) focal surveys of individual huckleberry plants were used to quantify the percentage of insects visiting huckleberry flowers. We conducted 359 focal surveys (approximately 90 hours of observations) during huckleberry flowering 2018–2020. Insect visitors included 569 individuals observed during 309 of the surveys. The group most frequently visiting flowers was bees (50.4%), followed by vespids (wasps and hornets, 18.3%), dipterans (flies, 18.1%), large ants (6.5%), and other [lepidopterans (moths and butterflies, 2.5%), coleopterans (beetles, 1.8%), and unidentified/other, 2.1%] (Figure 5). On multiple occasions, vespids were observed chewing through flower petals. Small ants were not counted because when observed there were usually large numbers moving around the plant. Bumble bees made up 85.6% of the total bees observed visiting flowers, the majority of bumble bees appearing to be queens based on size, which also aligns with the timing of queen bumble bee activity.
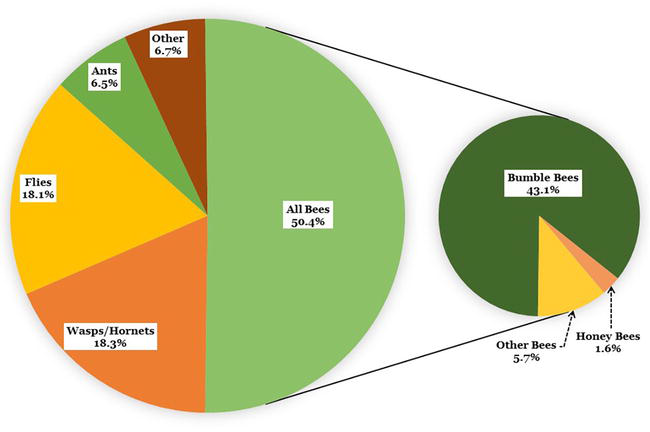
Figure 5.
Insects observed visiting huckleberry flowers at study sites on the Flathead reservation, Montana, USA.
Most flower visitors other than bees do not actively forage for, or collect, pollen as a source of food for developing larvae. Vespids typically visit flowers either for nectar or to find prey [69] and our observations of vespids chewing holes in flowers are a sign of nectar robbing [70]. Ants feed on nectar and may move pollen but they probably have a negligible role in pollination [71]. Some dipterans may feed on nectar, and some may be pollinators [72]. Lepidopterans are pollinators but were only occasionally observed visiting huckleberry flowers. Because bees were the most common huckleberry flower visitors and are known to be efficient pollinators, and many species specifically collect pollen as a food source for developing larvae; we concluded that bees, especially bumble bees, are the main pollinators of huckleberry flowers.
2.4 Bumble bees as primary probable pollinators
Bumble bees are capable of sonication (buzz pollination) which might be necessary to rapidly vibrate the globe-like flowers to access pollen from the recessed anthers [63, 64, 73]. Some other native bee species are also capable of sonication [19, 73] but only contributed to a small proportion of the total flower visitors. European honey bees can be efficient pollinators, but only nine individuals were observed during focal surveys at a single site and they are not capable of sonication [73]. Honey bees likely came from hives cultivated for honey at a lower elevation. This species is not common in huckleberry habitats in Montana where most cultivated hives are not located near montane forested areas.
Because we observed bumble bees to be the most frequent visitor of huckleberry flowers, we specifically captured them for non-lethal species identification. Bumble bees were captured with sweep nets at the huckleberry sites between the years 2018 and 2021 using 45-minute standardized surveys. Each captured bee was placed in a vial labeled with the flower they were observed visiting if applicable then chilled in a cooler. Once the 45-minute capture period was complete, chilled bees were placed on gray cardboard, photographed from multiple angles using a close-up lens with ring lighting, then released. The photographs were submitted to a bumble bee specialist for identification.
