Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
Increasing population warrants increasing food crop productivity with a minimum input cost. The usage of inorganic fertilizers is inevitable in modern agriculture cropping systems. Nitrogen, phosphorus, and potassium are the major nutrients used by agriculturalists worldwide since the effect of these nutrients is highly significant on crop productivity; therefore, it is crucial to use them in an optimized way to make farming economically sustainable. The capacity of crops to absorb nutrients and efficiently utilize them is known as nutrient use efficiency (NtUE). The NtUE of plants would reduce the usage of synthetic fertilizers, reduce the nutrient leaches into the environment, and increase crop productivity. The development of optimum biomass-producing and nutrient-efficient crop varieties are the key for rational agriculture.
Centre for Plant breeding and Genetics, Tamil Nadu Agricultural University (TNAU), Coimbatore, Tamil Nadu, India
Sumaiya Sulthana
Centre for Plant breeding and Genetics, Tamil Nadu Agricultural University (TNAU), Coimbatore, Tamil Nadu, India
Balaji Kannan
Department of Physical Sciences and Information Technology, TNAU, Coimbatore, Tamil Nadu, India
Sivakumar Rathinavelu
Department of Crop Physiology, CMS, TNAU, Coimbatore, Tamil Nadu, India
Anandhi Krishnan
Department of Pulses, CPBG, TNAU, Coimbatore, Tamil Nadu, India
Rajaprakasam Sudhagar*
CPBG, TNAU, Coimbatore, Tamil Nadu, India
*Address all correspondence to: sudhagar.r@tnau.ac.in
1. Introduction
To cater to the calorie requirement of the exploding population, especially in Asia, where legumes continue to be the main source of protein, the thrust to develop better-yielding, nutrient-rich, resource-use efficient, and stress-tolerant cultivars is increasing. Fertilizer application is required to augment the nutrient status of soils because the majority of arable soils are low in important nutrient(s). Most legumes are grown as rainfed crops in Asia, where inputs including water are used either in a minimal amount or not at all. Over time, after successful domestication and subsequent crop improvement activities, the nutrient use efficiency of legumes is altered. Enhanced NtUE in legume varieties is considered crucial, which otherwise reduces the production costs. Cultivars with high NtUE will protect the environment by minimizing the need for synthetic/inorganic fertilizers, lowering the rate of nutrient leaching into the ecosystems, increasing yield, and ensuring agricultural sustainability while preserving the quality of the soil and groundwater. The plant has the capacity to absorb and store nutrients in significant quantities. The presence of sufficient quantities of nutrients helps the plants combat the challenges posed by various stresses. Further, genotypes with good NtUE expand the area under legume production, which is otherwise classified as minimal-nutrient-available marginal lands. These lands are nevertheless amenable to cultivation. Genetic improvement for NtUE therefore is considered an important target in legume breeding because it has the potential to significantly enhance production and sustain the effects of climate change.
The perspective of NtUE varies according to the discipline. At each level, the input and the intended output vary, and different terminologies are used. Agro-ecologists, agronomists, and physiologists/breeders/biotechnologists are the major discipline of scientists involved in the NtUE research. Agroecologists perceive the agricultural system, encompassing associated ecosystems, as having a specific NtUE in relation to the agricultural goods it produces. Agronomists focus on the field and concentrate on factors such as yield, which is influenced by applied seeds; fertilizers; pesticides; and other inputs. They also include manpower costs for the specific area of land. Plant breeders/physiologists/biotechnologists, in their perspective, consider that the plant has an inherent NtUE, which is influenced by varied internal/external stimuli. In this chapter, we approach the NtUE from a breeder/physiologist/biotechnologist perspective.
