 Open access peer-reviewed chapter
Open access peer-reviewed chapter
Abstract
Major histocompatibility complex (MHC) molecule is highly polymorphic. Each MHC allele has a limited capacity to bind its susceptible T cell epitopes to form an MHC-peptide complex (pMHC) and subsequently activates a compatible T cell clone. The cytotoxic T cell plays the role of eliminating virally infected cells to heal the infected patient. Overall, this article will discuss why some viral-infected patients can recover from the infected virus regardless of any treatment while others cannot. During the emergent viral epidemics, the new-coming medicines claimed to be able to treat the infected viruses. We should reconsider that some individuals, perhaps most, can recover from the infected virus with their own immunity. The fundamental comprehension of the critical role of the MHC molecule in individual immunity should be helpful to reconsider the strategy to manipulate the public health system including the notion to generate emergent viral vaccines.
Keywords
- MHC
- pMHC
- MHC restriction
- viral persistent infection
- viral vaccine
- viral epidemics
1. Introduction
Viral epidemics, such as severe respiratory syndrome (SARS), and COVID-19, have influenced our lives for a recent period of time. The infected diseases are caused by viruses classified as members of the Coronaviridae family [1]. In fact, all of the previous epidemic viruses such as the Dengue virus [2, 3], Japanese encephalitis virus [4], Influenza virus [5, 6] and many others [7, 8, 9, 10, 11] still keep causing global epidemics. Is it possible to find any effective medicines to cure the infected virus and eradicate them all? How do viruses still exist and cause epidemics? It is believed that adaptive immunity could act effectively by the role of cytotoxic T cell (Tc) to clear the viral infected cell. If so, why do the viruses keep causing problems for global public health? Perhaps, there are some things in the air that we should look for to reach our commitment to overcome these epidemics. We should reconsider our knowledge of where we are and how to move in the right direction.
Remarkably, this article will describe by differentiation of the terminology between viral invasion and viral infection. Viral invasion means our body is exposed to a particular virus which subsequently can induce our immunity with the immunogenic property of the virus. Viral infection means the virus is able to attach the cellular molecule with its receptor-binding domain and entry into the target cell. Thus, the host that does not have the compatible cellular molecules for the viral entry would not be infected but invaded. With the objective of the book and a limitation for the permitted space of the article, the details concerning the viral receptor molecules will not be discussed herein. With the questions mentioned above, this article will inform and discuss mainly the association of MHC molecules in relation to the individual’s immunity to respond to the viral agents, which come out as survival or mortal.
2. Immuno-pathogenesis of viruses
Immune cells play a significant role in eliminating foreign substances that invaded a body. For clarification, immune cells respond to foreign agents as the non-selves substance. Immune cells cannot distinguish whether foreign substances are pathogenic or not. An example is allergens which act as foreign substances and cause allergy in some people although it is not a pathogen. The symptoms are based on the reaction of the immune cells by producing some kinds of immune substances, cytokines, to cause allergenic pathogenesis in those hypersensitive individuals [12, 13]. For most of the infected viruses, there is no evidence showing that the viruses directly cause pathogenesis. As same as allergens, the pathogenesis of the virally infected individuals is also caused by the roles of various cytokines that respond to the viral agents [14, 15]. However, a virus is different from an allergen in the way that it can regenerate in the compatible host but an allergen does not. The association between the viral load and the severity of the disease has been reported [16, 17]. Hence, viral replication enhances pathogenesis by inducing extremely high amounts of cytokines which are known as cytokine storms.
Reports show that most people worldwide have been exposed to some kinds of pathogenic viruses without self-notice. Various kinds of viral antibodies such as Dengue, Japanese encephalitis, yellow fever, etc. have been found in the major populations who have never been sick with the viruses [2, 3, 4, 5, 6, 7, 8, 9, 10, 11]. World health organization (WHO) reported that 80% of the population who are positive for the Dengue serological tests are mild or asymptomatic and the mortality rate is lower than 1% [18]. For the Japanese encephalitis virus, approximately 0.4% of the seropositive individuals are severe [19]. This includes COVID-19 which approximately 70–85% of the SARS-CoV-2 positive detected are symptomless. About 10–15% are severe and require hospitalization while the mortality rate is 1–5% depending on the population and region [20, 21]. For those with severe symptoms, some can survive spontaneously without any specific treatment, while others cannot although the premium treatment has been manipulated. Aging and underlying diseases have been suggested to be the factors for the cause of severity and mortality [21, 22, 23]. However, there are reports of survival among aging and underlying patients. Thus, underlying diseases and aging might be a part of the reason but cannot be accounted as the key factor for the severity and mortality. In addition, new viral strains by the genomic mutation have also been suggested to be the reason to explain the different symptoms and severity [21, 22, 24]. However, the records show that the new viral strain, SARS CoV-2 (O-Micron) and many others still cause a similar ratio of asymptomatic, severity and mortality as the original SARS CoV-2 did. Similarly, immune evading of a virus is another mechanism to explain the cause of pathogenicity and severity including evading the previous immunity and vaccination. Questionably, if the virus evades immunity why it does not evade everybody? Why do not all vaccinated people get infected? These should be discussed.
3. Human immune response to viral antigen
After invasion into a body, the viral agent is captured by innate white blood cells such as macrophages and dendritic cells which play the role of the primary antigen-presenting cells (APCs). APCs digest and present the viral epitopes to induce adaptive immune cells which are cytotoxic T cell (Tc) and helper T cell (Th) clones in secondary lymphoid organs such as the lymph nodes and spleen [25, 26, 27]. APCs randomly cleave antigens into short peptides of 8–20 amino acid residues. The short peptide then combines with the MHC (major histocompatibility complex) molecule to form the MHC-peptide complex (pMHC) which plays a significant role to activate a specific T cell clone on its receptor so-called T cell receptor (TCR).
There are two classes of MHC molecules, class I and II. The MHC class I molecules can be expressed by any nucleated cells, while MHC class II molecules can be found only in the APCs. MHC is the key molecule to present the viral Ag on the cell surface of APC [25, 26, 27]. There are two pathways of antigen processing, the so-called endogenous [class I Ag processing] and exogenous [class II Ag processing] pathways. The endogenous pathway creates pMHC-I to activate a specific Tc cell clone [27, 28, 29]. The pMHC-I is a complex molecule of a short peptide of approximately 8–12 amino acids and MHC class I. pMHC-I is the crucial molecule to induce a specific naïve Tc cell clone to be an activated Tc. It then differentiates to effective Tc to attack the viral infected host cell. The exogenous pathway creates pMHC-II which comprises a larger epitope size, 12–20 amino acids, and MHC class II. The pMHC-II is a key to inducing the compatible Th cell clone [27, 29, 30]. The Th cell then plays the role to activate Tc to be an effective Tc.
In addition, the B lymphocyte is also one of the adaptive immune cells to play the main role extracellularly by synthesizing specific antibodies. Unlike the receptors of Tc and Th cells which recognize only short peptides, the B cell receptor (BCR) recognizes the conformational molecule of any substances which are not just proteins but also carbohydrates, lipids and nucleic acids. In other words, B lymphocyte does not need APCs to induce since it recognizes the specific epitope (B cell epitope) based on the native form of the antigen. The B cell epitope with approximately 5–20 amino acids of the native structure of the antigen can directly induce a B cell clone through the BCR [31]. After being induced, the activated B cell can synthesize IgM antibodies. Meanwhile, to differentiate to be plasma cells of each immunoglobulin class, which are IgG, IgA and IgE plasma cells, the B lymphocyte clone requires the cognate Th clone to promote [32]. During this period, the B and Th cells play the reciprocal role to support each other [31, 32, 33]. B cell which also expresses the MHC II molecule plays an antigen presentation role to the cognate Th which also sends some signals to promote B lymphocyte for differentiation to plasma cells to synthesize various classes of immunoglobulin. Without Th cells, B cells cannot produce other classes of immunoglobulin except IgM. More importantly, it cannot differentiate to be a memory B cell for prevention of the future invasion of the virus. IgM has a low affinity to bind the viral antigen and has limited action to combat the virus during extracellular existence. IgG is the highest capacity to bind the virus with its strong affinity. An affinity of IgA is the second best of IgG but plays a great role in mucosal organs which is the main route of many viral transmissions [34], including the SARS-CoV-2 virus. Accordingly, the Th cell plays the central role to maturate both B and Tc lymphocytes including the memory cells for long-term protection from the secondary viral infection [31, 32, 33].