There were 192 bumble bees that were both observed visiting flowering huckleberry plants and identifiable to species with confidence. From these, we identified 10 bumble bee species visiting huckleberry flowers. These species were
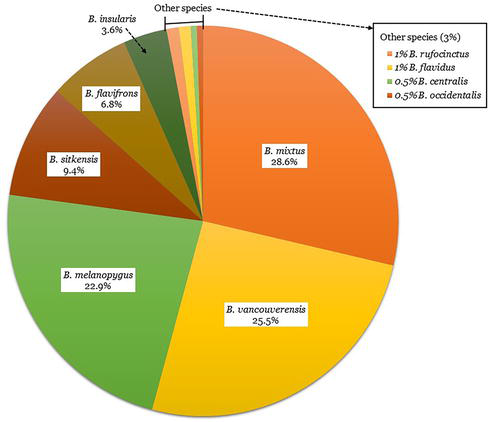
Figure 6.
Bumble bees captured from flowering huckleberry plants on the Flathead reservation, Montana, USA, and identified to species from photographs.
2.5 Huckleberries need bumble bee pollination
To determine the value of pollination for berry production, field experiments are needed and yet uncommon [74], particularly studies that consider the influence of pollination of wild plants using exclusion experiments [75]. We conducted a pollinator exclusion field experiment to quantify the potential importance of pollination for the development of huckleberries (Figure 7). We selected 120 huckleberry plants at one of the lower elevation (1250 m) study sites near Polson, Montana, USA, for this work. Half of these plants were covered with mesh bags sewn from nylon tulle netting to exclude natural pollinators and half were artificially hand-pollinated using pollen wands resulting in four treatments; (1) extra pollination: natural pollinators able to access flowers and plants also artificially pollinated by hand, (2) natural pollination: pollinators able to access flower but no additional pollination, (3) artificial pollination: natural pollinators excluded but plants artificially pollinated by hand, and (4) no pollination: natural pollinators excluded and no additional pollination. Pollination treatments (extra pollination [630 berries], natural pollination [545], and artificial pollination [365]) had far more berries than the no pollination plants (63 berries).

Figure 7.
Photograph of a subset of the 120 huckleberry plants (60 within tulle netting) included in a pollinator exclusion study on the Flathead reservation, Montana, USA (SKC photos).
Based on our observations, standardized focal surveys, and targeted captures, bumble bees were the primary visitors to huckleberry flowers. When bumble bees and other insects were excluded from pollinating huckleberry flowers, berry development was greatly limited. Even in this highly natural system at a site with overall high productivity in the year of our research, we saw a significant effect of pollen limitation on berry production.
3. Conclusions
Global climate change is resulting in warming temperatures and less precipitation in the Pacific Northwest [30]. Montana is also experiencing warmer temperatures that are predicted to increase by 3.1 to 5.4 degrees C by the end-of-the-century [67]. Precipitation in Montana has shown the greatest declines during winter months, particularly in northwest Montana. In Montana’s mountains, 62–65% of annual precipitation comes from snow. Snowpack in Montana has declined since the 1930s, particularly in the mountains surrounding the Continental Divide, and snowmelt is occurring earlier in the spring. Both of these trends are likely to continue with warming temperatures [67] and have led to large shifts in the phenological start of spring for vegetation, though the direction of change varies spatially [76]. In addition, forest pests and pathogens are predicted to increase [67]. Tribal elder observations further support shifts in weather patterns, less snow, and warmer temperatures in Montana, from observations within their lifetimes and knowledge gained from prior generations [77].
Climate-related changes have resulted in documented shifts in the range of huckleberry plants and timing of berry ripening in some locations [28, 33]. Models further suggest that the range of huckleberry will decrease at lower altitudes and at the southern edge of their distribution range and perhaps shift to higher altitudes and northward, although germination or other constraints may influence this [28, 33]. Such changes in location, number, health, and timing for accessing traditional foods may alter cultural connections and is a concern of tribal communities [77]. The Séliš and Q̓l̓ispe̓ people refer to August as St̓šá Spq̓ni (Month of the Huckleberry) yet at some sites we found ripe berries as early as June. Tribal elder interviews concerning climate change mention huckleberries specifically. For example, Sadie Saloway said, “The huckleberries are just disappearing so fast” and Stephen SmallSalmon said, “We changed that…about the weather. You could tell by the choke cherries getting smaller and the huckleberries getting smaller. I saw that happen.” Threats posed by climate change have the potential to threaten native cultures and traditional ways of life [77].