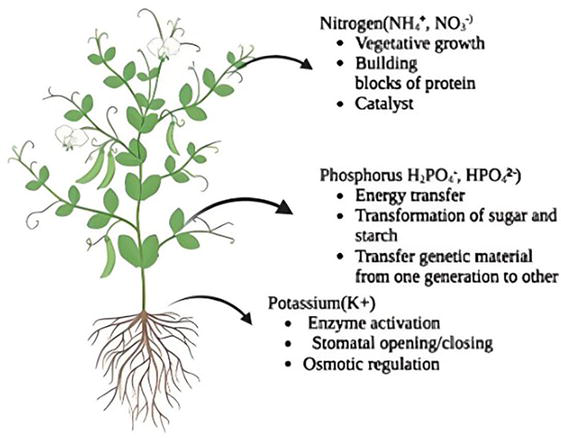
The nutrients nitrogen (N), potassium (K), and phosphorus (P) regulate plant growth and metabolism in significant ways and therefore are called macronutrients. Nitrogen, a key constituent of nucleic acids, and alkaloids regulate protein and chlorophyll synthesis. N, in association with a few other minor nutrients, improves the water and nutrient absorption efficiency of roots [1]. Potassium (K) plays an important role in water regulation, osmotic balance, enzymatic activation, stomatal opening and closing, plant defense, stress tolerance, yield, and quality improvement. Phosphorus (P) regulates many plant functions: (i) energy transfer; (ii) photosynthesis; (iii) nutrient, sugar, and starch transport; and (iv) character inheritance (Figure 1).
Figure 1.
Role of major nutrients in plants. (created with https://www.biorender.com).
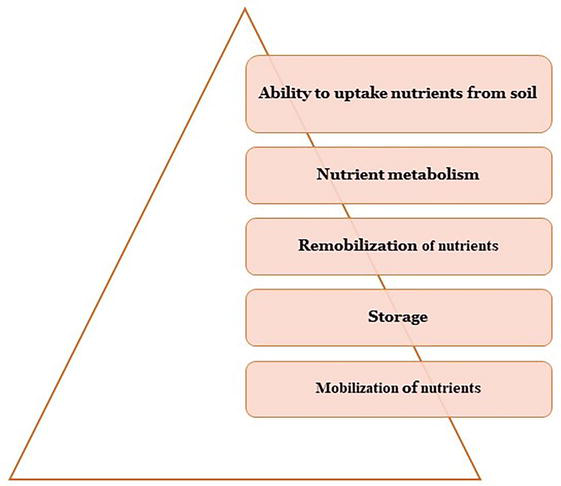
The NtUE represents the ability of plants to absorb and utilize nutrients to maximize yield. The NtUE involves three crucial plant processes: nutrient absorption, assimilation, and utilization. The key factors determining the NtUE are presented in Figure 2.
Figure 2.
Factors determining NtUE in legumes.
For the effective utilization of applied and accessible nutrients, plants primarily employ a few strategies like (i) increased root growth, (ii) changing the rhizosphere’s characteristics to increase nutrient availability, (iii) altering root architecture in all possible ways to harbor significant interactions between rhizosphere microbial populations, and (iv) regulating the expression of ion transporters.
4. Internal and external factors that affect nutrient use efficiency
The NtUE depends on both internal (plant) and external factors. Fertilizer-dependent factors, climate variables, microbial population, and other agronomic practices are examples of external factors. Internal factors include genetic, physiological, and biochemical regulations (Table 1).
Internal factors
External factors
Genetic factors:
Types of species/cultivar/genotypes
Physiological factors:
Length of the root
The density of root hair
Mobilization of nutrients within the plant
Presence of ion transporters.
Biochemical factors:
Availability of enzymes like nitrate reductase (N), phosphatase (P), etc. for the conversion of nutrients
Root exudates: citric acid, malic acid, and trans-aconitic acid
Fertilizer-dependent factors:
Source of fertilizer
Time of application
Depth and method of application
When applied in combination
Use of slow-release form.
Climatic factors:
Soil moisture
Temperature
Rainfall
Erosion
Microbial population:
Arbuscular mycorrhizae and other beneficial soil microbes
NUE is the productivity ability of plants in a unit of land with the available nitrogen (both naturally occurring and supplemented by N fertilizer) [2]. NUE is the result of (i) utilization efficiency (yield/absorbed N) and (ii) absorption efficiency (it is the ratio between absorbed and available N). Knowledge of the different forms of nitrogen that are preferred by the plants for uptake, enzymes involved in the conversion processes, transporters involved in N transport, and gene expression studies will help the breeders to formulate effective strategies to improve the NUE.
