Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
Perspective Chapter: Specific Predictors of the Autoimmune Reactions Formation in Case of Immunocompetent Organs Damage in Patients with Myasthenia Gravis and Hepatosplenomegaly
Written By
Elena Klimova, Larisa Drozdova, Olena Lavinska, Sergey Sushkov and Valery Boyko
Submitted: 24 May 2023Reviewed: 04 August 2023Published: 15 December 2023
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
The urgency of the problem is determined by the increasing prevalence and rapid progression of autoimmune diseases and autoimmune components in various nosologies. The aim is to study individual trigger factors, predictors of development, and the condition severity markers to substantiate complex treatment, including surgical tactics and the therapeutic target choice, in case of the immunocompetent organs (thymus and spleen) damage. In patients with myasthenia gravis the trigger markers were identified: the presence of herpes viruses persistence and mycoplasma; the relationship of certain human leucocyte antigen (HLA) molecules; high content of cytotoxic damage-associated molecular patterns (DAMPs); decreased expression of CD8+ T lymphocytes and co-stimulatory molecules CD3+CD4+CD28+. Some patients with myasthenia gravis had antibodies to α1 and α7 subunits nicotinic acetylcholine receptors (nAChR), etc. Patients with hepatosplenomegaly depending on the trigger factors (hepatitis HBV/HBC, herpes viruses (CMV/EBV)) and genetic predictors (hereditary enzymopathy) had specific markers, such as activation or inhibition of barrier function, reactive oxygen species (ROS) production, an increase in the concentration of cytokines, changes in the clusters of differentiation expression and specific autoantibodies. Thus, the creation of supplemented diagnostic protocols with additional markers for patients with various autoimmune reactions will make it possible to substantiate personalized immunocorrection.
State Institution “Zaycev V. T. Institute of General and Urgent Surgery of National Academy of Medical Sciences of Ukraine”, Kharkiv, Ukraine
V. N. Karazin National University, Kharkiv, Ukraine
Larisa Drozdova
State Institution “Zaycev V. T. Institute of General and Urgent Surgery of National Academy of Medical Sciences of Ukraine”, Kharkiv, Ukraine
Olena Lavinska
State Institution “Zaycev V. T. Institute of General and Urgent Surgery of National Academy of Medical Sciences of Ukraine”, Kharkiv, Ukraine
V. N. Karazin National University, Kharkiv, Ukraine
Sergey Sushkov
State Institution “Zaycev V. T. Institute of General and Urgent Surgery of National Academy of Medical Sciences of Ukraine”, Kharkiv, Ukraine
Valery Boyko
State Institution “Zaycev V. T. Institute of General and Urgent Surgery of National Academy of Medical Sciences of Ukraine”, Kharkiv, Ukraine
*Address all correspondence to: klimovalena53@gmail.com
1. Introduction
There is an increase in autoimmune diseases and autoimmune component formation in the pathogenesis of many nosologies. The causes of autoimmune diseases are not completely clear. Many autoimmune diseases are multifactorial and with their progression, various organs can be involved in the pathological process [1].
The implementation of immune markers into clinical practice is limited due to insufficient information of the mechanisms that lead to changes in the metabolism of immune cells under the trigger factors.
Central and peripheral mechanisms of self-tolerance loss, genomic and epigenomic predictors associated with the autoimmune pathology (mutations of the autoimmune regulator (AIRE); HLA polymorphism; impaired editing of immunoglobulin genes; changes in the receptors structure and epigenomic marks formation of certain genes, etc.) are studied [2].
Some nosologies may have organ-nonspecific and organ-specific manifestations of reactions. If autoimmune disorders develop in the immune organs, then diseases are severe and often accompanied by life-threatening symptoms and a high risk of mortality. Such pathologies include generalized myasthenia gravis and hepatosplenomegaly, which are characterized by autoimmune damage of the immune organs: thymus and spleen, resulting in structural and functional disorders of these organs, in the form of hyperplasia or neoplasia. In autoimmune disorders of the thymus and spleen, it is relevant to investigate the features mechanisms formation of autoimmune reactions in patients with autoimmune myasthenia gravis and hepatosplenomegaly of various origins. It is necessary to search for additional pathogenetic links in order to develop approaches for diagnosing the mechanisms of formation and treatment of autoimmune reactions of various geneses [3].
Trigger factors for the occurrence of myasthenia gravis and hepatosplenomegaly can often be infectious factors, various specificities of autoantibodies, or stress factors that provoke the cells activation of the immune-neuroendocrine complex (e.g., with glandular syndrome). The most important predictor of the development of autoimmune diseases is a genetic predisposition. The relationship between the presence of certain HLA diplotypes and haplotypes is known; polymorphism of candidate genes that determine hereditary enzymopathy and high genome mutability; an increase in the frequency of chromosomal aberrations with certain clinical phenotypes of autoimmune diseases. Also, the formation of myasthenia gravis can occur as a result of epigenomic changes—hypermethylation of individual genes, which affects the synthesis of mRNA (inhibits or activates) responsible for the neurotransmitters production that affects the potential of the end plate. Myasthenia gravis is a disease with a progenitor course, characterized by progressive muscle weakness, ptosis, shortness of breath, and often the development of a myasthenic crisis with a fatal outcome [3, 4].
Hepatosplenomegaly accompanies different severe pathology, such as cirrhosis and hepatitis; and hereditary diseases such as Gaucher’s syndrome, Pompe’s syndrome, cystic fibrosis (associated with recurrent infectious and inflammatory processes), and other orphan diseases manifested by impaired hematopoiesis and osteogenesis. The progression of hepatosplenomegaly manifests itself in the form of portal hypertension and recurrent gastrointestinal bleeding, which is a risk factor for mortality. Recurrent bleeding in hepatosplenomegaly can be against the background of impaired blood flow in this area, and hypertension of the portal vein against the background of hypersplenism, as a result of hereditary pathology, in particular, lysosomal enzymopathy [1, 5]. It is also important to take into account violations of proliferation and differentiation of immunocompetent cells, due to changes in intercellular interaction and various humoral factors.
It is relevant to study the markers of the autoimmune pathologies development in order to determine specific therapeutic targets. The treatment of different autoimmune diseases is to relieve symptoms by suppressing the immune system. For this, hormone therapy, monoclonal antibodies, and plasmapheresis are used. But the treatment of these pathologies is not always effective. The treatment of myasthenia gravis and hepatosplenomegaly should take into account the multifactorial nature of nosologies with an autoimmune component. To achieve better results of complex treatment, it is necessary to study individual trigger factors, development predictors, and markers of the condition severity to assess the risk of mortality and disease progression. An effective method of providing urgent care for patients with myasthenia gravis and hepatosplenomegaly is surgery (removal of the thymus and spleen).
The aim is to study individual trigger factors, predictors of development, and the condition severity markers to substantiate complex treatment, including surgical tactics and the therapeutic target choice, in the case of the immunocompetent organs (thymus and spleen) damage.
2. Markers of self-tolerance loss in patients with various clinical forms of generalized myasthenia gravis
Myasthenia gravis is a multifactorial autoimmune disease characterized by pathological progressive muscle weakness. The violation of neuromuscular synaptic transmission is a basis of pathogenesis [6, 7]. The heterogeneity of the clinical forms of myasthenia gravis depends on a variety of pathogenetic mechanisms that are formed at various levels of the body’s organization. Structural and functional disorders of the thymus are often manifested in autoimmune myasthenia gravis (in the form of hyperplasia or malignant thymoma) [8]. The pathogenetic development of myasthenia gravis is associated with the production of antibodies to the neuromuscular junction targets, to the structures of nicotinic acetylcholine receptors (nAChR), presynaptic ryanodine receptors (RyR), postsynaptic antigens – LRP4 (low-density lipoprotein receptor-related protein 4), agrin and tyrosine kinase receptors (MuSKR) [9, 10, 11]. Some authors have identified antibodies to extrasynaptic antigenic proteins—titin, actin, filamin, and actinin [12, 13, 14]. Depending on the presence of morphological and functional disorders of the thymus, myasthenia gravis is classified as thymus-independent or thymus-dependent (against the background of thymus hyperplasia or thymoma [15]. Neurotransmitter disorders in myasthenia gravis manifest themselves against the background of the self-tolerance loss to one’s own tissues [16].
Myasthenia gravis is based on damaged mechanisms of immunological autotolerance. Normally, during the formation of central self-tolerance, a program for the development of lymphocytes is normally implemented, in which aggressive autoreactive clones are eliminated in the thymus. The AIRE complex is involved in the differentiation of thymocytes, migration and apoptosis within the thymus, alternative mRNA splicing, miRNA expression, transactivation of HLA expression, and is important for testing thymocyte autoreactivity [17]. Autoaggressive clones of T lymphocytes and B lymphocytes are formed in the central organs of the immune system in the central mechanisms of self-tolerance violation. Structural disorders in the thymus in myasthenia gravis may be the result of central self-tolerance loss due to impaired selection of autoreactive lymphocytes in the thymus. The mechanisms of peripheral self-tolerance loss may be formed due to a lack of AIRE gene expression in immunocompetent cells [18, 19]. The second way of self-tolerance formation is normally carried out in peripheral lymphoid organs and limits the activation of autoreactive lymphocytes with the help of T regulatory cells (Treg) that have not been eliminated by the mechanisms of central tolerance. These mechanisms ensure the peripheral tolerance formation [20].
Treatment of myasthenia gravis is determined by standard protocols and includes anticholinesterase drugs, corticosteroids, plasmapheresis, and surgical treatment—removal of the thymus. The whole complex of modern therapeutic approaches does not always achieve an acceptable effect [21, 22]. Removal of the thymus, the target of autoimmune aggression, does not always allow avoiding anticholinesterase drugs and sometimes leads to the development of myasthenic and cholinergic crises [23, 24]. A search is underway for new approaches in diagnostics for personalized treatment choice, taking into account immunocorrection.
2.1 Materials and methods in the study of immune reactions in myasthenia gravis
We examined 492 patients (18–78 years) with thymus-dependent and thymus-independent myasthenia gravis. The first group with thymus-independent myasthenia gravis included 241 patients aged 18–35 years (138 women and 103 men); the second group with thymus-dependent myasthenia gravis against the background of thymic hyperplasia consisted of 59 patients aged 36–72 years (44 women and 15 men), the third group with thymus-dependent myasthenia gravis and thymoma included 192 patients aged 35–78 years old (106 women and 86 men). The diagnosis was made on the basis of determining muscle weakness and pathological muscle fatigue, taking into account the temporal reversibility of clinical and electromyography changes. According to the indications, patients underwent thymectomy. Myasthenic crisis developed in 18% of patients with thymoma after surgery.
To assess the change in the spectrum of factors leading to impaired self-tolerance, a number of indicators characterizing genomic and epigenomic trigger factors that cause the development of myasthenia gravis were studied by the following methods:
ELISA – the presence of viral persistence; concentration of total IgE; the cytokine profile IL-2, IL-4, IL-8, TNF-α; the presence of autoantibodies repertoire (the level of organ-specific antibodies, to cellular organelles (ANA), to α1 and α7 subunits of the nicotinic acetylcholine receptors (nAChR), and to the mitochondrial α7 subunit of neuronal nAChR in the thymus tissue).
Light microscopy – leukocyte allele polymorphism class II antigens were assessed serologically using HLA phenotyping: DR1, DR2, DR3, DR5, DR7, DR52; the indicators of chemotaxis, adhesion and endocytosis of phagocytes in oxygen-independent phagocytosis.
Fluorescence microscopy – the presence of autoantibodies ANA.
Flow cytometry – the level of NK cells CD3-CD56+CD16+; B lymphocytes CD3-CD20+/CD45+, T helpers CD3+CD4+; cytotoxic T lymphocytes CD3+CD8+/CD45+; marker of early activation of the inflammatory process CD3+CD4+CD25+, regulatory T lymphocytes CD3+CD4+CD25+CD127-, costimulatory molecules CD3+CD4+CD28+, marker of late activation of lymphocytes (CD3+HLA-DR+).
The relative content of IgG class autoantibodies to 24 antigens of the main human organs and systems were studied in myasthenia gravis in blood serum samples: native DNA, Fc fragment of IgG (Fc), beta-2-glycoprotein I (β2), myocardial cell membrane antigens (CoM), myocardial β1-adrenergic receptors (βAR), platelet membrane antigens (TrM), vascular endothelial anionic proteins (ANCA), renal tissue cytoplasmic antigens (KiS), renal tissue membrane antigens (KiM), membrane lung tissue antigens (LuM), lung tissue cytoplasmic antigens (LuS), gastric mucosal cell membrane antigens (GaM), small intestine mucosal cell membrane antigens (ItM), liver tissue cytoplasmic antigens (HeS), liver cell mitochondrial antigens (HMMP), thyroglobulin (TG), thyroid stimulating hormone receptors (TSH-R), insulin (Ins), insulin receptors (Ins-R), adrenal cell membrane antigens (Adr), prostate and sperm cell membrane antigens (Spr), protein S100 (S100), astrocyte intermediate filament protein (GFAP), and myelin basic protein (MBP).
