
Abstract
The amygdala is a core structure of the limbic system in the brain. Anatomically, the amygdaloid complex comprises ~13 nuclei in the mid-temporal lobe. The amygdaloid complex is important for regulating emotional behavior, anxiety, fear, learning and memory. It is involved in several neurological disorders such as post-traumatic stress syndrome, depression, and temporal lobe epilepsy. The lateral nucleus of the amygdala is the main sensory input station of the amygdala and receives sensory information from cortical and subcortical (thalamic) fields. Thalamic afferents project to the lateral amygdala medially from the internal capsule, whereas cortical afferents arrive from the internal capsule. These two input pathways converge on populations of principal neurons and interneurons, both of which can be identified by characteristic electrophysiological, neurochemical, and morphological properties. Pressing issues for our understanding of the organization and operation of the amygdala are the functional significance of modulatory inputs from various signaling systems and the plasticity of its synaptic circuitry in relation to its pathway-specific inputs. This chapter reviews progress in this regard as far as cellular processes and synaptic interactions in nuclei of the amygdala are concerned which will help with our understanding of neural mechanisms underlying fear, anxiety, and related clinical disorders.
Keywords
- amygdala
- cortex
- emotion
- fear
- intercalated cells
- memory
- synaptic plasticity
- thalamus
1. Introduction
The amygdala, also known as amygdaloid complex or amygdaloid body, consists of several related nuclei. The amygdala is involved in different functional contexts such as memory, learning, emotion, fear, and motivation. Moreover, neurons in the amygdala contribute to symptoms of temporal lobe epilepsy and spread of seizure discharges in models of epilepsy. The amygdala is a core structure of the limbic system and a key site for emotion, anxiety, fear, learning and memory [1, 2]. Anatomically, the amygdaloid complex comprises ~13 nuclei in the midtemporal lobe. The amygdaloid complex is important for regulating emotional behavior and learning and is involved in several neurological disorders such as post-traumatic stress syndrome, depression, and temporal lobe epilepsy [3, 4, 5]. Pressing issues for our understanding of the organization and operation of the amygdala are the functional significance of modulatory inputs from various signaling systems and the plasticity of its synaptic circuitry in relation to its pathway-specific inputs. Recent studies set out to determine (a) the role of signaling molecules and their receptors for nerve cell signaling and information processing and (b) the influence of these signaling systems on learning and memory processes in the amygdala. The long-term objective of these studies is to understand the neural mechanisms underlying fear, anxiety, and related clinical disorders. The amygdala is critically involved in mediating normal and pathological conditions of fear and anxiety. Furthermore, the amygdala serves as a relay station for sensory input and output to higher order brain centers and centers of autonomic control. The lateral amygdaloid nucleus (lateral amygdala, LA) is the initial site of synaptic integration for cortical and thalamic afferent input to the amygdala. Signal flow of lateral amygdala excitatory principal neurons to downstream basolateral and central amygdaloid nuclei is influenced by two populations of GABAergic interneurons, classic local interneurons scattered throughout the amygdala and clusters of paracapsular intercalated (ITC) cells. These interneurons control impulse traffic through the amygdala, are sites of synaptic plasticity, and control fear memory. Using genetically engineered mice in which specific neuronal populations are labeled with fluorescence markers, it is possible to classify interneurons by describing their molecular, cellular and electrophysiological properties. Patch-clamp electrophysiology can be used to determine activity patterns and plastic behavior of principal neurons and interneurons in acute brain slices. Results obtained in these studies aid in better understanding the mechanisms of fear and anxiety, and consequently anxiety disorders, and might yield a new approach in fighting anxiety and stress disorders.
2. Fear, stress, and anxiety disorders
Fear and stress responses are facets of a behavioral repertoire that contributes to the survival of an individual. However, pathological deviations from normal fear and stress responses manifest themselves as clinical disorders including mood, fear, and anxiety disorders such as panic disorder and post-traumatic stress disorder (PTSD). PTSD has a lifetime prevalence of 6.8% of the adult population in the United States [6]. The amygdala is an anatomical and functional brain substrate critically involved in mediating normal and pathological conditions. Despite the clinical and functional importance of the amygdala, it is only recently that general principles of intra-amygdaloid mechanisms of signal processing have emerged from electrophysiological, anatomical, and pharmacological studies.
The amygdaloid complex is important for regulating emotional behavior and learning [7, 8, 9, 10, 11, 12, 13]. It is involved in several neurological disorders. Some of these disorders may also lie at the root of the many well-documented race-based physical health disparities that affect African Americans and other minority populations [14]. For example, after hurricane Katrina, emotional and stress-related health problems affected primarily members of minority populations, and stress through racism affects primarily black men.
3. Structural and functional organization of the lateral amygdala
The amygdala is a central relay station for sensory input and output to higher order brain centers and centers of autonomic control. The lateral nucleus of the amygdala is the initial site of synaptic integration for cortical and thalamic afferent input to the amygdala. Synaptic information is then relayed to the basolateral nucleus with the central nucleus serving as the amygdala output station. In rat brain slices, sharp-electrode recordings in an interface chamber can be used to determine physiological and synaptic parameters of lateral amygdala neurons [15, 16]. Neurons that are labeled intracellularly and digitally reconstructed allow a correlation between structure and function (Figures 1 and 2). Principal neurons can be identified by morphological (after Biocytin injection and histological processing) and electrophysiological criteria. Projection neurons reveal complex morphologies with numerous spiny dendrites (Figures 1 and 2). Principal neurons in the amygdala generate slow membrane potential oscillations [1, 17]. Specifically, neurons produce theta activity (6–10 Hz) (Figure 1) during emotional arousal and various types of rhythmic activity during sleep.
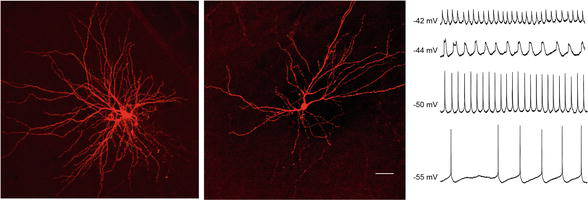
Figure 1.
Left and middle panels: confocal images of two principal neurons with many spine-rich dendrites in the lateral amygdala recorded intracellularly, stained with biocytin and incubated with Cy3-streptavidin. The neuron on the left has a pyramidal appearance, whereas the one in the center is more stellate in shape. Scale bar, for left image: 100 μm, center image: 150 μm. Right panel: intracellular recordings from a principal neuron in the lateral amygdala. Two types of oscillations of the membrane potential (low threshold [e.g., at −50 mV] and high threshold oscillations [at −42 mV and −44 mV]) are generated in principal neurons of the lateral amygdala.
