Open Access is an initiative that aims to make scientific research freely available to all. To date our community has made over 100 million downloads. It’s based on principles of collaboration, unobstructed discovery, and, most importantly, scientific progression. As PhD students, we found it difficult to access the research we needed, so we decided to create a new Open Access publisher that levels the playing field for scientists across the world. How? By making research easy to access, and puts the academic needs of the researchers before the business interests of publishers.
We are a community of more than 103,000 authors and editors from 3,291 institutions spanning 160 countries, including Nobel Prize winners and some of the world’s most-cited researchers. Publishing on IntechOpen allows authors to earn citations and find new collaborators, meaning more people see your work not only from your own field of study, but from other related fields too.
To purchase hard copies of this book, please contact the representative in India:
CBS Publishers & Distributors Pvt. Ltd.
www.cbspd.com
|
customercare@cbspd.com
HCV’s considerable genetic variability, which exists at various levels across viral populations in individual infected individuals at any given moment and during evolution, is a distinguishing feature of the virus. Because of this, it was discovered in 1993 through phylogenetic analysis of incomplete HCV sequences from several patient isolates worldwide that the virus could be divided into six major genotypes with significant subtypes. Based on a study of full-length ORF sequences, this categorisation was later verified. A seventh significant genotype has been identified, albeit only detected in a few people. An eight genotype has also been recently identified. The number of published ORF sequenced HCV isolates has dramatically increased because of breakthroughs in sequence analysis tools. This chapter seeks to identify the 7 main genotypes and 93 additional subtypes of HCV.
Department of Microbiology, Adeleke University, Ede, Nigeria
Chinonso Chinaza Okezie
Department of Microbiology, Adeleke University, Ede, Nigeria
Oluwaseyi Joy Alao
Department of Otorhinolaryngology, Babcock University Teaching Hospital, Ilishan-Remo, Nigeria
Elijah Oluwatosin Olopade
Department of Biochemistry, Adeleke University, Ede, Nigeria
Isaac Omotosho Komolafe
Department of Biological Sciences, Redeemers University, Ede, Nigeria
*Address all correspondence to: alaojude@adelekeuniversity.edu.ng
1. Introduction
Much emphasis has been paid to the phylogeny and molecular evolution of HCV over the last two decades. These events have far-reaching ramifications for viral taxonomy and disease epidemiology, as well as pathogen control and tracing. Furthermore, they are critical components in diagnoses, treatment regimen selection, and patient follow-up schedules, as well as vaccine development. In this paper, we outline the current perspective of HCV phylogeny and examine the genesis, distribution, and clinical significance of HCV genotypes. In addition, we describe the available evidence on HCV molecular evolution in the context of host-virus interplay at various biological levels, during the disease course, and after treatment.
The genetic diversity of the hepatitis C virus (HCV) is quite significant, and the variety of HCV genotypes and subtypes is growing. HCV was formerly divided into 7 different genotypes that varied by more than 30% at the nucleotide level [1]. Four individuals from the Indian state of Punjab who were epidemiologically unrelated were recently found to have the unique HCV genotype 8, which forms a different phylogenetic group from previously reported genomes [2]. Subtypes of genotypes having a sequence divergence of less than 15% are further subdivided [1]. HCV genotypes 1, 2, and 3 are present globally, albeit their distribution varies depending on the region [3].
The most common HCV genotype worldwide (46%) is genotype 1. While subtypes 1a and 1b of the HCV virus are more common in North America, Europe, and Australia, subtype 1b infection affects 73% of HCV-infected people in Japan. Regardless of location, persons who inject drugs (PWIDs) have a disproportionately large distribution of genotype 3, the second most common genotype in the world (30%) and is mainly found in South Asia. HCV genotype 4 infections are primarily prevalent in Africa and the Middle East, while genotypes 5 and 6 are only found in Southern Africa and Southeast Asia, respectively [4]. Multiple subtypes make up genotypes 1, 2, 3, 4, and 6, exhibiting a high genetic diversity level. HCV genotype 7a was discovered in a patient from the Democratic Republic of the Congo in 2006. Another patient from the same area was later found to have genotype 7b infection [1, 5]. Only one subtype has been documented for genotype 5 and the recently discovered genotype 8.
It is essential to describe novel subtypes and comprehend the potential effects of novel subtypes on treatment success, given the substantial genetic variety of HCV, both at the genotype and subtype levels. This chapter reviews a thorough investigation of viral diversity and sequence variation across genotypes to uncover uncharacterised subgroups and their impact on treatment outcomes.