2.2 Trigger factors of autoimmune myasthenia gravis
Violation of molecular events that cause intercellular interactions during the development of an immune response to various infectious agents–PAMPs (pathogen-associated molecular patterns), endogenous and exogenous DAMPs (damage-associated molecular patterns), and food antigens can lead to a violation of the mechanisms of self-tolerance formation.
Long-term studies have shown that in patients with various clinical phenotypes of myasthenia gravis, immunoresistance disorders are interrelated with the accumulation of a combined antigenic load–a bacterial and viral infection (CMV, EBV, hepatitis virus HBV/HCV, herpes virus HSV-1, HSV-2, HHV-6, and mycoplasma).
Some food allergens and food-specific IgE and IgG4 also contribute to the increase in body allergic reactions and autoimmune component formation [25, 26].
2.2.1 Viral infection as a trigger factor for the development of myasthenia gravis
Trigger factors for dysfunction of the neuroendocrine-immune complex in myasthenia gravis can be exogenous stress factors, bacterial and viral infections in target organs, including the thymus, which change the activity of body cells and contribute to the development of long-term inflammation due to the presence of persister cells [27, 28].
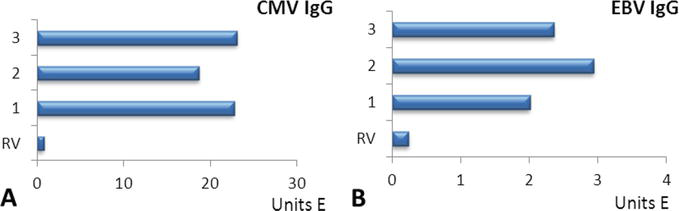
A high titer of antibodies to CMV was revealed in myasthenia gravis and thymoma by 30 times and in the thymic hyperplasia by 23 times (Figure 1A).
Figure 1.
The titer of IgG CMV (A) and IgG EBV (B) in patients with thymus-independent myasthenia gravis (1); thymus-dependent myasthenia gravis with thymic hyperplasia (2); thymus-dependent myasthenia gravis with thymoma (3); RV – Reference values.
The maximum incidence of CMV was in myasthenia gravis and thymoma–in 91.1% and 91.0%, respectively.
The titer of antibodies to EBV was significantly higher (by 10 times) than the reference values in all groups. The maximum concentration of antibodies to EBV was detected in thymic hyperplasia, and to CMV—in myasthenia gravis and thymoma (Figure 1B).
The highest incidence of antibodies to EBV was found in patients with thymic hyperplasia – in 100%. In myasthenia gravis and thymoma, the incidence of EBV was slightly lower and amounted to 89.0% and 87.4%, respectively.
Thus, the virus persistence can be considered as a trigger factor for this disease.
2.2.2 The content of total and allergen-specific IgE in patients with myasthenia gravis
In addition to infectious antigens, an exogenous trigger factor for the development of myasthenia gravis may be the presence of food allergens. We studied the concentration of total IgE in blood serum and allergen-specific IgE to food allergens in plasma. The panel of food allergens included antigens of animal proteins, cereals and flour, dairy products, mushrooms, nuts, carbohydrates, and drinks, in total 94 items.
In 72% of patients with generalized myasthenia gravis, congenital atopy was detected, as judged by an increased concentration of total IgE, which averaged 322.0 ± 34.0 IU/ml with a reference interval of 50–100 IU/ml.
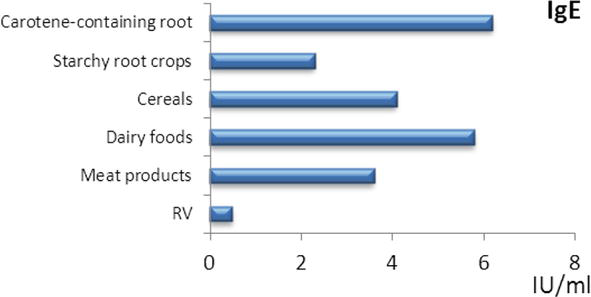
In patients with myasthenia gravis, a high degree of polyclonal sensitization of allergen-specific IgE to foodstuffs in average 5.8 ± 0.6 IU/ml at reference values 0.30 ± 0.01 IU/ml of protein origin, cereals, and starch root crops containing carotenoids was revealed. Most often, sensitization to dairy products, to meat and cereals was detected in patients with myasthenia gravis (Figure 2).
Figure 2.
The content of specific IgE antibodies to food allergens in patients with myasthenia gravis. RV – Reference values.
In myasthenia gravis and thymoma, an increased content of specific IgE antibodies to dairy products (increase by 11 times), to meat products (increase by 7 times), to cereals (by 8 times), starch-containing root crops, to carotene-containing products relative to the reference values, with thymoma—to dairy and meat food allergens.
In some patients with polyclonal sensitization to food allergens, the concentration of total IgE did not exceed the reference values (from 70 to 130 IU/ml).
The absence of specific IgE in peripheral blood serum does not exclude the possibility of its participation in the pathogenesis of the IgE-dependent mechanism of allergic reactions, since its local synthesis or binding of synthesized IgE by tissues is possible, which can occur without changing the concentration of IgE in peripheral blood [29].
Thus, one of the possible epigenomic factors in the formation of autoimmune myasthenia gravis may be food allergy.
2.3 Dysregulation of immunoreactivity and changes in the cytokine profile in different clinical phenotypes of myasthenia gravis
The activation of numerous and specialized immune cell subpopulations are under the control of the cellular regulation and cytokine network. Secondary activation of the expression of mitochondrial and nuclear genes is carried out by PAMPs (pathogen-associated molecular patterns) and DAMPs (damage-associated molecular patterns), which contribute to the synthesis of cytokines, nucleotides, nucleosides, RNA, and heat shock molecules [30]. The identified infectious trigger factors leading to the formation of molecular compounds PAMPs also contribute to the formation of various classes of endogenous cytotoxic molecules DAMPs, related to immunogenic molecules that are formed as a result of the breakdown of necrotic and apoptotic cells, as well as cells undergoing autophagy.
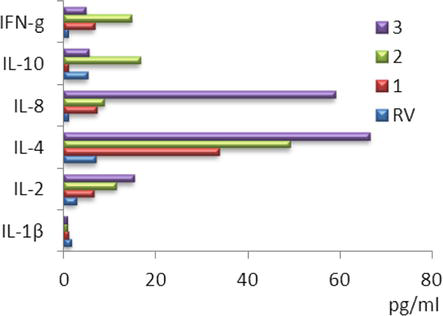
We revealed an increase of the IFN-γ content in all groups. The maximum increase in the IFN-γ concentration was observed in thymic hyperplasia.
In thymic hyperplasia and thymoma IL-4 content exceeded the reference values. The maximum value of IL-4 was found in thymoma – 66.4 ± 4.5 pg./ml; in thymic hyperplasia, this cytokine was increased sevenfold to 49.2 ± 5.2 pg./ml (Figure 3).
Figure 3.
The contents of cytokines in patients with thymus-independent myasthenia gravis (1); thymus-dependent myasthenia gravis with thymic hyperplasia (2); thymus-dependent myasthenia gravis with thymoma and (3); RV – Reference values.
Patients with thymoma had the maximum increase in IL-8. According to most authors, IL-8 manifests itself as a factor in enhancing tumor progression. This is explained by the action of IL-8 as an autocrine tumor growth factor and as a factor in enhancing angiogenesis by influencing the capillaries of metastases [22].
Synthesis of IL-10 was inhibited in the myasthenia gravis and amounted to 1.0 ± 0.6 pg./ml and in the thymoma – 5.5 ± 0.9 pg./ml. In the thymic hyperplasia, IL-10 was maximally elevated and exceeded reference values by three times.
The maximum increase (up to 15.3 ± 0.4 pg./ml) in IL-2 was revealed in thymoma.
IL-2 is an inducer of all cytotoxic cells that takes part in the formation of the Th1 phenotype and increases the production of IFN-γ. The synthesis of IL-2 forms the affinity of the TCR and activates HLA molecules. An increase of IL-2 was observed in the thymic hyperplasia and thymoma.
Thus, the most significant changes were found in the content of IL-4, IL-8, and IL-2.
2.4 An additional target of autoimmunization is the presence of nAChR in thymus mitochondria in patients with thymoma
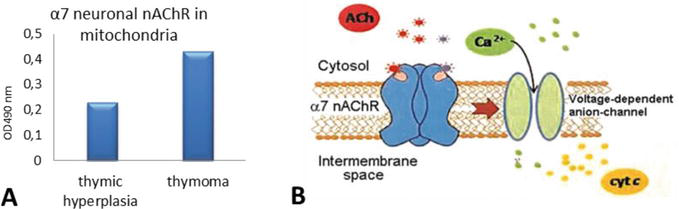
In the literature, changes in the thymus are practically not described, although there is a concept about the leading role of impaired central differentiation of T lymphocytes. Together with Professor Skok M.V. (Laboratory of Immunology cell receptors of the O.V. Palladin Institute of Biochemistry of the National Academy of Sciences of Ukraine) studies were carried out on the presence of α7 subunits of neuronal nicotinic acetylcholine receptors (nAChR) in mitochondria of thymus different layers obtained from patients with thymic hyperplasia and thymoma.
A significant difference was found in the content of α7 neuronal nAChR of thymus mitochondria in patients with thymic hyperplasia and thymoma (Figure 4A). The level of α7 neuronal nAChR mitochondria in the thymus in patients with thymic hyperplasia was 0.23 units E and in thymoma – 0.43 units E, which was 1.9 times higher than in thymus mitochondria not affected by the tumor process (Figure 4A).
Figure 4.
Changes in α7 nAChR level mitochondria in patients with thymic hyperplasia and thymoma (A) and the scheme of α7 neuronal nAChR localization and functioning in the outer membrane of mitochondria (B); adapted from [31].
The function of nAChRs in mitochondria is to control the formation of the mitochondrial transient conduction pore, which is the source of proapoptotic factors and reactive oxygen species (ROS) released in the cytosol (Figure 4B) [31, 32]. From this, it follows that the revealed difference between the mitochondria of patients with and without thymoma indicates that tumor transformation is accompanied by an increase in mitochondrial nAChRs, and this maintains the viability of tumor cells.
The presence of the α7 subunit nAChR in thymus mitochondria suggested that this structure can also serve as an additional target for inducing the formation of autoantibodies. Specific autoantibodies can bind to certain domains—subunits of nAChR and cause destruction of these receptors in the thymus.
2.5 Specific markers of self-tolerance loss in various clinical phenotypes of myasthenia gravis
The frequency of occurrence of autoantibodies to the α1 subunit of nicotinic acetylcholine receptors (nAChR) in all groups was 100% higher than the reference values.
Patients with thymic hyperplasia had the maximum increase (by 2.2 times) of antibodies to the α1 subunit nAChR.
The frequency of occurrence of antibodies to the α7 subunit nAChR was only in 20% of cases higher than in the control comparison group.
Patients with myasthenia gravis had the maximum increase (1.9 times) in antibodies to the α7 subunit nAChR.
2.5.1 The autoantibodies spectrum to various targets in various clinical phenotypes of myasthenia gravis
Patients with myasthenia gravis and thymoma had the widest range of autoantibodies (AABs)—12 and 11 specificities out of 24 studied, respectively.
Patients with thymic hyperplasia had 6 specificities out of 24 researched.
