
Abstract
The review presents data on the role of leaf epidermis, lignin, and its monolignols in the adaptation of higher plants to adverse environmental conditions. It discusses the localization and content of syringyl monolignin and guajacyl monolignin in cell walls and how they affect the elasticity of plant cell walls under different natural conditions. These data are presented using modern methods of laser scanning, confocal microscopy, and scanning electron microscopy. The review also discusses literature data and the results of our own experimental studies on the cellular mechanisms of lignin synthesis and its regulation, as well as the participation of monolignols in plant adaptation to environmental changes. It shows the role of monolignols in regulating water balance and transpiration of plants, as well as in protecting plant cells from abiotic stresses and environmental changes. Recent studies have also shown the influence of lignin on the expression of genes involved in the synthesis of secondary cell walls and metabolites with protective properties. This review indicates the potential for further research into the role of monolignols and confirms that the conservation of species characterized by increased lignin synthesis may be a conceptual basis for the protection and conservation of flora from abiotic stresses.
Keywords
- cell wall structure
- syringyl
- guajacyl
- flooding
- drought
- cold
- stress
- genes
1. Introduction
Over the past decade, research has shown that the structure and composition of plant epidermis cell walls change in response to various environmental factors. This applies to both terrestrial plants and hydrophytes, which grow in water or on the shore near water. Environmental changes can disrupt cell functions such as growth, water and sun photon absorption, transpiration, strength, and protection. The epidermis of leaves and stems serves as the first barrier between the plant and its environment, protecting it from biotic and abiotic factors such as flooding, extreme temperatures, and pathogen invasion [1, 2, 3]. The leaf epidermis is defined by a combination of structural characteristics and the chemical composition of cell walls, which can change under stress conditions. Trichomes and epidermal wax in the epidermal cells play a role in regulating the absorption of photon energy, decreasing photoinhibition, and reflecting ultraviolet rays. Additionally, the cells of the leaf epidermis play a key role in the development of plants [4, 5, 6]. It is known that one of the main components of the epidermal cell walls, responsible for mechanical strength, leaf elasticity, and water and aqueous solution transport, is lignin and its constituent monolignols. These properties of lignin depend on the content and ratio of its constituent monolignols, which are formed in independent ways [7].
Drought and soil flooding are factors that impact plant growth and species conservation. The study of these environmental factors began with the development of nature ecology and has led to new possibilities for investigating the effects of environmental stress on plant species conservation and vital plant functions. It has been established that the biogenesis and function of leaf epidermis are mediated by the activation of specific enzymes in plants. These studies are described in a series of reviews [7, 8, 9]. Understanding the structural and functional characteristics of leaf epidermis under normal conditions and adapting to stress factors is important. This chapter reviews current knowledge about the structure and function of the epidermis of hydrophytes and terrestrial plants, as well as the participation of lignin and its monolignols under the influence of environmental change.
2. The impact of flooding on leaf epidermis and monolignols
Flooding is a potentially harmful stress for land plants caused by both natural (river flooding, heavy precipitation, and sea tides) and artificial causes (construction of reservoirs and ponds). It can be short-term or long-term, leading to the death of some plant species and crops. Flooding occurs when soils are waterlogged or when irrigated lands are not properly maintained. The combination of changes in salinity and impaired oxygen respiration in the root system inhibits aerobic processes, impairs the absorption of ions from the soil, and affects growth processes. Aerenchyma is formed in the roots, aerobic processes are inhibited, and fermentation begins with the formation of lactic acid, leading to acidification of the environment around the root system [10]. When flooded, plants are affected by reduced lighting, changes in spectrum, a lack of oxygen, and CO2. Water absorbs light flux, and the attenuation is directly proportional to the water depth. Light is also absorbed by organic and inorganic particles in the water, with the long-wavelength parts of the spectrum being absorbed at greater depths [11].
Underwater plant leaves are impacted by a lack of light and CO2, which has high solubility but a low diffusion coefficient in fresh water (1.7 × 10−9 m2/c at 20°C). The pH of fresh water also affects gas solubility and the ratio of CO2 to O2 in water [12]. The air contains 0.03% carbon dioxide, 78.09% nitrogen, and 20.95% oxygen. In water, the amount of dissolved oxygen depends on atmospheric pressure, with higher pressure leading to greater solubility of oxygen. At 1 atm and 25°C, 0.023 g of oxygen is dissolved in 1 kg of water. The solubility of carbon dioxide in water is 200 times higher than that of oxygen [11]. The CO2 content in fresh water is 350 mM m−3, and the diffusion of gases in water is also lower. Underwater plants require 30 times more free CO2 to saturate their leaves [12] and therefore use CO2 ions as a carbon source [13]. Growth conditions in aquatic environments can cause hydrophytes to die when light is limited, and oxygen levels in the water and in plants decrease, leading to the inhibition of carbohydrate metabolism [14].
These adaptations allow hydrophytes to efficiently carry out photosynthesis and obtain the nutrients they need despite the challenging conditions. Additionally, some hydrophytes have specialized air channels and aerenchyma tissue that facilitate gas exchange and help them survive in waterlogged environments. These adaptations are essential for the survival of hydrophytes and marsh plants in their unique habitats [2, 15].
The features aid in acclimatization and support plant resistance to underwater existence, allowing for CO2 to enter photosynthetic cells through diffusion in the outer epidermal walls. The structure of cell walls in underwater leaves and stems differs from that of surface hydrophytes and mesophyte leaves, highlighting the importance of comparative structural and functional studies in understanding plant adaptation to aquatic environments.
2.1 The submerged leaf еpidermis structure
The outer cell walls of the epidermis and leaf stomata act as the first barrier and transport route for CO2 and water, as well as the point of contact with the environment for plant organs. The cuticle of mesophyte leaves’ epidermal walls contains pores that facilitate cuticular transpiration and water absorption. The functionality of cuticular pores in aerial leaves is influenced by the temperature and humidity of the surrounding air [16, 17]. In flooded plants, the cuticle is also synthesized in the epidermis and deposited in the periclinal cuticle sheaths. In certain species of higher aquatic plants (
Aquatic plants, also known as hydrophytes, have a super hydrophobic cuticle on the epidermis of their leaves and stems. This prevents the formation of a water film on the surface of submerged organs, which leads to normal gas exchange in the leaves and to photosynthesis [23, 24]. Recent studies have focused on the structure of the cuticle of epidermal cells in underwater plants, particularly. The presence of cuticular pores in the cell walls of the epidermis of underwater leaves and the functioning of the epidermal cell wall were studied [15, 25, 26].
The study of the ultrastructural peculiarities of wall epidermis in submerged leaves of

Figure 1.