From the 18 mentioned in 2005 (1), there are now more verified genotypes and subtypes: 67 in 2013 (2), 86 in 2017, 90 in May 2019, and 93 in March 2022 (Table 1). The first virtually entire genome sequence of the HCV was published in 1989. Prior to this discovery, HCV had developed and spread unnoticed across the human population for hundreds of years, giving birth to a diverse range of endemic and epidemic isolates capable of causing chronic liver disease. It quickly became apparent that isolates from various people or nations had significant genetic variability [63]. This variation was compiled after thorough research and surveys by organisations from around the world, and variants were assigned as genotypes and subtypes in a consensus classification and nomenclature system. Official standards were also established for the assignment and naming of future variants [6]. Phylogenetic groups distinct from previously described sequences, at least three epidemiologically unrelated isolates, one or more complete coding region sequences, and the exclusion of intergenotypic or intersubtypic recombination, regardless of whether the components were classified, are all requirements for genotype and subtype assignments [1]. Using these criteria validated the identification of six unique genotypes with 18 subtypes. In addition, 58 subtypes were temporarily designated, awaiting the discovery of further isolates or complete sequencing for the coding area. This consensus on nomenclature was mirrored by the creation of several curated databases, including the Los Alamos HCV Sequence Database, the euHCVdb [64], and the Hepatitis Virus Database (http://s2as02.genes.nig.ac.jp/), which organised HCV sequences as they became available and indicated which genotypes and subtypes were confirmed or provisionally assigned. A proposal to standardise the numbering of HCV concerning genotype 1a isolate H77 was made concurrently (AF009606) [65].
Confirmed HCV genotypes and subtypes (march, 2022) (adapted from Simmonds et al. [6]).
Gene/subtype names that have been unanimously suggested. Two sequences of an HCV genotype have been listed where more than one is available, with the sequences prioritised by (i) publication date, or (ii) submission date when unpublished.
Locus (or isolate name if locus is the same as the accession number).
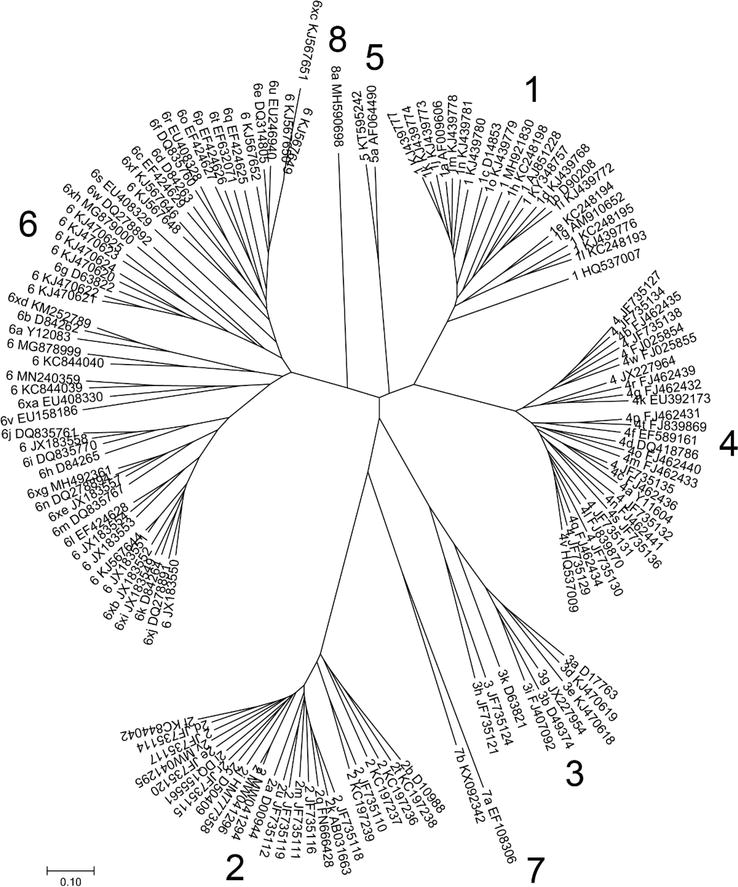
SSEv1.1 [66] and Muscle v3.8.31 [67] were used to align with unique HCV entire or nearly complete coding area sequences from NCBI Genome (969 sequences, http://www.ncbi.nlm.nih.gov/genome) and the Los Alamos HCV sequence database (1364 sequences >8000 nt from http://hcv.lanl.gov/content/index). Seven significant phylogenetic groups corresponding to genotypes 1 through 8 are revealed by phylogenetic analysis of sequences that comprise >95% of the coding area (Figure 1). 100% of bootstrap replications support clustering of the constituent subtypes within these genotypes.