4. The diversity of MHC molecules of human
MHC molecules are sets of molecules located on the cell membrane and are classified into 2 classes as mentioned. Each class has plural loci which are classified as the classical and non-classical loci. The MHC molecule of humans is HLA which stands for human leukocyte antigen (HLA) based on the fact that the MHC molecules were first found and studied in the white blood cell. Thus, HLA class I and II are synonyms for human MHC class I and II, respectively. The loci of classical HLA class I which have been known since 1950 are HLA-A, B and C while HLA-E, F and G were reported recently and have been called non-classical class I. In addition, the loci of classical HLA class II are HLA-DP, DQ and DR while those of the non-classical class II are HLA-DO and DM [26, 35]. The two classes of MHC molecules each have a different role in the adaptive cellular immune response. Each class of MHC genes comprises, at a minimum, three classical loci. MHC class I heterodimer is composed of alpha peptide and beta 2 microglobulins. Class I alpha peptide exhibits a high degree of polymorphism while beta 2 microglobulin does not. The two chains interact with one another non-covalently. As reported by the WHO Nomenclature Committee for Factors of the HLA System, the numbers of HLA-A, -B, and -C gene alleles and proteins are 7.644 (4.450), 9.097 (5.471), and 7.609 (4.218) × 103, respectively [36]. HLA molecules are inherited co-dominantly from the parents. Thus, each locus of the MHC genome in an individual could be either heterozygous or homozygous. A heterozygous individual has two different gene alleles, while a homozygous has the same gene allele in the locus. Accordingly, the number of gene alleles of MHC class I in any individual are limited to 3–6 gene alleles. For example, the individual who has all three loci as homozygous would have only three gene alleles, while those who have all heterozygous loci would have six gene alleles. As the MHC gene alleles are highly polymorphic, the possibility of two individuals having the same set of gene alleles would not be less than one in a million (mostly, identical twin).
MHC class II molecules are also heterodimers of alpha and beta chains, coded by A and B genes, respectively. Both alpha and beta chains of MHC class II are highly polymorphism, HLA-DPB, -DQB and -DRB genes exhibit a much higher degree of polymorphism than the HLA-DPA, -DQA, and DRA genes. Thus far, the numbers of HLA-DPB1, -DQB1, and -DRB1 gene alleles and proteins are 2.221 (1.325), 2.330 (1.455), and 4.256 (2.828) × 103, respectively while the numbers of alleles and proteins of DPA1, DQA1, and DRA are 491 (223), 508 (244), and 43 (5), respectively, in addition of a few gene alleles of DPB2, DQB2 [36]. Nevertheless, the combination of the alpha and beta peptides of HLA-II results in polymorphism more than HLA-I molecules.
5. The formation of pMHC molecule
The crystal structure of the MHC shows that each MHC molecule has a pocket to allow some of the amino acids of the peptide to fit in. As mentioned, individuals do not have more than six types for each class of MHC molecules. The studies found that each MHC variant has the ability to bind many different peptides [37, 38]. This makes MHC molecules have broad specificity to the T cell epitope presented by APCs. However, each MHC molecule can bind to only one peptide at once since there is only one cleft on an MHC molecule. To form pMHC, the MHC molecule requires only a few amino acids of T cell epitope peptide, the so-called anchor residue, for interaction. This allows each MHC allele to bind to many different peptides. Any processed peptides, which are derived from foreign substances by APCs, must contain the amino acids that can fit the MHC allele’s cleft to form pMHC [39, 40]. Subsequently, the pMHC becomes the crucial molecule to induce a specific TCR of the T cell clone although there are many other molecules involving the interaction between the T cell clone and APC. TCR requires interaction with both the peptide residue and the MHC molecule [41]. This conforms to the development of T cell clones, in the thymus, which requires passing positive selection by having the ability to work with the self-MHC alleles of the individuals [42, 43]. Noticeably, reports showed that the interaction between each MHC allele and different peptides has a different affinity. Each MHC alleles have a limitation to bind to some of the peptides [44, 45, 46]. It is unlikely that all the epitopes of the foreign peptides are able to form pMHC with a single MHC allelic molecule (Figure 1). Accordingly, the MHC allelic molecules of each person have a limitation to forming pMHC with some of the peptides if those peptides do not contain the anchor residues that are compatible with the individuals’ MHC alleles. There are studies showed that individuals who are MHC homozygous are more susceptible to pathogens than those who are heterozygous [47, 48, 49]. This explains that people who have fewer types of MHC alleles might have limitations to form pMHC molecules with some pathogen to induce naïve T cell clones. A good match of the peptide and the peptide-binding groove of the MHC molecule is a crucial factor in the induction of T-cell clones. Thus, the availability of the MHC allele and the antigen are the key MHC restriction factors for the induction of compatible T-cell clones. Besides the possibility of viral variants, a lack of available MHC alleles and antigens might explain why some individuals become infected and do not respond efficiently to gain seroprotection after viral vaccination [50, 51]. Therefore, the invasion of any particular antigen of a virus into different individuals does not guarantee the induction of the same level of immunity because of the limited varieties of MHC alleles in each person.
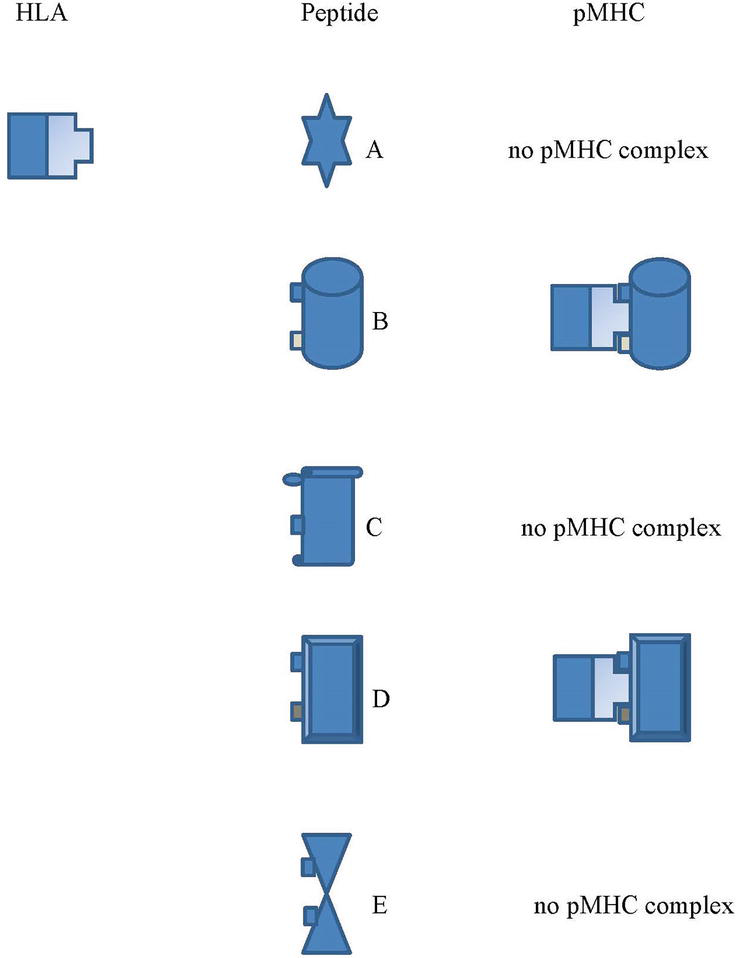
Figure 1.