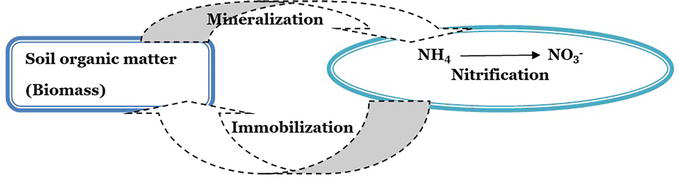
Forms of N and their conversion: N fertilizer supplies different forms of nitrogen upon soil application, and such forms are narrated in Table 2. Regardless of the source of N applied to soil, plants prefer the nitrate form. Excess application of N forms leads to certain environmental concerns (Figure 3).
Organic N
Ammonium N
Nitrate N
Plant Use
Not used
Usable
Usable
Loss through
Mineralization, erosion
Volatilization, erosion
Leaching and denitrification
Changes
Mineralization
Immobilization, nitrification
Immobilization, denitrification
Table 2.
Different forms of nitrogen.
Figure 3.
Conversion of different forms of nitrogen.
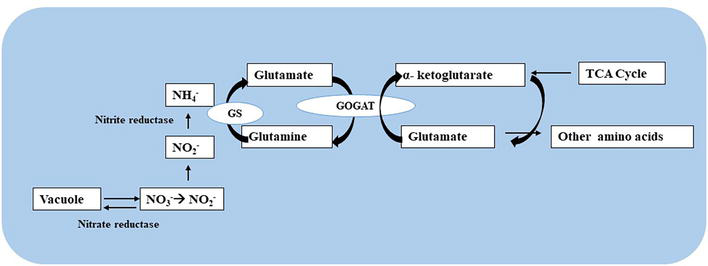
Enzymes involved in N assimilation: Plants with the help of certain transporters absorb these different forms of nitrogen, and a few enzymes support plants to use them. Nitrate reductase, nitrogenase, nitrite reductase, glutamate synthase, and glutamine synthetase are the primary enzymes associated with the reduction of nitrate and assimilation of ammonia. Ammonium is the main source of inorganic N in plants, which is converted into the organic molecules glutamine and glutamate by the enzymes GS (Glutamine synthase) and GOGAT (Glutamine synthetase-glutamate synthase). In plants, GS and GOGAT are present in several isoenzyme forms and distinctively expressed depending on the developmental stage of cells. The GS enzyme has two different forms: (i) a cytosolic form (GS1) found in phloem cells, roots, and leaves and (ii) a plastidic form (GS2) present in the chloroplast and mitochondria tissues used for photosynthesis. Each GS isoform plays unique roles in photo-respiratory, NH3 (ammonia) assimilation, No3− (nitrate) reduction, N translocation, and recycling. Variations in the proportions of GS1 and GS2 are noticed within plant organs and between plants. The enzyme GOGAT has two isoforms, each of which plays a specific role in the primary assimilation or recycling of N. A ferredoxin-dependent isoenzyme (Fd-GOGAT) is primarily involved in the re-assimilation of photo-respiratory ammonia, working in tandem with GS2. Glutamate synthesis (in photosynthetic and/or non-photosynthetic tissues) is regulated by a pyridine nucleotide-dependent isoform (NADH-GOGAT) (Figure 4).
Figure 4.
Nitrate assimilation.
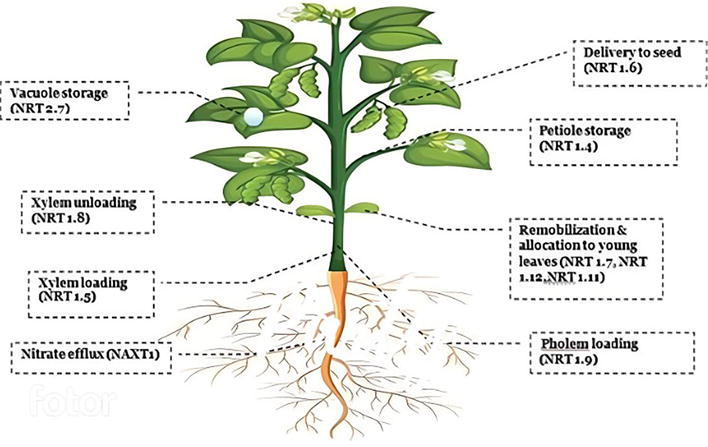
N transporters: To adapt to changes in soil N, plants must have highly developed absorption and signaling mechanisms. After the absorption and conversion of nutrients to available forms by the plants, the next important process will be the proper distribution of available nutrients to the plant parts for their conversion into biomass and grains (from source to sink) [3]. Certain transporters will also regulate this distribution of nutrients within the plants. Nitrate transporters are primarily responsible for nitrate absorption and translocation to the needy tissues [2, 4]. Transport and portioning of N are aided by xylem and phloem cells, respectively. Nitrate activates (i) transporter family genes like NRT1 and NRT2 and (ii) the assimilation pathway genes nitrate reductase (NR) and nitrite reductase (NiR) (Figure 5) [5].