Scanning electron microscopy of the epidermal cells of the submersed leaves of
The surface of the leaf epidermis of
2.2 Monolignols in submergenced leaves of hydrophytes
This section will discuss the presence of monolignols in the leaves of hydrophytes that are subjected to flooding. Lignin is essential for plant adaptation to stress, especially in response to flooding and siltation. It is a polymer of aromatic alcohols synthesized in cell walls that have completed stretching growth. Lignin helps make cell walls impermeable to water and aqueous solutions, forming a barrier to pathogens in the epidermis. It is composed of monolignins formed from p-hydroxyphenyl, guajacyl, and syringyl components, which differ in the degree of methoxylation [29]. Flooding and siltation impact the lignification of cell walls, with flooding being the main effect and mechanism of adaptation of the rice root system due to a lack of oxygen. The study investigated the impact of 50 cm deep flooding on the lignification of rice stems by comparing three rice varieties: flood-resistant (
According to researchers [31, 32], underwater (submerged) plant organs experience stress from the weight of the water column and the mechanical action of waves, leading to tension in flooded organs. The effect of this stress on the structure and functioning of cell walls in flooded plants are still not fully understood. While there is data on the impact of stress on the cell walls of land plants, the specific effects on flooded plants are not well documented. For example, in naked-seeded plants, the wood stress response occurs at the level of several vessel elements that have a small diameter, thick cell walls, high cellulose content, and low lignin [33]. These walls are characterized by a small amount of guajacol, a high content of syringin, and a three-layer structure [34].
A study examined the presence of lignin and monolignols in the leaves of the hydrophytes

Figure 2.
The general view of underwater leaves of
The presence and distribution of lignin in the tissues of underwater and above-water leaves of the hydrophyte
2.3 Floating and above-water leaves
The epidermal structure of floating leaves is adapted to their aquatic environment. These leaves have a thick waxy cuticle and a reduced number of stomata, which helps to minimize water loss. Additionally, the epidermal cells may contain air spaces to provide buoyancy.
The micromorphology and ultrastructure of the epidermis of floating leaves of

Figure 3.
The scanning electron microscopy images display the adaxial (a) and abaxial (b) surfaces of
The anticlinal walls are elevated above the periclinal walls, forming a cup-shaped depression in each cell. Stomata are located on the lower epidermis in no particular order (Figure 3b), with an average density of 33.1 ± 1.7 per 1 μm2 of area. They are of the parasitic type, with accompanying cells located along the guard cells. The usual epidermal cells of the lower epidermis, similarly to those of the upper epidermis, form cushion-like depressions. The epidermal cells vary in shape, with round cells having a diameter of about 10 ± 1.2 μm and oval cells having a long axis ranging from 10 ± 0.7 to 37 ± 2.3 μm and a short axis from 7 ± 0.3 to 22 ± 1.9 μm, respectively. The anticlinal walls are wavy, reaching a height of 7 ± 0.4 μm. Periclinal walls are concave within the leaf blade and are located in depressions. Using the Pascal program (LSM), it has been calculated that there is air under the lower surface of the leaf, in the recesses of the main epidermal cells, with a content of 119,700 ± 2300 μm3 on an area of 1 cm2. Aerenchyma was also revealed between cells of the mesophyll and epidermis. It was established that the leaves of
Our investigation found that the air content under the lower surface of the flooding leaf of
We demonstrated that syringyl and guajacyl monolignols, when combined with the specific fluorescent indicator diphenyl boric acid-2-aminoethyl ester, exhibited bright blue and green fluorescence in the periclinal and anticlinal walls of adaxial and abaxial epidermal cells of
Previous research has shown that an increase in the quantity of syringyl/guajacyl (S/G) acts as a chemical barrier, enhancing cell defense against water penetration and pathogens [50] while also increasing the mechanical durability of cells [51]. Based on our findings, we hypothesize that the elevated content of syringyl in the abaxial epidermis of the studied species can enhance the mechanical properties of the cell walls. Interestingly, the content of guajacyl was found to be similar in the upper and lower epidermal walls. Given the known function of guajacyl in wall softening, it is plausible that its high content in the cell walls of floating leaves of

Figure 4.
Fragments of epidermal cells of surface above-water (a, c, d) and submerged (b) leaves of
The mechanism of cell wall thinning and loosening, particularly in hydrophytes, has not been fully understood. There are several proposed models for cell wall loosening, including associations with decreased cell turgor, reduced activity of polysaccharide synthetases [54], apoplast acidification [55], and activation of phytohormones [56, 57]. It is revealed that phytohormones induce stretching and increase wall plasticity, depending on the apoplast pH [54, 56]. Additionally, phytohormones act as effectors of wall loosening. It has been established that one mechanism of cell wall loosening in higher plant stems is the participation of brassinosteroids, which regulate the expression of genes for loosening proteins, namely xyloglucan endotransglucosylases, and act at the transcriptional level [57].
Studies of lignin in the surface leaves of
The presence and distribution of lignin in the tissues of underwater and above-water leaves of the hydrophyte
A cytochemical analysis of

Figure 5.
The general view of the above-water (a) and submerged (g) leaves of
The relative content of syringyl and guajacyl varied in the walls of the studied tissues. The highest values of guajacyl are found in the epidermis, palisade parenchyma, and vessels. The ratio of syringyl to guajacyl content (S/G) was highest in the abaxial epidermis, followed by the adaxial epidermis, spongy parenchyma, palisade parenchyma, and vessels. Notably, the periclinal walls had higher S/G values compared to the anticlinal walls, regardless of the type of epidermis. An increase in the S/G ratio strengthens the chemical barrier to protect the cell from water penetration and pathogen invasion [50]. Additionally, the S/G ratio indicates an increase in the mechanical strength of cell walls [51]. Leaves immersed in water are constantly in contact with surrounding aquatic microflora and numerous algae, and they can withstand constant water pressure and wave action [15]. Both the periclinal and anticlinal epidermal walls of underwater leaves protect the leaf cell surface from the exogenous effects of the aquatic environment by regulating the synthesis of lignin components.
3. Effect of drought on epidermis structure of plant leaves and monolignols content
Environmental stressors such as drought, salinity, salt fog, solar radiation, high light intensity, low nutrient availability, and high soil temperature affect the structure of a plant’s leaves [60, 61]. These changes can impact the morphology of the leaves, which is directly regulated by primary stress signals that trigger mechanisms for adaptation, crucial for plant survival in stressful habitats [62]. Drought, for example, can lead to a loss of leaf turgor, which is the plant’s ability to maintain cell turgor pressure during dehydration and is a strong predictor of the plant’s response to drought [63]. Various factors, including structural and functional changes in both the roots and leaves, help preserve leaf turgor, particularly in the leaf structure. Drought stress can reduce water potential and turgor, leading to inhibited photosynthesis, metabolism, and potential cell death [64]. Lignin plays a role in maintaining tissue water potential by regulating cuticular and transcriptional transpiration, thus reducing water consumption [65]. Vegetation in coastal sand dunes faces environmental stress such as drought, salinization, solar radiation, high light intensity, low nutrient availability, high temperatures, sand instability, and sea or river breezes [60, 61, 66]. These stressors affect the structure of plant organs [67].