In the illustration for the process to form the pMHC complex, the anchor residues of the epitope peptide must be compatible with the groove of the MHC allelic molecule to form pMHC. The HLA allele can form a pMHC complex to the peptides B and D which subsequently induce the TCR of each specific T cell clone but cannot do with the peptides A, C and E.
6. Immune evading versus chronic viral infection
Symptomatic treatment is a major strategy for treating most viral-infected patients until adaptive immunity is induced for the effective elimination of the viral agent. Usually, patients can recover from acute viral infections if their immune responses are produced within a specific period. On average, this requires one to two weeks. However, most viruses cause not only acute but also chronic infections. The definition of chronic viral infection is that the infected virus persists within a host for a longer period, usually longer than six months. During chronic infection, individuals might be asymptomatic but can transmit the virus to others [52, 53]. There are two significant types of chronic viral infections: latent and persistent. Latent infections are caused by specific viruses that can evade the host’s immunity after causing pathogenesis during the primary infection. Examples include members of Herpesviridae, such as herpes simplex and varicella-zoster [54, 55]. HSV produces latency-associated transcript (LAT) to inhibit cellular apoptosis to avoid the host’s immunity. During this time, the virus can keep being dormant in its infected cell without any clinical symptoms to the host [56, 57, 58]. Besides the role of LAT, there were reports that herpes viruses also have a mechanism to interfere with the MHC class I expression of the target host cells [59]. Actually, this mechanism has been found in many other viruses as well. It explains that cytotoxic T cells [Tc] cannot recognize and eliminate the viral infected cell [60]. After acute HSV infection, the virus remains latently infected in all of the infected individuals who have not been treated properly during the primary acute infection. The viruses migrate to preferred cells or organs (such as nerve cells) where the host’s immunity cannot respond. Acyclovir has been reported to be an effective antiviral drug to treat acute HSV infection to avoid chronically latent infection [61]. Accordingly, after acute HSV infection, the virus remains latently infected in the infected individuals who have not been treated or have not been treated properly during the primary acute infection. Eventually, patients usually experience latent infection [61, 62]. Thus, the mechanism of chronic infection of HSV should be accounted as immune evading which the mechanism seems to be explainable to all of the infected individuals and accounted as the viral factor.
The other chronic viral infection is viral persistence. The chronically persistent infection could be found in both DNA and RNA viruses. RNA viruses are much higher in their genomic mutation according to the low efficacy of their RNA polymerase to proofread their genomic replication [63, 64]. HCV, an RNA virus, is prone to cause a high prevalence of chronic infection. WHO reported that only about 30% (15–45%) of HCV-infected individuals have immune clearance within 6 months without any treatment. Of the rest of the HCV-infected individuals, approximately 70% (55–85%), developed a chronic infection [65]. However, many other RNA viruses have been reported to have a much lower prevalence of chronic infection e.g. Ebola [66], Influenza virus [67], Sars-Cov-2 [68], and other RNA viruses [69, 70, 71, 72]. In addition, there were some reports showing that HCV mutation might not be the only cause of viral persistency [73, 74]. Accordingly, the genomic mutation of the virus might not be concluded as the only genuine cause of chronically persistent infection. HBV, a DNA virus, has been reported to cause chronic infection in approximately 10–15% of HBV-infected individuals [65]. Chronically infected HBV and HCV individuals tend to develop liver cirrhosis and hepatocellular carcinoma [75, 76]. Similar to Herpes viruses, HBV and HCV have also been reported to interfere with the MHC class I expression which was claimed to be the cause of their persistence [77].
Questionably, why do all the HBV and HCV-infected individuals not become chronically infected? As mentioned, approximately 85–90% of the HBV infected and 15–45% of the HCV infected can spontaneously clear the viral agent. As an example, the HBV and HCV persistently infected might not be explainable to be caused just by the virus factor as it does for the Herpes viruses. Like many other viruses including HSV, there are reports that HBV and HCV can interfere with MHC expression. In contrast to latent viral infection, chronically persistent viral infections vary by individual [69, 78, 79, 80, 81]. Notably, the persistent viral agent continues to exist in the target organ of infected individuals without any major symptoms. Obviously, the infected host cannot clear the virus, and the infection becomes persistent. This raises the question of why chronic viral persistence cannot be found in all HBV and HCV-infected individuals as HSV does. Besides HBV and HCV, persistent viral infections can also be reported in many other viruses [82, 83]. Should one say that a virus factor such as MHC interference of viruses is the only factor to cause chronic viral infection? Should a host factor based on various genetics of individuals be the crucial key to explaining the cause of chronic viral persistent infections?
Accordingly, there are at least two major approaches to explain the cause of chronic viral infection. Firstly, it is a virus factor. The virus has a kind of mechanism to avoid host immunity. This kind of chronic infection should be the same in every individual as in the case of Herpesvirus which could be accounted for as the immune evading of the virus. The other approach is the different efficiency of the individual’s immunity to fight against any particular infectious virus. In the latter case, some of the virally infected individuals can eventually clear the viral agents while others cannot and become persistently infected. This is expected to relate to the host factor, which is different and based on the individual’s genetics. Thus, the polymorphic of MHC molecules is purposed to be the major factor herein.
7. The association of the MHC molecule and persistent viral infection
Viral persistent infection can also be found in a low-evolved immune animal. Insects, which lack adaptive immune response, many of which become viral carriers of Arboviruses, such as dengue hemorrhagic fever [84], Japanese encephalitis virus [85], West Nile virus [86], and others [69, 78, 79, 80, 81]. The bee, which is an economic insect, can also be persistently infected with its pathologic viruses [87, 88]. Another example of viral persistent infection in low-evolved immune animals is penaeid shrimp which is a vital farming commodity in many countries. Thus, there are many reports concerning the viral epidemics in penaeid shrimp in addition to viral persistence.
Over two decades ago, the white spot virus and yellow head virus each caused pathogenesis and high mortality in infected shrimps when they emerged [89, 90, 91]. Notably, there is evidence of persistent viral infection in the shrimp, mortality declined sharply after several epidemic years once pond management systems such as water quality, feeding system, temperature, and rearing population size were optimized. In poorly managed farms, high mortality was reported [89, 91]. This could explain why shrimps perform a kind of unknown mechanism to tolerate infectious viruses. Alternatively, this might also be explained by the incidence of genomic mutation of the viruses which lowers their pathogenesis. The previous study, however, reported that the naïve shrimp showed acute infection and high mortality when challenged by a virus that was isolated from occluded, persistent-infected shrimp [89]. Accordingly, the viral genomic mutation should be excluded from the explanation for the cause of the persistent infection in shrimps. Insects and shrimps possess native immunity but not adaptive immunity. Naturally, innate immunity is not as sufficiently effective in clearing the virus and virus-infected cells as adaptive Tc.
In humans, there were reports showing the association of HLA variants and viral persistent infection. Bhaskaran et al. [92] reported that HLA-B*44 and DRB1*07 had a significant association with persistent HPV-16 infection (odds ratio, p-value = 26.3, 0.03 and 4.7, 0.01, respectively). HLA-B*27 and DRB1*12 were significantly associated with both HPV-16+ cervical cancer (CaCx) and persistent HPV-16 infection (23.8, 0.03; 52.9, 0.01; 9.8, 0.0009; and 13.8, 0.009; respectively). HLA-B*15 showed a negative association with HPV-16-positive CaCx (0.1, 0.01), whereas DRB1*04 exhibited protection to both HPV-16-positive CaCx and persistent HPV-16 infection (0.3, 0.0001 and 0.1, 0.0002, respectively). Besides, the associations of HLA variants and viral persistent infection of HBV [93, 94, 95] and HCV [96, 97, 98] have also been reported. These previous studies support the association between the HLA variants and the cause of viral persistent infection in some individuals which should be intended to evaluate the crucial role of MHC molecules concerning viral epidemics from the viral persistent carriers.