Figure 5.
Types of N transporters. (image adapted from https://www.freepik.com/).
Gene expression studies: Nitrate activates a few genes very quickly (within minutes) without the need for protein synthesis, which is referred to as “Primary Nitrate Response” (PNR) [6]. A number of kinases, transcription factors, and non-coding RNAs [4] control PNR. Nitrate is a signaling molecule that is essential for the development and growth of plants. Legumes have an advantage over non-legume plants in that they can fix nitrogen through the symbiotic interaction between N-fixing diazotrophs, requiring less nitrogen fertilizer [5]. Following the identification of a structural and/or regulatory gene connected to metabolic pathway regulation, information can be gained by creating over-expressers or choosing mutants with the gene of interest. Because legume leaves contain more nitrogen (N), there is higher photosynthesis as a result.
Phosphorus (P) is the macronutrient that regulates plant growth by its association with several metabolic cycles, nucleic acid synthesis, and the modulation of several enzyme activities. P is incorporated with enzymes, nucleic acids, phospholipids, phosphor-proteins, sugar phosphates, and adenosine triphosphate (ATP). P also supports the movement of other nutrients by providing energy to cell membranes through ATP molecules. Sufficient P is required for root development and its enhanced functions (Figure 6), which support crop sustainability in adverse situations. Up to 80% of soil-applied phosphorus will be inaccessible to plants because phosphorus is mostly available in the immobile state, which restricts plant growth and development. Access to sufficient P fertilizer is a barrier because the reserves of rock phosphate are localized and region-specific [7]. Legumes require a high P because symbiotic N fixation in nodules consumes energy, and root nodules also function as strong P sinks. Low P causes a 30–40% growth and yield reduction [8]. The absorption of P by a plant is facilitated by root hairs, tips, the outermost layers of root cells, and the root-colonized mycorrhizal fungi. The absorbed P is either stored in the root or transported and stored in other cells based on the requirement.
Figure 6.
Comparison of plant growth under high and low P availability conditions. (created with https://www.biorender.com).
In low soil P situations, to achieve a higher PUE, legumes modify their root morphology, organic acid exudation, associations with mycorrhizal fungi, and signaling mechanisms [8]. P-use efficiency mainly depends on two processes P acquisition efficiency and P utilization efficiency. P acquisition efficiency mainly depends on the ability of the roots to absorb P from the soil so it can be regulated by changing the root morphological characteristics, architecture, and organic acid exudation. The major modifications plants accomplish for a better PUE are associated with root modifications and transporter deployment, which are discussed hereunder.
Root morphological modifications: Root morphological alterations related to length, volume, density, shallowness, exudates, and hair influence the PUE. For example, to achieve efficient P absorption under the P deficit situations, the primary root growth is stimulated [9]. To ensure a greater Pi acquisition, the lateral roots proliferate and expand soil exploration [10], and such modifications are based on soil Pi availability [11]. Changes in the root epidermal cell-borne hairs aid in greater absorption of nutrients, and their role in significant Pi uptake is well documented in alfalfa [5, 12]. Cluster root, which is one of the modifications of root otherwise called proteinoid roots, also helps in P acquisition. Cluster root has numerous dense short rootlets, which increase PUE through (i) increasing the surface area between soil and plant and (ii) secreting a good quantity of phosphatases, organic acids, and protons. In legumes, this type of root modification is mainly observed in white lupin [13]. Root architecture is another factor that determines PUE [14]. The legume root developmental pattern depends on soil P availability. Under P-deficiency situations, legumes develop shallow roots to cope with the mineral stress [15]. Shallow roots with a greater lateral rooting are observed in soybean under P deficiency, and such modifications increase Pi uptake more than deep root systems [16], and these observations are also well documented in a cultivated soybean core collection [16]. Soil P is highly immobile; therefore, it needs to be converted to inorganic form for proper absorption by plants. This conversion can be accelerated by the root exudates like organic acids, phosphatases, and a few others. Various organic acids are secreted from different types of legumes like pigeon pea, soybean, common bean, and so on. [17]. For example, white lupin produces cluster roots that secrete malate and citrate, which help in increasing Pi concentration.