Drought causes changes in leaf structure and plays a significant role in leaf adaptation. Specific patterns in leaf morphology are regulated by primary stress signals, triggering mechanisms of structural and functional adaptation crucial for plant survival in stressful habitats [62]. Structural adaptations to drought conditions include water-storing parenchyma leaf blades, curling, deepening of stomata, and leaf pubescence. These adaptations help psammophytes maintain optimal water balance in plants. A study of 54 taxa in the Boraginaceae family found mineralized structures at the base of leaf trichomes in psammophytic species, as well as the presence of calcium and potassium ions [68].
For example, a study of species specific to sand dunes (Marina di Vecchiano coast, Italy), namely
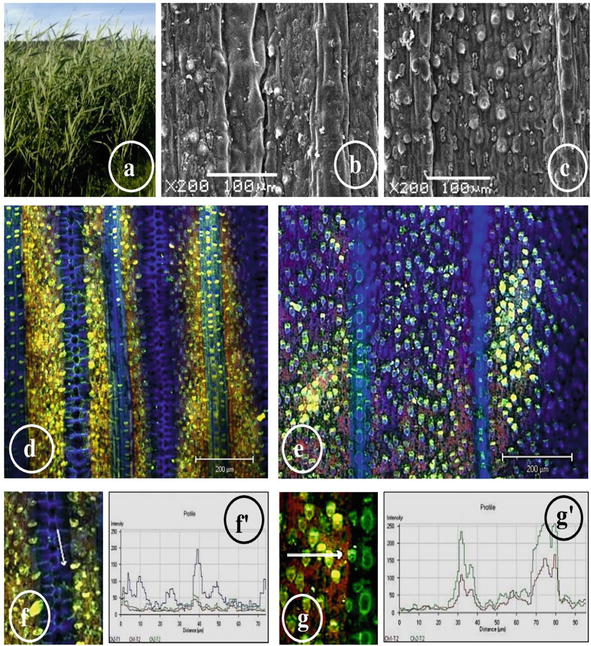
Figure 6.
The general view of
The study of both surfaces of the leaves of
Trichome density also increases during drought [83]. A dense coating of trichomes changes the optical properties of the leaf surface and can reflect or absorb a certain wavelength of light. Trichomes, which capture a layer of air on the leaf surface, can also preserve heat and moisture [73]. In the cells of the epidermis and hypodermis of
The study of the structural and functional characteristics of
A cytochemical and laser confocal microscopic study of monolignols in the leaf epidermis of the air-water and terrestrial ecotypes of
The differences in the relative content of monolignols in different epidermal cells of
It is known that the increase in lignin causes osmotic stress, leading to water loss or cell death and inhibiting plant growth and development [64]. Lignin can decrease water flow by cells, supporting osmotic balance and cell integrity [87, 88, 89]. Considering the literature and our results on the increased S/G ratio in the cells of conductive vessels and cells of the upper epidermis in leaves of the terrestrial ecotype of
According to the literature, drought, similar to soil salinity, has various negative effects on plant growth, such as osmotic stress and inhibition of organ growth [64, 87, 88, 89]. However, plants can adapt to unfavorable conditions by exhibiting phenotypic plasticity, which includes an increase in lignin synthesis and changes in leaf blade shape (twisting into tubes or folding) [90, 91, 92]. This helps the plant maintain optimal water balance [38, 89] and protects it from UV radiation. Lignin acts as an “aggressive” wall component that can displace other polysaccharides [93]. Researchers have proposed a biomechanical model demonstrating that crystalline cellulose is replaced by lignin in the stomatal end walls, serving to strengthen the wall.
Besides, lignin and its monomers are a chemical barrier that increases the protecting of cells from penetration of water and invasion by pathogens [50], and the sign (S/G) testifies to the increase of mechanical durability of cells [51]. The presence of lignin and crystalline cellulose in guard cells implies different biomechanical functions. Our experimental data suggests that in terrestrial plants, leaves of reeds receive a higher dose of ultraviolet radiation than leaves of reeds in plants growing in water. The high content of syringyl monolignol in terrestrial plants can serve as a marker for making strong bricks, while the increased content of other guajacyl monolignol in air-water plants can be a marker for getting soft paper from the reeds. The presence and ratio of monolignols in the cell walls of the epidermis and vessels correlate with early-received data for upland plants [94]. It is possible that the leaf cell walls of the plants studied also contain p-hydroxyphenyl, which plays a substantial role in lignin structure and can be determined using UV spectroscopy [95].
Thus, it was established that the changes at tissue and cellular levels are accompanied by an increase in lignin and cellulose in plants grown under drought stress. However, changes in lignin content in plants under the influence of the environment vary, with some experiments showing a decrease and others showing an increase. This may depend on the plant species and growth phase used for the study. Plant protection during drought occurs at both the cellular and molecular levels. Researchers have noted increased enzyme activity of CoA reductase and the expression of relevant genes in response to moderate water deficiency in the soil. This led to an increased synthesis of monolignols in the cell walls of
4. The influence of cold on leaf lignin
The main causes of plant death from cold are direct or prolonged exposure to low temperatures and the impact on crop growth and development. Researchers categorize plants into three types based on their response to cold: sensitive to cooling, resistant to cold, and adapted to low temperatures [96]. Plants that overwinter in temperate climates can tolerate the freezing of a significant amount of water on the cell surface. Ice crystal formation begins when exposed to temperatures ranging from −1 to −3°C for one hour. The receptors for cold perception in a plant cell have not yet been identified, but it is assumed that the cold signal is perceived by converting it into a calcium signal, which triggers further processes [97].
A study of the structural and functional features of plants growing in the Arctic may provide an answer to the natural mechanisms of adaptation of the plant world to the effects of low temperatures. Polar plants are characterized by the inhibition of respiration in the light, with ATP production occurring during the day through photorespiration and dark respiration observed only at night. This has been shown in polar species such as
Cold shock proteins and genes, including desaturases, and dehydrins are hydrophilic and are secreted into the cell walls of epidermal tissues. They are then placed on the surface of ice crystals, inhibiting their further growth [99, 100]. Freezing water from the outside of the cell prevents the formation of ice crystals in the protoplast but leads to dehydration of the cell’s macromolecular structures. This occurs due to increased outflow of water from the cell through the plasmalemma, resulting in changes to the composition of phospholipids. Water then goes into the intercellular spaces, causing severe dehydration of the protoplast [101, 102]. These events lead to a decrease in cell volume, deformation, and changes in the mechanical properties of cell walls [103, 104, 105].
Cold acclimatization leads to changes in the structure of the cell wall and the composition of polysaccharides, including thickening of the cell wall of the epidermis and alterations in the synthesis of cellulose, soluble polysaccharides, and pectins [105, 106]. Studies on oilseed rape seedlings exposed to cold conditions showed significant changes in the content of major polysaccharides [107]. The resistance of the cell walls of the epidermis to frost affects the protection of the protoplast from ice formation, resulting in changes in leaf structure and cell wall thickness in cold conditions [99, 108].