As mentioned, the host’s immunity requires T cells, especially Tc and Th cells, for the effective clearance of the viral agent. MHC restriction is a key to activating T cell clones through their TCR. The antigenic epitope of the viral agent requires an MHC molecule to form the pMHC at the MHC groove and induce the compatible T lymphocyte clone. Each of the MHC alleles expresses different forms of the grooves [99, 100]. The limited capacity of the MHC groove to bind the viral short peptides results in lacking the appropriate pMHC to activate the significant T cell clone. In other words, the affinity differs between each MHC variant, and the distinct antigenic epitopes subsequently result in varying levels of the immune response. Hence, it assumes that persistently infected patients cannot clear the viral infected cell because they cannot produce an effective Tc cell for viral clearance. As is the case in infected shrimps, persistently infected patients do not have the adaptive immunity of the cellular-mediated immune response (CMIR). This could be due to the lack of an appropriate MHC-I allelic molecule to interact with the viral epitope, which results in the pMHC not forming to induce appropriate Tc lymphocyte clones. In addition, it is also interesting to raise the question of why the persistent virus can live in a host without any pathogenesis.
Besides activating, cytotoxic T cell also requires induction of Th for differentiation to an effective Tc for the efficient clearance of the infected cell, in addition, to producing memory cells for long-term activity. Thus, the compatible MHC-II is also an additional molecule to clear the virally infected cells. Individuals who lack either compatible MHC-I or II or both with the crucial viral epitopes cannot clear the viral infected cell efficiently. These infected patients might become severe cases unless the virus compromises to live in the host comfortably, as same as the infected shrimps that live in the optimal environment and come out asymptomatically [3, 100, 101]. The mechanism to compromise might be a process of adaptation of both virus and host, in addition to an environment. This subject has been discussed in many different aspects which require further study [102, 103, 104].
8. Perspective to increase the effectiveness of viral vaccines
As mentioned above, the MHC molecules play an important role in the immune response by forming the pMHC I and pMHC II complex molecule to activate Tc and Th cells, respectively, through the action of APC-containing cell clusters. Given that MHC class I and II molecules have many different alleles [36], while each of us has only 3–6 MHC alleles in each class, this is a limited binding to various epitopes. Therefore, some clones of T cell clones were not induced due to the lack of compatible pMHC [47, 48, 49]. This likely explains why a number of people do not have effective immunity after vaccination against HBV [9, 10, 11, 12, 13, 14] and many other viruses [1, 2, 3, 4, 5, 6, 7, 8]. It could be assumed that those vaccinated individuals lack the appropriate MHC allele to form the compatible pMHC complex with the vaccine epitopes. Thus, the required T cell clones could not be induced.
Th cells are important immune cells in helping to achieve the functioning of both Tc and B lymphocytes. Without a suitable Th cell cloning known as cognate Th cell, the activated B lymphocyte produces only IgM antibody, which has low stability and lower efficiency than IgG and IgA. Cognate Th cells can induce specific B cell clones to develop into plasma cells (antibody-secreting B cell) to produce other antibody classes (IgG, IgA, IgE) and memory B lymphocytes. For viral protection, antibodies are more important in preventing viral infection than Tc lymphocytes because the antibody can bind and inhibit the viral particles from binding to the viral receptor molecule, preventing the virus from entering the target cell and multiplying. This is known as antibody neutralization. Tc cells become active only after the target cell has been invaded by the virus since the recognition of the Tc cell clone through its TCR requires pMHC derived from virus-infected target cells before Tc cells can be manipulated. Therefore, the induction of memory B cells is an important goal of viral vaccines that the body’s immune system must produce. Accordingly, viral vaccines should contain epitopes capable of binding to the MHC II alleles of individuals to form a pMHC-II as a capable molecule for inducing a suitable Th cell cognate clone. All these reasons suggest that any particular subunit viral vaccine may not be available to every community or the entire world. This is because viral vaccines should include an epitope appropriate for the type of MHC alleles in each person or population. Although the production of subunit viral vaccines is convenient with various technologies, especially with molecular genetic techniques, it raises problems and questions concerning the effectiveness of any particular vaccine for all global populations.
In the past, with much lesser knowledge concerning immunology, Dr. Edward Jenner and his associates had a way to initiate a successful battle against an epidemic of smallpox virus during the 18th century by the term of vaccination [105, 106]. Up till now, it seems we could not find any better outcome although a lot of medical scientific knowledge, including immunology, has been discovered. There are not sufficient kinds of viral vaccines to prevent the infection of many classic and emerging viruses. With a thought of safety reasons to prevent side effects, most of the scientists decided to work on subunit viral vaccines. This is different from the time when Edward Jenner used a whole particle of the Cowpox virus to vaccinate people to prevent the smallpox viral epidemic. On a different aspect, if the epitopes of any viral vaccines were cut down, an antigen-presenting cell would not be able to process induction to some particular helper T cell clones. Subsequently, the particular B cell clones cannot synthesize those particular antibodies to neutralize the infectious viral particles. Thus, could this be a reason that the subunit viral vaccines do not work in some people in the general public?
Some viral vaccines might show to be successful in the laboratory because the experimental animals were inbred to some specific strains. The viral vaccines that gave positive results at the experimental level do not mean it can work in human populations that contain various polymorphic MHC alleles. In addition, the viral vaccines that work in one population do not mean they always work in other populations. Perhaps, the subunit vaccine should be derived from all over the viral epitopes as if the wild-type virus. Of course, it is more appropriate to vaccinate people to gain viral protection on the receptor-binding domain (RBD). However, people who do not have the appropriate MHC alleles could not form the appropriate pMHC for the RBD which is the best target. Perhaps, their MHC’s alleles might be able to form pMHC with other parts of the virus. This could allow them to synthesize memory B cells for some other parts of the virus. The memory B cell of other viral components might perform some actions to attack the infected virus. The combination of the antibody and natural killer (NK) cell by the antibody-dependent cell cytotoxicity (ADCC) mechanism could be helpful. This will be similar to Dr. Janner’s manipulation and the manufacturing should not be too hard as we would produce the wild-type viral vaccines. This purpose should be considered for further study.
9. Conclusion
Some previous studies are convinced that chronically viral persistent infection is caused by viral factors such as viral mutation and some of the viral mechanisms. This article proposes that the cause of the chronic viral persistent infection should, also, associate with the existence of the MHC variants of the individual which is a host factor. Viral clearance requires the compatible MHC variants to induce Tc and Th for the effective eradication of the viral infected cell. There is no evidence that native immunity, such as natural killer cells and macrophages can clear the infectious virus efficiently by itself. The existence of a compatible MHC-I allele to the viral epitope is necessary for inducing the appropriate Tc clone to clear all the virally infected cells. The main fact is that the Tc clone [s] also requires compatible Th to support its efficiency. Thus, besides the MHC-I variants, the existence of the compatible MHC-II allele molecules is also another key to clearing the viral infected cell. The MHC molecules, as a key to preventing and restraining a viral epidemic, have been underestimated for their crucial role to eradicate the infected viral agent. Thus, this requires a better understanding of the distribution of MHC alleles in our global community. In addition, the Th cell is also a keystone for the regulation of immunoglobulin gene switching to synthesize more effective immunoglobulin which are IgG and IgA, not just IgM. More importantly, the Th cell is required for the B cell differentiation to be a memory B cell which makes viral prevention sustainable. Accordingly, the availability of the appropriate MHC class I manipulates the host to clear the viral infected cell effectively by the role of Tc. In addition, MHC class II is a key for the induction of the appropriate Th-cell clones to make adaptive immunity work properly in terms of prevention and clearance of the viral agent.