P transporters: Four subfamilies of P transporters, PHT1, PHT2, PHT3, and PHT4, regulate Pi uptake and transport in plants. On the basis of structural differences and subcellular localization, these regulators are classified into different subfamilies [18] (Table 3).
Transporters
PHT1
PHT2
PHT3
PHT4
Location
Plasma membranes
Chloroplast membranes (plastids)
Mitochondrial membranes
Golgi apparatus
Function
Helps in P acquisition under low and high P situations
Helps in P mobility
Helps in an effective P distribution
Helps in P movement between cytosols
Table 3.
Details of P transporter subfamilies.
Gene expression studies: Organic acid synthesis in legumes is regulated by a few genes. Among them, phosphodiesterase or hydrolase enzymes, particularly purple acid phosphatases (PAPs), play key roles in hydrolyzing organic P. The resultant product of hydrolyzation is inorganic Pi, which is readily absorbed by plants. The expression studies of these genes can be targeted for increased P uptake.
Potassium (K+) makes up about 10% of a plant’s dry weight [19] and is considered a vital macronutrient. K+ has numerous roles in cell physiology and metabolism. It increases root growth, improves drought resistance, activates enzymes, maintains turgor in cells, enhances translocation of sugars and starch, and imparts resistance against several biotic stresses [20]. The plants growing in K+ limited environments are stunted and underdeveloped. K+ also influences the maturity of various crops in different ways.
Potassium absorption and mobility: Plants maintain a required level of potassium content within. To achieve this, plants use various mechanisms for sensing the availability of K+ in the soil, but these mechanisms are yet to be deciphered [21]. In K+-sufficient conditions, the external K+ is transported into the plant via plasma membrane through several channels and transporters. The K+ uptake and utilization comprise three processes: (i) root uptake, (ii) in & out movement in the xylem/phloem, and (iii) cellular compartmentalization [22]. In K+-deficit conditions, plants expand their root systems to ensure sufficient absorption through enhanced root surface contact with soil. An increase in root volume is achieved by over-expressing transcription factors that are responsible for root formation. Plants use two systems of K+ uptake depending on the K+ availability [23]. When the available soil K+ is greater than 100 mM, plants utilize a low-affinity K+ uptake system where the inward rectifying channels are shaker-type K+. High affinity K+ uptake is used when the available soil K+ is less than 100 mM [11]. Plant Cyclic Nucleotide Gated Channels (CNGC) are the type of non-selective channels that are also employed by plants in certain situations; however, the reasons for such utilization are not well known [24]. Generally, in plants, for achieving an effective K+ transport, K+ uptake permeases (KT/HAK/KUP), the Trk/HKT transporter family, and the CPA (cation proton antiporters) are used [25]. After absorption, K+ is transported to the needy tissues through loading/unloading channels of the xylem and phloem. In Arabidopsis roots, the first shaker-like outward rectifying K+ channel ‘Stelar K+ Outward Rectifier’ (SKOR) is reported, which is associated with K+ transport from stellar cells to the xylem [26]. Intracellular movement of K+ is achieved through sequestration into vacuoles and guard cell movement. These movements are regulated by several K+ transporters and channels [27].
The focus has been on enhancing NtUE in legumes due to scarce nutrient sources and environmental issues in real field conditions. Nutrient deficiency stress can cause an adaptive reaction, such as increased root volume for improved access to nutrients. For increasing the nutrient use efficiency of crops, certain conditions need to be fulfilled. They include the presence of useful genetic variation, information on gene action and trait heritability, and an appropriate knowledge of physiological determinants of NtUE. Detailed information is to be generated on nutrition acquisition and utilization, for formulating breeding strategies to achieve an efficient NtUE. The nutrient acquisition involves (i) changes in the rhizosphere properties (both biological and chemical), (ii) enhanced root growth, (iii) changes in the root architecture, (iv) microbial interactions in the rhizosphere, and (v) expression and regulation of ion transporters. An effective utilization can be achieved through greater root-to-shoot nutrient translocation, compartmentalization, partitioning, and remobilization [28]. From the research findings available on public platforms, the following breeding strategies are suggested to enhance the NtUE (Figure 7).