A study of the epidermis ultrastructure in the leaves of

Figure 7.
Ultrastructure of epidermal cells of the leaves of
It was early established that low temperatures did not significantly affect the content of lignin, but there were changes in enzyme activity and an increase in the expression of genes coding for
Summing up the numerous literature data and the results of our experimental studies, we conclude that regardless of the type of extreme environmental impact on plants, the cells of aboveground organs undergo increased synthesis of lignin. The content and composition of lignin depend on the type of stress and its duration. Cellulose is also synthesized in the cell walls in parallel, regardless of the type of stress. The crystalline form of cellulose, along with monolignins, creates a natural barrier to water transport by inhibiting apoplastic transport, as well as cuticular and stomatal transpiration [30, 115]. It is known that the amorphous zones of cellulose are involved in the transport and absorption of water molecules, while the crystalline form of cellulose does not have this ability [116]. We believe that it is extremely interesting and valuable for fundamental biology and applied sciences to establish the interaction of lignin oligomers with the crystalline form of cellulose-1β [117, 118]. This was experimentally demonstrated using the leaves of
In practice, this invention is used in the development of natural wood adhesives. In my opinion, such natural adhesives with lignin oligomers and cellulose can also be used in medicine, textile, and building materials industry. In our opinion, the leaves of reeds and cattails that grow in the coastal strips of European rivers, including the rivers of Ukraine, can serve as a source of such adhesives. Why reeds and cattails? Because the leaves of these plants, in addition to the monolignols (guajacyl and syringyl) and crystalline cellulose, also contain a lot of silicon, which easily absorbs and reflects sunlight. Such properties of natural materials are valuable for their further use in practice.
5. Conclusions
This review indicates that the leaf epidermis plays a role in the adaptation of plants to submergence and drought at structural and biochemical levels. The adaptation mechanisms of the epidermis in leaves to flooding are expressed in the absence of stomata and wax in the leaf epidermis, the loosening of cell walls, and the presence of cuticle pores. Meanwhile, the adaptation mechanisms of the leaf epidermis to drought are expressed in an increase of wax and cuticle, as well as an increase in trichome size and density. The questions involving the epidermal walls of submerged and drought plants are so varied and numerous that this field of plant morphology, anatomy, and cell biology will continue to attract the attention of many investigators to establish not only regularities of fundamental biology but also to use theoretical-experimental data for constructing agricultural plants with resistance to environmental stress.
The adaptation mechanisms of the epidermis of leaves to flooding and drought are also expressed in the synthesis of monolignols syringyl and guajacyl, the content of which depends on environmental conditions. Syringyl and guajacyl monolignols were detected in the leaves of air-water and terrestrial plants using cytochemical methods and laser confocal microscopy. The Pascal program for laser confocal microscopic study of monolignols in the leaf epidermis of hydrophytes revealed that the external environment of the leaf epidermis (water or air) can change the content of monolignols in the epidermis wall. It is established that the anticlinal walls of usual epidermal cells are the main accumulator of guajacyl monolignol. An increase in the content of guajacyl monolignol in the anticlinal walls of the epidermis leads to an increase in buoyancy and the optimization of plant life support on the water surface. It was found that the decrease in soil moisture leads to an increase in the content of monolignols (syringyl and guajacyl, S + G, and ratio of S/G) in the epidermis of plants under drought. An increase in syringyl and guajacyl content in the cell walls of epidermal cells of the leaves of terrestrial plants can reduce cuticular and stomata transpiration and lead to optimization of the water balance of plants grown in moderate drought.
Our experimental data show that the high content of syringyl monolignol, which gives high reed strength in terrestrial plants, can serve as a marker for the use of terrestrial reed plants for making strong bricks for commercial use. The increased content of other guajacyl monolignol in the leaves of air-water plants gives softness to the leaves and can be a marker for getting soft paper from the reeds grown on the banks of the river.
The tolerance mechanism of the leaf to cold is expressed through an increase in syringyl synthesis in the outer wall of the leaf epidermis. Given that syringol monolignol increases the mechanical stability of cells and acts as a barrier to water transport [51] and the action of ice crystals, we can assume that syringol in the periclinal walls of the epidermis of artificial grasses is an active component of plant cold resistance.
Acknowledgments
The author thanks Dr. Klimchuk D.O. for help during scanning electron microscopy of the samples.
Funding
The preparation and writing of the manuscript chapter (title: “Influence of Environmental Change on Monolignols and the Micromorphology of Leaf Epidermis in Hydrophytes and Terrestrial Plants” for Advances Lignin Technologies) was not funded by M.G. Kholodny Institute of Botany of the National Academy of Sciences of Ukraine.
References
- 1.
Dietz K-J, Hartung W. The leaf epidermis: Its ecophysiological significance. Progress in Botany. Fortschritte der Botanik. 1996; 57 :32-53. DOI: 10.1007/978-3-642-79844-3_3 - 2.
Mommer L, Visser EJW. Underwater photosynthesis in flooded terrestrial plants: A matter of leaf plasticity. Annals of Botany. 2005; 96 :581-589. DOI: 10.1093/aob/mci212 - 3.
LoPresti EF. Chemicals on plant surfaces as a heretofore unrecognized, but ecologically informative, class for investigations into plant defence. Biological Reviews. 2015; 91 :1102-1117. DOI: 10.1111/brv.12212 - 4.
Kerstein G. Cuticular water permeability and its physiological significance. Journal of Experimental of Botany. 1996; 47 :1813-1832. DOI: 10.1093/jxb/47.12.1813 - 5.
Werker E. Trichome diversity and development. Advances in Botanical Research. 2000; 31 :1-35. DOI: 10.1016/S0065-2296(00)31005-9 - 6.
Bickford CP. Eco-physiology of leaf trichomes. Functional Plant Biology. 2016; 43 :807-814. DOI: 10.1071/FP16095 - 7.
Dixon RR, Chen F, Guo D, Parvathi K. The biosynthesis of monolignols: A “metabolic grid”, or independent pathways to guaiacyl and syringyl units? Phytochemistry. 2001; 57 :1069-1084. DOI: 10.1016/S0031-9422(01)00092-9 - 8.
Roland JC, Vian B. The wall of the growing cell: Its three dimension organization. International Review of Cytology. 1979; 61 :129-166. DOI: 10.1016/S0074-7696(08)61997-X - 9.
Schuetz M, Benske A, Smith R, Watanabe Y, Tobimatsu Y, et al. Laccases direct lignification in the discrete secondary cell wall domains of protoxylem. Plant Physiology. 2014; 166 :798-807. DOI: 10.1104/pp.114.245597 - 10.
Vartapetian BB, Jackson MB. Plant adaptations to anaerobic stress. Annals of Botany. 1997; 79 :3-20. DOI: www.jstor.org/stable/42764824 - 11.