If the natural selection theory, proposed by Charles Darwin in 1858, influences creatures and human beings, the MHC gene could be one of the answers and become the target for us to overcome. Individuals have distinguished MHC allelic molecules which make each person respond to the viral agent differently. Any particular viral vaccines especially subunit viral vaccines do not seem to be available to prevent viral transmission. For viral treatment, it should have further studies to determine the MHC alleles for their association with the recovery of the viral infected patients. Using antiviral drugs, especially COVID-19 requires more careful evaluation. Those infected patients might be able to recover by their own immunity which relates to their MHC alleles to process Tc to clear the viral infected cell, not the medicine. If the medicine is truly effective to cure viral infected diseases why cannot it help many of the COVID-19 who do not have any other underlying diseases? Why is the mortality ratio of the COVID-19 epidemic still more or less the same although new variants are being found all the time? This needs an answer.
Acknowledgments
The author would like to appreciate Mr. Rohn Meyer for his kindness in helping proofread this article.
References
- 1.
Leao JC, Gusmao TPL, Zarzar AM, Leao Filho JC, Santos B, de Faria A, et al. Coronaviridae-old friends, new enemy! Oral Diseases. 2022; 28 (Suppl. 1):858-866. DOI: 10.1111/odi.13447. Epub 2020 Jun 26 - 2.
Luo S, Cui W, Li C, et al. Seroprevalence of dengue IgG antibodies in symptomatic and asymptomatic individuals three years after an outbreak in Zhejiang Province, China. BMC Infectious Diseases. 2018; 18 (1):92 - 3.
Ly S, Fortas C, Duong V, Benmarhnia T, et al. Asymptomatic dengue virus infections, Cambodia, 2012-2013. Emerging Infectious Diseases. 2019; 25 (7):1354-1362 - 4.
Nealon J, Taurel AF, Yoksan S, et al. Serological evidence of Japanese encephalitis virus circulation in Asian children from dengue-endemic countries. The Journal of Infectious Diseases. 2019; 219 (3):375-381 - 5.
Ip DK, Lau LL, Leung NH, et al. Viral shedding and transmission potential of asymptomatic and paucisymptomatic influenza virus infections in the community. Clinical Infectious Diseases. 2017; 64 (6):736-742 - 6.
Melchior TB, Perosa AH, Camargo CN, Granato C, Bellei N. Influenza virus prevalence in asymptomatic and symptomatic subjects during pandemic and postpandemic periods. American Journal of Infection Control. 2015; 43 (5):460-464 - 7.
Galanti M, Birger R, Ud-Dean M, et al. Rates of asymptomatic respiratory virus infection across age groups. Epidemiology and Infection. 2019; 147 :e176 - 8.
Attaran MS, Hosseini SM, Fakhari J, Sharifi Z. Serological and molecular characterization of hepatitis B virus in asymptomatic blood donors in Iran. Iranian Journal of Microbiology. 2018; 10 (1):59-64 - 9.
Glynn JR, Bower H, Johnson S, et al. Asymptomatic infection and unrecognised Ebola virus disease in Ebola-affected households in Sierra Leone: A cross-sectional study using a new non-invasive assay for antibodies to Ebola virus. The Lancet Infectious Diseases. 2017; 17 (6):645-653 - 10.
Stoszek SK, Engle RE, Abdel-Hamid M, Mikhail N, Abdel-Aziz F, Medhat A, et al. Hepatitis E antibody seroconversion without disease in highly endemic rural Egyptian communities. Transactions of the Royal Society of Tropical Medicine and Hygiene. 2006; 100 (2):89-94 - 11.
Sejvar JJ. West Nile Virus infection. Microbiology Spectrum. 2016; 4 (3). DOI: 10.1128/microbiolspec.EI10-0021-2016 - 12.
Rogala B, Bozek A, Gluck J, Jarzab J. Prevalence of IgE-mediated allergy and evaluation of Th1/Th2 cytokine profiles in patients with severe bronchial asthma. Postępy Dermatologii i Alergologii. 2015; 32 (4):274-280. DOI: 10.5114/pdia.2015.53323. Epub 2015 Aug 12 - 13.
Pelikan Z. Cytokine profiles in tears accompanying the secondary conjunctival responses induced by nasal allergy. Current Eye Research. 2014; 39 (2):120-132. DOI: 10.3109/02713683.2013.836542. Epub 2013 Oct 21 - 14.
Oldstone MB, Rosen H. Cytokine storm plays a direct role in the morbidity and mortality from influenza virus infection and is chemically treatable with a single sphingosine-1-phosphate agonist molecule. Current Topics in Microbiology and Immunology. 2014; 378 :129-147. DOI: 10.1007/978-3-319-05879-5_6 - 15.
Yuan S, Jiang SC, Zhang ZW, Fu YF, Hu J, Li ZL. Quantification of cytokine storms during virus infections. Frontiers in Immunology. 2021; 12 :659419. DOI: 10.3389/fimmu.2021.659419 - 16.
Kwon JS, Kim JY, Kim MC, Park SY, Kim BN, Bae S, et al. Factors of severity in patients with COVID-19: Cytokine/chemokine concentrations, viral load, and antibody responses. The American Journal of Tropical Medicine and Hygiene. 2020; 103 (6):2412-2418. DOI: 10.4269/ajtmh.20-1110. Epub 2020 Oct 27 - 17.
Nakamura S, Iwanaga N, Hara S, Shimada S, Kashima Y, Hayasaka D, et al. Viral load and inflammatory cytokine dynamics associated with the prognosis of severe fever with thrombocytopenia syndrome virus infection: An autopsy case. Journal of Infection and Chemotherapy. 2019; 25 (6):480-484. DOI: 10.1016/j.jiac.2019.01.013. Epub 2019 Feb 26 - 18.
WHO. Dengue and Severe Dengue. 2022. Available from: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue - 19.
WHO. Japenese Encephalitis. 2019. Available from: https://www.who.int/news-room/fact-sheets/detail/japanese-encephalitis - 20.
WHO. 2022. Available from: https://covid19.who.int/?adgroupsurvey={adgroupsurvey}&gclid =Cj0KCQiAsd KbBhDHARIsANJ6-jdtyWU-UD_ZYkm0yVwxk-PEepbZ673LOJt4F6i6ceuMQbjgLTM7JhgaAkKwEALw_wcB - 21.
Salzberger B, Buder F, Lampl B, Ehrenstein B, Hitzenbichler F, Holzmann T, et al. Epidemiology of SARS-CoV-2. Infection. 2021; 49 (2):233-239. DOI: 10.1007/s15010-020-01531-3. Epub 2020 Oct 8 - 22.
Williams N, Radia T, Harman K, Agrawal P, Cook J, Gupta A. COVID-19 severe acute respiratory syndrome coronavirus 2 [SARS-CoV-2] infection in children and adolescents: A systematic review of critically unwell children and the association with underlying comorbidities. European Journal of Pediatrics. 2021; 180 (3):689-697. DOI: 10.1007/s00431-020-03801-6. Epub 2020 Sep 10 - 23.
Bartleson JM, Radenkovic D, Covarrubias AJ, Furman D, Winer DA, Verdin E. SARS-CoV-2, COVID-19 and the ageing immune system. Nature Aging. 2021; 1 (9):769-782. DOI: 10.1038/s43587-021-00114-7. Epub 2021 Sep 14 - 24.
Meo SA, Meo AS, Al-Jassir FF, Klonoff DC. Omicron SARS-CoV-2 new variant: Global prevalence and biological and clinical characteristics. European Review for Medical and Pharmacological Sciences. 2021; 25 (24):8012-8018. DOI: 10.26355/eurrev_202112_27652 - 25.
Hume DA. Macrophages as APC and the dendritic cell myth. Journal of Immunology. 2008; 181 (9):5829-5835. DOI: 10.4049/jimmunol.181.9.5829 - 26.
Kelly A, Trowsdale J. Genetics of antigen processing and presentation. Immunogenetics. 2019; 71 (3):161-170. DOI: 10.1007/s00251-018-1082-2. Epub 2018 Sep 13 - 27.