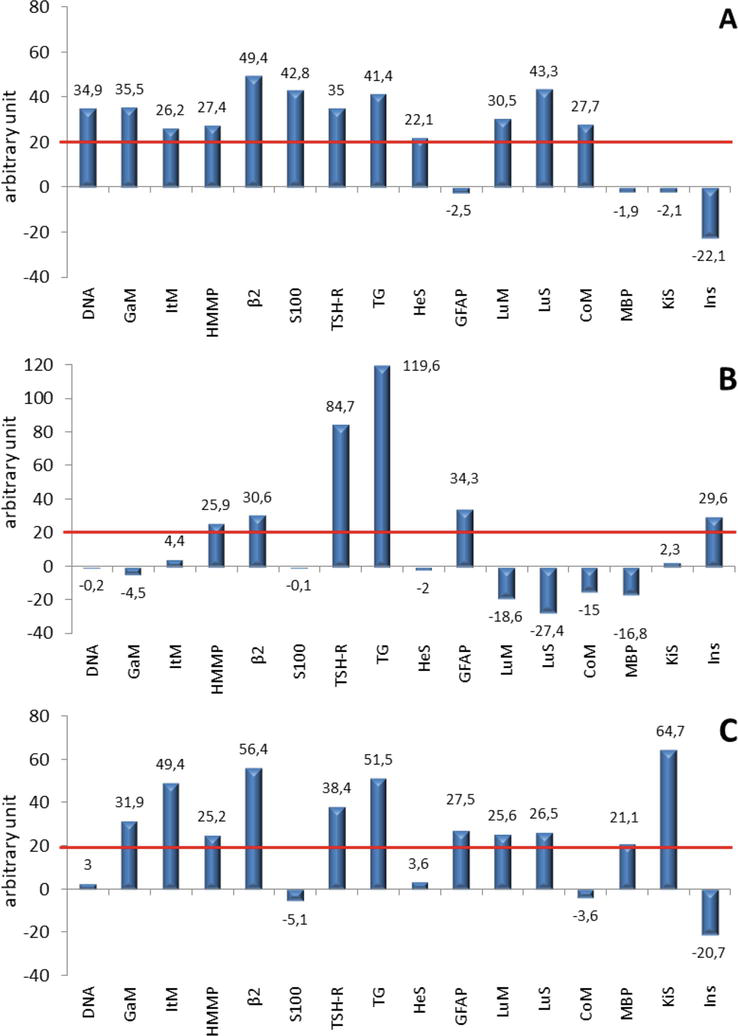
Four specificities of autoantibodies were detected with high frequency in all forms of myasthenia gravis: AABs to the TSH receptor (myasthenia gravis—with a frequency of 55.6%, thymic hyperplasia—66.7%, thymoma—70%), to thyroglobulin (myasthenia gravis—33.3%, thymic hyperplasia—66.7%, thymoma—40%), to β2-glycoprotein I (myasthenia gravis—33.3%, thymic hyperplasia—33.3%, thymoma—20%) and to hepatocyte mitochondria (myasthenia gravis—55.6%, thymic hyperplasia—33.3%, thymoma—10%). In all patients, organ-specific AABs at different levels were detected with different frequency (from 33 to 56%) (Figure 5).
Figure 5.
Change in the relative autoantibodies content from the individual average level of immunoreactivity (red line) in patients with myasthenia gravis (A), thymic hyperplasia (B), and thymoma (C).
Thus, in patients with myasthenia gravis and thymoma, the same additional targets for AABs were identified: membrane antigens of the mucous cells membrane of the stomach and small intestine, membrane and cytoplasmic antigens of the lung tissue, which may indicate possible identical mechanisms for the formation of myasthenia gravis these forms.
The progression of myasthenia gravis in ontogenesis may lead to the formation of myasthenia gravis on the background of thymoma in elderly patients. In myasthenia gravis with elevated concentrations of autoantibodies in the gastric mucosa (parietal cells), the presence of food allergy (high concentration of specific IgG4 and IgE) was revealed as a possible pathogenetic factor in the development of this disease.
Organ-specific antibodies in patients with thymic hyperplasia had other cellular targets and were represented by the following localization: AABs to insulin protein and astrocytes. In addition, in thymic hyperplasia, the concentration of AABs to the TSH receptor and thyroglobulin significantly exceeded the content of these antibodies in the myasthenia gravis and thymoma.
In patients with myasthenia gravis and thymoma, the concentration of autoantibodies to β2-glycoprotein I was significantly higher than in the patients with thymic hyperplasia.
The selective rise in the level of relative immunoreactivity in reaction with the above antigens can be considered as a factor in existing or emerging disorders in the relevant organs and systems.
2.6 Relationship of class II HLA various alleles antigens with different clinical phenotypes of myasthenia gravis
In addition to trigger exogenous factors identified in the examined patients with myasthenia gravis, the assessment of leukocyte antigens of class II HLA (Human Leukocyte Antigens) revealed the relationship between certain HLA DR alleles. HLA molecules play a central role in the immune response, determining the severity and direction of the reactions to antigenic exposure by forming a macromolecular complex of recognized antigens (HLA and T-cell receptor TCR) [33]. Many mononucleotide differences in the HLA genes determine their polymorphic variants. Some alleles may be associated with the development of certain pathological conditions, such as endocrine, reproductive, thrombolytic, and cardiovascular. Expression of HLA molecules on the immune system cell surface is a complex dynamic process, on which their functional activity largely depends [34].
In patients with a genetic predisposition the pathological activation of the immune system is associated with the functioning of the HLA-antigen complex, and it leads to the autoimmune disease development [35]. HLA-B27 heavy chains can bind to innate immune receptors on natural killer NK cells [36, 37].
The polymorphism of the HLA genes determines the selection of T lymphocytes during maturation in the thymus. In the presence of certain alleles of the HLA system genes, the elimination of T lymphocytes, which carry receptors for certain autoantigens on their surface, is impaired, while in a healthy body such T lymphocytes are destroyed at the maturation [38]. The function of class II HLA receptors is to present antigen peptides from the extracellular space. Pathological activation of the immune system in patients with a genetic predisposition can lead to autoimmune processes and a severe course of other diseases and a negative prognosis.
We revealed the presence of various allelic phenotypes of HLA associated with various forms of myasthenia gravis. Among the entire cohort of patients, the HLA DR5 phenotype was the most common.
In patients with myasthenia gravis, the allele encountered with the maximum frequency revealed HLA DR5 and HLA DR2.
In patient with thymic hyperplasia the phenotypes HLA DR5 (in 70%) and HLA DR1 (in 60%) were detected with high frequency.
In patients with thymoma, the frequency of occurrence for the HLA DR7 allele was revealed. In locally advanced thymomas, HLA DR3 and HLA DR52 were also detected.
Thus, a high frequency of detection of the HLA DR5 allele, both in homozygous and heterozygous states, was shown in all patients.
Patients with thymoma had high frequency of HLA DR7. The presence of this allele can serve as a diagnostic and prognostic marker for the thymoma development in patients included in cohort with myasthenia gravis without thymoma and also have the HLA DR7 allele.
2.7 Expression of cell surface clusters differentiation on lymphocytes in myasthenia gravis
To assess the effectiveness of interaction between T cells and B cells during the immune response development to various thymus-dependent bacterial and viral antigens, the expression of different subpopulation of lymphocytes was determined.
In 45% of patients with various phenotypes of myasthenia gravis, a decrease in CD8+ expression was noted.
An increase in aggressive clones’ natural killers CD3-CD56+CD16+ was revealed against the background of a decrease in cytotoxic CD8+. And it can lead to excessive cytotoxicity.
Patients with myasthenia gravis had the minimum level (38.4%) of CD4+CD28+ expression.
In patients with thymic hyperplasia, CD4+CD28+ expression was 1.2 times higher than in myasthenia gravis.
Patients with thymoma had the greatest increase (54.3%) in positive CD4+CD28+ cells.
In 18% of the patients, there was a decrease in the expression of CD8+ and CD4+CD28+.
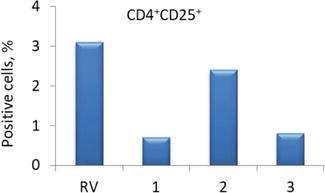
Patients with myasthenia gravis and thymoma had a fourfold decrease in the expression of CD25+ on CD4+ T cells compared to the reference values (Figure 6).
Figure 6.
Expression of CD4+CD25+ in patients with various types of self-tolerance loss: 1 – Myasthenia gravis; 2 – Thymic hyperplasia; and 3 – Thymoma; RV – Reference values.
Patients with thymic hyperplasia had the level of CD4+CD25+ cells on average 30% lower than reference values (Figure 6).
Probably, in thymoma, CD4+ T cells with a high expression of the CD25 molecule predominated, since it is these cells that have suppressor activity [39].
A significant increase in the lymphocytes late activation marker (CD3+HLA-DR+) was revealed in thymic hyperplasia (by 34%) and thymoma (by 87%) groups.
Violation of molecular processes that determine intercellular interactions during the development of an immune response to various infectious agents can lead to disruption of the mechanisms of self-tolerance formation [40].
2.8 The content of antinuclear autoantibodies (ANA) in patients with thymoma
It was revealed the presence of antinuclear autoantibodies (ANA, ELISA method) in patients with locally advanced thymoma (who developed myasthenic crisis in the postoperative period), the content of which was four times higher than the reference level and averaged 4.2 ± 0.2 units.
In myasthenia gravis and thymic hyperplasia ANA was not detected in the blood serum.
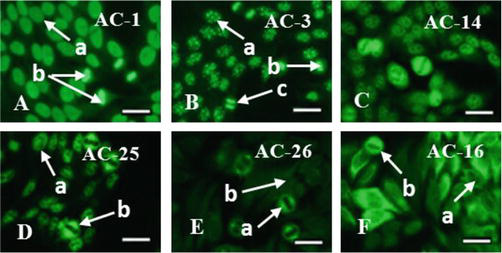
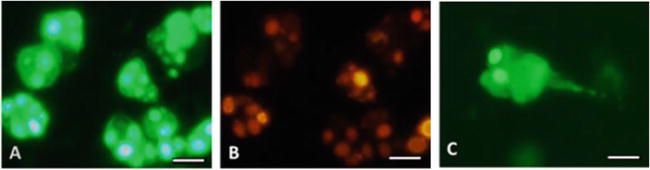
To visualize the specificity of ANA, immunofluorescent analysis was performed, which made it possible to identify targets for autoantibodies (AABs) in the form of various components of cell nuclei in patients with thymoma—ANA to histones, double-stranded DNA, and other chromatin-associated antigens (Figure 7A). Since due to the action of various factors, reparative processes in DNA are disrupted, a lot of erroneous codes appear in the structure of the DNA helix, which upon the next reading triggers the disturbed mechanism of histone formation.
Figure 7.
Antinuclear autoantibodies (ANA) in myasthenia associated with thymoma: A – To histones and double-stranded DNA (a – Stained nucleoli, b – Mitotic cells); B – To the centromeres (a – Uniform distribution of granules in the nucleus, b – Metaphase stage, c – Anaphase stage); C – To centromeric protein F (CTNPF); D – To the achromatin spindle protein NuMa (a – no staining of the nucleoli, b - fluorescence of the achromatin spindle fibers); E – To the MSA-2 antigen of mitotic spindle fibers (a – Mitotic spindle fibers, b – Unstained nuclei of interphase cells); F – To cytoskeletal proteins (a – Microtubules and intermediate fibers, b – Cytoskeletal proteins in mitotic cells). Fluorescent microscopy. Scale bar 20 μm. Codes AC (AC – Anti-cell pattern) in accordance with the international consensus on ANA patterns (ICAP) are presented. FITC staining of HEp-2 standard antigenic substrates after interaction with serum antibodies; magnification ×1000.
In patients with lymphoepithelial thymoma, ANA to chromosome centromeres was detected (Figure 7B). Centromeres play an important role in the regulation and functioning of the cell cycle. Ordered linkage and separation of centromeres is a prerequisite for the normal distribution of chromosomes in a mitotic cell. Violation of this due to the presence of antibodies leads to chromosomal instability and cancerous transformation.
Also, patients with thymoma had ANA to the centromeric protein F (CTNPF) (Figure 7C); to the protein involved in the formation of the mitotic spindle (NuMa), which is associated with the centrosome (Figure 7D); to the MSA-2 antigen of mitotic spindle fibers (Figure 7E), which is involved in the regulation of transcription during the G1/S transition of the mitotic cell cycle; to the cytoskeleton, represented by proteins cytokeratins and tropomyosin (Figure 7F).
It is known that cytokeratins are part of the intermediate filaments of the cell cytoskeleton, and tropomyosin is a fibrous protein that interacts with actin in muscle tissue and is involved in the process of muscle contraction.
2.9 Changes in immunoreactivity and structural organization of the thymus during the development of the immunopathological process with various types of self-tolerance loss in myasthenia gravis
There is a self-sustaining immune response to self-antigens in myasthenia gravis, which leads to cell damage. The study of the self-tolerance loss mechanisms in myasthenia gravis is of fundamental and practical importance; it can be used to predict the course of the disease and select treatment tactics.
A range of trigger factors for myasthenia gravis, thymic hyperplasia, and thymoma, such as persistence infection and IgG4 and IgE antibodies to food antigens was determined.