Figure 1.
The typical full coding area sequences of HCV are arranged in this phylogenetic tree. As proposed in the 2005 consensus proposal (1), for an HCV isolate to be considered as a new confirmed genotype or subtype, a complete coding region sequence that: (a) forms a distinct phylogenetic group from previously described sequences, (b) is represented by at least three epidemiologically unrelated isolates, and (c) does not represent a recombinant between other genotypes or subtypes should be obtained.
According to the consensus criteria, confirmed subtypes (indicated by a letter after the genotype) need sequence data from at least two other isolates in core/E1 (>90% of the sequence corresponding to positions 869 to 1292 of the H77 reference sequence [accession number AF009606] numbered according to reference [65]) and NS5B (>90% of pos (Table 1) [6].
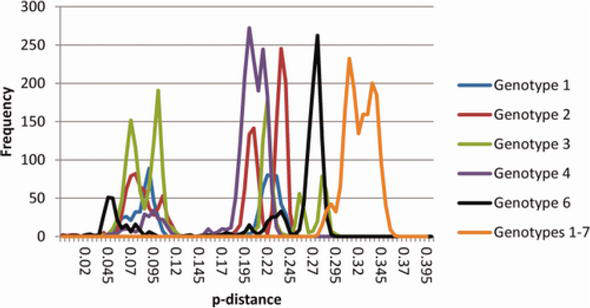
Analysis of the many possible subtypes that have been sequenced (Figure 2) lends credence to using a 15% criterion throughout the coding area. Except for the distances of 14 and 14.2% between JX227963 and two subtype 4 g sequences, this shows significant and regular gaps in the pairwise distances within and between each genotype’s subtype distribution, which were dispersed as follows.: genotype 1: 12.9–17.0%, genotype 2: 13.1–17.6%, genotype 3: 12.5–19.6%, genotype 4: 12.7–15.3%, and genotype 6: 9.9–14.9% (with the exception of the 13.1–13.7% between EU246931 and three subtype 6e sequences). Therefore, a substantial distinction between isolates that differ by 13% throughout their full coding area sequences (members of the same subtype) and those that differ by >15% can be determined for all genotypes with very few exceptions (different genotypes or subtypes). Sequences that are not currently represented by three or more independent isolates of recognised HCV subtypes but are different from any of those subtypes are included in this chapter. It is uncertain if the reported outliers result from different epidemiological histories or technological issues [1].
Figure 2.
Distribution of p-distances between sequences with entire coding regions. Using SSE, the frequency of p-distances within and between genotypes was determined. With the exception of subtypes 1a, 1b, and 2b, where 20 random sequences were employed [66], intra-genotypes pairwise distances were determined for all accessible full coding area sequences. Frequencies were adjusted to get the maximum frequency down to under 300 for p-distances >0.15 (corresponding to a percent difference of 15%). The frequencies scaled as above, and distances between genotypes were determined using one or two samples of each confirmed and unassigned subtype (adapted from [1]).
A few variations with contradictory assignments were discovered during the production of these tables. Isolates P026, P212, and P245 (FJ025854-6) are classified as subtype 4b [38], although their full coding region sequences only share 85% similarity with isolate Z1 (U10235, L16677), which is provisionally designated as 4b [69] and more closely linked to isolate QC264’s core/E1 (FJ46243516 [34]). A third isolate (P213, GU049362) has the NS5B sequence for the same new subtype that P212 and P245 belong to, making this verified subtype 4w. Despite being represented by a single nucleotide that varies from all other genotype 4 sequences by more than 17.5%, isolate P026 is presently unassigned.
Similar to this, isolates KM45 and KM41 (DQ278891,3) have been identified as subtype 6 k [58]. However, they vary from each other and isolate VN405 (D84264) by 6.7 and > 17%, respectively, in the entire coding area sequence, leaving them to be a genotype 6 undefined subtype. Subtype 6u has been assigned to two different groups of isolates: EU408330-2 [51] and EU246940 [47]. The latter was submitted to GenBank first and is represented by NS5B sequences from two additional isolates; as a result, it was given subtype 6u, whereas EU408330, EU408331, and EU408332 were given the subtype 6xa designation.