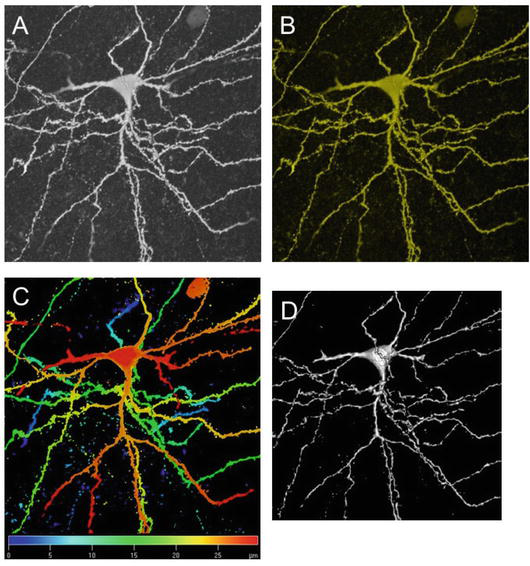
Figure 2.
Four images of the same principal neuron in the lateral amygdala, stained with biocytin and incubated with Cy3-streptavidin. (A) Laser scanning confocal image of the neuron. (B) False color image of the neuron. (C) Color-coded spatial image of the neuron. (D) Surface-rendered image of the neuron to generate 3-D effect.
An acute brain slice preparation that contains the amygdala and adjacent regions of the brain, e.g., cortical and thalamic areas (Figure 3) is an advantageous experimental system for studies of amygdala signaling [15, 16]. This slice preparation preserves the synaptic circuitry in the amygdala to the extent that one can study synaptic processing by electrically stimulating afferent input (stimulation electrodes) while recording from neurons (recording electrode) in the lateral amygdala or other amygdaloid nuclei. It is possible to record and intracellularly stain principal neurons and interneurons in the amygdala while selectively activating thalamic or cortical input fibers (Figure 3). Thereby, one can synaptically drive postsynaptic neurons and induce neuroplasticity in an input-specific manner. Such frontal amygdaloid slices, prepared from deeply anesthetized rats of either sex (postnatal days 25–30) [16], allow to evoke synaptic responses from principal neurons in the lateral amygdala with two bipolar tungsten electrodes placed in the external capsule and close to the internal capsule for stimulation of putative cortical and thalamic afferents, respectively [18, 19]. Stimulus intensity can be adjusted to produce a synaptic response 30–50% of maximum amplitude without triggering action potentials. In such experiments, neurons are held at resting membrane potential.
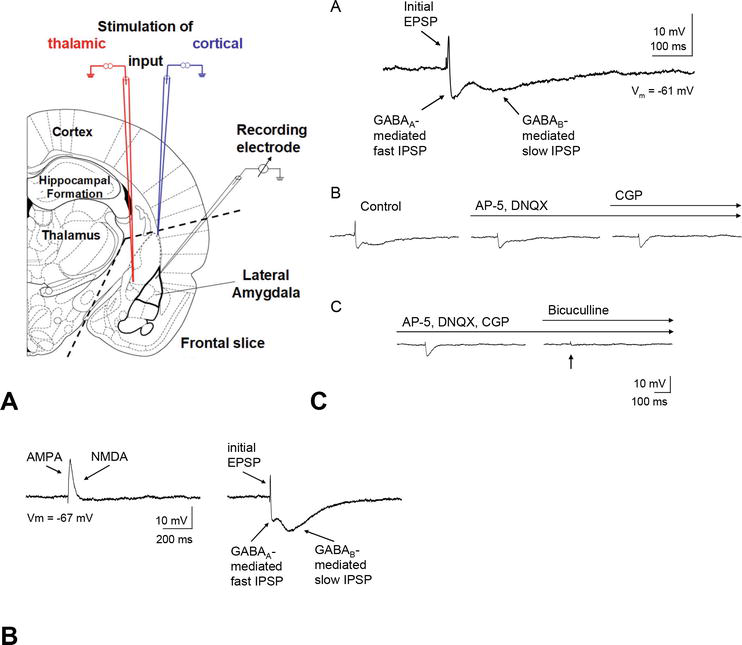
Figure 3.
(A) Amygdala slice preparation and activation of afferent input fibers to the lateral amygdala
Recordings from principal neurons in the lateral amygdala have confirmed that electrical stimulation of afferents that project to the lateral amygdala evokes excitatory and inhibitory postsynaptic potentials (PSPs) and thus results in a dual effect with direct excitation of principal neurons coupled with concurrent feedforward inhibition via interneurons [17, 20, 21, 22]. The excitatory PSPs (EPSPs) are mediated by feedforward glutamate receptor-mediated excitation. The inhibitory PSPs (IPSPs) are either polysynaptic in origin or they are monosynaptically mediated by feedforward inhibition via local GABAergic interneurons since (a) the IPSPs can be evoked after blocking excitatory transmission (Figure 3) [15], (b) connections between different basolateral amygdala nuclei primarily consist of excitatory connections [23], and (c) lesions deafferenting the basolateral complex lead to minor decreases in glutamic acid decarboxylase levels [24]. Thus, the excitability of principal neurons in the lateral amygdala depends upon the relative strength of the inputs to projection neurons and interneurons.
4. Synaptic plasticity and fear learning in the lateral amygdala
The lateral amygdala is the main sensory input station of the amygdala and receives sensory information from cortical and subcortical (thalamic) fields [25]. Thalamic afferents project to the lateral amygdala medially from the internal capsule, whereas cortical afferents arrive from the external capsule [26]. These two input pathways converge on principal neurons and interneurons, both of which have characteristic electrophysiological, neurochemical, and morphological properties [20, 21, 27, 28, 29, 30]. Theta rhythm activity (Figure 1) has been implicated in lateral amygdala synaptic plasticity, retrieval of fear memory, and transmission in the amygdalo-hippocampal network [31]. Therefore, theta frequency stimulation can be used as a paradigm to test for intercellular signaling and synaptic plasticity in the lateral amygdala. We have reported that homosynaptic long-term depression (LTD) of excitatory responses can be induced in the lateral amygdala after stimulation of putative thalamic inputs but not of cortical inputs (Figure 4) [16]. This form of LTD is induced by theta frequency stimulation and involves postsynaptic Ca2+-dependent mechanisms and metabotropic glutamate receptors. The reader is referred to [2] for an excellent review of plastic synaptic networks of the amygdala for the acquisition, expression, and extinction of conditioned fear.
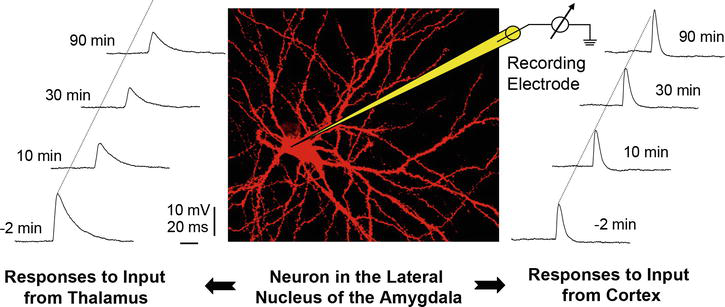
Figure 4.