Figure 7.
Strategies to be targeted to enhance the NtUE.
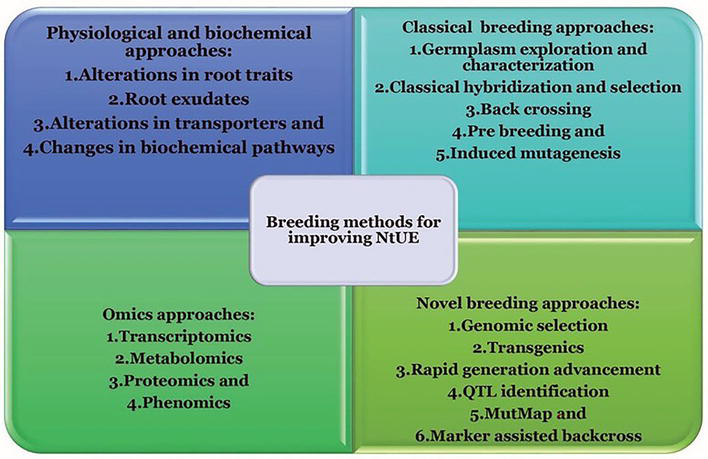
To ensure success in the NtUE programs, an integrated approach involving physiological and biochemical methods, classical breeding approaches, novel breeding methods, and omics strategies are to be formulated. Germplasm exploration, characterization, cataloging, and quantifying the existing genetic variability for NtUE are the basic plant breeding steps to be accomplished, which will further help in formulating the type of advanced research to be considered. When the required quantum of variability does not exist in the germplasm, induced mutagenesis can be employed to evolve variability. Thereon, MutMap can be considered for tagging genes or QTLs. In the situation of sufficient variability for NtUE, the genotypes can be screened under sub-optimal conditions of the nutrients. After the identification of specific genetic stocks, either MAGIC population development or classical hybridization followed by pedigree selection can be considered. When nutrient-specific donors are identified, mapping populations can be developed for the identification of QTLs and their further utilization. Different genome editing strategies can be employed to normalize various regulators (positive and negative) associated with NtUE. The CRISPR/dead (d) Cas9 (dCas9)-aided promoter engineering can be employed for cytosine/adenine base editing. To achieve the over-expression of targeted genes, this system uses transcriptional factors (repressors/activators) [29]. Different ‘omics’ strategies can be employed to understand the genetic architecture of NtUE and their consequent effects.
An adequate supply of critical nutrients in balanced ways is the key to increasing crop yield. As farmers strive to increase production and maintain profitability in the face of rising fertilizer costs and changing climate conditions, NtUE is considered important. In a production system where the food crops are nutrient-starved, improving soil fertility using fertilizers is necessarily required to sustain productivity. To achieve this requirement, synthetic fertilizers were used in enormous quantities after the Green Revolution. Indiscriminate use of synthetic fertilizers leads to environmental pollution and, therefore, warrants better management of the soil nutrient equilibrium where NtUE offers scope to achieve this target. However, progress has been slow due to the complexity of soil and plant genomes affecting crop nutrition, a lack of complete knowledge of the genetics of NtUE, and an inconsistent conceptual understanding of NtUE. With a better understanding of NtUE, scientists can make significant progress in creating more nutrient-efficient cultivars with the use of different cutting-edge technologies.
The mechanisms to increase NtUE of the plant species may vary according to nutrients and crops. The following concluding remarks shall be considered to achieve an effective NtUE.
NUE: One of the significant N sources in farming is obtained through biological nitrogen (N2) fixation. Therefore, it is suggested to identify genotypes with enhanced biological nitrogen-fixing ability that can otherwise help in effective NUE. The varieties developed in the past 50 years were bred and/or selected under high mineral fertilization inputs, which otherwise narrowed the chance of using genetic variations under low mineral fertilization conditions, thus warranting the inception of targeted selection programs. The best-performing genotypes for varied N situations shall not be selected based on the interactions between genotypes and levels of N since lines that perform best at high/ideal N levels may not be promising well at deficit/low N level(s). Evolution of new plant varieties that (i) absorb more N from the soil irrespective of form and (ii) effective utilization of absorbed N. Breeding for more active rhizobial and Arbuscular mycorrhizal (AM) symbioses in legumes can be an intriguing alternative to boosting plant yield.