The potential shifts in huckleberry range, numbers, and phenology could also impact the crucial interactions between bumble bees and huckleberries. In Colorado, there is documentation of a growing phenological mismatch between glacier lily (
Changes in huckleberry availability and phenology could also have concerning impacts for bumble bees in montane forest habitats. Huckleberries are some of the first and only flowering plants available in montane forest sites in northwest Montana when bumble bee queens emerge from hibernation in the spring. Recent research highlights the role of forests during the queen lifestage [80]. Bumble bees at these sites likely rely on early-flowering plants such as huckleberry for nectar and pollen to begin new colonies. Seed production can be reduced when flowering of spring ephemeral plants proceeds the emergence of bumble bee pollinators [46].
Declining numbers of bumble bees could also be of concern to huckleberry ecology. It is estimated that as many as one in four North American bumble bee species are being negatively impacted by some level of stress [81]. Habitat changes may be the primary stressor threatening bumble bee populations as well as stress from parasites, pesticides, and lack of flowers [82, 83] and climate change is predicted to lead to further bumble bee declines and range losses in the future [84, 85].
This chapter has illustrated the exceptional value of wild huckleberry to the ecology and people of the Pacific Northwest of North America. Our work in Montana has further added documentation of the birds, mammals, and insects that are at least incidental visitors to huckleberry plants. We added to the knowledge of species that specifically visit huckleberry flowers for pollen and nectar. We were able to provide further support that the primary probable pollinators of huckleberry flowers are bumble bees. Finally, our research used exclusion experiments in the wild to definitively quantify the significance of bumble bee pollination for the development of huckleberries. More information is needed to better understand the ideal conditions for huckleberry health and productivity, ability to spread to newly suitable areas, potential impacts of fire suppression and climate change, ecological role of interacting species, potential phenological mismatches, and conservation actions to ensure healthy huckleberry harvests into the future.
Acknowledgments
We are grateful to the Confederated Salish and Kootenai Tribes of the Flathead Reservation for permissions to conduct this work on tribal lands. We would like to thank all of the people that contributed to these projects. Dr. Laura Burkle and Casey Delphia provided feedback on our initial pollinator exclusion design and Amy Dolan for identifying bumble bees from field photos. Many people helped with the set-up, data collection, and lab work related to the exclusion study including Morgan Schultz, Madison Dimacle, Marcy Mead, Dennis Lichtenberg, Anthony Lichtenberg, Dion Lichtenberg, and Brett Stevenson. In the following years, Rebekah Brassfield served as a crew leader for an array of student research internship projects that included maintaining remote field cameras and data loggers as well as additional bumble bee focal studies and captures. Additional students that contributed to this project were Amanda Spencer, Rustin Bielski, Aspen Jaeger, Kendra Melanson, and Sarah Zurkee. Much of this work was funded by the National Science Foundation (NSF) Tribal College and University Small Grant for Research award number NSF TCP 1721243. Additional funding sources included the NSF Research Experience for Undergraduates Sustainable Land and Water Resources award number NSF EAR 1757451. The U.S. Geological Survey Species Management Program supported Tabitha Graves. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
References
- 1.
Montana Natural Heritage Program. Montana field guide. 2022. Available from: https://fieldguide.mt.gov/displaySpecies.aspx?family=Ericaceae [Accessed November 11, 2022] - 2.
Keefer M, Cocksedge W, Munro R, Meuleman J, MacPherson N. What about the Berries? Managing for Understory Species. British Columbia: The Centre for Livelihoods and Ecology Royal Roads University; 2010. p. 89 - 3.
Vander Kloet SP. Vaccinium membranaceum . Flora of North America Association [Internet]. 2020. Available from:https://floranorthamerica.org/Vaccinium_membranaceum [Accessed November 26, 2022] - 4.