Theta frequency stimulation of thalamic afferents evokes an LTD of EPSP amplitude evoked with single thalamic control stimuli while it has no effect on cortically evoked EPSPs. Theta stimulation of cortical input fibers does not result in a change of EPSP amplitude of either afferent input pathway demonstrating input specific plasticity, namely, depression of EPSPs in response to stimulation of thalamic afferents. Modified from [
5. Principles of signal processing in the amygdala
Three principles of signal processing have emerged from studies of amygdala circuitry [1, 2]: (1) oscillatory activity in the theta frequency range, (2) dominance of inhibition and (3) sustained synaptic plasticity. One principle of information processing in the amygdala is the stereotypic responses in the form of slow oscillating activity in the theta frequency range [1, 32]. Studies of the membrane properties of projection neurons in the lateral amygdala revealed that these neurons are capable of generating intrinsic stable oscillations in the frequency range of 6–10 Hz (Figure 1) which is reminiscent of theta waves in the limbic system. The frequency of the oscillations is relatively constant even with excitatory synaptic input of different strength. The functional consequence is a synchronization of intrinsic and synaptic input in this defined frequency and a temporal structuring of synaptic input activity toward slow rhythmic output [33, 34]. These oscillations form the basis of phase-coupled oscillatory response patterns that are evoked in neuronal networks of the amygdala. In amygdalohippocampal circuits, rhythmic synchronization at theta frequencies increases between the hippocampal CA1 region and the lateral amygdala after fear conditioning and presentation of fear stimuli [31]. Therefore, synchronized theta activities in the amygdalo-hippocampal network serve as a neuronal correlate of conditioned fear.
A second principle of information processing is the dominance of synaptic inhibition in the amygdala (Figure 3). Mainly feedforward inhibitory responses are observed to stimulation of major afferent systems through activation of GABAergic or glycinergic receptors [21, 22, 35, 36, 37, 38]. Powerful IPSPs regulate the responses of projection neurons in the lateral amygdala relatively irrespective of the stimulation site, i.e., perirhinal, entorhinal, basomedial or lateral amygdala stimulation [35]. Thus, the lateral amygdala seems to be equipped with an inhibitory gating mechanism regulating information flow through the amygdala [27, 39]. We found that adenosine may participate in these neuronal processes related to fear conditioning, learning and memory in the amygdala [15], e.g., by directly interacting with the inhibitory mechanisms and/or by modulating the potentiation of GABAA-mediated synaptic currents in pyramidal neurons after tetanic stimulation of inputs to interneurons [28]. A decrease of GABAergic interneuron activity in the lateral amygdala is correlated with the development of epileptiform discharges. In the kindling model of epilepsy, GABA receptor-mediated inhibitory transmission is reduced [40, 41, 42] and glutamatergic transmission is enhanced in the amygdala [41, 42].
Another principle of processing in the lateral amygdala is the prevalence of synaptic plasticity, including input-specific neural plasticity of its afferent pathways (Figure 4). Different forms of activity-dependent synaptic plasticity have been reported to occur in the amygdala, one of which, long-term potentiation, has been suggested to be an important cellular mechanism for conditioned fear [9]. During fear conditioning the synapses of thalamic afferent pathways in the lateral amygdala undergo synaptic plasticity [43, 44] illustrating the relevance of associative LTP for learning processes in a behavioral context. The molecular mechanisms of LTP have been studied as well as other forms of plasticity such as LTD and bidirectional synaptic plasticity (depression and potentiation) that is sensitive to the type and sequence of the stimulation paradigm used [16, 45, 46, 47]. Modulation of synaptic transmission in the amygdala by activity-dependent processes is controversially discussed in the literature, and it is not yet clear how the different input systems relate to synaptic plasticity in the amygdala and to conditioned fear. Short-term and long-term synaptic plasticity in the amygdala appear to play a critical role in conditioned fear and anxiety.
6. Synaptic signaling mechanisms and plasticity in amygdala interneurons
Fear learning involves the lateral and basolateral nuclei (BLA) of the amygdala. Here the association between incoming fearful and neutral stimuli leads to potentiation of synaptic transmission. Projections from the basolateral amygdala nucleus synapse on neurons in the central nucleus which in turn projects its axons to the hypothalamus and brainstem to trigger the autonomic expression of fear [9]. However, this classic view was recently challenged [48] in favor of a model in which afferent input reaches both the lateral and central amygdala. Lateral amygdala neurons do not directly synapse on central amygdala output neurons, but rather lateral amygdala neurons are thought to synapse on interneurons which in turn disinhibit brainstem projecting central amygdala neurons. The central amygdala itself can be a site of synaptic plasticity. The GABAergic interneurons of the paracapsular intercalated (ITC) cell masses that surround the lateral amygdala and basolateral amygdala have a pivotal role in mediating signal flow from lateral to central amygdala [39, 49, 50, 51, 52, 53]. Therefore, pressing issues for understanding the organization and operation of the amygdala are the functional significance of modulatory input (e.g., [54, 55]), the functional role of its distinct neuronal populations, and the plasticity of its synaptic circuitry in relation to its pathway-specific inputs.
One of the emerging principles of synaptic processing in the amygdala is the dominance of inhibition. In addition to excitatory principal neurons, the amygdala houses two populations of GABAergic interneurons: (1) classic local interneurons scattered throughout the amygdala and (2) clusters of paracapsular intercalated (ITC) cells (Figure 5). Afferent synaptic input converges onto single principal neurons as well as interneurons in the lateral amygdala. The relay from lateral amygdala principal neurons to downstream amygdala nuclei is regulated by GABAergic interneurons through feedforward and feedback inhibition. Classic interneurons in the lateral amygdala respond with EPSCs of constant latencies to minimal stimulation of both cortical and thalamic fibers, indicating direct convergent monosynaptic input [29]. Based on the data, a population of interneurons in the lateral amygdala is likely to receive convergent input from thalamic and cortical fibers through principal neurons both directly and indirectly (in a feedforward and feedback manner, respectively). Irrespective of the route of excitation, interneurons in the lateral amygdala can be regarded as homogeneous with respect to their afferent connections considering thalamic and cortical input pathways and mediate inhibitory control of postsynaptic principal neurons. This symmetrically built GABAergic circuitry can be of functional significance, given the distinctive role of the two afferent input systems for the mediation of different components of fear responses and the importance of GABAergic mechanisms for limitation of excessive neuronal activity [1, 29].