Doronin Y. Ocean Physics. St. Petersburg: Russian State Hydrometeorological University. Publ. RSHU; 2000. 340 p. (in Russ.) - 12.
Madsen TV, Maberty SC. Diurnal variation in light and carbon limitation of photosynthesis by two species of submergered freshwater macrophytes with a differential ability to use bicarbonate. Freshwater Biology. 1991; 26 :175-187. DOI: 10.1111/j.1365-2427.1991.tb01727.x - 13.
Sand-Jensen K, Pedersen MF, Nielsen SL. Photosynthetic use of inorganic carbon among primary and secondary water plants in streams. Freshwater Biology. 1992; 27 :283-293. DOI: 10.1111/J.1365-2427.1992.TB00540.X - 14.
Mommer L, Pederson O, Visser EJW. Acclimation of a terrestrial plant to submergence facilitates gas exchange under water. Plant Cell and Environment. 2004; 27 :1281-1287. DOI: 10.1111/j.1365-3040.2004.01235.x - 15.
Nedukha OM. Heterophylly in Plants. Kyiv: Publ.: Alt Press; 2011. 191 p. (in Ukrainian). ISBN 978-066-542-494-9 - 16.
Schönherr J. Resistance of plant surfaces to water loss: Transport properties of cutin, suberin and associated lipids. In: Lange О., Nobel P., Osmond C., et al., editors. Physiological Plant Ecology. II. Encyclopedia of Plant Physiology. Heidelberg: Springer; 1982. 2B. p. 153-179. - 17.
Schönherr J. Characterization of aqueous pores in plant cuticles and permeation of ionic solutes. Journal of Experimental Botany. 2006; 57 :2471-2491. DOI: 10.1093/jxb/erj217 - 18.
Bruni NC, Yang JP, Dengler NG. Leaf development plasticity of Ranunculus aquatilis in response to terrestrial and submerged environments. Canadian Journal of Botany. 1996;74 :823-827. DOI: 10.1139/b96-103 - 19.
Rascio N, Cuccato F, Dalla Vecchia F, et al. Structural and functional features of the leaves of Ranunculus trichphyllus Chaix., a freshwater submerged macrophyte. Plant Cell and Environment. 1999;22 :205-212. DOI: 10.1046/j.1365-3040.1999.00394.x - 20.
Frost-Christensen H, Bolt Jorgensen L, Flotoo F. Species specificity of resistance to oxygen diffusion in thin cuticular membranes from amphibious plants. Plant, Cell and Environment. 2003; 26 :561-569. DOI: 10.1046/j.1365-3040.2003.00986.x - 21.
Frost-Christensensen H, Floto F. Resistance to CO2 diffusion in cuticular membranes of amphibious plants and the implication for CO2 acquisition. Plant Cell and Environment. 2007; 30 :12-18. DOI: 10.1111/j.1365-3040.2006.01599.x - 22.
Hoffmannbenning S, Kend H. On the role of abscisic-acid and gibberellin in the regulation of growth in rice. Plant Physiology. 1992; 99 :1156-1161. DOI: 10.1104/pp.99.3.1156 - 23.
Armstrong W, Brande R, Jackson MB. Mechanisms of flood tolerance in plants. Acta Botanica Neerlandica. 1994; 43 :307-358. DOI: 10.1111/j.1438-8677.1994.tb00756.x - 24.
Jackson MB, Colmer TD. Response and adaptation by plants to flooding stress. Annals of Botany. 2005; 96 :501-505. DOI: 10.1093/aob/mci205 - 25.
Nedukha OM. Epidermis leaf structural responses of some aquatic plants to constant water environment. Advances of Agricultural Sciences. Problem Issue. 2010; 545 (545):169-178. DOI: 10.13140/RG.2.1.2174.0562 - 26.
Nedukha OM. Structural features of the cell walls in the leaves of Sagittaria sagittifolia andTrapa natans . Modern Phytomorphology. 2013;3 :259-265. УДК 57.012.4:581.821 - 27.
Nedukha OM. Role of cell walls in plant adaptation to flooding. Journal of Biology and Nature . 2017;7 :38-62. ISSN: 2395-5376 (print), 2395-5384 - 28.
Yates D, Hutley LB. Polar uptake of water by wet leaves of Sloanea woollsii an Australian subtropical rainforest trees. Australian Journal of Botany. 1995;43 :157-167. DOI: 10.1071/BT9950157 - 29.
Leisola M, Pastinen O, Axe DD. Lignin—Designed. Bio-complexity.org . 2012;3 :1-11. DOI: 10.5048/BIO-C.2012.3.c - 30.
Panda D, Sarkar RK. Structural carbohydrates and lignifications associated with submergence tolerance in race ( Oryza sativa L.). Journal of Stress Physiology and Biochemistry. 2013;9 :299-306. ISSN: 1997-0838 - 31.
Tyree MT, Cheung YNS. Resistance to water flow in Fagus grandifolia leaves. Canadian Journal of Botany. 1997; 55 :2591-2599. DOI: 10.1139/b77-296 - 32.
Niklas KJ. Plant Biomechanics: An Engineering Approach to Plant Form and Function. University of Chicago Press; 1992. 622 p - 33.
Wilson K, White DJB. Reaction wood: Its structure, properties and functions. In: Wilson K, Stobart WDJB, editors. The Anatomy of Wood: Its Diversity and Variability. London, UK; Stobart; 1986. pp. 222-250. ISBN 10: 0854420339 /ISBN 13:9780854420339 - 34.
Joseleau J-P, Imai T, Kuroda K, Ruel K. Detection in situ and characterization of lignin in the G-layer of tension wood fibres of Populus deltoides . Planta. 2004;219 :338-345. DOI: 10.1007/s00425-004-1226-5 - 35.
Nedukha OM. The participation of wall monolignols in leaf tolerance to nature flooding of hydrophytes. Advances in Bioscience and Biotechnology. 2015; 6 :546-555. DOI: 10.4236/abb.2015.68058 - 36.
Nedukha OM. Cytochemical study of monolignols in cell walls of hydrophytes leaves. Ukrainian Journal of Botany. 2015; 72 :385-392. (In Ukrainian). DOI:nbuv.gov.ua/UJRN/UBJ _ - 37.
Moura JC, Bonine CA, Viana J, Dornelas MC, Mazzafera P. Abiotic and biotic stresses and changes in the lignin content and composition in plants. Journal Integrative Plant Biology. 2010; 52 :360-376. DOI: 10.1111/j.1744-7909.2010.00892.x - 38.
Hilal M, Parrado M, Rosa M, Gallardo M, Orce L, et al. Epidermal lignin deposition in quinoa cotyledons in response to UV-B radiation. Photochemistry and Photobiology. 2004;79 :205-210. DOI: 10.1562/0031-8655(2004)079 <0205:ELDIQC>2.0.CO;2 - 39.