USDA (United States Department of Agriculture). NRCS (Natural Resources Conservation Service), PLANTS Database [Internet]. 2022. Available from: https://plants.usda.gov/home [Accessed November 26, 2022] - 5.
USDA Forest Service. Fire Effects Information System (FEIS), Species: Vaccinium membranaceum [Internet]. 2022. Accessible from: https://www.fs.usda.gov/database/feis/plants/shrub/vacmem/all.html#27 [Accessed November 26, 2022] - 6.
Proctor M, Lamb C, MacHutchon G. The Grizzly Dance of Berries and Bullets: The Relationship between Bottom up Food Resources, Huckleberries, and Top Down Mortality Risk on Grizzly Bear Population Processes in Southeast British Columbia. Kaslo (British Columbia): Trans-border Grizzly Bear Project; 2017 - 7.
Horowitz E. Huckleberry hounds: Sniffing out Montana's perfect gem. Montana Outdoors. 2004 - 8.
Martin P. Factors influencing globe huckleberry fruit production in Northwestern Montana. Bears: Their Biology and Management. 1980; 5 :159-165 - 9.
Simonin KA. Fire effects information system: Vaccinium membranaceum. [Internet]. 2018. Available from:https://www.fs.fed.us/database/feis/plants/shurb/vacmem/all.html . [Accessed May 29, 2018] - 10.
Richards RT, Alexander SJ. 2A Social History of Wild Huckleberry Harvesting in the Pacific Northwest. Portland (Oregon): Pacific Northwest Research Station; 2006. p. 113. DOI: 10.2737/PNW-GTR-657 - 11.
Trusler S, Johnson LM. Berry patch as a kind of place- the ethnoecology of black huckleberry in Northwest Canada. Human Ecology. 2008; 36 :553-568. DOI: 10.1007/s10745-008-9176-3 - 12.
Turner NJ, Douglas D, Mellott CR. Up on the mountain: Ethnobotanical importance of montane sites in the Pacific coastal North America. Journal of Ethnobiology. 2011; 31 (1):4-43. DOI: 10.2993/8-0771-31.1.4 - 13.
Salish C, Tribes K. Fire on the Land: An Interactive DVD [DVD]. Omaha (NB): University of Nebraska Press; 2005 - 14.
Thomas MG, Schumann DR. Chapter 2. Berries and wild fruit. In: Thomas MG. Schumann DR. Business Opportunities in Special Forest Products: Self-Help Suggestions for Rural Entrepreneurs, Agriculture Information Bulletin AIB-666. Lincoln (NB): Nebraska Forest Service; 1993. pp. 17-24. https://digitalcommons.unl.edu/nebforestpubs/46 - 15.
Carroll MS, Blatner KA, Cohn PJ. Somewhere between: Social embeddedness and the spectrum of wild edible huckleberry harvest and use. Rural Sociology. 2010; 68 (3):319-342. DOI: 10.1111/j.1549-0831.2003.tb00140.x - 16.
Holden ZA, Kasworm WF, Servheen C, Hahn B, Dobrowski S. Sensitivity of berry productivity to climatic variation in the Cabinet–Yaak grizzly bear recovery zone, Northwest United States, 1989-2010. Wildlife Society Bulletin. 2012; 36 (2):226-231. DOI: 10.1002/wsb.128 - 17.
Martin AC, Zim HS, Nelson AL. American Wildlife and Plants: A Guide to Wildlife Food Habits. New York: Dover Publications; 1980. p. 2051 - 18.
Dolan AC. Insects Associated with Montana’s Huckleberry (Ericaceae: Vaccinium globulare ) Plants and the Bumble Bees (Hymenoptera: Apidae) of Montana. Bozeman: Montana State University; 2016 - 19.
Cane JH, Payne JA. Regional, annual, and seasonal variation in pollinator guilds: Intrinsic traits of bees (Hymenoptera: apoidea) underlie their patterns of abundance at Vaccinium ashei (Ericaceae). Annals of the Entomological Society of America. 1993; 86 :577-588. DOI: 10.1093/aesa/86.5.577 - 20.