Finally, Smith et al. [1] analysis of sequence divergence and phylogenetic groupings opines that several isolates [52] classified as “subtype k-related” (TV257, KM35, QC273, TV476), “subtype l-related” (L349, TV533), “intermediate between subtypes 6m and 6n” (DH027), or “intermediate among subtypes 6j and 6i” (QC271) in their GenBank accessions should be regarded as unassigned novel subtypes.
2.1 Further levels of taxonomy
There are challenges in imposing a discrete categorisation method on a complicated taxonomy when defining this taxonomic difference between viral genotypes and subtypes. There are probably many taxonomic hierarchies, particularly for genotypes 3 and 6. For instance, a clade formed by many genotype 6 isolates with subtypes 6 k and 6 l [52]. These sequences and subtypes 6 m and 6n are part of a higher-level clade, while these subtypes and subtypes 6i and 6j are part of another grouping (Figure 1). The discontinuous distribution of p-distances across full coding area sequences (Figure 2), which consists of three practically overlapping ranges (approximately 15–20%, 20–25%, and 25–30%), reflects these evolutionary hierarchies.
2.2 Anticipated developments
The approach for categorising variations into genotypes and subtypes has proven unexpectedly reliable despite the increased amount and diversity of HCV sequences. The partitioning of the seven verified genotypes into subtypes that differ across a whole coding area sequence by >15% represents a natural break in the distribution of sequence distances, and the seven confirmed genotypes show significant bootstrap support (Figure 2). There are still some questions regarding the endemic region of genotype 5, which is represented by a single subtype that has been isolated in Europe, Brazil, North Africa, and South Africa, and genotype 7, which has been isolated from a Congolese immigrant. We may also expect to find more HCV-like viruses in the genus Hepacivirus [70, 71, 72, 73], as well as variations that are more genetically related to HCV than the non-primate Hepacivirus that appears to be an endemic infection of horses globally [70].
This chapter established that there are 8 genotypes of HCV, and 93 subtypes. Of the 8 genotypes, 7 were highlighted in this chapter. The chapter also attempted to link HCV genotypes to their endemicity. Utilising the phylogenetic classification of HCV can provide a greater insight into eradicating the virus by 2030.
We thank Dr. Donald B. Smith of the University of The University of Edinburgh for granting us permission to use the figures and tables in his earlier published paper.
References
1.Smith DB, Bukh J, Kuiken C, Muerhoff AS, Rice CM, Stapleton JT, et al. Expanded classification of hepatitis C virus into 7 genotypes and 67 subtypes: Updated criteria and genotype assignment web resource. Hepatology (Baltimore, Md.). 2014;59(1):318-327
2.Borgia SM, Hedskog C, Parhy B, et al. Identification of a novel hepatitis C virus genotype from Punjab, India: Expanding classification of hepatitis C virus into 8 genotypes. The Journal of Infectious Diseases. 2018;218:1722-1729
3.Messina JP, Humphreys I, Flaxman A, et al. Global distribution and prevalence of hepatitis C virus genotypes. Hepatology. 2015;61:77-87
4.Asselah T, Hassanein T, Waked I, et al. Eliminating hepatitis C within low-income countries—The need to cure genotypes 4, 5, 6. Journal of Hepatology. 2017;68:814-826
5.Schreiber J, McNally J, Chodavarapu K, et al. Treatment of a patient with genotype 7 hepatitis C virus infection with sofosbuvir and velpatasvir. Hepatology. 2016;64:983-985
6.Simmonds P, Bukh J, Combet C, Deléage G, Enomoto N, Feinstone S, et al. Consensus proposals for a unified system of nomenclature of hepatitis C virus genotypes. Hepatology. 2005;43:962-973