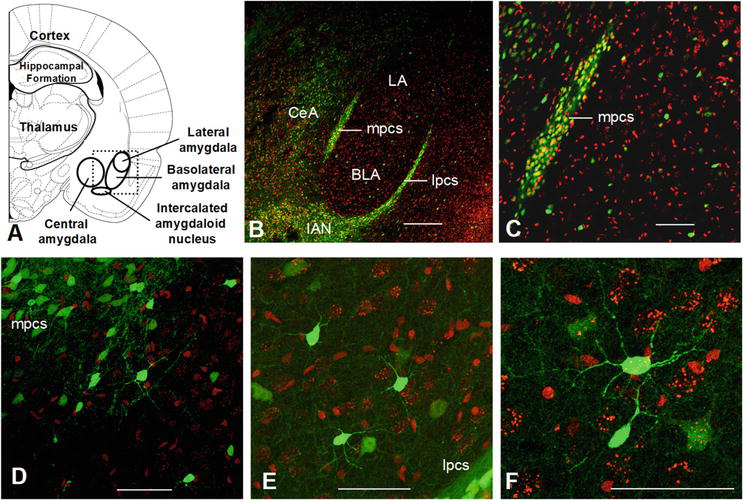
Figure 5.
Confocal images of amygdala slices from GAD-67-GFP mice counterstained with a nuclear stain (Sytox orange). (A) Schematic diagram of parts of the amygdala. Dotted rectangle indicates outlines of panel B. (B) Paracapsular intercalated cells form an inhibitory sheath surrounding the basolateral amygdala (BLA). Clusters of intercalated cells are located primarily along the external capsule (lateral paracapsular cells, lpcs) and along the border of BLA and central amygdala (CeA) (medial pcs, mpcs). Scattered GABAergic interneurons are found throughout the BLA and adjacent areas. The intercalated nucleus is located immediately ventro-medial to the BLA. (C) The medial cluster of paracapsular cells contains many GABAergic INs; same slice as in B. (D) Higher magnification of parts of the medial paracapsular cell cluster and scattered INs in the BLA; same slice as in B. (E) Scatterd INs and parts of the lateral paracapsular cell cluster, same slice as in B. (F) Dendritic branching pattern of an IN in BLA; same slice as in B. Scale bars: B—200 μm, C—100 μm, D—50 μm, E—50 μm, F—50 μm. BLA—basolateral amygdala; CeA—central amygdala; IAN—intercalated amygdaloid nucleus; LA—lateral amygdala; lpcs—lateral paracapsular cells; mpcs—medial paracapsular cells.
7. Intercalated cells of the amygdala
Another principle of processing in the lateral amygdala is the prevalence of synaptic plasticity, including input-specific neural plasticity of its afferent pathways. Based on the findings discussed above, one hypothesis is that Pavlovian fear conditioning and fear memory involve synaptic plasticity also of the inhibitory GABAergic interneurons. Specifically, (a) activation of interneurons in the lateral amygdala might regulate impulse traffic of its principal neurons, and (b) interneurons in the amygdala are sites of input-specific and location-specific neural plasticity themselves. These interneurons include scattered interneurons in the amygdala and the paracapsular ITC neurons [56]. Genetically engineered mice, GAD-67-GFP mice [57, 58], house specific neuronal populations, namely GABAergic interneurons, that are labeled with fluorescence markers to identify and classify GABAergic neurons in slice preparations
In the GAD-67-GFP mouse line, GABAergic interneurons are fluorescently labeled with GFP such that GABAergic cells in slices from these mice can be directly visualized with fluorescence microscopy in the patch-clamp slice microscope. In this transgenic mouse line, the promoter for GAD-67 drives the expression of GFP. In addition to their fluorescent property, amygdala interneurons can be recognized by electrophysiological criteria. Classic interneurons respond with regular sustained firing to depolarizing current pulses, they possess high resting input resistance and generate fast spikes compared with those of principal cells [17, 27, 28, 29, 60, 61]. Interneurons typically have non-spiny or sparsely spiny dendrites (Figure 5).
Few studies have addressed synaptic signaling mechanisms and plasticity in amygdala interneurons. After tetanic electrical stimulation of the external capsule, The authors in [28] observed long-term potentiation (LTP) mediated by calcium-permeable AMPA receptors in putative GABAergic interneurons in the basolateral amygdala. This LTP is reflected in an increased GABAA-mediated inhibitory synaptic current in principal neurons. In another study, tetanization of thalamic afferents resulted in NMDA-dependent heterosynaptic LTP of excitatory postsynaptic currents (EPSCs) in interneurons which in turn was reflected as larger IPSCs in postsynaptic principal neurons [62]. However, the potentiation was found in both input pathways, thalamic and cortical. Interneurons of ITC cell masses located at the border between central amygdala and basolateral amygdala (putative medial paracapsular cells, mpcs) displayed bidirectional synaptic plasticity (LTD, LTP) depending on the stimulus paradigm used [63, 64]. More recent evidence points to a role of ITC cell masses in fear extinction [65, 66] and a specific role of neuropeptide S in interneuron-related fear processes [65, 67]. ITC cells form a sheath of GABAergic interneurons surrounding the basolateral amygdala [2, 68]. Additionally, a prominent intercalated nucleus of GABAergic cells is located ventro-medially of the basolateral (Figure 5). Intercalated cells of the medial group, which is located at the border of basolateral amygdala and central amygdala, display distinct short-term plastic synaptic responses that vary dramatically between different connected cell pairs [69]. Intercalated cells exhibit an unusual pattern of receptor expression and do not stain for typical interneuron markers such as calbindin, calretinin, parvalbumin, NPY, CCK, somatostatin [59]. Furthermore, only study has attempted to systematically analyze the large number of anatomically distinct ITC interneurons in terms of electrophysiological parameters [59]. Overall, these studies reveal an exciting, yet inconclusive picture of interneuron function in the amygdala and do not provide a clear functional assignment and/or classification of GABAergic cells. The data available suggest that, as in other neural systems [70], interneurons in the amygdala present themselves as diverse types of neurons [30, 71] and possibly participate in spatiotemporal division of labor. This idea is supported by two studies of amygdala neural circuits [54, 72]. Interneurons in the amygdala are found in specific locations and patterns of aggregation, either in ITC clusters or distributed as individual interneurons throughout amygdaloid nuclei. These interneurons can also be distinguished based on morphological features. In confocal images, interneurons of ITC cell masses have smaller soma size, and their dendritic branching is less readily visible than the more extensive branching of scattered interneurons. This suggests that differences in dendritic receptive fields exist that in turn could result in fewer synaptic connections of ITC cells and a possible dominance of short-range synaptic interactions with neighboring ITC cells. Thus, one possible role of ITC cells could be to mediate lateral inhibition in ITC cell clusters whereas classic interneurons with longer dendrites could function in longer range feedforward or feedback inhibition of principal neurons. Furthermore, afferent input from thalamic and cortical regions can either converge on individual interneurons and principal neurons or reach amygdaloid neurons in a more distributed manner [29]. All of this suggests that subpopulations of interneurons exhibit distinct functional roles in amygdaloid networks. The basis of these functional roles rests with distinct intrinsic and synaptic properties of interneurons which are supported by two studies that showed how distinct neuronal circuits can differentially affect amygdala function [54, 72].