Various clinical types of myasthenia gravis have relationship with specific HLA alleles. Patients with myasthenia gravis had the highest frequency of occurrence of the HLA DR5 and HLA DR2 allele. In thymic hyperplasia, in addition to the HLA DR5 allele, the HLA DR1 allele was found with a high frequency. Patients with thymoma had a high frequency of HLA DR7 and HLA DR2 alleles.
In 50% of all patients, a decrease in the subpopulation of T killer cells CD8+ and an increase in NK cells CD3-CD56+CD16+ were observed.
Violation of dual recognition by changing the expression of CD4+CD28+ led to a violation of peripheral self-tolerance.
Patients with myasthenia gravis had the minimum level of CD4+CD28+ expression.
Patients with thymoma had the maximum increase in CD4+CD28+ cells.
In myasthenia gravis and thymoma was a fourfold decrease in the early marker of the inflammatory response CD4+CD25+.
In thymic hyperplasia and thymoma was a significant increase in the marker of lymphocytes late activation CD3+HLA-DR+.
Peripheral disorders of humoral self-tolerance in the form of an increase in the concentration of various autoantibodies were detected in most patients with myasthenia gravis, thymic hyperplasia, and thymoma. Autoantibodies react with cell surface molecules and have both blocking and stimulating effects (depending on the features of the target molecule and the signaling pathways associated with it).
An increase in the concentration of autoantibodies to the α1 subunit of nicotinic acetylcholine receptors (nAChRs) was shown in all patients.
The α7 subunit of neuronal nAChRs is an additional target for inducing the formation of autoantibodies mitochondrial structures. Mitochondria of thymocytes in patients with thymoma had the maximum level of α7 nAChR.
In addition to nAChR antibodies, a wide range of organ-specific antibodies were detected in all patients: out of 24 antigenic specificities of the panel, autoimmune antibodies were detected to 16. For all examined patients with myasthenia gravis (myasthenia gravis, thymic hyperplasia, and thymoma) is characterized by the presence of a high titer of autoantibodies to the TSH receptor and thyroglobulin, to hepatocyte mitochondria and β2-glycoprotein I.
Young patients with myasthenia gravis and elderly patients with thymoma had autoantibodies to the same targets (membrane antigens of mucous membrane cells stomach and small intestine, membrane and cytoplasmic antigens of lung tissue).
In addition to the main four antigenic targets characteristic of all group’ patients with thymic hyperplasia also formed an organ-specific antibodies to other targets: the insulin protein and the protein of astrocyte intermediate filaments.
Only patients with thymoma had antinuclear antibodies (ANA): to histones and double-stranded DNA, to chromosomal centromere proteins, to centromeric protein F (CTNPF), to the achromatin spindles protein NuMa, to the mitotic spindle fiber antigen MSA-2, to cytoskeletal proteins.
Perhaps the reason for the diversity of antibodies is a violation of the negative selection of autoreactive T lymphocytes and the presentation of self-antigens. The detection of autoantibodies with different characteristics in the blood serum of patients with myasthenia gravis proves the immunological heterogeneity of this disease and the different mechanisms of self-tolerance loss.
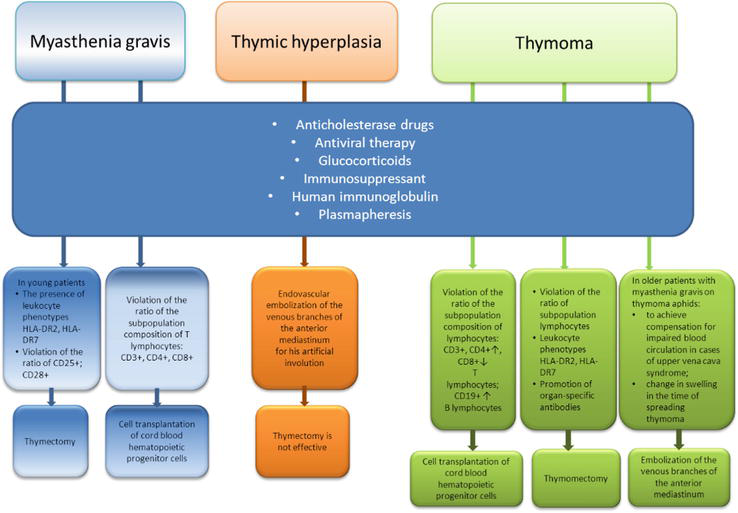
The results of some regularity in the autoimmune pathology formation make it possible to determine additional targeted for their treatment. The prognosis of myasthenia gravis progression and remission development can be carried out using the identified markers of self-tolerance loss. Specialists of the Institute (State Institution “Zaycev V. T. Institute of General and Urgent Surgery of National Academy of Medical Sciences of Ukraine) carried out the complex treatment in patients with myasthenia gravis, thymic hyperplasia, and thymoma, perform thymectomy in patients with thymic hyperplasia and locally advanced thymoma. Treatment methods are used in various combinations and sequences in patients with thymoma (embolization of thymus vessels in order to reduce the tumor size; endovascular transcatheter sclerotherapy in patients with locally advanced thymoma in the superior vena cava syndrome with transvenous destruction of the thymus parenchyma as a preoperative preparation). Thymomectomy and thymectomy often lead to the development of postoperative complications in the form of myasthenic and cholinergic crises (Figure 8).
Figure 8.
The strategy of treatment for patients with myasthenia gravis.
Indications for surgical treatment of thymoma are close to absolute, but as a rule, without taking into account the course nature and the disease severity.
In 12% young patients with progressive myasthenia gravis, surgical treatment is indicated.
Determination of immunological markers in myasthenia gravis made it possible to substantiate new approaches to treatment in young patients with myasthenia gravis.
In elderly patients with locally advanced thymoma, it is necessary to reduce the tumor volume via applying new X-ray endovascular interventions.
But, unfortunately, surgical treatment of patients with thymoma leads to satisfactory remission only in a third of patients, since it basically eliminates some of the disease symptoms.
The age-dependent development of the disease is not always taken into account for choosing treatment tactics. It has been shown that the disease onset in young patients with myasthenia gravis is often characterized by a low degree of severity.
The progression of myasthenia gravis in ontogenesis can lead to myasthenia gravis and thymic hyperplasia in elderly patients. In this case, surgical treatment is not effective and can lead to complications and death [41, 42].
If genetic predictors of HLA DR2, HLA DR7, and markers of central autotolerance loss are detected in young patients with myasthenia gravis, then it is advisable to recommend thymectomy in the early stages of the disease. This is necessary to avoid further thymus-dependent myasthenia gravis, in the form of locally advanced thymomas and postoperative complications during surgical intervention at a later stage of the disease—an attack of autoantibodies on other target organs, bleeding.
If markers of peripheral autotolerance loss are detected in patients with myasthenia gravis (possible thymus hyperplasia formation), then thymectomy is not advisable.
We also have a positive experience in the complete cure of generalized myasthenia gravis in non-operated patients after a long 35-day fast followed by stimulation of the phagocytic function of neutrophils (since their endoitosis, according to our data, was suppressed in all patients with myasthenia gravis [43]) using a composite preparation containing enzymes.
It is relevant to determine additional markers in patients with myasthenia gravis, thymic hyperplasia, and thymoma, leading to changes in the afferent and efferent links of immunity for complex treatment, including targeted immunocorrection.
Individual methods of correction for self-tolerance loss in myasthenia gravis should take into account the state of the immunity main organ and include the use of diet, immunosuppressants, specific neutralizing immunoglobulins, massive IgG therapy, and the use of anti-inflammatory recombinant interleukins.
3. Immunometabolic markers in patients with hepatosplenomegaly of various etiologies
An enlarged spleen is often combined with liver pathology. Both organs are involved in the pathological process, and hepatosplenomegaly (or hepatolienal syndrome) develops. Hepatosplenomegaly (HSM) accompanies a number of pathological conditions (hepatitis cirrhosis, parasitic lesions, storage diseases, malignant and benign tumors, acute and chronic diseases) and is often complicated by portal hypertension, recurrent bleeding from esophageal varices. Causes of enlargement of the spleen, indicating the presence of hepatolienal syndrome are bacterial infections, protozoal infections, helminthiases, immune thrombocytopenic purpura, splenogenic neutropenia, agranulocytosis, polyarthritis nodosa, portal hypertension, portal vein occlusion, Nimmann-Pick disease, Gaucher disease, Felty’s syndrome, sarcaidosis, amyloidosis, hepatocerebral dystrophy, and mononucleosis [5]. Hepatosplenomegaly disrupts the main function of the spleen to eliminate defective senescent cells (that have changed their phenotype) and PAMPs (pathogen-associated molecular patterns) [44]. The liver mediates detoxification reactions at the molecular level, and during infection, pathological molecular processes occur with the formation of cytotoxic molecular patterns of cellular debris (DAMPs—damage-associated molecular patterns) [45, 46]. Thus, it is necessary for additional screening and differential diagnosis of hepatolienal syndrome.
3.1 Trigger factors in the development of hepatosplenomegaly
Hepatosplenomegaly trigger factors are infectious antigens in the form of PAMPs viral structures of microorganisms and protozoa [30, 47]. The study of the bacterial contamination degree and viral load in patients with HSM are contradictory. It is considered that viral hepatitis and alcohol are trigger factors for the development of a HSM complicated course. Alcohol and infectious antigens exacerbate the development of hepatomegaly [48, 49]. However, there is a pronounced clinical heterogeneity of etiological factors and features of reactions occurring in the liver and spleen under the influence of other triggers and predictors [50, 51]. In this work, the studies made it possible to identify trigger factors, as well as genetic predictors in the form of specific immunological and metabolic markers for HSM development.
3.2 Bacterial and viral persisters of hepatosplenomegaly
There is a long-term presence of infectious antigens in accordance with the hypothesis of hidden antigens, the presence of superantigens, molecular mimicry, antigen complementarity, and idiotype-antiidypic interactions [52]. Against the background of immunosuppression, viral and bacterial persistent cells can form, which generates irreversible inflammatory reactions. These contribute to the development of irreversible autoimmune pathologies, including in hepatosplenomegaly [53, 54]. In prolonged persistent infection, the spectrum of specific T lymphocytes and antibodies expands, and more and more new epitopes of the same protein or new proteins can be recognized [55]. And here there is a risk that some of the lymphocytes will be able to react with their own antigens, eventually leading to autoimmune disorders. This is especially true for persistent infections and the release of self-proteins, which leads to the development of an immune response against self-epitopes [52]. In the case of immunosuppression of innate and adaptive immunity due to the relationship of infection with immunocompetent cells, the existing basic postulates were expanded with additional markers. Today, the relationship of infectious agents with 30 autoimmune diseases has been established [56, 57].
3.3 Structural and functional features of the hepatobiliary system organs to trigger factors
During the immune reactions formation in response to various antigens in the spleen and liver, activation and redistribution of cellular elements occur, which affects their mass, size, and function. Morphological and functional changes in the spleen also depend on the migratory properties of immunocompetent cells in the spleen compartments. Infectious antigens change the profile of immune cells. The spleen accumulates activated macrophages expressing CD68 receptors; CD4 and CD8 T lymphocytes; CD20 B lymphocytes; CD57 NK cells, which should normally leave the spleen [58]. Immune cells change phenotype and move to the spleen in accordance with the stages of the immune response. The spleen, in addition to activating macrophages and differentiating lymphoid cells, takes part in the formation of antibodies, also destroys erythrocytes and platelets, synthesizes mediators that affect hematopoiesis in the bone marrow, and ensures the maturation of immunocompetent cells [59]. Hyperplasia of the spleen develops with an increase in the phagocytic function of the spleen against the background of autoimmune diseases. The spleen increases against the background of inflammation associated with a violation of the immune system. The presence of an immune response directed against antigens or self-tissues may be a primary independent process in relation to tissue damage. Enlargement of the spleen can be the cause of hypersplenism, that is, sequestration and high destruction of blood cells of two or more cell lines by splenic macrophages. In hypersplenism, cytopenia is observed, and in the bone marrow— enhanced hematopoiesis [44].
The liver contains resident immunocompetent cells, hepatic stellate cells (Ito cells), and resident macrophages (Kupffer cells), which are actively involved in changing the functional state of hepatocytes [60, 61]. Ito cells can induce the development of liver fibrosis under the influence of exogenous factors [62]. Activated fibroblasts act as producers of excess collagen. Fat-storing Ito cells transform into myoblasts and synthesize extracellular collagen. In response to damage by antigens, Kupffer cells produce pro-inflammatory cytokines, which are an additional factor in apoptosis and autophagy of hepatocytes against the background of a large amount of reactive oxygen species (ROS) production. It is the cytokine-producing Kupffer cells that are the activators of this negative process [63, 64].