7.Choo QL, Richman KH, Han JH, Berger K, Lee C, Dong C, et al. Genetic organization and diversity of the hepatitis C virus. Proceedings of the National Academy of Sciences of the United States of America. 1991;88:2451-2455
8.Inchauspe G, Zebedee S, Lee DH, Sugitani M, Nasoff M, Prince AM. Genomic structure of the human prototype strain H of hepatitis C virus: Comparison with American and Japanese isolates. Proceedings of the National Academy of Sciences of the United States of America. 1991;88:10292-10296
9.Kato N, Hijikata M, Ootsuyama Y, Nakagawa M, Ohkoshi S, Sugimura T, et al. Molecular cloning of the human hepatitis C virus genome from Japanese patients with non-A, non-B hepatitis. Proceedings of the National Academy of Sciences of the United States of America. 1990;87:9524-9528
10.Takamizawa A, Mori C, Fuke I, Manabe S, Murakami S, Fujita J, et al. Structure and organization of the hepatitis C virus genome isolated from human carriers. Journal of Virology. 1991;65:1105-1113
11.Okamoto H, Kojima M, Sakamoto M, Iizuka H, Hadiwandowo S, Suwignyo S, et al. The entire nucleotide sequence and classification of a hepatitis C virus isolate of a novel genotype from an Indonesian patient with chronic liver disease. Journal of General Virology. 1994;75:629-635
12.Lu L, Li C, Xu Y, Murphy DG. Full-length genomes of 16 hepatitis C virus genotype 1 isolates representing subtypes 1c, 1d, 1e, 1g, 1h, 1i, 1j and 1k, and two new subtypes 1m and 1n, and four unclassified variants reveal ancestral relationships among subtypes. Journal of General Virology. 2014;95:1479-1148
13.Li C, Njouom R, Pépin J, Nakano T, Bennett P, Pybus OG, et al. Characterization of full-length HCV sequences for subtypes 1e, 1h, and 1l, and a novel variant revealed Cameroon as an area in origin for genotype 1. Journal of Virology. 2013;94:1780-1790
14.Bracho MA, Saludes V, Martró E, Bargalló A, González-Candelas F, Ausina V. Complete genome of a European hepatitis C virus subtype 1g isolate: Phylogenetic and genetic analyses. Journal of Virology. 2008;5:72
15.Okamoto H, Okada S, Sugiyama Y, Kurai K, Iizuka H, Machida A, et al. Nucleotide sequence of the genomic RNA of hepatitis C virus isolated from a human carrier: Comparison with reported isolates for conserved and divergent regions. Journal of General Virology. 1991;72:2697-2704
16.Kato T, Furusaka A, Miyamoto M, Date T, Yasui K, Hiramoto J, et al. Sequence analysis of hepatitis C virus isolated from a fulminant hepatitis patient. Journal of Medical Virology. 2001;64:334-339
17.Okamoto H, Kurai K, Okada S, Yamamoto K, Lizuka H, Tanaka T, et al. Full-length sequence of a hepatitis C virus genome having poor homology to reported isolates: Comparative study of four distinct genotypes. Virology. 1992;188:331-341
18.Murakami K, Abe M, Kageyama T, Kamoshita N, Nomoto A. Down-regulation of translation driven by hepatitis C virus internal ribosomal entry site by the 3′ untranslated region of RNA. Archives of Virology. 2001;146:729-741
19.Nakao H, Okamoto H, Tokita H, Inoue T, Iizuka H, Pozzato G, et al. Full-length genomic sequence of a hepatitis C virus genotype 2c isolate (BEBE1) and the 2c-specific PCR primers. Archives of Virology. 1996;141:701-704
20.Li C, Cao H, Lu L, Murphy D. Full-length sequences of 11 hepatitis C virus genotype 2 isolates representing five subtypes and six unclassified lineages with unique geographical distributions and genetic variation patterns. Journal of General Virology. 2012;93:1173-1184
21.Xu R, Tong W, Gu L, Li C, Fu Y, Lu L. A panel of 16 full-length HCV genomes was characterized in China belonging to genotypes 1-6 including subtype 2f and two novel genotype 6 variants. Infection, Genetics and Evolution. 2013;20:225-229
22.Noppornpanth S, Lien TX, Poovorawan Y, Smits SL, Osterhaus ADME, Haagmans BL. Identification of a naturally occurring recombinant genotype 2/6 hepatitis C virus. Journal of Virology. 2006;80:7569-7577