8. Fear conditioning, functional imaging, and theta rhythms
Functional imaging studies, such as fMRI (functional magnetic resonance imaging) studies, have confirmed the central role of the amygdala in fear conditioning paradigms. Fear conditioning is a classical experimental paradigm used to investigate associative learning, particularly the formation and expression of fear-related memories. In this paradigm, participants are exposed to neutral stimuli (such as tones or images) paired with aversive stimuli (such as electric shocks or unpleasant images). Over time, the neutral stimuli become associated with fear responses [73, 74, 75]. The amygdala is critically involved in the acquisition and consolidation of fear-related memories during fear conditioning. Studies have consistently shown that activity in the amygdala increases when animal models (rodents) or humans are exposed to conditioned stimuli associated with fear-inducing events. This increased activity reflects the encoding and storage of fear memories [4, 5, 9, 76, 77]. In fear conditioning paradigms, the amygdala also plays a role in the expression of fear responses. Once fear memories are formed, the amygdala becomes activated when individuals encounter the conditioned stimuli, even in the absence of the aversive stimuli. This activation leads to the expression of fear responses, such as increased arousal, physiological changes, and behavioral reactions. Functional neuroimaging techniques such as fMRI examine the neural correlates of fear conditioning in humans. Studies using fMRI have consistently implicated the amygdala in the processing of fear-related stimuli during fear conditioning tasks. Increased amygdala activity is observed when participants are exposed to conditioned stimuli associated with fear, providing evidence for its involvement in fear learning and expression [77, 78, 79, 80]. Beyond its role as a primary hub for fear processing, the amygdala interacts with other brain regions to modulate fear responses. fMRI studies have revealed functional connectivity between the amygdala and various cortical and subcortical regions involved in emotional regulation, attention, and memory. These network interactions contribute to the complex processing of fear-related information and the regulation of emotional responses [77, 78, 79, 80].
Fear conditioning represents a most reproducible paradigm to study the neurobiological mechanisms underlying emotions [81]. Studies of the synaptic plasticity underlying fear conditioning in both humans and rodent models focus on neural circuits in the amygdala and prefrontal cortex, cingulate gyrus, and hippocampus [82, 83]. Plastic changes involve cortical representations and the refinement of contextual input as part of circuit modulation. Theta oscillatory activity across several brain regions has been found to be critical in this regard [31, 84, 85, 86, 87], e.g., in relation to long-term fear memory, modulation of synchronized oscillations in the hippocampal-prefrontal-amygdala circuit in contextual fear, and changes in brain rhythms and connectivity tracking fear acquisition and reversal. Projection neurons of the lateral amygdala exhibit resonant/oscillatory behavior of their membrane potential and provide a cellular correlate of coherent theta activity in amygdalo-hippocampal pathways (see Figure 1). This can be a neural correlate of conditioned fear [84, 85]. Synaptic plasticity observed in these networks has been proposed to rely on correlated activity, e.g., input-specific long-term depression of thalamo-amygdaloid signals and consolidation of long-term potentiation in the dentate gyrus. Moreover, increased theta rhythm in the cingulate cortex was found to subserve fear acquisition and is transmitted to other cortical regions via increased functional connectivity, which allows for fast theta rhythm synchronization [87].
9. Conclusions
A long-term objective of research in the amygdala is to understand the neural mechanisms underlying fear, anxiety, and related clinical disorders. The amygdala is critically involved in mediating normal and pathological conditions of fear and anxiety. The amygdala serves as a relay station for sensory input and output to higher order brain centers and centers of autonomic control. The lateral amygdaloid nucleus is the initial site of synaptic integration for cortical and thalamic afferent input to the amygdala. Signal flow of lateral amygdala excitatory principal neurons to downstream basolateral and central amygdaloid nuclei is influenced by two populations of GABAergic interneurons, classic local interneurons scattered throughout the amygdala and clusters of paracapsular intercalated cells. Interneurons control impulse traffic through the amygdala, are sites of synaptic plasticity, and control fear memory.
A basic principle of information processing in the amygdala is the stereotypic responses in the form of oscillating activity in the theta frequency range. Input specific plastic changes seen with the theta frequency stimulation paradigm make them extremely relevant for synaptic plasticity and help to integrate the intrinsic properties of the projection neurons into synaptic and network properties in the amygdala. Presumably, synaptic depression in the lateral amygdala, like other forms of enduring synaptic weakening, provides a means of reducing the relative contribution of a high synaptic input. Changes in synaptic strength are input specific and could mediate balance shifts of thalamic vs. cortical influences, possibly with effects on fear conditioning neuronal circuits. The functional consequence could be a reduction of reflex-like components of conditioned fear responses, the direct thalamic influences, with a shift toward a stronger cortical component which is linked to analysis of contextual stimuli. These studies determine the functional significance of neurotransmitter systems and provide insights into amygdala network properties. Furthermore, the results could help to explain and suggest treatments for pain, anxiety, phobias, stress, and neurological conditions associated with amygdala networks.
Acknowledgments
This work was supported in part by the National Science Foundation [NSF IOS-1355034], Howard University College of Medicine, and the District of Columbia Center for AIDS Research, an NIH funded program [P30AI117970], which is supported by the following NIH Co-Funding and Participating Institutes and Centers: NIAID, NCI, NICHD, NHLBI, NIDA, NIMH, NIA, NIDDK, NIMHD, NIDCR, NINR, FIC and OAR. The content is solely the responsibility of the author and does not necessarily represent the official views of the NIH.
I wish to express my sincere gratitude to Prof. Dr. Hans-Christian Pape for his guidance and support during my studies of the amygdala in his institute at the Otto-von-Guericke University, Magdeburg, Germany. I am grateful to Dr. Michael T. Shipley and Dr. Adam Puche (both University of Maryland School of Medicine, Baltimore, Maryland) for sharing GAD67-GFP labeled brain material. I would like to thank Prof. Dr. Yuchio Yanagawa (Gunma University, Gunma, Japan) for allowing me to use the GAD67-GFP knock-in mice that he generated. I wish to acknowledge the Integrated Imaging Center, Johns Hopkins University (Baltimore, Maryland) for using the facility, including the Zeiss LSM 510 VIS/Confocor 3 FCS.
Conflict of interest
The author declares no conflict of interest.
References
- 1.
Pape H-C, Driesang RB, Heinbockel T, Laxmi TR, Meis S, Seidenbecher T, et al. Cellular processes in the amygdala: Gates to emotional memory? Zoology. 2001; 104 :232-240 - 2.