Pierce JD. Shiras moose forage selection in relation to browse availability in north-Central Idaho. Canadian Journal of Zoology. 1984; 62 :2404-2409. DOI: 10.1139/z84-354 - 21.
Edge WD, Marcum CI, Olson-Edge SI. Summer forage and feeding site selection by elk. Journal of Wildlife Management. 1988; 52 (4):573-577. DOI: 10.2307/3800909 - 22.
Klebenow DA. A montane forest winter deer habitat in western Montana. Journal of Wildlife Management. 1985; 29 (1):27-33. DOI: 10.2307/3798627 - 23.
McLellen BN, Hovey FW. The diet of grizzly bears in the Flathead River drainage of Southern British Columbia. Canadian Journal of Zoology. 1995; 73 (4):704-712. DOI: 10.1139/z95-082 - 24.
Welch CA, Keay J, Kendall KC, Robbins CT. Constraints on frugivory by bears. Ecology. 1997; 78 :1105-1119. DOI: 10.2307/2265862 - 25.
McLellan BN. Some mechanisms underlying variation in vital rates of grizzly bears on a multiple use landscape. Journal of Wildlife Management. 2015; 79 :749-765. DOI: 10.1002/jwmg.896 - 26.
Kendall KC. Grizzly and Black Bear Feeding Ecology in Glacier National Park, Montana: Progress Report. West Glacier (MT): Science Center, Glacier National Park; 1986 - 27.
McCall BS, Mitchell MS, Schwartz MK, Hayden J, Cushman SA, Zager P, et al. Combined use of mark-recapture and genetic analyses reveals response of a black bear population to changes in food productivity. Journal of Wildlife Management. 2013; 77 (8):1572-1582. DOI: 10.1002/jwmg.617 - 28.
Prevéy JS, Parker LE, Harrington CA, Lamb CT, Proctor MF. Climate change shifts in habitat suitability and phenology of huckleberry ( Vaccinium membranaceum ). Agriculture and Forest Meteorology. 2020;280 :1-12. DOI: 10.1016/j.agrformat.2019.107803 - 29.
Pyne SJ. Fire: A Brief History. Seattle (WA): University of Washington Press; 2001. p. 204. https://ebookcentral-proquest-com.weblib.lib.umt.edu:2443/lib/msoumt/detail.action?docID=3444376 - 30.
Abatzoglu JT, Rupp DE, Mote PW. Seasonal climate variability and change in the Pacific Northwest of the United States. Journal of Climate. 2013; 27 :2125-2142. DOI: 10.1175/JCLI-D-13-002181.1 - 31.
Dobkins R, Hummel SS, Lewis C, Pochis G, Dickey E. Tribes of the Oregon Country: Cultural plant harvests and indigenous relationships with ancestral lands in the twenty-first century. Oregon Historical Quarterly. 2017; 118 (4):488-517. DOI: 10.5403/oregonhistq.118.4.0488 - 32.
Shores CR, Mikle N, Graves TA. Mapping a keystone shrub species, huckleberry ( Vaccinium membranaceum ), using seasonal colour change in the Rocky Mountains. International Journal of Remote Sensing. 2019;40 (15):5695-5715. DOI: 10.1080/01431161.2019.1580819 - 33.
Roberts DR, Nielsen SE, Stenhouse GB. Idiosyncratic responses of grizzly bear habitat to climate change based on projected food resource changes. Ecological Applications. 2014; 24 :1144-1154. DOI: 10.1890/13-0829.1 - 34.
Lamb CT, Mowat G, McLellan BN, Nielson SE, Boutin S. Forbidden fruit: Human settlement and abundant fruit create an ecological trap for an apex omnivore. Journal of Animal Ecology. 2017; 86 :55-65. DOI: 10.1111/1365-2656.12589 - 35.