23.Samokhvalov EI, Hijikata M, Gylka RI, Lvov DK, Mishiro S. Full-genome nucleotide sequence of a hepatitis C virus variant (isolate name VAT96) representing a new subtype within the genotype 2 (arbitrarily 2k). Virus Genes. 2000;20:183-187
24.Jordier F, Deligny ML, Barré R, De Micco P, Cantaloube JF. Evidence for two phylogenetic clusters within hepatitis C virus (HCV) genotype 2 inferred from analysis of complete coding sequences of 15 HCV strains. Journal of Medical Virology. 2013;85:1754-1764
25.Martró E, Valero A, Jordana-Lluch E, Saludes V, Planas R, González-Candelas F, et al. Hepatitis C virus sequences from different patients confirm the existence and transmissibility of subtype 2q, a rare subtype circulating in the metropolitan area of Barcelona, Spain. Journal of Medical Virology;83:820-826
26.Rajhi M, Haddad-Boubaker S, Chouikha A, Bourquain D, Michel J, Hammami W, et al. Identification of two novel hepatitis C virus subtype 2 from Tunisia (2v and 2w). PLoS One. 2021;16:e0248249
27.Sakamoto M, Akahane Y, Tsuda F, Tanaka T, Woodfield DG, Okamoto H. Entire nucleotide sequence and characterization of a hepatitis C virus of genotype V/3a. Journal of General Virology. 1994;75:1761-1768
28.Yamada N, Tanihara K, Mizokami M, Ohba K, Takada A, Tsutsumi M, et al. Full-length sequence of the genome of hepatitis C virus type 3a: Comparative study with different genotypes. Journal of General Virology. 1994;75:3279-3284
29.Chayama K, Tsubota A, Koida I, Arase Y, Saitoh S, Ikeda K, et al. Nucleotide sequence of hepatitis C virus (type 3b) isolated from a Japanese patient with chronic hepatitis C. Journal of General Virology. 1994;75:3623-3628
30.Li C, Lu L, Murphy DG, Negro F, Okamoto H. Origin of hepatitis C virus genotype 3 in Africa as estimated through an evolutionary analysis of the full-length genomes of nine subtypes, including the newly sequenced 3d and 3e. Journal of General Virology. 2014;95:1677-1688
31.Lu L, Li C, Yuan J, Lu T, Okamoto H, Murphy DG. Full-length genome sequences of five hepatitis C virus isolates representing subtypes 3g, 3h, 3i and 3k, and a unique genotype 3 variant. Journal of General Virology. 2013;94:543-538
32.Newman RM, Kuntzen T, Weiner B, Berical A, Charlebois P, Kuiken C, et al. Whole genome pyrosequencing of rare hepatitis C virus genotypes enhances subtype classification and identification of naturally occurring drug resistance variants. The Journal of Infectious Diseases. 2013;208:17-31
33.Chamberlain RW, Adams N, Saeed AA, Simmonds P, Elliott RM. Complete nucleotide sequence of a type 4 hepatitis C virus variant, the predominant genotype in the Middle East. Journal of General Virology. 1997;78:1341-1347
34.Li C, Lu L, Wu X, Wang C, Bennett P, Lu T, et al. Complete genomic sequences for hepatitis C virus subtypes 4b, 4c, 4d, 4g, 4k, 4l, 4m, 4n, 4o, 4p, 4q, 4r and 4t. Journal of General Virology. 2009;90:1820-1826
35.Kuntzen T, Berical A, Ndjomou J, Bennett P, Schneidewind A, Lennon N, et al. A set of reference sequences for the hepatitis C genotypes 4d, 4f, and 4k covering the full open reading frame. Journal of Medical Virology. 2008;80:1370-1378
36.Lu L, Xu Y, Yuan J, Li C, Murphy DG. The full-length genome sequences of nine HCV genotype 4 variants representing a new subtype 4s and eight unclassified lineages. Virology. 2015;482:111-116
37.Demetriou VL, Kostrikis LG. Near-full genome characterization of unclassified hepatitis C virus strains relating to genotypes 1 and 4. Journal of Medical Virology. 2011;83:2119-2127
38.Koletzki D, Dumont S, Vermeiren H, Peixe P, Nina J, Camacho RJ, et al. Full genome sequence of three isolates of hepatitis C virus subtype 4b from Portugal. Archives of Virology. 2009;154:127-132