Pape HC, Pare D. Plastic synaptic networks of the amygdala for the acquisition, expression, and extinction of conditioned fear. Physiological Reviews. 2010; 90 (2):419-463. DOI: 10.1152/physrev.00037.2009. Erratum in: Physiological Reviews 2010;90 (3):1269 - 3.
Johansen JP, Cain CK, Ostroff LE, LeDoux JE. Molecular mechanisms of fear learning and memory. Cell. 2011; 147 :509-524 - 4.
LeDoux JE. Rethinking the emotional brain. Neuron. 2012; 73 :653-676 - 5.
LeDoux JE. Coming to terms with fear. PNAS USA. 2014; 111 :2871-2878 - 6.
NIH website: Post-Traumatic Stress Disorder (PTSD). Available from: https://www.nimh.nih.gov/health/statistics/post-traumatic-stress-disorder-ptsd [Accessed: January 02, 2024] - 7.
Davis M. Role of NMDA receptors and MAP kinase in the amygdala in extinction of fear: Clinical implications for exposure therapy. The European Journal of Neuroscience. 2002; 16 :395-398 - 8.
Herry C, Ciocchi S, Senn V, Demmou L, Muller C, Luthi A. Switching on and off fear by distinct neuronal circuits. Nature. 2008; 454 :600-606 - 9.
LeDoux JE. Emotion circuits in the brain. Annual Review of Neuroscience. 2000; 23 :155-184 - 10.
Levine J, Chengappa KN, Gershon S, Drevets W. Differentiating primary pathophysiologic from secondary adaptational processes. Depression and Anxiety. 2001; 14 :105-101 - 11.
Phelps EA, LeDoux JE. Contributions of the amygdala to emotion processing: From animal models to human behavior. Neuron. 2005; 48 (2):175-187. DOI: 10.1016/j.neuron.2005.09.025 - 12.
Ressler KJ, Mayberg HS. Targeting abnormal neural circuits in mood and anxiety disorders: From the laboratory to the clinic. Nature Neuroscience. 2007; 10 (9):1116-1124. DOI: 10.1038/nn1944 - 13.
Shin LM, Rauch SL, Pitman RK. Amygdala, medial prefrontal cortex, and hippocampal function in PTSD. Annals of the New York Academy of Sciences. 2006; 1071 :67-79 - 14.
Mays VM, Cochran SD, Barnes NW. Race, race-based discrimination, and health outcomes among African Americans. Annual Review of Psychology. 2007; 58 :201-225 - 15.
Heinbockel T, Pape H-C. Modulatory effects of adenosine on postsynaptic potentials of projection neurons in the lateral amygdala of the rat. British Journal of Pharmacology. 1999; 128 :190-196 - 16.
Heinbockel T, Pape H-C. Input-specific long-term depression in the lateral amygdala evoked by theta frequency stimulation. The Journal of Neuroscience. 2000; 20 (RC68):1-5 - 17.
Washburn MS, Moises HC. Electrophysiological and morphological properties of rat basolateral amygdaloid neurons in vitro. The Journal of Neuroscience. 1992; 12 :4066-4079 - 18.
Mahanty NK, Sah P. Excitatory synaptic inputs to pyramidal neurons of the lateral amygdala. The European Journal of Neuroscience. 1999; 11 :1217-1222 - 19.
Weisskopf MG, LeDoux JE. Distinct populations of NMDA receptors at subcortical and cortical inputs to principal cells of the lateral amygdala. Journal of Neurophysiology. 1999; 81 :930-934 - 20.
Rainnie DG, Asprodini EK, Shinnick-Gallagher P. Excitatory transmission in the basolateral amygdala. Journal of Neurophysiology. 1991; 66 :986-998 - 21.
Rainnie DG, Asprodini EK, Shinnick-Gallagher P. Inhibitory transmission in the basolateral amygdala. Journal of Neurophysiology. 1991; 66 :999-1009 - 22.
Washburn MS, Moises HC. Inhibitory responses of rat basolateral amygdaloid neurons recorded in vitro. Neuroscience. 1992; 50 :811-830 - 23.
Smith Y, Paré D. Intra-amygdaloid projections of the lateral nucleus in the cat: PHA-L anterograde labeling combined with postembedding GABA and glutamate immunocytochemistry. The Journal of Comparative Neurology. 1994; 342 (2):232-248. DOI: 10.1002/cne.903420207 - 24.
Le Gal LSG, Paxinos G, Ben-Ari Y. Neurochemical mapping of GABAergic systems in the amygdaloid complex and bed nucleus of the stria terminalis. Brain Research. 1978; 155 (2):397-403. DOI: 10.1016/0006-8993(78)91037-5 - 25.
Pitkanen A, Savander V, LeDoux JE. Organization of intra-amygdaloid circuitries in the rat: An emerging framework for understanding functions of the amygdala. Trends in Neurosciences. 1998; 20 :517-523 - 26.
LeDoux JE, Farb CR, Milner TA. Ultrastructure and synaptic associations of auditory thalamo-amygdala projections in the rat. Experimental Brain Research. 1991; 85 :577-586 - 27.
Lang EJ, Paré D. Synaptic responsiveness of interneurons of the cat lateral amygdaloid nucleus. Neuroscience. 1998; 83 (3):877-889. DOI: 10.1016/s0306-4522(97)00420-x - 28.
Mahanty NK, Sah P. Calcium-permeable AMPA receptors mediate long-term potentiation in interneurons in the amygdala. Nature. 1998; 394 :683-687 - 29.
Szinyei C, Heinbockel T, Montagne J, Pape HC. Putative cortical and thalamic inputs elicit convergent excitation in a population of GABAergic interneurons of the lateral amygdala. The Journal of Neuroscience. 2000; 20 (23):8909-8915. DOI: 10.1523/JNEUROSCI.20-23-08909.2000 - 30.
Sah P, Faber ESL, Lopez de Armentia M, Power J. The amygdaloid complex: Anatomy and physiology. Physiological Reviews. 2003; 83 :803-834 - 31.
Seidenbecher T, Laxmi TR, Stork O, Pape H-C. Amygdalar and hippocampal theta rhythm synchronization during fear memory retrieval. Science. 2003; 301 :846-850 - 32.
Paré D, Pape H-C, Dong J. Bursting and oscillating neurons of the cat basolateral amygdaloid complex in vivo: Electrophysiological properties and morphological features. Journal of Neurophysiology. 1995; 74 :1179-1191 - 33.
Collins DR, Paré D. Reciprocal changes in the firing probability of lateral and central medial amygdala neurons. The Journal of Neuroscience. 1999; 19 :836-844 - 34.
Paré D, Collins DR, Pelletier JG. Amygdala oscillations and the consolidation of emotional memories. Trends in Cognitive Sciences. 2002; 6 :306-314 - 35.