3.4 Materials and methods in the study of immunological reactions in hepatosplenomegaly
We examined 148 patients with hepatosplenomegaly (HSM), age 22–71 years. Patients, depending on the severity, underwent sclerotherapy of esophageal varices; embolization of the splenic and left gastric arteries, and other methods of surgical treatment. Depending on the etiological factors, all patients with HSM were divided into three groups.
The first group (I) included 74 patients with cirrhosis of the liver against the background of the hepatitis virus, of which 60 people (34 men and 26 women) aged 22 to 71 years with cirrhosis of the liver against the background of the hepatitis C virus (HCV), 6 men aged 49 to 68 years with cirrhosis associated with hepatitis B virus (HBV), and 8 patients (5 men and 3 women) aged 38 to 57 years old with cirrhosis of the liver on the background of combined HCV + HBV infection. The duration of the disease was 2–5 years. The presence of hepatitis B and C was established based on the detection of HBsAg and total anti-HCV antibodies respectively in the blood serum by ELISA.
The second group (II) included 65 people (39 women and 26 men) aged 24 to 81 years. Patients of this group were diagnosed with cirrhosis of the liver of unknown etiology, during which the persistence of herpes viruses—cytomegalovirus (CMV) and Epstein-Barr virus (EBV) was detected. The group did not include patients with lymphoproliferative diseases. The presence of herpes group viruses is detected when anti-infection antibodies to CMV and EBV are detected in the blood serum, in the absence of persistence of hepatitis A, B, C, and D viruses. The response of patients to infectious antigens was heterogeneous. We also detected in patients with liver cirrhosis and hepatosplenomegaly, autoantibodies to carotene-containing root crops were detected, which can be regarded as a trigger factor for this condition [65].
The third group (III) included 9 people (7 women and 2 men) aged 23 to 61 years. In patients, fatty hepatosis was diagnosed; the group was distinguished by frequent recurrent bleeding. In two patients of this group, the manifestation of bleeding began in childhood; this made it possible to suggest the presence of congenital storage diseases in this group patients. Further studies of the activity of lysosomal enzymes made it possible to confirm the presence of congenital genetically determined enzymopathy. Thus, the third group was characterized by hepatosplenomegaly against the background of a genetically determined deficiency of lysosomal enzymes (congenital enzymopathy).
In patients with hepatosplenomegaly, we used the following methods:
ELISA – the presence of viral persistence: hepatitis HBV/HCV, CMV, EBV, Helicobacter pylori; the cytokine profile (IL-1β, IL-2, IL-6, IL-18, IFN-γ, TNF-α, IL-4, IL-10).
Spectrophotometry – the level of various DAMP fractions (λ = 238 nm – oligopeptides, λ = 254 nm – peptides, λ = 260 nm – nucleotides, λ = 280 nm – aromatic amino acids), the content of ceruloplasmin and enzymes.
Immunoturbidimetry – the concentration of C3 and C4 complement components.
Light microscopy – the activity of oxygen-independent and oxygen-dependent phagocytosis; the level of lymphocytotoxicity in Terasaki test.
Fluorescence microscopy – the presence of antiplatelet, antinuclear autoantibodies (ANA), and antineutrophil autoantibodies.
Confocal microscopy – the reactive oxygen species (ROS) production.
Flow cytometry – the state of adaptive immunity—the content of various subpopulations of T and B lymphocytes: B lymphocytes CD22+; T helpers CD3+CD4+; cytotoxic T lymphocytes CD3+CD8+/CD45+; early activation marker of the inflammatory process CD3+CD4+CD25+; regulatory T lymphocytes CD3+CD4+CD25+CD127-, costimulatory molecules CD3+CD4+CD28+, marker of T lymphocytes late activation CD3+HLA-DR+.
Fluorimetry – the determination of enzymes β-glucocerebrosidal, tartrate-resistant acid phosphatase.
Liquid chromatography – the content of amino acids.
3.5 The innate immunity factors in patients with hepatosplenomegaly
Some humoral (complement components) and cellular (activity of phagocytic neutrophils) factors that perform the main barrier function were studied.
In group I (HSM, hepatitis), the concentration of the C3 complement component was 117.0 ± 14.1 mg/dl and did not differ from the reference values (105.0 ± 7.1 mg/dl). There was a decrease in the activity of the C4 component (15.7 ± 3.11 mg/dl and reference values – 25.0 ± 1.12 mg/dl) due to its consumption in opsonization observations.
In group II (HSM, herpes viruses), a significant increase in the activity of the C3 component (up to 153.0 ± 6.3 mg/dL) was revealed, which led to the formation of a membrane attack complex, an additional tissue alteration factor.
The concentration of the C3 complement component in group III (enzymopathies) showed a slight increase (122.0 ± 8.4 mg/dl) against the background of the highest concentration of circulating immune complexes.
In group I was activation of the all stages of oxygen-independent phagocytosis (increased chemotaxis, adhesion, and absorptive capacity of neutrophils, as evidenced by significantly high values of the phagocytic index and phagocytic number) and pathological decrease in phagocyte endocytosis.
An increase in the phagocytic number by 42% indicated a high viral load or bacterial contamination, leading to an increase in the concentration of toxic DAMPs.
With intense adhesive and absorptive capacity of neutrophils, a pronounced pathological decrease (by 57%) in the phagocytosis completion index (0.71 ± 0.02 with a reference level of 1.21 ± 0.12) was revealed – endocytosis of antigens through lysosomal granzymes (Table 1).
Indicators
RV – reference values
Group І (HCV/HBV)
Group ІІ (CMV/VEB)
Group ІІІ (congenital enzymopathy)
Oxygen-independent phagocytosis
Phagocytic index,%
85.0 ± 5.1
98.8 ± 2.1*
91.0 ± 3.2
45.2 ± 2.7*
Phagocytic number
3.62 ± 0.2
5.1 ± 0.1*
4.77 ± 0.3*
2.1 ± 0.3*
Phagocytosis completion index
1.21 ± 0.12
0.71 ± 0.02*
0.95 ± 0.01*
0.8 ± 0.01*
Oxygen-dependent phagocytosis
Spontaneous level of NADPH oxidase reactions (SP, %)
10.1 ± 1.1
44.48 ± 4.3*
29.23 ± 5.6*
25.0 ± 2.5*
Stimulated level of NADPH oxidase reactions (ST, %)
65.2 ± 3.2
66.69 ± 7.2
68.39 ± 5.9
71.5 ± 6.4
Stimulation index (SI=ST/SP)
6.5 ± 0.9
1.39 ± 0.4*
3.82 ± 0.6*
2.7 ± 0.3
Table 1.
Indicators of oxygen-independent and oxygen-dependent phagocytosis in patients with hepatosplenomegaly.
Note: * - the differences are reliable relative to the reference values, p ≤ 0.05.
A pronounced fourfold increase in the spontaneous level of NADPH-oxidase reactions of neutrophils was revealed in group I. Patients with HSM and hepatitis had a decrease in oxygen reserve, that is, an imbalance in the ratio of spontaneous and induced ROS production in NADPH-oxidase reactions of oxygen-dependent phagocytosis (Table 1).
To visualize the processing of antigen in neutrophils, we also used acridine orange dye, which made it possible to monitor the stages of DNA denaturation of the antigen (Saccharomyces cerevisiae cells), in the lysosomes of phagocytic neutrophils (Figure 9A, B).
Figure 9.
Phagocytic activity of neutrophils: A – Native DNA, green color, λmax = 530 nm; B – Denatured DNA (incompleteness of the digestive function in oxygen-independent phagocytosis due to dysfunction of lysosomal enzymes, red color λmax = 640 nm); C – Formation of a neutrophil trap. The fluorescence color changed to red when the acridine orange dye was bound to denatured DNA of model antigen (S. cerevisiae). The intensity of red fluorescence characterized various functional stages of the activity of neutrophils lysosomal enzymes. The transition of double-stranded DNA to single-stranded DNA characterizes the intensive digestion of the antigen by neutrophil lysosomal enzymes, and undigested antigens retained the green luminescence of native DNA. Scale bar 10 μm. Fluorescence microscopy, magnification ×1000.
In some patients of group II, neutrophil traps were detected (Figure 9C).
The number of neutrophil traps in patients of group II was the highest and amounted to 53.5%.
In group II (HSM, herpes viruses) was an excessive activation due to granzyme and mitochondrial enzyme complexes. And the magnitude of induced reactions was low, due to which there was a pathological ratio of spontaneous and induced reactions.
The stimulation index significantly decreased (Table 1).
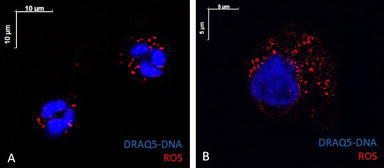
Hyperactivation of the immune response in group II during inflammatory processes led to the development of oxidative stress. The production of reactive oxygen species (ROS) in neutrophils was five times higher than the reference level (measured in relative fluorescence units RFU = 3786.19), which caused increased cell apoptosis (Figure 10).
Figure 10.
Activation of neutrophils NADPH-reactions in oxygen-dependent phagocytosis due to mitochondrial enzyme complexes and massive formation of ROS, which contributed to the further free radical process. A – Intense ROS luminescence on phagocyte membranes during spontaneous unstimulated phagocytosis (scale bar 10 μm). B – A significant activation of the ROS formation after stimulation of NADPH oxidase reactions, an increase in the concentration of ROS on the membranes of the cytoplasmic structures of neutrophils (scale bar 5 μm). Nuclear DNA stained blue (DRAQ5 dye). Reactive oxygen species are stained red. Confocal microscopy.
The production of ROS caused cell apoptosis, an increase in the activity of the neutrophil trap formation, and the activation of the gamma-glutamyl cycle enzymes involved in microsomal oxidation. The maximum increase (5.5 times) in the activity of gamma-glutamyltransferase indicated a violation of the gamma-glutamyl cycle antioxidant function, which is catalyzed by enzyme systems of the hepatocytes endoplasmic reticulum membranes with the participation of cytochrome p450.
Phagocytic reactions of neutrophils in group III (enzymopathy) differed significantly from the two previous groups in low ability to chemotaxis, adhesion, and endocytosis of antigens (Table 1). The failure of all stages of oxygen-independent phagocytosis indicated a defect in lysosomal enzymes, in particular beta-glucocerebrosidal, tartrate-resistant acid phosphatase, due to genetic deficiencies.
A threefold decrease in the catalytic activity of lysosomal enzymes is a diagnostic marker. A decrease in the activity of the enzymes lactate dehydrogenase and cholinesterase was also found.
3.6 Expression of lymphocyte differentiation clusters to enhance intercellular cooperation of immunocompetent cells
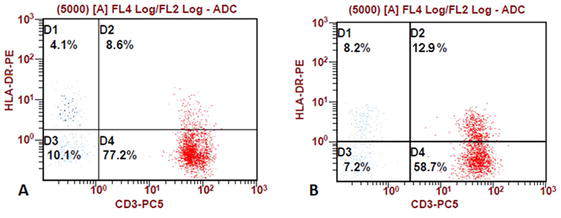
A chronic inflammatory response can be formed by DAMPs in the absence of acute infection or activation of endogenous PAMPs—cellular persisters [52, 66]. The cellular debris formed as a result of destruction, presented in the form of various endogenous cytotoxic DAMPs against the background of insufficiency of cellular and humoral factors of innate immunity, is also the cause of a change in the function of immunocompetent cells and a trigger factor for the autoimmune conditions development [67]. With ineffective reactions of innate immunity and the formation of an irreversible chronic inflammatory process in group I (hepatosplenomegaly, hepatitis), an increase in the expression of T lymphocytes late activation marker CD3+HLA-DR+ by 55% was revealed due to the long-term persistence of HBV and/or HCV viruses (Figure 11).
Figure 11.
Representative flow cytometry plots to determine the percentage of CD3+ T cells expressing HLA-DR. the number of cells with the CD3+HLA-DR+ phenotype is indicated by the D2 quadrant. As an example, individual values of the relative amount of the T lymphocytes late activation marker (CD3+HLA-DR+) are given: in a healthy donor (A), in a patient with hepatosplenomegaly on the background of HBV/HCV (B).
In group I, there was an increase in the concentration of profibrogenic pro-inflammatory cytokines IL-2 (8 times), IL-4 (6 times), IL-6 (18 times), and anti-inflammatory cytokine IL-10 (6 times), which initiate regeneration of destroyed hepatocytes by fibrotic type.