Momburg F, Hengel H. Corking the bottleneck: The transporter associated with antigen processing as a target for immune subversion by viruses. Current Topics in Microbiology and Immunology. 2002; 269 :57-74. DOI: 10.1007/978-3-642-59421-2_4 - 28.
Lu X, Gibbs JS, Hickman HD, David A, Dolan BP, Jin Y, et al. Endogenous viral antigen processing generates peptide-specific MHC class I cell-surface clusters. Proceedings of the National Academy of Sciences of the United States of America. 2012; 109 (38):15407-15412. DOI: 10.1073/pnas.1208696109. Epub 2012 Sep 4 - 29.
Shastri N, Schwab S, Serwold T. Producing nature’s gene-chips: The generation of peptides for display by MHC class I molecules. Annual Review of Immunology. 2002; 20 :463-493. DOI: 10.1146/annurev.immunol.20.100301.064819. Epub 2001 Oct 4 - 30.
Lázaro S, Gamarra D, Del Val M. Proteolytic enzymes involved in MHC class I antigen processing: A guerrilla army that partners with the proteasome. Molecular Immunology. 2015; 68 (2 Pt A):72-76. DOI: 10.1016/j.molimm.2015.04.014. Epub 2015 May 23 - 31.
Roche PA, Furuta K. The ins and outs of MHC class II-mediated antigen processing and presentation. Nature Reviews. Immunology. 2015; 15 (4):203-216. DOI: 10.1038/nri3818. Epub 2015 Feb 27 - 32.
Murin CD, Wilson IA, Ward AB. Antibody responses to viral infections: A structural perspective across three different enveloped viruses. Nature Microbiology. 2019; 4 (5):734-747. DOI: 10.1038/s41564-019-0392-y. Epub 2019 Mar 18 - 33.
Berzofsky JA. T-B reciprocity. An Ia-restricted epitope-specific circuit regulating T cell-B cell interaction and antibody specificity. Survey of Immunologic Research. 1983; 2 (3):223-229. DOI: 10.1007/BF02918417 - 34.
McHeyzer-Williams LJ, Malherbe LP, McHeyzer-Williams MG. Checkpoints in memory B-cell evolution. Immunological Reviews. 2006; 211 :255-268. DOI: 10.1111/j.0105-2896.2006.00397.x - 35.
Rock KL, Reits E, Neefjes J. Present yourself! By MHC class I and MHC class II molecules. Trends in Immunology. 2016; 37 (11):724-737. DOI: 10.1016/j.it.2016.08.010. Epub 2016 Sep 7 - 36.
Marsh SGE. Nomenclature for factors of the HLA system, update July, August and September 2021. HLA. 2021; 98 (6):579-618. DOI: 10.1111/tan.14464 - 37.
Marzella DF, Parizi FM, van Tilborg D, Renaud N, Sybrandi D, Buzatu R, et al. PANDORA: A fast, anchor-restrained modelling protocol for peptide: MHC complexes. Frontiers in Immunology. 2022; 13 :878762. DOI: 10.3389/fimmu.2022.878762 - 38.
Perez MAS, Cuendet MA, Röhrig UF, Michielin O, Zoete V. Structural prediction of peptide-MHC binding modes. Methods in Molecular Biology. 2022; 2405 :245-282. DOI: 10.1007/978-1-0716-1855-4_13 - 39.
Rammensee HG. Chemistry of peptides associated with MHC class I and class II molecules. Current Opinion in Immunology. 1995; 7 (1):85-96. DOI: 10.1016/0952-7915[95]80033-6 - 40.
Nielsen M, Lundegaard C, Lund O. Prediction of MHC class II binding affinity using SMM-align, a novel stabilization matrix alignment method. BMC Bioinformatics. 2007; 8 :238. DOI: 10.1186/1471-2105-8-238 - 41.
Szeto C, Lobos CA, Nguyen AT, Gras S. TCR recognition of peptide-MHC-I: Rule makers and breakers. International Journal of Molecular Sciences. 2020; 22 (1):68. DOI: 10.3390/ijms22010068 - 42.
Lucas B, McCarthy NI, Baik S, Cosway E, James KD, Parnell SM, et al. Control of the thymic medulla and its influence on αβT-cell development. Immunological Reviews. 2016; 271 (1):23-37. DOI: 10.1111/imr.12406 - 43.
Tinsley KW, Hong C, Luckey MA, Park JY, Kim GY, Yoon HW, et al. Ikaros is required to survive positive selection and to maintain clonal diversity during T-cell development in the thymus. Blood. 2013; 122 (14):2358-2368. DOI: 10.1182/blood-2012-12-472076. Epub 2013 Aug 1 - 44.
Margulies DH, Corr M, Boyd LF, Khilko SN. MHC class I/peptide interactions: Binding specificity and kinetics. Journal of Molecular Recognition. 1993; 6 (2):59-69. DOI: 10.1002/jmr.300060204 - 45.
Sinigaglia F, Hammer J. Defining rules for the peptide-MHC class II interaction. Current Opinion in Immunology. 1994; 6 (1):52-56. DOI: 10.1016/0952-7915[94]90033-7 - 46.
Jensen KK, Andreatta M, Marcatili P, Buus S, Greenbaum JA, Yan Z, et al. Improved methods for predicting peptide binding affinity to MHC class II molecules. Immunology. 2018; 154 (3):394-406. DOI: 10.1111/imm.12889. Epub 2018 Feb 6 - 47.
Lipsitch M, Bergstrom CT, Antia R. Effect of human leukocyte antigen heterozygosity on infectious disease outcome: The need for allele-specific measures. BMC Medical Genetics. 2003; 4 :2. DOI: 10.1186/1471-2350-4-2. Epub 2003 Jan 24 - 48.
Arora J, Pierini F, McLaren PJ, Carrington M, Fellay J, Lenz TL. HLA heterozygote advantage against HIV-1 is driven by quantitative and qualitative differences in HLA allele-specific peptide presentation. Molecular Biology and Evolution. 2020; 37 (3):639-650. DOI: 10.1093/molbev/msz249 - 49.
Carrington M, Nelson GW, Martin MP, Kissner T, Vlahov D, Goedert JJ, et al. HLA and HIV-1: Heterozygote advantage and B*35-Cw*04 disadvantage. Science. 1999; 283 (5408):1748-1752. DOI: 10.1126/science.283.5408.1748 - 50.
Faneye AO, Adeniji JA, Olusola BA, Motayo BO, Akintunde GB. Measles virus infection among vaccinated and unvaccinated children in Nigeria. Viral Immunology. 2015; 28 (6):304-308. DOI: 10.1089/vim.2014.0118. Epub 2015 Jun 23 - 51.
Smith LL, Milstone AM, Jibowu M, Luo CH, Morris CP, Mostafa HH, et al. Transmission of severe acute respiratory coronavirus virus 2 (SARS-CoV-2), delta variant, between two fully vaccinated healthcare personnel. Infection Control and Hospital Epidemiology. Dec 2022; 43 (12):1983-1985. DOI: 10.1017/ice.2021.469. Epub: 2021 Nov 8. PMID: 34743764; PMCID: PMC8632451 - 52.
Sharma SK, Saini N, Chwla Y. Hepatitis B virus: Inactive carriers. Virology Journal. 2005; 2 :82. DOI: 10.1186/1743-422X-2-82 - 53.
Furuya-Kanamori L, Yakob L. Heterogeneous and dynamic prevalence of asymptomatic influenza virus infections. Emerging Infectious Diseases. 2018; 24 (5):951. DOI: 10.3201/eid2405.180075 - 54.
Bello-Morales R, López-Guerrero JA. Extracellular vesicles in herpes viral spread and immune evasion. Frontiers in Microbiology. 2018; 9 :2572. DOI: 10.3389/fmicb.2018.02572 - 55.