Little ST. Adaptation and acclimation of populations of Ludwigia repens to growth in high- and lower CO2 springs [a dissertation presented to the graduate school of the University of Florida in partial fulfillment of the requirements for the degree of doctor of philosophy]. USA: Univ. Florida; 2003. pp. 1-157 - 40.
Vanholme R, De Meester B, Ralph J, Wout BW. Lignin biosynthesis and its integration into metabolism. Current Opinion in Biotechnology . 2019;56 :230-239. DOI: 10.1016/j.copbio.2019.02.018 - 41.
Alain-M B. Towards an understanding of the supramolecular organization of the lignified wall. In: Rose JKC, editor. The Plant Cell Wall. Ithaca, NY, USA: Dep. Plant Biol. Cornell Univ.; 2001. pp. 155-182 - 42.
Lai YZ, Funaoka M, Chen HT. Chemical heterogeneity in woody lignins. In: Argyropoulos DS, editor. Advances in Lignocellulosics Characterization. Atlanta: TAPPI Press; 1999. pp. 43-53 - 43.
Chabannes M, Ruel K, Yoshinaga A. In situ analysis of lignins from tobacco transgenic lines down-regulated for several enzymes of monolignol synthesis reveals a differential impact of transformation on the spatial patterns of lignin deposition at the cellular and subcellular levels. Plant Journal. 2001;28 :271-282. DOI: 10.1046/j.1365-313x.2001.01159.x - 44.
Ancibor E. Systematic anatomy of vegetative organs of the Hydrocharitaceae. Botanical Journal of the Linnean Society. 1970; 78 :237-266. DOI: 10.1111/j.1095-8339.1979.tb02570.x - 45.
Nedukha OM. Heterophylly in Sagittaria sagittiflia . II. Surface ultrastructure of above=water and submerged leaves. Ukrainian Botanical Journal. 2012;69 :596-603. (In Ukrainian) - 46.
Bercu R. Some histological and cytological features on four hydrophytes occurring in the Danube Delta (Romania). Contributii Botanice, (Grădina Botanică “Alexandru Borza” Cluj-Napoca). 2003; 38 :127-134 - 47.
Sanduescu EB, Stavrescu- Bedivan MM, Scaeteanu GV, Schiopu T. Morpho-anatomic features and chemical compounds in some aquatic plant species – preliminary data. Scientific Papers. Series A. Agronomy. 2014; LVII :441-447. ISSN 2285-5785, ISSN CD-ROM 2285-5793 - 48.
Jung J, Lee JSC, Chu H-K. Anatomical patterns of aerenchyma in aquatic and wetland plants. Journal of Plant Biology. 2008; 51 :428-439. DOI: 10.1007/bf03036065 - 49.
Vogel S. Contributions to the functional anatomy and biology of Nelumbo nucifera (Nelumbonaceae) I. Pathways of air circulation. Plant Systematics and Evolution. 2004;249 :9-25. DOI: 10.1007/s00606-004-0201-8 - 50.
Menden B, Kohlhoff M, Moerschbacher BM. Wheat cell accumulate a syringil-rich lignin during the hypersensitive resistance response. Phytochemistry. 2007; 68 :513-529. DOI: 10.1016/j.phytochem.2006.11.011 - 51.
Christiernin M. Composition of lignin in outer cell-wall layers [PhD thesis]. Stockholm: Division of Wood Chemistry and Pulp Technology, Royal Institute of Technology, TRITA-FPT-Report; 2006. p. 153. ISSN1652-2443 ISRN KTH/FPT/R-2006/16-SE - 52.
Nedukha OM. Cell Wall and Environmental Factors. Kyiv: Alt Press; 2015. 288 p. (In Ukrainian. ISBN 978-966-542-575-5) - 53.
Nedukha OM, Kordyum EL. The plasticity of anatomical structure and cell wall lignin in Trapa natans adaptation to nature flooding. Annals of the Romanian Society for Cell Biology. 2016;21 :27-34. DOI: annalsofrscb.ro/index.php/journal/article/view/13 - 54.
Cosgrove DJ. Molecular regulation of plant cell wall extensibility. Gravitational and Space Biology Bulletin. 1998; 11 :61-70 - 55.
Rayle DL, Cleland RE. The acid growth theory of auxin-induced cell elongation. Plant Physiology. 1992; 99 :1271-1274. DOI: 10.1104/pp.99.4.1271 - 56.
Cosgrove DJ. Wall structure and wall loosening. A look backwards and forwards. Plant Physiology. 2001; 125 :131-134. DOI: 10.1104/pp.125.1.131 - 57.
Clouse SD. Molecular genetic analysis of brassinosteroid action. Physiology Plantarum. 1997; 100 :702-709. DOI: /10.1111/j.1399-3054.1997.tb03077.x - 58.
Nedukha OM. The participation of cell wall polysaccharide in cellular mechanisms of leaf tolerance to nature flooding of plants In: M.T. Grazeslak. Plant Functioning Under Environmental Stress. Cracow, Poland: DRUKROL LTD; 2013. p. 137-152. ISBN 978.83.86978-30-7 - 59.
Rastogi S, Dwivedi U. Manipulation of lignin in plants with special reference to O-methyltransferase. Plant Science. 2008; 174 :264-277. DOI: 10.1016/j.plantsci.2007.11.014 - 60.
Maun MA. Adaptations of plants to burial in coastal sand dunes. Canadian Journal of Botany. 1997; 6 :713-738. DOI: 10.1139/b98-058 - 61.
Grigore MN, Toma C. Histo-anatomical strategies of Chenopodiaceae halophytes; adaptive, ecological and evolutionary implications. WSEAS Transactions on Biology and Biomedicine. 2007;12 :204-218. ISSN:1109-9518 - 62.
Catoni R, Gratani L. Morphological and physiological adaptive traits of Mediterranean narrow endemic plants: The case of Centaurea gymnocarpa (Capraia Island, Italy). Flora. 2013;208 :174-183. DOI: 10.1016/j.flora.2013.02.010 - 63.
Zhu SD, Chen YJ, Ye Q , He PC, Liu H, et al. Leaf turgor loss point is correlated with drought tolerance and leaf carbon economics traits. Tree Physiology. 2018; 38 :658-663. DOI: 10.1093/treephys/tpy013 - 64.
Chaves MM, Flexas J, Pinheiro C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Annals of Botany. 2009; 103 :551-560. DOI: 10.1093/aob/mcn125 - 65.
Hu Y, Li WC, Xu YQ , Li GJ, Liao Y, et al. Differential expression of candidate genes for lignin biosynthesis under drought stress in maize leaves. Journal of Applied Genetics. 2009; 50 :213-223. DOI: 10.1007/BF03195675 - 66.
Hesp P. Ecological processes and plant adaptations on coastal dunes author links open overlay panel. Journal of Arid Environments. 1991; 21 :165-191. DOI: 10.1016/S0140-1963(18)30681-5 - 67.