A twofold decrease in the expression of differentiation clusters of CD4+ T helpers and co-stimulating CD3+CD4+CD28+ cells led to a decrease in the activity of antibody-producing CD22+ B lymphocytes (by 13%) and impaired formation of specific clones of plasma cells.
In patients with a high content of the anti-inflammatory cytokine IL-10, the expression of regulatory Treg lymphocytes CD3+CD4+CD25+CD127- was reduced by 2.5 times due to the balance of pro-inflammatory cytokines.
In group I, a maximum twofold increase in the serum concentration of IgA was revealed, as a strong opsonizer and participant in the antigen-antibody-complement complex. This was probably due to the entry of large amounts of IgA contained in the intestinal mucosa through portosystemic anastomoses directly into the bloodstream during abdominal infection [68].
Group II (hepatosplenomegaly, herpes viruses) was the highest titer of antibodies to persistent VEB and CMV and a threefold increase in the number of CD22+ plasma B lymphocytes—antibody producers. Some patients had a high concentration of IgM and IgG, which characterizes the onset of inflammation and the active synthesis of virus-neutralizing antibodies and immune complexes.
Some patients of group II had a compensatory increase in regulatory Treg cells CD3+CD4+CD25+CD127- in response to hyperactivation of pro-inflammatory cytokines.
From the revealed changes, it can be concluded that in group II patients with HSM against the background of herpes infection, humoral reactions prevailed and autoimmune processes were formed.
The high degree of intensity and severity of the inflammatory response in group II was characterized by another change in the profile of immune markers.
An increase in IL-1β (3 times), antiviral chemokine INF-γ (4 times), TNF-α (10 times)—the primary activators of acute inflammatory response —was revealed.
Some patients of group II had a multiple increase in IL-6 (18 times) and IL-18 (2 times). An increase in the synthesis of IL-6 leads to an increased entry of macrophages into the spleen. And an increase in IL-18 levels leads to the activation of macrophages that acquire a pro-inflammatory phenotype. Activated macrophages, together with CD8+ T lymphocytes infiltrate tissues, in particular the bone marrow and liver, and lead to cytopenia, liver dysfunction, and coagulopathies [69].
In group III, hepatosplenomegaly against the background of enzymopathy due to a defect in lysosomal enzymes was a decrease in primary inflammatory activators—IL-1β (2 times) and TNF-α (10 times). Anti-inflammatory IL-6 was increased by 20 times and characterized an increase in the intensity of inflammation against the background of bleeding.
In patients with hereditary enzymopathies, the highest iron content and a twofold increase in the vascular growth factor VEGF was revealed, which, in addition to stimulating angiogenesis, is a chemoattractant of Gaucher cells and an additional factor in their accumulation in the spleen and liver.
Also in group III, against the background of a normal content of serum immunoglobulins, a 1.5-fold increase in the subpopulation of CD22+ B lymphocytes was revealed.
3.7 The cytotoxic factors and DAMP fractions in patients with hepatosplenomegaly
In group I, an increase in the peptide fraction (λ = 254 nm) was revealed due to cytotoxic molecules of cell debris (DAMPs) and pathogen-associated molecular patterns (PAMPs), acting as autoimmunization inducers, as evidenced by the highest level of lymphocytotoxicity (in Terasaki test).
A twofold increase in the nucleotide fraction (λ = 260 nm) of average mass cytotoxic molecules was revealed due to the destruction of both the nuclear and mitochondrial genomes and their release into the intercellular space.
The concentration of ceruloplasmin, a plasma antioxidant, was reduced (by 40%) in patients with HSM on the background of hepatitis B and C (group I).
An increase in lactate and an adaptive fourfold increase in lactate dehydrogenase may also indicate in favor of mitochondrial dysfunction, which indicates a transition to glycolysis.
The increase in the fraction of aromatic amino acids (λ = 280 nm) is due to the high content of phenylalanine and tyrosine detected in the blood serum, which was detected in group II. In contrast to groups I and II, the oligopeptide fraction of average mass molecules (λ = 238 nm) in group III was increased.
The lipid profile of group III was characterized by an increased concentration of total cholesterol and a reduced concentration of high-density lipoproteins, which is characterized by a hereditary decrease in the lipoprotein lipase enzyme.
3.8 The repertoire of autoantibodies in hepatosplenomegaly
All examined patients with hepatosplenomegaly (HSM) had a large repertoire of autoantibodies: antiplatelet, antinuclear, and antineutrophil.
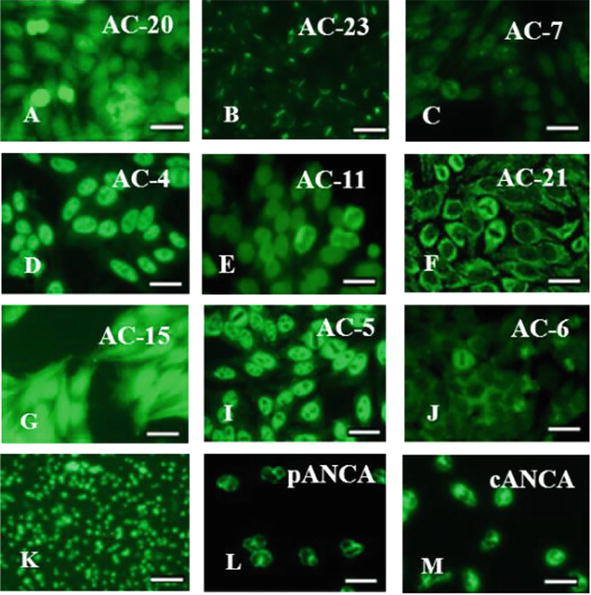
In group I, antinuclear autoantibodies (ANA) of 9 specificities with different frequency of occurrence were detected: to histidyl-tRNA synthetase in 12% (which is associated with antisynthetase syndrome); to dsDNA and histones in 5.9% (marker of high viral load); to centromeres in 5.8%; to Cajal bodies in 5.8% (containing small nuclear ribonucleoproteins involved in the processing of mRNA and histones, the restoration of telomeres, RNA splicing, and are also involved in reactions viral infection); to IMPDH2 (inosine monophosphate dehydrogenase 2) in 11.6%; to tropomyosin in 5.8%; to filamentous structures in 12%; to F-actin in 5.8% (controls and regulates the structural and functional features of the cytoskeleton) (Figure 12). Autoantibodies to Cajal bodies and tropomyosin were typical only for patients of group I.
Figure 12.
Types of autoantibodies in patients with HSM of various etiologies. Antinuclear autoantibodies (ANA): A – To histidyl-tRNA synthetase; B – To filamentous structures; C – To the Cajal bodies; D – To SS-A/Ro, SS-B/La; E – To Lamin B; F – To mitochondria; G – To tropomyosin; I – To U1-RNP; J – To Sp-100, PML proteins; K – Antiplatelet antibodies; L – Perinuclear fluorescence type antineutrophil autoantibodies (pANCA); M – Cytoplasmic fluorescence type antineutrophil autoantibodies (cANCA). Fluorescent microscopy. Scale bar 20 μm. Codes AC (AC – Anti-cell pattern) in accordance with the international consensus on ANA patterns (ICAP) are presented. FITC staining of HEp-2 standard antigenic substrates after interaction with serum antibodies; magnification ×1000.
The spectrum of autoimmune antibodies in group II (antiplatelet, antinuclear, antineutrophil, antimitochondrial, etc.) included 11 specificities.
Patients of group II had autoantibodies to platelets with the maximum frequency (in 95% of patients). The presence of autoantibodies to platelets in patients with HSM indicates their possible pathogenetic role in the occurrence of bleeding and thrombocytopenia. Thrombocytopenia found in group II due to the presence of autoantibodies to platelets, compensatory led to an increase in the production of plasma coagulation factors due to an increase in the concentration of fibrin and fibrinogen, which increase the risk of disseminated intravascular coagulation (DIC) condition.
In group II were detected antibodies to such antigens: dsDNA and histones (5.9%), centromeres (5.9%), histidyl-tRNA synthetase (5.9%), F-actin protein (5.9%), to soluble nuclear ribonucleoproteins SS-A/Ro and SS-B/La (18.2%), to protein MSA-2 (9.3%), to lamin B, PL7 (threonyl-tRNA synthetases) and PL12 (alanyl-tRNA synthetases) (9.1%), to mitochondria (17.6%) (Figure 12).
Since mitochondria are regulators of immunity, they control cell differentiation and induction. In addition, a significant contribution to the development of autoimmune reactions in HSM is made by mitochondrial dysfunction, which can be judged by the presence of antimitochondrial autoantibodies and dysfunction of mitochondrial complexes, leading to disruption of biochemical reactions cascades (accumulation of lactate, transition to glycolysis, disruption of cellular energy metabolism, Krebs cycle, cellular respiration, and β-oxidation).
Also in group II, the number of ANCA-positive patients was higher than in group I, and amounted to 85.7%. Among them, patients with the autoantibodies to the myeloperoxidase (pANCA – 71.4%) and to the proteinase 3 (cANCA – 14.3%) also prevailed (Figure 12).
Such a wide range of autoantibodies is the result of various factors of liver destruction and spleen tissues. Antibodies can have destructive processes; take part in cell killing, exerting a depressing effect on fibrogenic factors.
In group III (congenital enzymopathy), antinuclear antibodies ANA of two specificities were detected: to ribonucleoprotein U1-RNP (14.3%), involved in the initiation of pre-mRNA splicing, and Sp-100 and PML proteins (14.3%), which do not occur with viral etiology of HSM (Figure 12). Autoantibodies to PML proteins participate in transcription and apoptosis, in DNA repair and determining resistance to viruses. And the detected anti-PML autoantibodies prevent the inhibition of IL-6 secretion in this group.
3.9 Markers of autoimmune reactions in the choice of therapeutic targets for hepatosplenomegaly of various etiologies
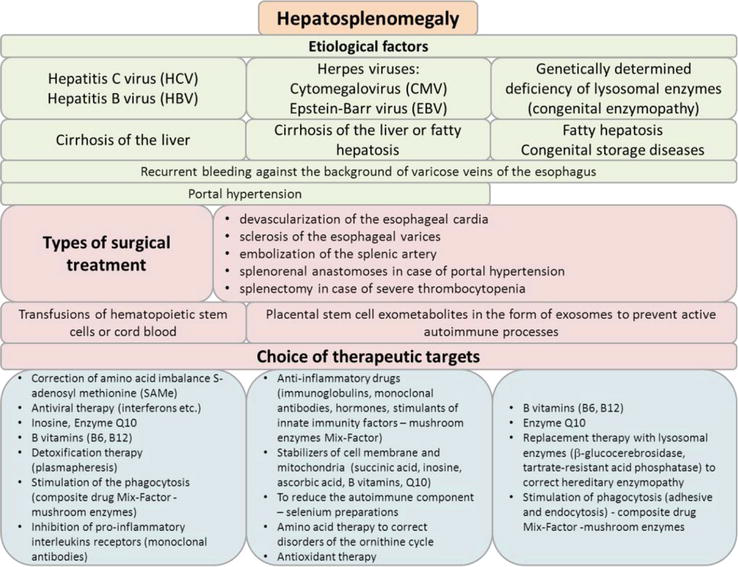
Hepatosplenomegaly (HSM) is characterized by simultaneous enlargement of the spleen and liver in many hepatobiliary diseases. The dynamics of changes in HSM immune processes in response to various antigens occur with the activation and redistribution of cellular elements and changes in the functioning and reactivity primarily of the immunocompetent organ—the spleen [70, 71]. The method of choice for HSM against the background of portal hypertension and recurrent bleeding is splenectomy. But the effectiveness of splenectomy is not always appropriate, since a large number of specialized immune cells are removed from an important peripheral immunocompetent organ, which also performs the function of eliminating antigens.
A differential approach is needed to the choice of treatment tactics for patients in this category, taking into account reconstructive surgeries and targeted immunocorrection. In the Institute clinic, in patients with hepatosplenomegaly complicated by bleeding, the following types of operations are performed: devascularization of the esophageal cardia, sclerosis of the esophageal varices, splenorenal anastomoses in case of portal hypertension, embolization of the splenic artery, and in case of severe thrombocytopenia—splenectomy. Along with surgical treatment to eliminate the threat to the life of the patient due to bleeding, it is advisable to use transfusions of hematopoietic stem cells or cord blood [72] in group I. And in the ІІ and ІІІ groups, it is advisable to use placental stem cell exometabolites in the form of exosomes to prevent active autoimmune processes.