Lang EJ, Paré D. Similar inhibitory processes dominate the responses of cat lateral amygdaloid projection neurons to their various afferents. Journal of Neurophysiology. 1997; 77 (1):341-352. DOI: 10.1152/jn.1997.77.1.341 - 36.
Nose I, Higashi H, Inokuchi H, Nishi S. Synaptic responses of Guinea pig and rat central amygdala neurons in vitro. Journal of Neurophysiology. 1991; 65 :1227-1241 - 37.
Danober L, Pape H-C. Mechanisms and functional significance of a slow inhibitory potential in neurons of the lateral amygdala. The European Journal of Neuroscience. 1998; 10 :853-867 - 38.
Danober L, Pape H-C. Stychnine-sensitive glycine responses in neurons of the lateral amygdala: An electrophysiological and immunocytochemical characterization. Neuroscience. 1998; 85 :427-441 - 39.
Royer S, Martina M, Paré D. An inhibitory interface gates impulse traffic between the input and output stations of the amygdala. The Journal of Neuroscience. 1999; 19 (23):10575-10583. DOI: 10.1523/JNEUROSCI.19-23-10575.1999 - 40.
Gean PW, Shinnick-Gallagher P, Anderson AC. Spontaneous epileptiform activity and alteration of GABA- and of NMDA-mediated neurotransmission in amygdala neurons kindled in vivo. Brain Research. 1989; 494 :177-181 - 41.
Asprodini EK, Rainnie DG, Shinnick-Gallagher P. Epileptogenesis reduces the sensitivity of presynaptic gamma-aminobutyric acidB receptors on glutamatergic afferents in the amygdala. The Journal of Pharmacology and Experimental Therapeutics. 1992; 262 :1011-1021 - 42.
Rainnie DG, Asprodini EK, Shinnick-Gallagher P. Kindling-induced long-lasting changes in synaptic transmission in the basolateral amygdala. Journal of Neurophysiology. 1992; 67 :443-454 - 43.
McKernan MG, Shinnick-Gallagher P. Fear conditioning induces a lasting potentiation of synaptic currents in vitro. Nature. 1997; 390 (6660):607-611. DOI: 10.1038/37605 - 44.
Rogan MT, Stäubli UV, LeDoux JE. Fear conditioning induces associative long-term potentiation in the amygdala. Nature. 1997; 390 (6660):604-607. DOI: 10.1038/37601. Erratum in: Nature 1998;391 (6669):818 - 45.
Huang YY, Kandel ER. Postsynaptic induction and PKA-dependent expression of LTP in the lateral amygdala. Neuron. 1998; 21 :169-178 - 46.
Huang YY, Kandel ER. Low-frequency stimulation induces a pathway-specific late phase of LTP in the amygdala that is mediated by PKA and dependent on protein synthesis. Learning & Memory. 2007; 14 :497-503 - 47.
Li H, Weiss SR, Chuang DM, Post RM, Rogawski MA. Bidirectional synaptic plasticity in the rat basolateral amygdala: Characterization of an activity-dependent switch sensitive to the presynaptic metabotropic glutamate receptor antagonist 2S-alpha-ethylglutamic acid. The Journal of Neuroscience. 1998; 18 :1662-1670 - 48.
Paré D, Quirk GJ, Ledoux JE. New vistas on amygdala networks in conditioned fear. Journal of Neurophysiology. 2004; 92 (1):1-9. DOI: 10.1152/jn.00153.2004 - 49.
Royer S, Martina M, Paré D. Polarized synaptic interactions between intercalated neurons of the amygdala. Journal of Neurophysiology. 2000; 83 :3509-3518 - 50.
Asede D, Bosch D, Lüthi A, Ferraguti F, Ehrlich I. Sensory inputs to intercalated cells provide fear-learning modulated inhibition to the basolateral amygdala. Neuron. 2015; 86 (2):541-554. DOI: 10.1016/j.neuron.2015.03.008. Epub 2015 Apr 2 - 51.
Asede D, Doddapaneni D, Bolton MM. Amygdala intercalated cells: Gate keepers and conveyors of internal state to the circuits of emotion. The Journal of Neuroscience. 2022; 42 (49):9098-9109. DOI: 10.1523/JNEUROSCI.1176-22.2022 - 52.
Hagihara KM, Bukalo O, Zeller M, Aksoy-Aksel A, Karalis N, Limoges A, et al. Intercalated amygdala clusters orchestrate a switch in fear state. Nature. 2021; 594 (7863):403-407. DOI: 10.1038/s41586-021-03593-1. Epub 2021 May 26 - 53.
Stern DB, Wilke A, Root CM. Anatomical connectivity of the intercalated cells of the amygdala. eNeuro. 2023; 10 (10):ENEURO.0238-23.2023. DOI: 10.1523/ENEURO.0238-23.2023 - 54.
Huber D, Veinante P, Stoop R. Vasopressin and oxytocin excite distinct neuronal populations in the central amygdala. Science. 2005; 308 (5719):245-248. DOI: 10.1126/science.1105636 - 55.
Neugebauer V, Mazzitelli M, Cragg B, Ji G, Navratilova E, Porreca F. Amygdala, neuropeptides, and chronic pain-related affective behaviors. Neuropharmacology. 2020; 170 :108052. DOI: 10.1016/j.neuropharm.2020.108052. Epub 2020 Mar 15 - 56.
Paré D, Smith Y. GABAergic projection from the intercalated cell masses of the amygdala to the basal forebrain in cats. The Journal of Comparative Neurology. 1994; 344 (1):33-49. DOI: 10.1002/cne.903440104 - 57.
Tamamaki N, Yanagawa Y, Tomioka R, Miyazaki J, Obata K, Kaneko T. Green fluorescent protein expression and colocalization with calretinin, parvalbumin, and somatostatin in the GAD67-GFP knock-in mouse. The Journal of Comparative Neurology. 2003; 467 (1):60-79. DOI: 10.1002/cne.10905 - 58.
Ono M, Yanagawa Y, Koyano K. GABAergic neurons in inferior colliculus of the GAD67-GFP knock-in mouse: Electrophysiological and morphological properties. Neuroscience Research. 2005; 51 (4):475-492. DOI: 10.1016/j.neures.2004.12.019 - 59.
Marowsky A, Yanagawa Y, Obata K, Vogt KE. A specialized subclass of interneurons mediates dopaminergic facilitation of amygdala function. Neuron. 2005; 48 (6):1025-1037. DOI: 10.1016/j.neuron.2005.10.029 - 60.
Sugita S, Tanaka E, North RA. Membrane properties and synaptic potentials of three types of neurone in rat lateral amygdala. Journal of Physiology (London). 1993; 460 :705-718 - 61.
Danober L, Heinbockel T, Driesang RB, Pape H-C. Synaptic mechanisms of NMDA-mediated hyperpolarization in lateral amygdaloid neurons. Neuroreport. 2000; 11 :2501-2506 - 62.