Le Bagousse-Pinguet Y, Forey E, Touzard B, Michalet R. Disentangling the effects of water and nutrients for studying the outcome of plant interactions in sand dune ecosystems. Journal of Vegetation Science. 2013; 24 :375-383. DOI: 10.1111/j.1654-1103.2012.01462.x - 68.
Selvi F, Bigazzi MM. Leaf surface and anatomy in Boraginaceae tribe Boragineae with respect to ecology and taxonomy. Flora. 2001; 196 :269-285. DOI: 10.1016/S0367-2530(17)30056-7 - 69.
Hua Cong Ci, Xing Dong He, Rong Li, Wei Wu, PingPing Xue, et al. Characteristics of plant calcium fractions for 25 species in Tengger Desert. Sciences in Cold and Arid Regions. 2010;2:168-174. DOI: www.researchgate.net/publication/267692985 - 70.
Perronea R, Salmeri C, Brullo S, Colombo P. What do leaf anatomy and micro-morphology tell us about the psammophilous ? Flora. 2015;Pancratium maritimum L. (Amaryllidaceae) in response to sand dune conditions213 :20-31. DOI: 10.1016/j.flora.2015.03.001 - 71.
Nedukha OM. Micromorphology and monolignols of leaf epidermis in Phragmites australis (Poaceae) of air-aquatic and terrestrial ecotypes. Protoplasma. 2021;258 :949-961. DOI: 10.1007/s00709-021-01614-4 - 72.
Caine R, Yin X, Sloan J, Harrison E, Umar Mohammed U, et al. Rice with reduced stomatal density conserves water and has improved drought tolerance under future climate conditions. New Phytologist. 2019; 221 :371-384. DOI: 10.1111/nph.15344 - 73.
Peter AJ, Shanower TO, Romeis J. The role of plant trichomes in insect resistance: A selective review (International Crops Research Institute for the semi-arid tropics). Journal: Phytophaga. 1995; 7 :41-63. Available from:https://www.scirp.org›refer - 74.
Hauke V, Schreiber L. Ontogenetic and seasonal development of wax composition and cuticular transpiration of ivy ( Hedera helix L.) sun and shade leaves. Planta. 1998;207 :67-75. DOI: 10.1007/s004250050456 - 75.
Barlott W, Mattias M, Bharat B, Koch K. Plant surface: Structures and functions for biomimetic innovations. Nano-Micro Letters. 2017; 9 :1-40. DOI: 10.1007/s40820-016-0125-1 - 76.
Ashraf M, Harris PJC. Photosynthesis under stressful environments: An overview. Photosynthetica. 2013; 51 :163-190. DOI: 10.1007/s11099-013-0021-6 - 77.
Ruocco M, Bertoni D, Sarti G, Ciccarelli D. Mediterranean coastal dune systems: Which abiotic factors have the most influence on plant communities? Estuarine, Coastal and Shelf Science. 2014; 149 :213-222. DOI: 10.1016/j.ecss.2014.08.019 - 78.
Futorna OA, Badanina VA, Zigalova CL. Ecologo-anatomic characterization of some species Tragopogon (Asteraceae ) of Ukraine Flora. Biosystems Diversity. 2017;25 :274-281. (In Ukrainian). DOI: 10.15421/011742 - 79.
Nedukha OM. Appearance of phenotypic plasticity of leaves in psammophyte Corynephorus canescens during flooding. Plant Introduction. 2021; 91 (92):24-35. DOI: 10.46341/PI2021011 - 80.
Abd E, Mona E, Abo-Alatta OK, Habib SA, Abd Elba Ola H. The anatomical features of the desert halophytes Zygophyllum album L.F. andNitraria retusa . Annals of Agricultural Science. 2016;61 :1-8. DOI: 10.1016/j.aoas.2015.12.001 - 81.
Hameed BH, Krishni RR, Sata SA. A novel agricultural waste adsorbent for the removal of cationic dye from aqueous solutions. Journal of Hazardous Materials. 2009; 162 :305-311. DOI: 10.1016/j.jhazmat.2008.05.036 - 82.
Jianu LD, Bercu R,Popoviciu DR. Silene thymifolia Sibth. et Sm. (Caryophyllaceae) – A vulnerable species in Romania: Anatomical aspects of vegetative organs. Notulae Scientia Biologicae. 2021;13 :1-7. DOI: 10.15835/nsb13110875 - 83.
Bourland FM, Hornbeck JM, McFall AB, Calhoun SD. A rating system for leaf pubescence of cotton. The Journal of Cotton Science. 2003; 7 :8-15. Available from:http://journalofcotton.org - 84.
Ciccarelli D, Forino L, Costantina M, Balestri M, Maria PA. Leaf anatomical adaptations of Calystegia soldanella ,Euphorbia paralias andOtanthus maritimus to the ecological conditions of coastal sand dune systems. Caryologia. 2009;62 :142-151. DOI: 10.1080/00087114.2004.10589679 - 85.
Berthet S, Demontcaulet N, Pollet B, Bidzinski P, Cezard L, et al. Disruption of LACCASE4 and 17 results in tissue-specific alterations to lignification ofArabidopsis thaliana stems. The Plant Cell. 2011;23 :1124-1137. DOI: ff10.1105/tpc.110.082792ff. ffhal-01000145f - 86.
Fan L, Linker R, Gepstein S, Tanimoto E, Yamamoto R, et al. Progressive inhibition by water deficit of cell wall extensibility and growth along the elongation zone of maize roots is related to increased lignin metabolism and progressive stelar accumulation of wall phenolics. Plant Physiology. 2006; 140 :603-612. DOI: DOI; 10.1104/pp.105.073130 - 87.
Agrawal A. Induced responses to herbivory and increased plant performance. Science. 1998; 279 :1201-1202. DOI: 10.1126/science.279.5354.1201 - 88.
Begovic L, Abičić I, Alojzije L, Lepeduš H, Cesar V, et al. Lignin synthesis and accumulation in barley cultivars differing in their resistance to lodging. Plant Physiology and Biochemistry. 2018; 133 :142-148. DOI: 10.1016/j.plaphy.2018.10.036 - 89.
Monties B, Fukushima K. Occurrence, function and biosynthesis of lignins. In: Hofrichter M, Steinbuchel A, editors. Biopolymers. Lignin, Humic Substances and Coal. Vol. 1. Weinheim: Wiley; 2001. pp. 1-64. DOI: 10.1002/3527600035.bpol1001 - 90.
Micol JL, Hake S. The development of plant leaves. Plant Physiology. 2003; 131 :389-394. DOI: 10.1104/pp.015347 - 91.
Kadioglu A, Terzi R. A dehydration avoidance mechanism: Leaf rolling. The Botanical Review. 2007; 73 :290-302. DOI: 10.1663/0006- 8101(2007)73[290:ADAMLR]2.0.CO;2 - 92.