PUE: Phosphorus, is highly immobile in the soil; therefore, proper P absorption is very difficult. Increasing PUE is therefore achieved by increasing root volume, root architecture, root exudates, and transporters. Significant variations are identified for different mechanisms to improve PUE in legumes. Development of transgenic soybean plants with over-expressed Gm6PGDH1 ensures greater Pi uptake under P-deficient conditions through significant lateral root growth [16]. Based on a RIL-based QTL analysis in beans under P-deficit situations, it is concluded that the QTL for basal root growth angle is co-segregated with grain yield and whose further introgression would facilitate more PUE [19]. Genome-wide association studies can be helpful in identifying important genes for improving PUE. Other methods, such as alternate gene splicing, overexpression of certain genes, and utilizing long noncoding RNA, have also been used to enhance PUE nutrient response in crops.
KUE: Development of mapping population and QTL identification, genome broadening, hereditary and evolutionary analysis, and gene mapping and identification are the commonly used techniques to improve KUE breeding. The genes for low-K tolerance and good shoot dry weight are identified via QTL analyses and GWAS, which are considerably facilitated by large amounts of genome-wide re-sequencing data, and high-quality reference genomes can further be exploited. Transcriptomic information offers insight into gene expression patterns in response to various K treatments; as a result, it has been widely used to research the genes, biological processes, and metabolic processes that determine low K tolerance. Using transcriptome profiling and QTL mapping together, it is possible to find potential genes that regulate low K tolerance. Proteomics, metabolomics, and ionomics investigations on K consumption efficiency in crops have been conducted, but they are significantly less prevalent than transcriptome research. Alternative mRNA splicing (AS) and noncoding RNAs are the main topics of epigenomics studies. The nutshell of the above-described methodologies is presented in Figure 8 for better understanding of the readers.
Figure 8.
Different breeding methodologies for improving NtUE in legumes.
References
1.Andrews M, Lea PJ, Raven JA, Azevedo RAD. Nitrogen use efficiency. 3. Nitrogen fixation: Genes and costs. The Annals of Applied Biology. 2009;155(1):1-13
2.Tegeder M, Masclaux-Daubresse C. Source and sink mechanisms of nitrogen transport and use. The New Phytologist. 2018;217(1):35-53. DOI: 10.1111/nph.14876
3.Zhang B, Meng S, Gong M. The expected and unexpected roles of nitrate transporters in plant abiotic stress resistance and their regulation. International Journal of Molecular Sciences. 2018;19(11):9-11. DOI: 10.3390/ijms19113535
4.Krouk G, Crawford NM, Coruzzi GM, Tsay YF. Nitrate signaling: Adaptation to fluctuating environments. Current Opinion in Plant Biology. 2010;13(3):265-272
5.Wang YY, Cheng YH, Chen KE, Tsay YF. Nitrate transport, signaling, and use efficiency. Annual Review of Plant Biology. 2018;69:85-122
6.Hao QN, Zhou XA, Sha AH, Wang C, Zhou R, Chen SL. Identification of genes associated with nitrogen-use efficiency by genome-wide transcriptional analysis of two soybean genotypes. BMC Genomics. 2011;12:1-15
7.Smit AL, Bindraban PS, Schröder JJ, Conijn JG, Van der Meer HG. Phosphorus in agriculture: Global resoources, trends and developments: Report to the steering committee technology assessment of the Ministery of agriculture, nature and food quality. In: The Netherlands, and in Collaboration with the Nutrient Flow Task Group (NFTG), Supported by DPRN (Development Policy Review Network). Wageningen: Plant Research International; 2009