39.Bukh J, Apgar CL, Engle R, Govindarajan S, Hegerich PA, Tellier R, et al. Experimental infection of chimpanzees with hepatitis C virus of genotype 5a: Genetic analysis of the virus and generation of a standardized challenge pool. The Journal of Infectious Diseases. 1998;178:1193-1197
40.Adams NJ, Chamberlain RW, Taylor LA, Davidson F, Lin CK, Elliott RM, et al. Complete coding sequence of hepatitis C virus genotype 6a. Biochemical and Biophysical Research Communications. 1997;234:393-396
41.Tokita H, Okamoto H, Iizuka H, Kishimoto J, Tsuda F, Miyakawa Y, et al. The entire nucleotide sequences of three hepatitis C virus isolates in genetic groups 7-9 and comparison with those in the other eight genetic groups. Journal of General Virology. 1998;79:1847-1857
42.Lu L, Li C, Fu Y, Gao F, Pybus OG, Abe K, et al. Complete genomes of hepatitis C virus (HCV) subtypes 6c, 6l, 6o, 6p and 6q: Completion of a full panel of genomes for HCV genotype 6. Journal of General Virology. 2007;88:1519-1525
43.Li C, Fu Y, Lu L, Ji W, Yu J, Hagedorn CH, et al. Complete genomic sequences for hepatitis C virus subtypes 6e and 6g isolated from Chinese patients with injection drug use and HIV-1 co-infection. Journal of Medical Virology. 2006;78:1061-1069
44.Lu L, Li C, Fu Y, Thaikruea L, Thongswat S, Maneekarn N, et al. Complete genomes for hepatitis C virus subtypes 6f, 6i, 6j and 6m: Viral genetic diversity among Thai blood donors and infected spouses. Journal of General Virology. 2007;88:1505-1518
45.Tokita H, Okamoto H, Iizuka H, Kishimoto J, Tsuda F, Lesmana LA, et al. Hepatitis C virus variants from Jakarta, Indonesia classifiable into novel genotypes in the second (2e and 2f), tenth (10a) and eleventh (11a) genetic groups. Journal of General Virology;77:293-301
46.Li C, Lu L, Zhang X, Murphy D. Entire genome sequences of two new HCV subtypes, 6r and 6s, and characterization of unique HVR1 variation patterns within genotype 6. Journal of Viral Hepatology. 2009;16:406-417
47.Noppornpanth S, Poovorawan Y, Lien TX, Smits SL, Osterhaus ADME, Haagmans BL. Complete genome analysis of hepatitis C virus subtypes 6t and 6u. Journal of General Virology. 2008;89:1276-1281
48.Lu L, Murphy D, Li C, Liu S, Xia X, Pham PH, et al. Complete genomes of three subtype 6t isolates and analysis of many novel hepatitis C virus variants within genotype 6. Journal of General Virology. 2008;89:444-452
49.Wang Y, Xia X, Li C, Maneekarn N, Xia W, Zhao W, et al. A new HCV genotype 6 subtype designated 6v was confirmed with three complete genome sequences. Journal of Clinical Virology. 2009;44:195-199
50.Lee YM, Lin HJ, Chen YJ, Lee CM, Wang SF, Chang KY, et al. Molecular epidemiology of HCV genotypes among injection drug users in Taiwan: Full-length sequences of two new subtype 6w strains and a recombinant form_2b6w. Journal of Medical Virology. 2010;82:57-68
51.Xia X, Zhao W, Tee KK, Feng Y, Takebe Y, Li Q , et al. Complete genome sequencing and phylogenetic analysis of HCV isolates from China reveals a new subtype, designated 6u. Journal of Medical Virology. 2008;80:1740-1746
52.Wang H, Yuan Z, Barnes E, Yuan M, Li C, Fu Y, et al. Eight novel hepatitis C virus genomes reveal the changing taxonomic structure of genotype 6. Journal of General Virology. 2013;94:76-80
53.Li C, Pham VH, Abe K, Lu L. Nine additional complete genome sequences of HCV genotype 6 from Vietnam including new subtypes 6xb and 6xc. Virology. 2014;468-470:172-177
54.Ye M, Chen X, Wang Y, Duo L, Zhang C, Zheng YT. Identification of a new HCV subtype 6xg among injection drug users in Kachin, Myanmar. Frontiers Microbiology. 2019;10:814
55.Li C, Barnes E, Newton PN, Fu Y, Vongsouvath M, Klenerman P, et al. An expanded taxonomy of hepatitis C virus genotype 6: Characterization of 22 new full-length viral genomes. Virology. 2015;476:355-363