Candela H, Johnston R, Gerhold A, Foster T, Hake S. The milkweed pod 1 gen encode a KANADI protein that is required for abaxial/adaxial patterning in maize leaves. The Plant Cell. 2008; 20 :2073-2087. DOI: 10.1105/tpc.108.059709 - 93.
Shtein H, Shelef Y, Marom Z, Zelinger E, Schwartz A, et al. Stomatal cell wall composition: Distinctive structural patterns associated with different phylogenetic groups. Annals of Botany. 2017; 119 :1021-1033. DOI: 10.1093/aob/mcw275 - 94.
Baucher M, Monties B, Van Montagu M, Boerjan W. Biosynthesis and genetic engineering of lignin. Critical Reviews in Plant Sciences. 1998; 17 :125-197. DOI: 10.1080/ 07352689891304203 - 95.
Fath L, Arno Frьhwald A, Koch G. Distribution of lignin in vascular bundles of coconut wood ( Cocos nucifera ) by cellular UV-spectroscopy and relationship between lignification and tensile strength in single vascular bundles. Holzforschung. 2014;68 :915-925. DOI: www.researchgate.net/publication /269989221 - 96.
Thakur P, Nayyar H. Facing the cold stress by plants in the changing environment: Sensing, signaling, and defending mechanisms. In: Tuteja N, Gill SS, editors. Plant Acclimation to Environmental Stress. New York: Springer Science, Business Media; 2013. pp. 29-69 - 97.
Miura K, Furumoto T. Cold signaling and cold response in plants. International Journal of Molecular Sciences. 2013; 14 :5312-5337. DOI: 10.3390/ijms14035312 - 98.
Heskel M. Environmental Controls of Foliar Respiration in Arctic Tundra Plants. Submitted in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy in the Graduate School of Arts and Sciences. Columbia University; 2013. 222 p - 99.
Griffith M, Brown G. Cell wall deposition in winter rye Secale cereale L. “puma” during cold acclimatization. Botanical Gazette. 1982;143 :486-490. Available from:www.jstor.org/stable/2474764 - 100.
Griffith M, Lumb C, Wiseman SB. Antifreeze proteins modify the freezing process in planta. Plant Physiology. 2005; 138 :330-340. DOI: 10.1104/pp.104.058628 - 101.
Steponkus PL, Uemura M, Webb MS. A contrast of the cryostability of the plasma membrane of winter rye and spring oat-two species that widely differ in their freezing tolerance and plasma membrane lipid composition. In: Steponkus PL, editor. Advances in Low-Temperature Biology. Vol. 2. London: JAI Press; 1993. pp. 211-312 - 102.
Steponkus PL, Uemura M, Webb MS. Membrane destabilization during freeze-induced dehydration. Current Topics Plant Physiology. 1993; 10 :37-47. DOI: escholarship.org - 103.
] Pearce RS. Extracellular ice and cell shape in frost-stressed cereal leaves: A low-temperature scanning-electron-microscopy study. Planta. 1988; 175 :313-324. DOI: 10.1007/BF00396336 - 104.
Pearce RS, Ashworth EN. Cell shape and localization of ice in leaves of overwintering wheat during frost stress in the field. Planta. 1992; 188 :324-331. DOI: 10.1007/BF00192798 - 105.
Pearce Rogen S. Plant freezing and damage. Annals of Botany. 2001; 87 :417-424. DOI: 10.1006/anbo.2000.1352 - 106.
Wallner SJ, Wu M, Anderson-Krengel SJ. Changes in extracellular polysaccharides during cold acclimation on cultured pear cells. Journal of the American Society for Horticultural. 1986; 111 :769-773 - 107.
Kubacka-Zebalska M, Kacperska A. Low temperature-induced modifications of cell wall content and polysaccharide composition in leaves of winter oilseed rape ( Brassica napus L. var.oleifera L.). Plant Science. 1999;148 :59-67 - 108.
Rajashekar C, Lafta A. Cell-wall changes and cell tension in response to cold acclimation and exogenous abscisic acid in leaves and cell cultures. Plant Physiology. 1996; 111 :605-612. DOI: 10.1104/pp.111.2.605 - 109.
Kordyum EL, Nedukha OM, Ovcharenko YV, Jadko SI, Ivanenko GF, et al. Anatomical and functional features of Deschampsia antarctica (Poaceae) leaf blade growing on the Argentine islands. Ukrainian Antarctic Journal. 2017;16 :143-149. DOI: 10.33275/1727-7485.16.2017.72 - 110.
Gielwanowska I, Szczuka E, Bednara J, Gorecki R. Anatomical features and ultrastructure of Deschampsia antarctica (Poaceae) leaves from different growing habitats. Annals of Botany. 2005;96 :1109-1119. DOI: 10.1093/aob/mci262 - 111.
Romero M, Casanova A, Iturra G, Reyes A, Montenegro G, et al. Leaf anatomy of Deschampsia antarctica (Poaceae) from the maritime Antarctic and its plastic response to changes in the growth conditions. Revista Chilena de Historia Natural. 1999;72 :411-425 - 112.
Szczuka E, Gielwanowska I, Leszczuk A, Domaciuk M, Pietrusiewicz J, et al. Specific ultrastructure of the leaf mesophyll cells of Deschampsia antarctica Desv. (Poaceae). Annales Universitatis Mariae-Curie-Sklodowska, Sectio C – Biologia. 2013;8 :25-33. DOI: 10.2478/v10067-012-0031-y - 113.
El Kayal W, Keller G, Debayles C. Regulation of tocopherol biosynthesis through transcriptional control of tocopherol cyclase during cold hardening 873 in Eucalyptus gunnii . Physiology Plantarum. 2006;126 :212-223. DOI: 10.3389/fpls.2021.743993 - 114.
Ford CW, Morrison IM, Wilson JR. Temperature effects on lignin, hemicellulose and cellulose in tropical and temperate grasses. Australian Journal of Agricultural Research . 1979;30 :621-633. DOI: 10.1071/AR9790621 - 115.
Hausman JF, Evers D, Thiellement H, Jouve L. Compared responses of poplar cuttings and in vitro raised shoots to short-term chilling treatments. Plant Cell Reports. 2000;19 :954-960. DOI: 10.1007/s002990000229 - 116.
Brown RM. The biosynthesis of cellulose. Journal of Macromolecular Science. 1996; 33 :1345-1373. DOI: 10.1080/10601329608014912 - 117.
Sedano-Mendoza M, Lopez-Albarran P, Pizzi A. Natural lignans as adhesives for cellulose: Computational interaction energy vs experimental results. Journal of Adhesion Science and Technology. 2010;24 :1769-1786 - 118.
Lopez-Albarran P, Pizzi P, Navarro-Santos P, Hernandez-Esparza R. Oligolignols within lignin-adhesive formulations drive their Young's modulus: A ReaxFF-MD study. International Journal of Adhesion and Adhesives. 2017; 78 :227-233. DOI: 10.1016/j.ijadhadh.2017.08.003