8.Chen Z, Wang L, Cardoso JA, Zhu S, Liu G, Rao IM, et al. Improving phosphorus acquisition efficiency through modification of root growth responses to phosphate starvation in legumes. Frontiers in Plant Science. 2023;14:1094157. DOI: 10.3389/fpls.2023.1094157
9.Zhang H, Yang Y, Sun C, Liu X, Lv L, Hu Z, et al. Up-regulating GmETO1 improves phosphorus uptake and use efficiency by promoting root growth in soybean. Plant, Cell & Environment. 2020;43(9):2080-2094. DOI: 10.1111/pce.13816
11.Zhou J, Xie J, Liao H, Wang X. Overexpression of b-expansin gene GmEXPB2 improves phosphorus efficiency in soybean. Physiologia Plantarum. 2014;150(2):194-204. DOI: 10.1111/ppl.12077
12.Li C, Li K, Liu X, et al. Transcription factor GmWRKY46 enhanced phosphate starvation tolerance and root development in transgenic plants. Frontiers in Plant Science. 2021;12:700651. DOI: 10.3389/fpls.2021.700651
13.Lynch JP, Brown KM. Topsoil foraging - An architectural adaptation of plants to low phosphorus availability. Plant and Soil. 2001;237(2):225-237. DOI: 10.1023/A:1013324727040
14.Jha UC, Nayyar H, Parida SK, et al. Breeding and genomics approaches for improving phosphorus-use efficiency in grain legumes. Environmental and Experimental Botany. 2023;205:105120
15.Pang J, Ryan MH, Lambers H, Siddique KH. Phosphorus acquisition and utilisation in crop legumes under global change. Current Opinion in Plant Biology. 2018;45:248-254
16.Liang Q , Cheng X, Mei M, Yan X, Liao H. QTL analysis of root traits as related to phosphorus efficiency in soybean. Annals of Botany. 2010;106(1):223-234
17.Ramaekers L, Remans R, Rao IM, Blair MW, Vanderleyden J. Strategies for improving phosphorus acquisition efficiency of crop plants. Field Crops Research. 2010;117(2-3):169-176
18.Adams E, Shin R. Transport, signaling, and homeostasis of potassium and sodium in plants. Journal of Integrative Plant Biology. 2014;56(3):231-249. DOI: 10.1111/jipb.12159
19.Shin R. Strategies for improving potassium use efficiency in plants. Molecules and Cells. 2014;37(8):575
20.Gierth M, Mäser P. Potassium transporters in plants–involvement in K+ acquisition, redistribution and homeostasis. FEBS Letters. 2007;581(12):2348-2356
21.Liu K, Li L, Luan S. Intracellular K+ sensing of SKOR, a shaker-type K+ channel from Arabidopsis. The Plant Journal. 2006;46(2):260-268
22.Wang W, Zou J, White PJ, et al. Identification of QTLs associated with potassium use efficiency and underlying candidate genes by whole-genome resequencing of two parental lines in Brassica napus. Genomics. 2021;113(2):755-768
23.He B, Hu F, Du H, et al. Omics driven crop potassium use efficiency breeding. Frontiers in Plant Science. 2022;13:1076193. DOI: 10.3389/fpls.2022.1076193
24.Chen Y, Zhang S, Du S, Jiang J, Wang G. Transcriptome and metabonomic analysis of Tamarixramosissima potassium (K+) channels and transporters in response to NaCl stress. Genes. 2022;13(8):1313
25.Fageria NK, Barbosa Filho MP, Da Costa JGC. Potassium-use efficiency in common bean genotypes. Journal of Plant Nutrition. 2001;24(12):1937-1945
26.Li X, Zeng R, Liao H. Improving crop nutrient efficiency through root architecture modifications. Journal of Integrative Plant Biology. 2016;58(3):193-202. DOI: 10.1111/jipb.12434
27.Le Thanh T, Hufnagel B, Soriano A, Divol F, Brottier L, Casset C, et al. Dynamic development of white lupin rootlets along a cluster root. Frontiers in Plant Science. 2021;12:738172
28.McDonald G, Bovill W, Huang C, Lightfoot D. Nutrient use efficiency. In: Genomics and Breeding for Climate-Resilient Crops. Vol. 2. Target Traits. Germany: Springer; 2013. pp. 333-393
29.Gajardo HA, GÃ3mez-Espinoza O, Boscariol Ferreira P, Carrer H, Bravo LA. The potential of CRISPR/Cas technology to enhance crop performance on adverse soil conditions. Plants. 2023;12(9):1892
 Open access peer-reviewed chapter
Open access peer-reviewed chapter