Inouye DW. The ecological and evolutionary significance of frost in the context of climate change. Ecology Letters. 2000; 3 :457-463. DOI: 10.1046/j.1461-0248.2000.00165.x - 36.
Inouye DW. Effects of climate change on phenology, frost damage, and floral abundance of montane wildflowers. Ecology. 2008; 89 :353-362. DOI: 10.1890/06-2128.1 - 37.
Augspurger CK. Reconstructing patterns of temperature, phenology, and frost damage over 124 years: Spring damage risk is increasing. Ecology. 2013; 94 :41-50. DOI: 10.1890/12-0200.1 - 38.
Straka JR, Starzomski BM. Fruitful factors: What limits seed production of flowering plants in the alpine? Oecologia. 2015; 178 :249-260. DOI: 10.1007/s00442-014-3169-2 - 39.
Boggs CL, Inouye DW. A single climate driver has direct and indirect effects on insect population dynamics. Ecology Letters. 2012; 15 :502-508. DOI: 10.1111/j461-0248.2012.01766.x - 40.
Bale JS, Hayward SAL. Insect overwintering in a changing climate. Journal of Experimental Biology. 2010; 213 :980-994. DOI: 10.1242/jeb.037911 - 41.
Mainka SA, Howard GW. Climate change and invasive species: Double jeopardy. Integrated Zoology. 2010; 5 :102-111. DOI: 10.1111/j.1749-4877.2010.00193.x - 42.
Schuh RT, Slater JA. True Bugs of the World (Hemiptera:Heteroptera). Classification and Natural History. Ithica (NY): Comstock Publishing Associates; 1995. p. 336 - 43.
Langille AB, Arteca EM, Newman JA. The impacts of climate change on the abundance and distribution of the Spotted Wing Drosophila ( Drosophila suzukii ) in the United States and Canada. PeerJ. 2017, 2017;5 :e3192. DOI: 10.7717/peerj.3192 - 44.
McCloskey SPJ, Daniels LD, McLean JA. Potential impacts of climate change on western hemlock looper outbreaks. Northwest Science. 2009; 83 (3):225-238. DOI: 10.3955/046.083.0306 - 45.
Donnelly A, Caffarra A, O'Neill BF. A review of climate-driven mismatches between interdependent phenophases in terrestrial and aquatic ecosystems. International Journal of Biometeorology. 2011; 55 :805-817. DOI: 10.1007/s00484-011-0426-5 - 46.
Kudo G, Ida TY. Early onset of spring increases the phenological mismatch between plants and pollinators. Ecology. 2013; 94 (10):2311-2320. DOI: 10.1890/12-2003.1 - 47.
Post E, Forchhammer MC. Climate change reduces reproductive success of an Arctic herbivore through trophic mismatch. Philosophical Transactions of the Royal Society of British Biological Sciences. 2008; 363 :2369-2375. DOI: 10.1098/rstb.2007.2207 - 48.
Doiron M, Gauthier G, Levesque E. Trophic mismatch and its effects on the growth of young in an Arctic herbivore. Global Change Biology. 2015; 21 :4364-4376. DOI: 10.1111/gcb.13057 - 49.
Kreyling J. Winter climate change: A critical factor for temperate vegetation performance. Ecology. 2010; 91 :1939-1948. DOI: 10.1890/09-1160.1 - 50.
Wipf S, Rixen C. A review of snow manipulation experiments in the Arctic and alpine tundra ecosystems. Polar Research. 2010; 29 :95-109. DOI: 10.1111/j.1751-8369.2010.00153.x - 51.
Cooper EJ. Warmer shorter winters disrupt arctic terrestrial ecosystems. Ecological and Evolutionary Systems. 2014; 45 :271-295. DOI: 10.1146/annurev-ecolsys-120213-091620 - 52.
Kudo G, Cooper EJ. When spring ephemerals fail to meet pollinators: Mechanisms of phenological mismatch and its impact on plant reproduction. Proceedings of the Royal Society B: Biological Sciences. 2019; 286 :20190573. DOI: 10.6084 - 53.