The immunopathological markers found in patients indicate different mechanisms of HSM formation, depending on trigger factors and genetic predictors. Metabolic disorders in patients with HSM complicated by bleeding depend on the severity and duration of the stages of the inflammatory process, and the presence of an autoimmune component.
The identified immunological markers make it possible to substantiate a personalized diagnostic and treatment algorithm using specific antiviral antibodies, amino acids, transfusion of stem cells of various origin, anti-inflammatory cytokines and inhibitors of their receptors, and enzyme replacement therapy.
In group I patients (HSM against the background of HBV/HCV), the following was detected: activation of the secretion of profibrogenic pro-inflammatory cytokines IL-2, IL-4, IL-6; inhibition of the expression of the T lymphocytes early activation marker CD3+CD25+ and induction of the expression of the T lymphocytes late activation marker CD3+HLA-DR+; decrease in the C4 complement component due to its consumption as a cytolytic factor, followed by an increase in the peptide fraction of DAMP (λ = 254 nm); inhibition of phagocytic neutrophils endocytosis, which indicated the development of a chronic irreversible inflammatory reaction. These disorders contributed to the pronounced vascularization of the spleen tissues, which contributed to the development of the HSM syndrome. A high frequency of ANA autoantibodies occurrence, including those to Cajal bodies and tropomyosin, was revealed. In patients with HSM on the background of hepatitis viruses, an increase in the content of straight-chain amino acids and a multiple increase in the concentration of methionine were revealed, which indicates a violation of its metabolism to the end product, cysteine, which was significantly reduced.
For patients of group I, along with the correction of amino acid imbalance (disruption of the methionine cycle), it is advisable to use s-adenosyl methionine (SAMe), inosine, B vitamins (B6, B12), antiviral therapy with the use of interferons and other targeted drugs to support mitochondrial function, enzyme Q10, detoxification therapy (plasmapheresis), drugs to stimulate the adhesive function and endocytosis of phagocytes (composite drug Mix-Factor [73], stimulation of the synthesis of anti-inflammatory cytokines (recombinant IL-10, monoclonal antibodies to receptors of pro-inflammatory interleukins) (Figure 13).
Figure 13.
The strategy of the treatments for patients with hepatosplenomegaly.
In group II, patients in response to the persistence of CMV/EBV viruses, a multiple increase in the secretion of inducers of the inflammatory process first level—INF-γ, ІL-1β, IL-18, TNF-α, and IL-6 was revealed, which reflected the severity of inflammatory reactions; induction of expression of T lymphocytes early activation marker CD3+CD4+CD25+; excessive stimulation of the ROS production and enhancement of NADPH reactions, which contributed to an increase in the frequency of neutrophil traps (NETs) formation; an increase in the cytolytic factor C3 complement component against the background of a multiple increase in the concentration of IL-1β and IL-18, it contributed to the intensive production of ROS and caspase-1-dependent cell pyroptosis. Patients in this group showed a high content of the cyclic amino acids fraction in the blood serum at λ = 280 nm. And a different pattern of amino acid metabolism disorders was found, which was expressed in a significant increase in the titer of aromatic amino acids (phenylalanine, tyrosine), and a violation of the ornithine cycle. All patients of group II had antiplatelet antibodies, antineutrophil antibodies, and a wide range of antinuclear autoantibodies (ANA), including those to nuclear and mitochondrial nucleotides, causing mitochondrial dysfunction.
For patients of group II, it is advisable to use anti-inflammatory drugs (hormones, immunoglobulins, monoclonal antibodies, stimulants of innate immunity factors – Mix-Factor), cell membrane stabilizers (succinic acid, inosine, ascorbic acid, B vitamins), to reduce the autoimmune component–selenium preparations, therapy with amino acid preparations to correct disorders of the ornithine cycle, antioxidant therapy, stimulators of differentiation of the CD8+ killer subpopulation and CD16+ natural killers (Figure 13).
In group III, there was a lack of enzymes activity—β-glucocerebrosidase, tartrate-resistant acid phosphatase, and a pronounced decrease in lactate dehydrogenase and cholinesterase. Against the background of enzymopathy, all stages of oxygen-dependent and oxygen-independent phagocytosis were inhibited due to a defect in lysosomal enzymes; an increase in the concentration of inflammatory activators IL-1 and TNF-α was revealed. Pro-inflammatory IL-6 was elevated and characterized an increase in inflammation against the background of bleeding, and due to the development of an adaptive response, endothelial growth factor VEGF was increased. There was an increase in the DAMP oligopeptide fraction (λ = 238 nm), and a multiple increase in the serum iron content.
For patients of group III against the background of frequent recurrent bleeding—replacement therapy with lysosomal enzymes (beta-glucocerebrosidase, tartrate-resistant acid phosphatase) to correct hereditary enzymopathy, stimulation of phagocytosis (adhesive and digesting properties of phagocytes), it is advisable to use the composite drug Mix-Factor (Figure 13).
Thus, most patients with hepatosplenomegaly syndrome of varying severity, as a rule, require emergency surgical interventions. But it is necessary to take into account the presence of trigger factors and immunophysiological disorders in order to develop a comprehensive treatment and diagnostic algorithm, including targeted therapy of immune checkpoints, which will be the best option to prevent recurrence of bleeding in patients with hepatosplenomegaly.
Hepatosplenomegaly syndrome and myasthenia gravis are multifactorial diseases with an autoimmune component in pathogenesis. In these pathologies, immunocompetent organs are damaged. Myasthenia gravis is often accompanied by damage of the immunity central organ—the thymus, and structural and functional changes in the immunity peripheral organ—spleen, are associated with various diseases of the hepatobiliary system.
Myasthenia gravis and hepatosplenomegaly, which are often accompanied by portal vein hypertension and recurrent bleeding, are serious diseases. And their conservative treatment is not always effective and often these patients require emergency surgical care (thymectomy and splenectomy). Removal of immunocompetent organs may have negative consequences.
We have revealed trigger factors and predictors of autoimmune reactions in patients with various clinical phenotypes of myasthenia gravis and hepatosplenomegaly. The specific markers can be used to develop new treatment and diagnostic protocols and individual approaches to targeted therapy.
The identified specific markers characterize changes in the immune response and make it possible to choose individual treatment tactics and selectively apply surgical intervention in patients with thymus-independent myasthenia gravis, including certain HLA-DR polymorphism and a specific repertoire of autoantibodies; with splenomegaly— to correct autoimmune processes and excessive cellular proliferation in the spleen, and in the genetically determined orphan storage diseases to carry out enzyme replacement therapy.
1.Sharif K, Watad A, Coplan L, Lichtbroun B, Krosser A, Lichtbroun M, et al. The role of stress in the mosaic of autoimmunity: An overlooked association. Autoimmunity Reviews. 2018;17(10):967-983. DOI: 10.1016/j.autrev.2018.04.005
2.Mathis D, Benoist C. Aire. Annual Review of Immunology. 2009;27:1,287-1,312. DOI: 10.1146/annurev.immunol.25.022106.141532
3.Giraud M, Beaurain G, Yamamoto AM, Eymard B, Tranchant C, Gajdos P, et al. Linkage of HLA to myasthenia gravis and genetic heterogeneity depending on anti-titin antibodies. Neurology. 2001;57(9):1555-1560. DOI: 10.1212/wnl.57.9.1555
4.Bach JF. The etiology of autoimmune diseases: The case of myasthenia gravis. Annals of the New York Academy of Sciences. 2012;1274:33-39. DOI: 10.1111/j.1749-6632.2012.06774.x
5.Barreto R, Fagundes C, Guevara M, Solà E, Pereira G, Rodríguez E, et al. Type-1 hepatorenal syndrome associated with infections in cirrhosis: Natural history, outcome of kidney function, and survival. Hepatology. 2014;59(4):1505-1513. DOI: 10.1002/hep.26687
6.Scarpino S, Di Napoli A, Stoppacciaro A, Antonelli M, Pilozzi E, Chiarle R, et al. Expression of autoimmune regulator gene (AIRE) and T regulatory cells in human thymomas. Clinical and Experimental Immunology. 2007;149(3):504-512. DOI: 10.1111/j.1365-2249.2007.03442.x
7.Peterson P, Org T, Rebane A. Transcriptional regulation by AIRE: Molecular mechanisms of central tolerance. Nature Reviews. Immunology. 2008;8(12):948-957. DOI: 10.1038/nri2450
8.Anderson MS, Venanzi ES, Chen Z, Berzins SP, Benoist C, Mathis D. The cellular mechanism of Aire control of T cell tolerance. Immunity. 2005;23(2):227-239. DOI: 10.1016/j.immuni.2005.07.005
9.Xing Y, Hogquist KA. T-cell tolerance: Central and peripheral. Cold Spring Harbor Perspectives in Biology. 2012;4(6):a006957. DOI: 10.1101/cshperspect.a006957
10.Lindstrom J, Anand R, Gerzanich V, Peng X, Wang F, Wells G. Structure and function of neuronal nicotinic acetylcholine receptors. Progress in Brain Research. 1996;109:125-137. DOI: 10.1016/s0079-6123(08)62094-4
11.Conti-Fine BM, Milani M, Kaminski HJ. Myasthenia gravis: Past, present, and future. The Journal of Clinical Investigation. 2006;116(11):2843-2854. DOI: 10.1172/JCI29894
12.Berrih-Aknin S, Frenkian-Cuvelier M, Eymard B. Diagnostic and clinical classification of autoimmune myasthenia gravis. Journal of Autoimmunity. 2014;48-49:143-148. DOI: 10.1016/j.jaut.2014.01.003
13.Drachman DB. Myasthenia gravis. The New England Journal of Medicine. 1994;330:1797-1810. DOI: 10.1056/nejm199406233302507
14.Vincent A. Unravelling the pathogenesis of myasthenia gravis. Nature Reviews. Immunology. 2002;2(10):797-804. DOI: 10.1038/nri916
15.Verschuuren JJ, Huijbers MG, Plomp JJ, Niks EH, Molenaar PC, Martinez-Martinez P, et al. Pathophysiology of myasthenia gravis with antibodies to the acetylcholine receptor, muscle-specific kinase and low-density lipoprotein receptor-related protein 4. Autoimmunity Reviews. 2013;12(9):918-923. DOI: 10.1016/j.autrev.2013.03.001
16.Stergiou C, Lazaridis K, Zouvelou V, Tzartos J, Mantegazza R, Antozzi C, et al. Titin antibodies in "seronegative" myasthenia gravis-a new role for an old antigen. Journal of Neuroimmunology. 2016;292:108-115. DOI: 10.1016/j.jneuroim.2016.01.018
17.Gasperi C, Melms A, Schoser B, Zhang Y, Meltoranta J, Risson V, et al. Anti-agrin autoantibodies in myasthenia gravis. Neurology. 2014;82(22):1976-1983. DOI: 10.1212/WNL.0000000000000478
18.Lazaridis K, Tzartos SJ. Autoantibody specificities in myasthenia gravis; implications for improved diagnostics and therapeutics. Frontiers in Immunology. 2020;11:212. DOI: 10.3389/fimmu.2020.00212
19.Savino W, Dardenne M. Neuroendocrine control of thymus physiology. Endocrine Reviews. 2000;21(4):412-443. DOI: 10.1210/edrv.21.4.0402