56.Wu T, Xing Z, Yuan M, Ge J, Yuan G, Liang K, et al. Analysis of HCV isolates among the Li ethnic in Hainan Island of South China reveals their HCV-6 unique evolution and a new subtype. Cellular Physiology and Biochemistry. 2018;50:1832-1839
57.Yue W, Feng Y, Jia Y, Liu Y, Zhang Y, Geng J, et al. Identification of a new HCV subtype 6xi among chronic hepatitis C patients in Yunnan, China. Journal of Infection. 2020;80:469-496
58.Lu L, Nakano T, Li C, Fu Y, Miller S, Kuiken C, et al. Hepatitis C virus complete genome sequences identified from China representing subtypes 6k and 6n and a novel, as yet unassigned subtype within genotype 6. Journal of General Virology. 2006;87:629-634
59.Jia Y, Yue W, Gao Q , Tao R, Zhang Y, Fu X, et al. Characterization of a novel hepatitis C subtype, 6xj, and its consequences for direct-acting antiviral treatment in Yunnan, China. Microbiology Spectrum. 2021;9:e0029721
60.Murphy DG, Sablon E, Chamberland J, Fournier E, Dandavino R, Tremblay CL. Hepatitis C virus genotype 7, a new genotype originating from Central Africa. Journal of Clinical Microbiology. 2015;53:967-972
61.Salmona M, Caporossi A, Simmonds P, Thélu MA, Fusillier K, Mercier-Delarue S, et al. First next-generation sequencing full-genome characterization of a hepatitis C virus genotype 7 divergent subtype. Clinical Microbiology and Infection. 2016;22:947.e941-947.e948
62.Borgia SM, Hedskog C, Parhy B, Hyland RH, Stamm LM, Brainard DM, et al. Identification of a novel hepatitis C virus genotype from Punjab, India: Expanding classification of hepatitis C virus into 8 genotypes. Journal of Infectious Diseases. 2018;218:1722-1729
63.Choo Q , Kuo G, Weiner A, Overby L, Bradley D, Houghton M. Isolation of a cDNA clone derived from a blood-borne non-A, non-B viral hepatitis genome. Science. 1989;80(244):359-362
64.Combet C, Garnier N, Charavay C, Grando D, Crisan D, Lopez J, et al. euHCVdb: The European hepatitis C virus database. Nucleic Acids Research. 2007;35:D363-D366
65.Kuiken C, Combet C, Bukh J, Shin-I T, Deleage G, Mizokami M, et al. A comprehensive system for consistent numbering of HCV sequences, proteins and epitopes. Hepatology. 2006;44:1355-1361
66.Simmonds P. SSE: A nucleotide and amino acid sequence analysis platform. BMC Research Notes. 2012;5:50
67.Edgar RC. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Research. 2004;32:1792-1797
68.Classification and genotype/subtype assignments of hepatitis C virus. Flaviviridae: Hepacivirus C Classification. 2022. Available from: https://ictv.global/sg_wiki/fla viviridae/hepacivirus [Accessed: December 20, 2022]
69.Bukh J, Purcell RH, Miller RH. At least 12 genotypes of hepatitis C virus predicted by sequence analysis of the putative E1 gene of isolates collected worldwide. Proceedings of the National Academy of Sciences of the United States of America. 1993;90:8234-8238
70.Kapoor A, Simmonds P, Cullen JM, Scheel TKH, Medina JL, Giannitti F, et al. Identification of a pegivirus (GB virus-like virus) that infects horses. Journal of Virology. 2013;87:7185-7190
71.Kapoor A, Simmonds P, Scheel T, Hjelle B, Cullen J, Burbelo P, et al. Identification of rodent homologs of hepatitis C virus and pegiviruses. MBio. 2013;4:e00216-e00213
72.Quan P-L, Firth C, Conte JM, Williams SH, Zambrana-Torrelio CM, Anthony SJ, et al. Bats are a major natural reservoir for hepaciviruses and pegiviruses. Proceedings of the National Academy of Sciences of the United States of America. 2013;110:8194-8199
73.Drexler JF, Corman VM, Müller MA, Lukashev AN, Gmyl A, Coutard B, et al. Evidence for novel hepaciviruses in rodents. PLoS Pathogens. 2013;9:e1003438
Written By
Jude Oluwapelumi Alao, Chinonso Chinaza Okezie, Oluwaseyi Joy Alao, Elijah Oluwatosin Olopade and Isaac Omotosho Komolafe
Submitted: 23 December 2022Reviewed: 24 December 2022Published: 17 February 2023
 Open access peer-reviewed chapter
Open access peer-reviewed chapter