Ogilvie JE, Griffin SR, Gezon KJ, Inouye BD, Underwood N, Inouye DW, et al. Interannual bumble bee abundance is driven by indirect climate impacts on floral resource phenology. Ecology Letters. 2017; 20 :1507-1515. DOI: 10.1111/ele.12854 - 54.
Biesmeijer JC, Roberts PM, Reemer M, Ohlemuller R, Edwards M, Peeters T, et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science. 2006; 313 :351-353. DOI: 10.1126/science.1129551 - 55.
Isaacs R, Kirk AK. Pollination services provided to small and large highbush blueberry fields by wild and managed bees. British Ecological Society. 2010; 47 (4):841-849. DOI: 10.1111/j.1365-2664.2010.01823.x - 56.
Kearns CA, Inouye DW, Waser NM. Endangered mutualisms: Conservation of plant-pollinator interactions. Annual Review of Ecology and Systematics. 1998; 29 :83-112. DOI: 10.1146/annurev.ecolsys.29.1.83 - 57.
Withgott J. Pollination migrates to top of conservation agenda. Bioscience. 1990; 49 (11):857-862. DOI: 10.2307/1313643 - 58.
Thomann M, Imbert E, Devaux C, Cheptou PO. Flowering plants under global pollinator declines. Trends in Plant Science. 2013; 18 (7):353-359. DOI: 10.1016/j.tplants.2013.04.002 - 59.
Graves TA, Janousek WM, Gaulke SM, Nicholas AC, Keinath DA, Bell CM, et al. Western bumble bee: Declines in the continental United States and range-wide information gaps. Ecosphere. 2020; 11 (6):1-13. DOI: 10.1002/ecs2.3141 - 60.
Klein AM, Vaissier BE, Cane JH, Steffan-Dewenter I, Cunningham SA, Kremen C, et al. Importance of pollinators in changing landscapes for world crops. Proceedings of the Royal Society Bulletin. 2006; 270 (1608):955-961. DOI: 10.1098./rspb.2006.3721 - 61.
Burd M. Bateman's principle and plant reproduction: The role of pollen limitation in fruit and seed set. Botanical Review. 1994; 60 (1):83-139. DOI: 10.1007/bf02856594 - 62.
Aras P, DeOliveira D, Savoie L. Effect of a honey bee (Hymenoptera: Apidae) gradient on the pollination and yield of lowbush blueberry. Journal of Economic Entomology. 1996; 89 (5):1080-1083. DOI: 10.1093/jee/89.5.1080 - 63.
Javorek S, Mackenzie K, Vander KS. Comparative pollination effectiveness among bees (Hymenoptera: Apoidea) on lowbush blueberry (Ericaceae: Vaccinium augustifolium ). Annals of the Entomological Society of America. 2002;95 :345-351. DOI: 10.1603/0013-8746(2002)095%5B0345:CPEABH%5D2.0.CO;2 - 64.
Ratti CM, Higo HA, Griswold TL, Winston ML. Bumble bees influence berry size in commercial Vaccinium spp. cultivation in British Columbia. Canadian Entomologist. 2008; 140 (3):348-363. DOI: 10.4039/n07-065 - 65.
MacKenzie KE. Pollination requirements of three highbush blueberry (Vaccinium corymbosum L.) cultivars. Journal of the American Horticultural Society. 1997; 122 (6):891-896. DOI: 10.21273/jashs.122.6.891 - 66.
Rogers SR, Tarpy DR, Burrack HJ. Bee species diversity enhances productivity and stability in a perennial crop. PLoS One. 2014; 9 (5):e97307. DOI: 10.1371/journal.pone.0097307 - 67.
Whitlock C, Cross W, Maxwell B, Silverman N, Wade AA. Montana Climate Assessment. Bozeman and Missoula, MT: Montana State University and University of Montana, Montana Institute on Ecosystems; 2017. p. 318. DOI: 10.15788/m2ww8w - 68.