Bauer EP, LeDoux JE. Heterosynaptic long-term potentiation of inhibitory interneurons in the lateral amygdala. The Journal of Neuroscience. 2004; 24 :9507-9512 - 63.
Royer S, Paré D. Bidirectional synaptic plasticity in intercalated amygdala neurons and the extinction of conditioned fear responses. Neuroscience. 2002; 115 :455-462 - 64.
Royer S, Paré D. Conservation of total synaptic weight through balanced synaptic depression and potentiation. Nature. 2003; 422 :518-522 - 65.
Jüngling K, Seidenbecher T, Sosulina L, Lesting J, Sangha S, Clark SD, et al. Neuropeptide S-mediated control of fear expression and extinction: Role of intercalated GABAergic neurons in the amygdala. Neuron. 2008; 59 :298-310 - 66.
Likhtik E, Popa D, Apergis-Schoute J, Fidacaro GA Jr, Pare D. Amygdala intercalated neurons are required for expression of fear extinction. Nature. 2008; 454 :642-645 - 67.
Meis S, Bergado-Acosta JR, Yanagawa Y, Obata K, Stork O, Munsch T. Identification of a neuropeptide S responsive circuitry shaping amygdala activity via the endopiriform nucleus. PLoS One. 2008; 3 :e2695 - 68.
Millhouse OE. The intercalated cells of the amygdala. The Journal of Comparative Neurology. 1986; 247 (2):246-271. DOI: 10.1002/cne.902470209 - 69.
Geracitano R, Kaufmann WA, Szabo G, Ferraguti F, Capogna M. Synaptic heterogeneity between mouse paracapsular intercalated neurons of the amygdala. The Journal of Physiology. 2007; 585 :117-134 - 70.
Klausberger T, Somogyi P. Neuronal diversity and temporal dynamics: The unity of hippocampal circuit operations. Science. 2008; 321 :53-57 - 71.
Sosulina L, Meis S, Seifert G, Steinhauser C, Pape H-C. Classification of projection neurons and interneurons in the rat lateral amygdala based upon cluster analysis. Molecular and Cellular Neurosciences. 2006; 33 :57-67 - 72.
Herry C, Ciocchi S, Senn V, Demmou L, Müller C, Lüthi A. Switching on and off fear by distinct neuronal circuits. Nature. 2008; 454 (7204):600-606. DOI: 10.1038/nature07166. Epub 2008 Jul 9 - 73.
Luchkina NV, Bolshakov VY. Mechanisms of fear learning and extinction: Synaptic plasticity-fear memory connection. Psychopharmacology. 2019; 236 (1):163-182. DOI: 10.1007/s00213-018-5104-4. Epub 2018 Nov 10 - 74.
Meulders A. Fear in the context of pain: Lessons learned from 100 years of fear conditioning research. Behaviour Research and Therapy. 2020; 131 :103635. DOI: 10.1016/j.brat.2020.103635. Epub 2020 Apr 30 - 75.
Labrenz F, Woud ML, Elsenbruch S, Icenhour A. The good, the bad, and the ugly-chances, challenges, and clinical implications of avoidance research in psychosomatic medicine. Frontiers in Psychiatry. 2022; 13 :841734. DOI: 10.3389/fpsyt.2022.841734 - 76.
Meisner OC, Nair A, Chang SWC. Amygdala connectivity and implications for social cognition and disorders. Handbook of Clinical Neurology. 2022; 187 :381-403. DOI: 10.1016/B978-0-12-823493-8.00017-1 - 77.
Alvarez RP, Biggs A, Chen G, Pine DS, Grillon C. Contextual fear conditioning in humans: Cortical-hippocampal and amygdala contributions. The Journal of Neuroscience. 2008; 28 (24):6211-6219. DOI: 10.1523/JNEUROSCI.1246-08.2008 - 78.
Geng H, Xu P, Aleman A, Qin S, Luo YJ. Dynamic organization of large-scale functional brain networks supports interactions between emotion and executive control. Neuroscience Bulletin. 2024. DOI: 10.1007/s12264-023-01168-w - 79.
Tassone VK, Gholamali Nezhad F, Demchenko I, Rueda A, Bhat V. Amygdala biomarkers of treatment response in major depressive disorder: An fMRI systematic review of SSRI antidepressants. Psychiatry Research: Neuroimaging. 2024; 338 :111777. DOI: 10.1016/j.pscychresns.2023.111777. Epub 2024 Jan 3 - 80.
Battaglia S, Di Fazio C, Mazzà M, Tamietto M, Avenanti A. Targeting human glucocorticoid receptors in fear learning: A multiscale integrated approach to study functional connectivity. International Journal of Molecular Sciences. 2024; 25 (2):864. DOI: 10.3390/ijms25020864 - 81.
Traina G, Tuszynski JA. The neurotransmission basis of Post-traumatic stress disorders by the fear conditioning paradigm. International Journal of Molecular Sciences. 2023; 24 (22):16327. DOI: 10.3390/ijms242216327 - 82.
Ritger AC, Parker RK, Trask S, Ferrara NC. Elevated fear states facilitate ventral hippocampal engagement of basolateral amygdala neuronal activity. Frontiers in Behavioral Neuroscience. 2024; 18 :1347525. DOI: 10.3389/fnbeh.2024.1347525 - 83.
Plas SL, Tuna T, Bayer H, Juliano VAL, Sweck SO, Arellano Perez AD, et al. Neural circuits for the adaptive regulation of fear and extinction memory. Frontiers in Behavioral Neuroscience. 2024; 18 :1352797. DOI: 10.3389/fnbeh.2024.1352797 - 84.
Pape HC, Stork O. Genes and mechanisms in the amygdala involved in the formation of fear memory. Annals of the New York Academy of Sciences. 2003; 985 :92-105. DOI: 10.1111/j.1749-6632.2003.tb07074.x - 85.
Pape HC, Narayanan RT, Smid J, Stork O, Seidenbecher T. Theta activity in neurons and networks of the amygdala related to long-term fear memory. Hippocampus. 2005; 15 (7):874-880. DOI: 10.1002/hipo.20120 - 86.
Stubbendorff C, Hale E, Bast T, Cassaday HJ, Martin SJ, Suwansawang S, et al. Dopamine D1-like receptors modulate synchronized oscillations in the hippocampal-prefrontal-amygdala circuit in contextual fear. Scientific Reports. 2023; 13 (1):17631. DOI: 10.1038/s41598-023-44772-6 - 87.
Pirazzini G, Starita F, Ricci G, Garofalo S, di Pellegrino G, Magosso E, et al. Changes in brain rhythms and connectivity tracking fear acquisition and reversal. Brain Structure & Function. 2023; 228 (5):1259-1281. DOI: 10.1007/s00429-023-02646-7. Epub 2023 May 2