Abendroth A, Kinchington PR, Slobedman B. Varicella zoster virus immune evasion strategies. Current Topics in Microbiology and Immunology. 2010; 342 :155-171. DOI: 10.1007/82_2010_41 - 56.
Carpenter D, Hsiang C, Jiang X, Osorio N, BenMohamed L, Jones C, et al. The herpes simplex virus type 1 [HSV-1] latency-associated transcript [LAT] protects cells against cold-shock-induced apoptosis by maintaining phosphorylation of protein kinase B [AKT]. Journal of Neurovirology. 2015; 21 (5):568-575. DOI: 10.1007/s13365-015-0361-z. Epub 2015 Jun 13 - 57.
Jaggi U, Matundan HH, Tormanen K, Wang S, Yu J, Mott KR, et al. Expression of murine CD80 by herpes simplex Virus 1 in place of latency-associated transcript [LAT] can compensate for latency reactivation and anti-apoptotic functions of LAT. Journal of Virology. 2020; 94 (6):e01798-e01719. DOI: 10.1128/JVI.01798-19 - 58.
Imai Y, Apakupakul K, Krause PR, Halford WP, Margolis TP. Investigation of the mechanism by which herpes simplex virus type 1 LAT sequences modulate preferential establishment of latent infection in mouse trigeminal ganglia. Journal of Virology. 2009; 83 (16):7873-7882. DOI: 10.1128/JVI.00043-09. Epub 2009 Jun 3 - 59.
Piguet V. Receptor modulation in viral replication: HIV, HSV, HHV-8 and HPV: Same goal, different techniques to interfere with MHC-I antigen presentation. Current Topics in Microbiology and Immunology. 2005; 285 :199-217. DOI: 10.1007/3-540-26764-6_7 - 60.
Stone JD, Aggen DH, Chervin AS, Narayanan S, Schmitt TM, Greenberg PD, et al. Opposite effects of endogenous peptide-MHC class I on T cell activity in the presence and absence of CD8. Journal of Immunology. 2011; 186 (9):5193-5200. DOI: 10.4049/jimmunol.1003755. Epub 2011 Mar 30 - 61.
Kłysik K, Pietraszek A, Karewicz A, Nowakowska M. Acyclovir in the treatment of herpes viruses—A review. Current Medicinal Chemistry. 2020; 27 (24):4118-4137. DOI: 10.2174/0929867325666180309105519 - 62.
Menendez CM, Carr DJJ. Defining nervous system susceptibility during acute and latent herpes simplex virus-1 infection. Journal of Neuroimmunology. 2017; 308 :43-49. DOI: 10.1016/j.jneuroim.2017.02.020. Epub 2017 Mar 8 - 63.
Duffy S. Why are RNA virus mutation rates so damn high? PLoS Biology. 2018; 16 (8):e3000003. DOI: 10.1371/journal.pbio.3000003 - 64.
Malpica JM, Fraile A, Moreno I, Obies CI, Drake JW, García-Arenal F. The rate and character of spontaneous mutation in an RNA virus. Genetics. 2002; 162 (4):1505-1511. DOI: 10.1093/genetics/162.4.1505 - 65.
WHO. Hepatitis C. 2020. Available from: https://www.who.int/news-room/fact-sheets/detail/hepatitis-c - 66.
Heeney JL. Ebola: Hidden reservoirs. Nature. 2015; 527 (7579):453-455. DOI: 10.1038/527453a - 67.
Morens DM, Taubenberger JK, Fauci AS. The persistent legacy of the 1918 influenza virus. New England Journal of Medicine. 2009; 361 (3):225-229. DOI: 10.1056/NEJMp0904819. Epub 2009 Jun 29. Erratum in: New England Journal of Medicine. 2009;361 (11):1123 - 68.
Kang H, Wang Y, Tong Z, Liu X. Retest positive for SARS-CoV-2 RNA of “recovered” patients with COVID-19: Persistence, sampling issues, or re-infection? Journal of Medical Virology. 2020; 92 (11):2263-2265. DOI: 10.1002/jmv.26114. Epub 2020 Jun 9 - 69.
Doi T, Kwon HJ, Honda T, Sato H, Yoneda M, Kai C. Measles virus induces persistent infection by autoregulation of viral replication. Scientific Reports. 2016; 6 :37163. DOI: 10.1038/srep37163 - 70.
Hoarau JJ, Jaffar Bandjee MC, Krejbich Trotot P, Das T, Li-Pat-Yuen G, Dassa B, et al. Persistent chronic inflammation and infection by chikungunya arthritogenic alphavirus in spite of a robust host immune response. Journal of Immunology. 2010; 184 (10):5914-5927. DOI: 10.4049/jimmunol.0900255. Epub 2010 Apr 19 - 71.
Lin WH, Kouyos RD, Adams RJ, Grenfell BT, Griffin DE. Prolonged persistence of measles virus RNA is characteristic of primary infection dynamics. Proceedings of the National Academy of Sciences of the United States of America. 2012; 109 (37):14989-14994. DOI: 10.1073/pnas.1211138109. Epub 2012 Aug 7 - 72.
Meiring T, Prozesky L, Du Preez ER, Verwoerd DJ. The diagnosis and prevalence of persistent infection with bovine viral diarrhoea virus in south African feedlot cattle. The Onderstepoort Journal of Veterinary Research. 2011; 78 (1):323. DOI: 10.4102/ojvr.v78i1.323 - 73.
Fuller MJ, Shoukry NH, Gushima T, Bowen DG, Callendret B, Campbell KJ, et al. Selection-driven immune escape is not a significant factor in the failure of CD4 T cell responses in persistent hepatitis C virus infection. Hepatology. 2010; 51 (2):378-387. DOI: 10.1002/hep.23319 - 74.
Lapa D, Garbuglia AR, Capobianchi MR, Del Porto P. Hepatitis C virus genetic variability, human immune response, and genome polymorphisms: Which is the interplay? Cell. 2019; 8 (4):305. DOI: 10.3390/cells8040305 - 75.
Zhao SN, Liu LL, Lv ZP, Wang XH, Wang CH. Network analysis of HBV- and HCV-induced hepatocellular carcinoma based on Random Forest and Monte Carlo cross-validation. Molecular Medicine Reports. 2017; 16 (3):2411-2416. DOI: 10.3892/mmr.2017.6861. Epub 2017 Jun 27 - 76.
Urabe Y, Ochi H, Kato N, Kumar V, Takahashi A, Muroyama R, et al. A genome-wide association study of HCV-induced liver cirrhosis in the Japanese population identifies novel susceptibility loci at the MHC region. Journal of Hepatology. 2013; 58 (5):875-882. DOI: 10.1016/j.jhep.2012.12.024. Epub 2013 Jan 12 - 77.
Hewitt EW. The MHC class I antigen presentation pathway: Strategies for viral immune evasion. Immunology. 2003; 110 (2):163-169. DOI: 10.1046/j.1365-2567.2003.01738.x - 78.
Boot HJ, Hahné S, Cremer J, Wong A, Boland G, van Loon AM. Persistent and transient hepatitis B virus [HBV] infections in children born to HBV-infected mothers despite active and passive vaccination. Journal of Viral Hepatitis. 2010; 17 (12):872-878. DOI: 10.1111/j.1365-2893.2009.01247.x - 79.
Gokhale NS, Vazquez C, Horner SM. Hepatitis C virus. Strategies to evade antiviral responses. Future Virology. 2014; 9 (12):1061-1075. DOI: 10.2217/fvl.14.89 - 80.
Kanno T, Ishihara R, Hatama S, Uchida I. A long-term animal experiment indicating persistent infection of bovine coronavirus in cattle. The Journal of Veterinary Medical Science. 2018; 80 (7):1134-1137. DOI: 10.1292/jvms.18-0050. Epub 2018 May 18 - 81.
Teunis PF, Sukhrie FH, Vennema H, Bogerman J, Beersma MF, Koopmans MP. Shedding of norovirus in symptomatic and asymptomatic infections. Epidemiology and Infection. 2015; 143 (8):1710-1717. DOI: 10.1017/S095026881400274X. Epub 2014 Oct 22 - 82.