20.Armstrong SM, Schumann L. Myasthenia gravis: Diagnosis and treatment. Journal of the American Academy of Nurse Practitioners. 2003;15(2):72-78. DOI: 10.1111/j.1745-7599.2003.tb00354.x
21.Melzer N, Ruck T, Fuhr P, Gold R, Hohlfeld R, Marx A, et al. Clinical features, pathogenesis, and treatment of myasthenia gravis: A supplement to the guidelines of the German neurological society. Journal of Neurology. 2016;263:1473-1494. DOI: 10.1007/s00415-016-8045-z
22.Aydin Y, Ulas AB, Mutlu V, Colak A, Eroglu A. Thymectomy in myasthenia gravis. The Eurasian Journal of Medicine. 2017;49:48-52. DOI: 10.5152/eurasianjmed.2017.17009
23.Farmakidis C., Pasnoor M., Dimachkie MM., Barohn R.J. Treatment of myasthenia gravis. Neurologic Clinics 2018; 36(2): 311-337. DOI:10.1016/j.ncl.2018.01.011
24.Watanabe A, Watanabe T, Obama T, Mawatari T, Ohsava H, Ichmiya Y, et al. Prognostic factors for myasthenic crisis after transsternal thymectomy in patients with myasthenia gravis. Journal of thoracic and cardiovascular surgery. 2004;127(3):868-876. DOI: 10.1016/j.jtcvs.2003.07.036
25.Bojko VV, Klimova EM, Kudrevich AN. Lechenie Miastenii s Uchetom Immunofiziologicheskih Fenotipov [Treatment of Myasthenia Gravis Taking into Account Immunophysiological Phenotypes]. Kharkov: Publishing House Sheinina E.V. 2008; p. 424
26.Klimova EM, Lavinskaya EV, Minukhin DV, Syrovaya AO, Drozdova LA, Samoilova AP, et al. On forming central and peripheral markers of self-tolerance loss in diverse clinical Myasthenic phenotypes. Der Pharmacia Lettre. 2017;9(6):8-17. Available from: http://scholarsresearchlibrary.com/archive.html
27.Sfriso P, Ghirardello A, Botsios C, Tonon M, Zen M, Bassi N, et al. Infections and autoimmunity: The multifaceted relationship. Journal of Leukocyte Biology. 2010;87(3):385-395. DOI: 10.1189/jlb.0709517
28.Leopardi V, Chang Y-M, Pham A, Luo J, Garden OA. Systematic review of the potential implication of infectious agents in myasthenia gravis. Frontiers in Neurology. 2021;12:618021. DOI: 10.3389/fneur.2021.618021
29.Eckl-Dorna J, Pree I, Reisinger J, Marth K, Chen K-W, Vrtala S, et al. The majority of allergen-specific IgE in the blood of allergic patients does not originate from blood-derived B cells or plasma cells. Clinical and Experimental Allergy. 2012;42(9):1347-1355. DOI: 10.1111/j.1365-2222.2012.04030.x
30.Kumar H, Kawai T, Akira S. Pathogen recognition by the innate immune system. International Reviews of Immunology. 2011;30(1):16-34. DOI: 10.3109/08830185.2010.529976
31.Skok MV, Koval LM, Lykhmus OY, Kalashnyk OM, Gergalova GL, Komisarenko SV. Nicotinic acetylcholine receptors: Specific antibodies and functions in humoral immunity Ukr. The Biochemical Journal. 2013;85(6):134-143. DOI: 10.15407/ubj85.06.134
32.Hurst R, Rollema H, Bertrand D. Nicotinic acetylcholine receptors: From basic science to therapeutics. Pharmacology & Therapeutics. 2013;137(1):22-54. DOI: 10.1016/j.pharmthera.2012.08.012
33.Bodis G, Toth V, Schwarting A. Role of human leukocyte antigens (HLA) in autoimmune diseases. Rheumatology and Therapy. 2018;5:5-20. DOI: 10.1007/s40744-018-0100-z
34.Choo SY. The HLA system: Genetics, immunology, clinical testing, and clinical implications. Yonsei Medical Journal. 2007;48(1):11-23. DOI: 10.3349/ymj.2007.48.1.11
35.Gough SC, Simmonds MJ. The HLA region and autoimmune disease: Associations and mechanisms of action. Current Genomics. 2007;8(7):453-465. DOI: 10.2174/138920207783591690
36.Colbert RA, Tran TM, Layh-Schmitt G. HLA-B27 misfolding and ankylosing spondylitis. Molecular Immunology. 2014;57(1):44-51. DOI: 10.1016/j.molimm.2013.07.013
37.Giraud M, Vandiedonck C, Garchon HJ. Genetic factors in autoimmune myasthenia gravis. Annals of the New York Academy of Sciences. 2008;1132:180-192. DOI: 10.1196/annals.1405.027
38.Franciotta D, Cuccia M, Dondi E, Piccolo G, Cosi V. Polymorphic markers in MHC class II/III region: A study on Italian patients with myasthenia gravis. Journal of the Neurological Sciences. 2001;190(1-2):11-16. DOI: 10.1016/s0022-510x(01)00573-1
39.Baecher-Allan C, Hafler DA. Suppressor T cells in human diseases. The Journal of Experimental Medicine. 2004;200(3):273-276. DOI: 10.1084/jem.20040812
41.Comacchio GM, Marulli G, Mammana M, Natale G, Schiavon M, Rea F. Surgical decision making: Thymoma and myasthenia gravis. Thoracic Surgery Clinics. 2019;29(2):203-213. DOI: 10.1016/j.thorsurg.2018.12.007
42.Klimova EM, Minuchin DV, Drozdova LA, Lavinskaya EV, Kordon TI, Kalashnykova YV. Features of self-tolerance loss in patients with different clinical phenotypes of myasthenia. Regulatory Mechanisms in Biosystems. 2018;9(4):561-567. DOI: 10.15421/021884
43.Klimova EM, Drozdova LA, Lavinskaya EV, Minukhin DV, Kudrevych IO, Kudrevych OM. Genomic and epigenomic predictors for various clinical phenotypes of myasthenia gravis. Wiadomości Lekarskie. 2021;LXXIV(3):1. DOI: 10.36740/WLek202103117
44.Bronte V, Pittet MJ. The spleen in local and systemic regulation of immunity. Immunity. 2013;39(5):806-818. DOI: 10.1016/j.immuni.2013.10.010
45.Mihm S. Danger-associated molecular patterns (DAMPs): Molecular triggers for sterile inflammation in the liver. International Journal of Molecular Sciences. 2018;19(10):3104. DOI: 10.3390/ijms19103104
46.Chentoufi AA, Serov YA, Alazmi M, Baba K. Immune components of liver damage associated with connective tissue diseases. Journal of Clinical and Translational Hepatology. 2014;2(1):37-44. DOI: 10.14218/JCTH.2014.00001
47.Mogensen TH. Pathogen recognition and inflammatory signaling in innate immune defenses. Clinical Microbiology Reviews. 2009;22(2):240-273. DOI: 10.1128/CMR.00046-08
48.Wu MY, Kao WY, Chan CY, Yiang GT, Liao WT, Chen CS. Spontaneous splenic rupture as a rare initial presentation in an acute lymphoblastic Leukemia patient. Diagnostics (Basel). 2019;9(4):152. DOI: 10.3390/diagnostics9040152
49.Wong JL, Arango-Viana JC, Squires T. Heart, liver and spleen pathology in chronic alcohol and drug users. Journal of Forensic and Legal Medicine. 2008;15(3):141-147. DOI: 10.1016/j.jflm.2007.07.011
50.Torimura T. Spleen and liver. The Liver in Systemic Diseases. 2016;3:35-50. DOI: 10.1007/978-4-431-55790-6_3
51.Klimova EM, Bozhkov AI, Lavinska OV, Drozdova LA, Kurhuzova NI. Low molecular weight cytotoxic components (DAMPs) form the post-COVID-19 syndrome. Immunobiology. 2023;228(1):152316. DOI: 10.1016/j.imbio.2022.152316
52.Vanderlugt CL, Miller SD. Epitope spreading in immune-mediated diseases: Implications for immunotherapy. Nature Reviews. Immunology. 2002;2(2):85-95. DOI: 10.1038/nri724
53.Martinecz A, zur Wiesch PA. Estimating treatment prolongation for persistent infections. Pathogens and Disease. 2018;76, 6:fty065. DOI: 10.1093/femspd/fty065
54.Koehli S, Naeher D, Galati-Fournier V, Zehn D, Palmer E. Optimal T-cell receptor affinity for inducing autoimmunity. Proceedings of the National Academy of Sciences USA. 2014;111:17248-17253. DOI: 10.1073/pnas.1402724111
55.Mahanty S, Prigent A, Garraud O. Immunogenicity of infectious pathogens and vaccine antigens. BMC Immunology. 2015;16:31. DOI: 10.1186/s12865-015-0095-y
56.Wucherpfennig KW. Mechanisms for the induction of autoimmunity by infectious agents. The Journal of Clinical Investigation. 2001;108(8):1097-1104. DOI: 10.1172/JCI14235
57.Ercolini AM, Miller SD. The role of infections in autoimmune disease. Clinical and Experimental Immunology. 2009;155(1):1-15. DOI: 10.1111/j.1365-2249.2008.03834.x
58.Lv Y, Lau WY, Li Y, Deng J, Han X, Gong X, et al. Hypersplenism: History and current status. Experimental and Therapeutic Medicine. 2016;12(4):2377-2382. DOI: 10.3892/etm.2016.3683
59.Lewis SM, Williams A, Eisenbarth SC. Structure and function of the immune system in the spleen. Science Immunology. 2019;4(33):eaau6085. DOI: 10.1126/sciimmunol.aau6085
60.Zheng M, Tian Z. Liver-mediated adaptive immune tolerance. Frontiers in Immunology. 2019;10:2525. DOI: 10.3389/fimmu.2019.02525
61.Dhar D, Baglieri J, Kisseleva T, Brenner DA. Mechanisms of liver fibrosis and its role in liver cancer. Experimental Biology and Medicine (Maywood, N.J.). 2020;245(2):96-108. DOI: 10.1177/1535370219898141
62.Bozhkov AI, Novikova AV, Klimova EM, Ionov IA, Akzhyhitov RA, Kurhuzova NI, et al. Vitamin a reduces the mortality of animals with induced liver fibrosis by providing a multi-level body Defense system. Journal of Clinical and Experimental Hepatology. 2023;13(1):48-63. DOI: 10.1016/j.jceh.2022.09.006
63.Bennett H, Troutman TD, Sakai M, Glass CK. Epigenetic regulation of Kupffer cell function in health and disease. Frontiers in Immunology. 2021;11:609618. DOI: 10.3389/fimmu.2020.609618
64.Klimova EM, Drozdova LA, Lavinska OV, Bychenko EA, Kot YH, Kordon TI. Hepatosplenomegaly in liver cirrhosis is caused by reactive oxygen species formation, an increase in apoptosis and autophagy, and pronounced autoimmune reactions. Regulatory Mechanisms in Biosystems. 2021;12(3):408-418. DOI: 10.15421/022156
65.Klimova EM, Kalashnikova YV, Merezhko OS, Kordon TI. Individual correction of the treatment of patients with hepatosplenomegaly syndrome, taking into account the parameters of lipid and protein metabolism. Kharkiv Surgical School. 2020;5-6:14-21. DOI: 10.37699/2308-7005.5-6.2020.03
66.Chen L, Deng H, Cui H, Fang J, Zuo Z, Deng J, et al. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget. 2017;9(6):7204-7218. DOI: 10.18632/oncotarget.23208
67.Roh JS, Sohn DH. Damage-associated molecular patterns in inflammatory diseases. Immune Network. 2018;18(4):e27. DOI: 10.4110/in.2018.18.e27
68.Klimova OM, Kordon TI, Sushkov SV, Drozdova LA, Lavinska OV, Merezhko OS, et al. Features of the immunoreactivity T and B lymphocytes subpopulations and cytokine imbalance in patients with hepatosplenomegaly of different etiology. Pathologia. 2021;18(2):174-182. DOI: 10.14739/2310-1237.2021.2.229454
69.Grant S, Schulert A, Grom A. Macrophage activation syndrome and cytokine directed therapies. Best Practice & Research. Clinical Rheumatology. 2014;28(2):277-292. DOI: 10.1016/j.berh.2014.03.002
70.Prasad AS, Halsted JA, Nadimi M. Syndrome of iron deficiency anemia, hepatosplenomegaly, hypogonadism, dwarfism and geophagia. The American Journal of Medicine. 1961;31:532-546. DOI: 10.1016/0002-9343(61)90137-1
71.Liu P, Li P, He W, Zhao LQ. Liver and spleen volume variations in patients with hepatic fibrosis. World Journal of Gastroenterology. 2009;15(26):3298-3302. DOI: 10.3748/wjg.15.3298
72.Boiko VV, Klymova OM, Vasylovska SV, Sushkov SV, Kordon TI, Shakina LO. Method for treatment of patients with the hepatocirrhosis and prevention of its complications. Pat. No. 37688, A61K 45/00. Published 10.12.2008, Bulletin No. 23. 2008:1-6
73.Bozhkov A, Ivanov I, Klimova E, Kurguzova N, Bozhkov A, Goltvyanskiy A, et al. “Mix-factor” is involved in the regulation of the organism’s redox systems in the late stages of ontogenesis and affects the lifespan of animals. Ageing and Longevity. 2021;2(2):24-36. DOI: 10.47855/jal9020-2021-2-4
Written By
Elena Klimova, Larisa Drozdova, Olena Lavinska, Sergey Sushkov and Valery Boyko
Submitted: 24 May 2023Reviewed: 04 August 2023Published: 15 December 2023
 Open access peer-reviewed chapter
Open access peer-reviewed chapter