USDA NRCS National Water and Climate Center. Report Generator 2.0. [Internet]. 2016. Available from https://wcc.sc.egov.usda.gov/reportGenerator/ [Accessed January 12, 2023] - 69.
Akre RD, Greene A, MacDonald F, Landolt PJ, Davis HG. Yellowjackets of America North of Mexico. Washington, DC: U.S. Department of Agriculture. p. 102 - 70.
Irwin RE, Bronstein JL, Manson JS, Richardson L. Nectar robbing: Ecological and evolutionary perspectives. Annual Review of Ecology, Evolution and Systematics. 2010; 41 :271-292. DOI: 10.1146/annurev.ecolsys.110308.120330 - 71.
Chacoff NP, Aschero V. Frequency of visits by ants and their effectiveness as pollinators of Condalia microphylla. Journal of Arid Environments. 2014; 2014 :91-91. DOI: 10.1016/j.jaridenv.2014.03.001 - 72.
Larson B, Kevan P, Inouye D. Flies and flowers: Taxonomic diversity of anthophiles and pollinators. The Canadian Entomologist. 2001; 133 :439-465. DOI: 10.4039/ent133439-4 - 73.
Cardinal S, Buchmann SL, Russell AL. The evolution of floral sonication, a pollen foraging behavior used by bees (Anthophila). Evolution. 2018; 72 (3):590-600. DOI: 10.1111/evo.13446 - 74.
Chacoff NP, Aizen MA, Aschero V. Proximity to forest edge does not affect crop production despite pollen limitation. Proceedings: Biological Sciences. 2008; 275 (1637):907-913. DOI: 10.1098/rspb.2007.1547 - 75.
Kearns CA, Inouye DW. Techniques for Pollination Biologists. Niwot (CO): University of Colorado Press; 1993. p. 583 - 76.
Berman E, Graves TA, Mikle N, Merkle J, Johnston A, Chong G. Comparing quality and trend of remotely sensed phenology and productivity metrics across the Western United States. Remote Sensing. 2020; 12 (16):2538. DOI: 10.3390/rs12162538 - 77.
Salish C, Tribes K. Climate Change Strategic Plan. Pablo (MT): Confederated Salish and Kootenai Tribes of the Flathead Nation; 2016. p. 86 - 78.
Thomson JD. Flowering phenology, fruiting success and progressive deterioration of pollination in an early-flowering genophyte. Proceedings of the Royal Society of London B. 2010; 365 :3187-3199. DOI: 10.1098/rstb.2010.0115 - 79.
USFWS (United States Fish and Wildlife Service). Federal Register Documents Proposed Rules. 2016; 81 (51) - 80.
Mola J, Hemberger J, Kochanski J, Richardson L, Pearse IS. The importance of forests in bumble bee ecology and conservation. Bioscience. 2008; 71 (12):1234-1248. DOI: 10.1093/biosci/biab121 - 81.
Colla SR, Gadallah F, Richardson L, Wagner D, Gall L. Assessing declines of north American bumble Bees (Bombus spp.) using museum specimens. Biodiversity and Conservation. 2012; 21 :3583-3595. DOI: 10.1007/s10531-0112-0383-2 - 82.
Goulson D, Hanley ME, Darvill B, Ellis JS, Knight ME. Causes of rarity in bumblebees. Biological Conservation. 2005; 122 :1-8. DOI: 10.1016/j.biocon.2004.06.017 - 83.
Goulson D, Nicholls E, Botias C, Rotheray EL. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science. 2015; 347 :1435-1444. DOI: 10.1126/science.1255957 - 84.
Janousek WM, Douglas M, Cannings S, Clement M, Delphia C, Everett J, et al. Recent and future declines in a historically widespread pollinator linked to climate, land cover, and pesticides. Proceedings North American Academy of Sciences. 2023; 120 (5):1-9. DOI: 10.1073/pnas.2211223120 - 85.
Sirois-Delisle C, Kerr JT. Climate change-driven range losses among bumblebee species poised to accelerate. Scientific Reports. 2018; 8 :14464. DOI: 10.1038/s41598-018.32665-y