McCarthy MK, Morrison TE. Persistent RNA virus infections: Do PAMPS drive chronic disease? Current Opinion in Virology. 2017; 23 :8-15. DOI: 10.1016/j.coviro.2017.01.003. Epub 2017 Feb 17 - 83.
Randall RE, Griffin DE. Within host RNA virus persistence: Mechanisms and consequences. Current Opinion in Virology. 2017; 23 :35-42. DOI: 10.1016/j.coviro.2017.03.001. Epub 2017 Mar 17 - 84.
Porter KR, Beckett CG, Kosasih H, Tan RI, Alisjahbana B, Rudiman PI, et al. Epidemiology of dengue and dengue hemorrhagic fever in a cohort of adults living in Bandung, West Java, Indonesia. American Journal of Tropical Medicine and Hygiene. 2005; 72 (1):60-66 - 85.
Thenmozhi V, Rajendran R, Ayanar K, Manavalan R, Tyagi BK. Long-term study of Japanese encephalitis virus infection in Anopheles subpictus in Cuddalore district, Tamil Nadu, South India. Tropical Medicine & International Health. 2006; 11 (3):288-293. DOI: 10.1111/j.1365-3156.2006.01567.x - 86.
Nielsen CF, Reisen WK, Armijos MV, Maclachlan NJ, Scott TW. High subclinical West Nile virus incidence among nonvaccinated horses in northern California associated with low vector abundance and infection. The American Journal of Tropical Medicine and Hygiene. 2008; 78 (1):45-52 - 87.
Chen Y, Zhao Y, Hammond J, Hsu HT, Evans J, Feldlaufer M. Multiple virus infections in the honey bee and genome divergence of honey bee viruses. Journal of Invertebrate Pathology. 2004; 87 (2-3):84-93. DOI: 10.1016/j.jip.2004.07.005 - 88.
Locke B, Semberg E, Forsgren E, de Miranda JR. Persistence of subclinical deformed wing virus infections in honeybees following Varroa mite removal and a bee population turnover. PLoS One. 2017; 12 (7):e0180910. DOI: 10.1371/journal.pone.0180910 - 89.
Flegel TW. The shrimp response to viral pathogens. In: Proceedings of the Special Session on Sustainable Shrimp Aquaculture, World Aquaculture [190-214]. Orlando, Boca Raton, USA: World Aquaculture Society; 2001 - 90.
Sritunyalucksana K, Srisala J, McColl K, Nielsen L, Flegel TW. Comparison of PCR testing methods for white spot syndrome virus [WSSV] infections in penaeid shrimp. Aquaculture. 2006; 255 :95-104. DOI: 10.1016/j.aquaculture.2005.12.002 - 91.
Walker PJ, Mohan CV. Viral disease emergence in shrimp aquaculture: Origins, impact and the effectiveness of health management strategies. Reviews in Aquaculture. 2009; 1 (2):125-154. DOI: 10.1111/j.1753-5131.2009.01007.x. Epub 2009 May 15 - 92.
Bhaskaran M, Murali SV, Rajaram B, Krishnasamy S, Devasena CS, Pathak A, et al. Association of HLA-A, -B, DRB, and DQB alleles with persistent HPV-16 infection in women from Tamil Nadu, India. Viral Immunology. 2019; 32 (10):430-441. DOI: 10.1089/vim.2019.0094. Epub 2019 Dec 3 - 93.
Ramezani A, Aghakhani A, Kalantar E, Banifazl M, Eslamifar A, Velayati AA. HLA-A *3303* and *3301 predispose patients to persistent hepatitis B infection. Journal of Gastrointestinal and Liver Diseases. 2009; 18 (1):117-118 - 94.
Pan N, Chen K, Qiu J, Sun H, Xu J, Miao F, et al. Human leukocyte antigen class I alleles and haplotypes associated with primary hepatocellular carcinoma in persistent HBV-infected patients. Human Immunology. 2013; 74 (6):758-763. DOI: 10.1016/j.humimm.2013.02.007. Epub 2013 Mar 1 - 95.
Zhu M, Dai J, Wang C, Wang Y, Qin N, Ma H, et al. Fine mapping the MHC region identified four independent variants modifying susceptibility to chronic hepatitis B in Han Chinese. Human Molecular Genetics. 2016; 25 (6):1225-1232. DOI: 10.1093/hmg/ddw003. Epub 2016 Jan 13 - 96.
Kondo Y, Ueno Y, Shimosegawa T. Dysfunction of immune systems and host genetic factors in hepatitis C virus infection with persistent normal ALT. Hepatitis Research and Treatment. 2011; 2011 :713216. DOI: 10.1155/2011/713216. Epub 2011 Jun 14 - 97.
McKiernan SM, Hagan R, Curry M, McDonald GS, Kelly A, Nolan N, et al. Distinct MHC class I and II alleles are associated with hepatitis C viral clearance, originating from a single source. Hepatology. 2004; 40 (1):108-114. DOI: 10.1002/hep.20261 - 98.
Mosaad YM, Farag RE, Arafa MM, Eletreby S, El-Alfy HA, Eldeek BS, et al. Association of human leucocyte antigen class I [HLA-A and HLA-B] with chronic hepatitis C virus infection in Egyptian patients. Scandinavian Journal of Immunology. 2010; 72 (6):548-553. DOI: 10.1111/j.1365-3083.2010.02468.x - 99.
Fan S, Wang Y, Wang S, Wang X, Wu Y, Li Z, et al. Polymorphism and peptide-binding specificities of porcine major histocompatibility complex [MHC] class I molecules. Molecular Immunology. 2018; 93 :236-245. DOI: 10.1016/j.molimm.2017.06.024. Epub 2017 Jul 24 - 100.
Provenzano M, Panelli MC, Mocellin S, Bracci L, Sais G, Stroncek DF, et al. MHC-peptide specificity and T-cell epitope mapping: Where immunotherapy starts. Trends in Molecular Medicine. 2006; 12 (10):465-472. DOI: 10.1016/j.molmed.2006.08.008. Epub 2006 Sep 7 - 101.
Fares-Gusmao R, Rocha BC, Sippert E, Lanteri MC, Áñez G, Rios M. Differential pattern of soluble immune markers in asymptomatic Dengue, West Nile and Zika virus infections. Scientific Reports. 2019; 9 (1):17172. DOI: 10.1038/s41598-019-53645-w - 102.
Flegel TW. Research progress on viral accommodation 2009 to 2019. Developmental and Comparative Immunology. 2020; 112 :103771. DOI: 10.1016/j.dci.2020.103771. Epub 2020 Jul 4. Erratum in: Developmental and Comparative Immunology. 2021;118 :104016 - 103.
Pasharawipas T. Inducible viral receptor, a possible concept to induce viral protection in primitive immune animals. Virology Journal. 2011; 8 :326. DOI: 10.1186/1743-422X-8-326 - 104.
Zambon RA, Vakharia VN, Wu LP. RNAi is an antiviral immune response against a dsRNA virus in Drosophila melanogaster . Cellular Microbiology. 2006;8 (5):880-889. DOI: 10.1111/j.1462-5822.2006.00688.x - 105.
Barquet N, Domingo P. Smallpox: The triumph over the most terrible of the ministers of death. Annals of Internal Medicine. 1997; 127 (8 Pt 1):635-642. DOI: 10.7326/0003-4819-127-8_part_1-199710150-00010. Erratum in: Annals of Internal Medicine. 1998;128 (9):787 - 106.
Damaso CR. Revisiting Jenner’s mysteries, the role of the Beaugency lymph in the evolutionary path of ancient smallpox vaccines. The Lancet Infectious Diseases. 2018; 18 (2):e55-e63. DOI: 10.1016/S1473-3099[17]30445-0. Epub 2017 Aug 18